
Abstract
NEAT1 long noncoding RNA orchestrates paraspeckle assembly and impacts tumorigenesis, fertility and immunity. Its maturation requires RNase P cleavage yielding an unstable transfer RNA-like multiple endocrine neoplasia-β tRNA-like transcript (menRNA) due to CCACCA addition. Here we report the crystal structure of human menRNA, which partially mimics tRNAs to drive RNase P and ELAC2 processing. Biophysical analyses uncover an RNA-centric, riboswitch-like mechanism whereby the nascent CCA reshapes the RNA folding landscape and propels a spontaneous conformational isomerization that directs repeat CCA addition, marking the menRNA and defective tRNAs for degradation.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others


Data availability
Atomic coordinates and structure factor amplitudes for the human menRNA have been deposited at the Protein Data Bank under accession code 8VT5 (https://www.rcsb.org/structure//8VT5). Source data are provided with this paper.
References
Hutchinson, J. N. et al. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genomics 8, 39 (2007).
Clemson, C. M. et al. An architectural role for a nuclear noncoding RNA: NEAT1 RNA is essential for the structure of paraspeckles. Mol. Cell 33, 717–726 (2009).
Sasaki, Y. T., Ideue, T., Sano, M., Mituyama, T. & Hirose, T. MENepsilon/beta noncoding RNAs are essential for structural integrity of nuclear paraspeckles. Proc. Natl Acad. Sci. USA 106, 2525–2530 (2009).
Sunwoo, H. et al. MEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspeckles. Genome Res. 19, 347–359 (2009).
Butler, A. A., Johnston, D. R., Kaur, S. & Lubin, F. D. Long noncoding RNA NEAT1 mediates neuronal histone methylation and age-related memory impairment. Sci. Signal. 12, eaaw9277 (2019).
Hirose, T. et al. NEAT1 long noncoding RNA regulates transcription via protein sequestration within subnuclear bodies. Mol. Biol. Cell 25, 169–183 (2014).
Adriaens, C. et al. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat. Med. 22, 861–868 (2016).
Imamura, K. et al. Long noncoding RNA NEAT1-dependent SFPQ relocation from promoter region to paraspeckle mediates IL8 expression upon immune stimuli. Mol. Cell 53, 393–406 (2014).
Zhang, B. et al. Identification and characterization of a class of MALAT1-like genomic loci. Cell Rep. 19, 1723–1738 (2017).
Wilusz, J. E., Freier, S. M. & Spector, D. L. 3′ end processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA. Cell 135, 919–932 (2008).
Skeparnias, I. et al. Structural basis of MALAT1 RNA maturation and mascRNA biogenesis. Nat. Struct. Mol. Biol. https://doi.org/10.1038/s41594-024-01340-4 (2024).
Wilusz, J. E., Whipple, J. M., Phizicky, E. M. & Sharp, P. A. tRNAs marked with CCACCA are targeted for degradation. Science 334, 817–821 (2011).
Kuhn, C. D., Wilusz, J. E., Zheng, Y., Beal, P. A. & Joshua-Tor, L. On-enzyme refolding permits small RNA and tRNA surveillance by the CCA-adding enzyme. Cell 160, 644–658 (2015).
Betat, H. & Morl, M. The CCA-adding enzyme: a central scrutinizer in tRNA quality control. Bioessays 37, 975–982 (2015).
Zhang, J. & Ferré-D’Amaré, A. R. Direct evaluation of tRNA aminoacylation status by the T-box riboswitch using tRNA–mRNA stacking and steric readout. Mol. Cell 55, 148–155 (2014).
Suddala, K. C. et al. Direct observation of tRNA-chaperoned folding of a dynamic mRNA ensemble. Nat. Commun. 14, 5438 (2023).
Jean, J. M. & Hall, K. B. 2-Aminopurine fluorescence quenching and lifetimes: role of base stacking. Proc. Natl Acad. Sci. USA 98, 37–41 (2001).
Hermann, T. & Patel, D. J. Stitching together RNA tertiary architectures. J. Mol. Biol. 294, 829–849 (1999).
Hood, I. V. et al. Crystal structure of an adenovirus virus-associated RNA. Nat. Commun. 10, 2871 (2019).
Murray, M. H., Hard, J. A. & Znosko, B. M. Improved model to predict the free energy contribution of trinucleotide bulges to RNA duplex stability. Biochemistry 53, 3502–3508 (2014).
Hopper, A. K. & Zhang, J. Captured: the elusive eukaryotic tRNA splicing enzyme. Nat. Struct. Mol. Biol. 30, 711–713 (2023).
Hayne, C. K. et al. Structural basis for pre-tRNA recognition and processing by the human tRNA splicing endonuclease complex. Nat. Struct. Mol. Biol. 30, 824–833 (2023).
Sekulovski, S., Susac, L., Stelzl, L. S., Tampe, R. & Trowitzsch, S. Structural basis of substrate recognition by human tRNA splicing endonuclease TSEN. Nat. Struct. Mol. Biol. 30, 834–840 (2023).
Zhang, J. & Landick, R. A two-way street: regulatory interplay between RNA polymerase and nascent RNA structure. Trends Biochem. Sci. 41, 293–310 (2016).
Trachman, R. J. 3rd, Passalacqua, L. F. M. & Ferre-D’Amare, A. R. The bacterial yjdF riboswitch regulates translation through its tRNA-like fold. J. Biol. Chem. 298, 101934 (2022).
Zhang, J. Probing RNA structures and interactions using fluorescence lifetime analyses. Methods Mol. Biol. 2568, 13–23 (2023).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Liebschner, D. et al. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Acta Crystallogr. D 75, 861–877 (2019).
Lavery, R., Moakher, M., Maddocks, J. H., Petkeviciute, D. & Zakrzewska, K. Conformational analysis of nucleic acids revisited: Curves+. Nucleic Acids Res. 37, 5917–5929 (2009).
Acknowledgements
We thank I. Botos for computational support; G. Piszczek and D. Wu for support in biophysical analyses; D. Anastasakis, M. Hafner and Y. He for cell culture support; C. Stathopoulos, C. Bou-Nader, A. Umuhire Juru, A.-N. Shaukat and A. Brasington for discussions. This work was supported by the Intramural Research Program of the National Institutes of Health, the National Institute of Diabetes and Digestive and Kidney Diseases (ZIADK075136 to J.Z.).
Author information
Authors and Affiliations
Contributions
I.S. and J.Z. conceived and designed the work. I.S. prepared all RNA and protein samples and crystals, collected diffraction data and performed all biochemical and biophysical assays. I.S. and J.Z. determined, refined and analyzed the structure. Both authors contributed to data interpretation and manuscript preparation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural & Molecular Biology thanks Ling-Ling Chen, Sebastian Glatt and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Sara Osman was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
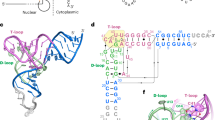
Extended Data Fig. 1 Secondary structures of human pre-tRNAGln, pre-mascRNA and pre-menRNA harboring their natural leaders and trailers.
a, b, c, Secondary structures of pre-tRNAGln (a) pre-mascRNA (b) and pre-menRNA (c). All three RNAs harbor 30-nucleotide (nt) natural 5′-leader sequences (red). pre-tRNAGln and pre-mascRNA contain 30-nt 3′-trailer sequences (red) while the pre-menRNA has a 5-nt 3′-trailer (red). RNase P and ELAC2 cleavage sites are indicated by blue arrowheads.
Extended Data Fig. 2 Secondary and crystal structures of human menRNA.
a, Secondary structure of the crystallization construct corresponding to the post-cleavage menRNA prior to 3′ CCA addition. To facilitate crystallization, the acceptor stem was extended by one G-C base pair (gray). b, c, Size exclusion chromatography (b) and non-denaturing PAGE electrophoresis (c) of the menRNA showing it exists primarily as a monomer in solution, based on its elution volume (n = 2 biologically independent replicates). d-f, Representative X-ray crystallographic electron density maps, including composite simulated anneal-omit 2|Fo | -|Fc| electron density calculated using the final model (contoured at 1.0 s.d.) superimposed with the final refined model of the menRNA (d), and portions of the map showing the elbow (e) or the acceptor stem (f) regions. g, Overall crystal-packing arrangements in the menRNA crystals. A reference molecule is shown in blue, while its symmetry-related molecules are in salmon. h, i, Detailed packing interfaces between menRNA molecules at the termini (h) and elbow (i) regions. j, Front, back, and side views of menRNA structure colored as in Fig. 1. The nucleobases of A25 and G27 in the anticodon loop lack clear electron density and only their backbones are modelled, and shown in gray. k, menRNA structure colored by crystallographic B-factors.
Extended Data Fig. 3 SHAPE and DMS chemical probing of human menRNA.
a, SHAPE probing of menRNA (n = 2 biologically independent replicates). Lanes 1-3: dideoxy sequencing ladders. Lanes 4 and 5: reverse transcription reactions using control (untreated) samples and samples treated by 2-methylnicotinic acid imidazolide (NAI). Modified nucleotides are indicated with red color while green, blue and purple colors denote A, C and U nucleotides, respectively. b, DMS probing of the menRNA (n = 2 biologically independent replicates). Lanes 1-3: dideoxy sequencing ladders. Lanes 4 and 5: reverse transcription reactions using control (untreated) samples and samples treated by DMS. Modified nucleotides are indicated with orange color while nucleotides are colored as in (a). c, Secondary structure of the human menRNA with a 22-nt 3′-trailer (gray) with which the Cy5-labeled probe annealed. Modified nucleotides were in red (SHAPE) and orange (DMS). d, e Secondary (d) and tertiary (e) structures of menRNA showing the location of the chemically modified nucleotides (red: SHAPE; orange: DMS).
Extended Data Fig. 4 CD analyses of menRNA and mascRNA thermostabilities.
Temperature-dependent Circular Dichroism (CD) spectra (left) and unfolding transitions (right) of menRNA and mascRNA variants in the presence of 10 mM Mg2+. The measured melting temperature (Tm) for each RNA is indicated. The RNA species are human except when explicitly labeled as mouse. AccStem: acceptor stem.
Extended Data Fig. 5 Purification of RNase P, ELAC2 and CCA-adding enzymes and kinetic analyses of cleavage of pre-tRNAGln, pre-mascRNA and pre-menRNA.
Schematics of enzyme purification of the human RNase P holoenzyme (a) and human ELAC2 (b). The purity of proteins was assessed by SDS-PAGE and silver stain (a) or Coomassie Blue staining (b). Protein annotations are based on the approximate molecular weights and previous assignments (n = 2 biologically independent replicates). c, d, Representative in vitro RNase P (c) and ELAC2 (d) cleavage analyses using the pre-tRNAGln (upper panels), pre-mascRNA (middle panels), and pre-menRNA (lower panels). Cleavage reaction progressions and rates are shown in Figs. 1h, 1i (n = 3 biologically independent replicates). e, SDS-PAGE analysis of purified bacterial, archaeal, human WT and human ΔTail CCA-adding enzymes (n = 2 biologically independent replicates).
Extended Data Fig. 6 Secondary structures of human pre-menRNA, pre-mascRNA and pre-tRNAGln used in in-cell RNase P and ELAC2 processing assays.
a, b, Secondary structures of human pre-menRNA, pre-mascRNA and pre-tRNAGln having either 5′-leaders (a) or 3′-trailers (b). The gray residues (5′-GG or 3′-UU) were appended during cloning into destination vectors. RNase P and ELAC2 cleavage sites are indicated by blue arrowheads. c, d, qRT-PCR analysis of unprocessed pre-menRNA, pre-mascRNA and pre-tRNAGln harboring either 5′-leaders (c) or 3′-trailers (d). Ordinary one-way ANOVA with Bonferroni’s multiple comparison test was performed, and P values are indicated (significant: p value < 0.05, non-significant: ns). Data are mean ± s.d. (n = 3 biologically independent replicates, indicated as individual data points).
Extended Data Fig. 7 2-aminopurine (2AP) lifetime analyses of human and mouse menRNA.
a-d, Time-resolved 2AP fluorescence decay time traces of mouse menRNA (a), the acceptor stem of the human menRNA (b) and its U57C/U58C (c) and G1A/G2A (d) mutants. IRF: instrument response function. e, Deconvoluted individual lifetimes (indicated in ns) and relative amplitudes derived from (a-d), also tabulated in Supplementary Table 1. The amplitudes for the short, intermediate, and long lifetimes are shown in violet, green, and yellow, respectively, as stacked columns. Norm.: Normalized. The amplitude-weighted averaged lifetimes τavg are summarized in Fig. 2f. Data collected on the PTI EasyLife spectrofluorometer.
Extended Data Fig. 8 2-aminopurine (2AP) lifetime analyses of human menRNA.
a-d, Time-resolved 2AP fluorescence decay time traces of the acceptor stem of the human menRNA (a), and of its C3U (b), C3A (c), and C3G (d) mutants. IRF: instrument response function (gray). e, f, Tailed and bulged conformers of various human menRNA constructs bearing 3′ CCA. The examined mutations were highlighted in red boxes. Red star denotes 2-aminopurine (2AP) in place of G56. g, Amplitude-weighted average lifetimes τavg of 2AP-containing RNAs, derived from a-d. Ordinary one-way ANOVA with Bonferroni’s multiple comparison test was performed, and P values are indicated (significant: p value < 0.05, non-significant: ns). Data are mean ± s.d. (n = 3 biologically independent replicates, indicated as individual data points). h, Deconvoluted individual lifetimes (indicated in ns) and relative amplitudes derived from a-d, also tabulated in Supplementary Table 2. The amplitudes for the short, intermediate, and long lifetimes are shown in violet, green, and yellow, respectively, as stacked columns. Norm.: Normalized. i, j Time-resolved 2AP fluorescence decay time traces of the full length (i) and acceptor stem (j) of the human menRNA in the presence of 0.25 mM Mg2+. k, Amplitude-weighted average lifetimes τavg of 2AP-containing RNAs derived from i and j. Ordinary one-way ANOVA with Bonferroni’s multiple comparison test was performed, and P values are indicated (significant: p value < 0.05, non-significant: ns). Data are mean ± s.d. (n = 3 biologically independent replicates, indicated as individual data points). l, Deconvoluted individual lifetimes (indicated in ns) and relative amplitudes derived from i and j, also tabulated in Supplementary Table 3. Norm.: Normalized. Data collected on the Horiba FluoroMax Plus spectrofluorometer.
Extended Data Fig. 9 CD analyses of Mg2+ and 2AP effects on menRNA thermostabilities.
a, b, Temperature-scanning Circular Dichroism (CD) spectra (upper) and unfolding transitions (lower) of menRNA acceptor stem variants without 2AP modifications in the presence of 0.25 mM Mg2+ (a) or 2 mM Mg2+ (b). c, Temperature-scanning CD spectra and unfolding transitions (lower) of menRNA acceptor stem variants that contain 2AP at the 56 position in the presence of 10 mM Mg2+. The measured melting temperature (Tm) for each RNA is indicated. d, Summary of Tm values of various menRNA acceptor stems in 0.25 mM Mg2+ (blue), 2 mM Mg2+ (green), or 10 mM Mg2+ (light orange, with or without 2AP). The data show that 3′ CCA and 3′ CCACCA tails, but not 3′ GGA tails, increase menRNA thermostability across different Mg2+ concentrations and in the presence or absence of 2AP modifications.
Extended Data Fig. 10 2-aminopurine (2AP) lifetime analyses of human menRNA in the presence of the CCA-adding enzyme.
a, A structural model of the archaeal CCA-adding enzyme bound to the full-length human menRNA based on the crystal structure of the A. fulgidus CCA-adding enzyme in complex with a tRNAArg minihelix (PDB: 4X4O). b, In vitro CCA addition assays of the human menRNA microhelix using the CCA-adding enzymes from E. coli, A. fulgidus or H. sapiens. A human ΔTail mutant was also examined, which removes the menRNA elbow contact with the enzyme tail domain (orange, in a). First and last lanes of each gel contain RNA length markers without proteins (n = 3 biologically independent replicates). c, In vitro CCA addition assays of the human menRNA microhelix containing a 2AP at the G56 position using the human WT CCA-adding enzyme (left) or its ΔTail mutant (right; n = 3 biologically independent replicates). d, e, Reaction progression analyses monitoring time-dependent changes of the 2AP lifetimes of the full-length menRNA (no CCA) in the presence of either WT human CCA-adding enzyme (d) or its ΔTail mutant (e) and NTPs. f, g, Amplitude-weighted average lifetimes τavg of 2AP-containing full-length menRNA (no CCA) during the time courses of CCA addition by either WT human CCA-adding enzyme (f) or its ΔTail mutant (g), derived from d and e. Data are mean ± s.d. (n = 3 biologically independent replicates). h, i, Deconvoluted individual lifetimes (indicated in ns) and relative amplitudes derived from d and e, respectively, also tabulated in Supplementary Table 4. The amplitudes for the short, intermediate, and long lifetimes are shown in violet, green, and yellow, respectively, as stacked columns. Norm.: Normalized. Data collected on the Horiba FluoroMax Plus spectrofluorometer.
Supplementary information
Source data
Source Data Figs. 1 and 2 and Extended Data Figs. 2, 4 and 6–10
Statistical source data for all main figures and extended data figures, where applicable.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Skeparnias, I., Zhang, J. Structural basis of NEAT1 lncRNA maturation and menRNA instability. Nat Struct Mol Biol 31, 1650–1654 (2024). https://doi.org/10.1038/s41594-024-01361-z
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41594-024-01361-z
This article is cited by
-
Structures of complete HIV-1 TAR RNA portray a dynamic platform poised for protein binding and structural remodeling
Nature Communications (2025)
-
Structural basis of double-stranded RNA recognition by the J2 monoclonal antibody
Nature Communications (2025)
-
iPSC-derived ITGA6-positive cells restore aqueous humor outflow in glaucoma eyes
Nature Communications (2025)
