
Abstract
Intestinal-colonizing Candida albicans is a primary source of systemic infection where it translocates across intestinal barriers into the bloodstream leading to disseminated candidiasis. To persist in the gastrointestinal tract, C. albicans must adapt to complex environments, including extreme hypoxic conditions (EHC). Here, we performed a functional genomic screen to identify genes important for C. albicans fitness under EHC. We discovered that one of the two C. albicans sterol C4-methyl oxidases, Erg251, is specifically required for producing ergosterol, an essential component for fungal membrane, in low oxygen conditions. Deleting Erg251 or mutating key amino acid residues for its function under EHC impaired C. albicans virulence and colonization in mouse models of systemic infection and commensalism, respectively. Selective inhibitors of fungal sterol C4-methyl oxidases, inhibit C. albicans growth in vitro and in a nematode infection model, showing therapeutic potential.
Similar content being viewed by others
Introduction
On a global scale, fungal pathogens are responsible for more than 6.5 million life-threatening infections, leading to 2.5 million deaths each year1. One of the most important contributors to systemic fungal disease is Candida albicans, a human commensal that primarily exists within the healthy human microbiota, colonizing various mucosal surfaces, including the skin, gastrointestinal, and reproductive tracts2,3. However, under conditions such as immune suppression or microbial dysbiosis, it can cause life-threatening systemic infections with mortality rates as high as 50%, despite antifungal treatment1. This threat is further exacerbated by the limited arsenal of antifungal drugs and the rising incidence of antimicrobial resistance4.
Intestinal-colonizing C. albicans is reported to serve as the main source of systemic infection2,3,5. During pathogenesis, C. albicans can translocate across the intestinal barrier into the bloodstream, leading to disseminated candidiasis. Within the gastrointestinal tract, C. albicans encounters complex environments including fluctuating pH and nutrient availability; host immune pressure; competition with other commensals; and importantly, hypoxic conditions. Oxygen levels in the gut are non-uniform, ranging from ~0.04% to 10%, and can even drop to near-zero levels6. During systemic infection, C. albicans also evades and persists in a variety of anatomical sites that are characterized by low oxygen levels, such as the renal medullae.
The ability of C. albicans to shift between normoxic and hypoxic growth is essential for survival and colonization in a human host, and hypoxic conditions have broad impact on C. albicans metabolism, cell wall structure, morphology, and susceptibility to antifungals7,8,9,10,11,12,13,14. Previous studies identified key transcriptional regulators involved in hypoxic adaptation in C. albicans9,10,15. Notably, under low-oxygen conditions, the transcription factor Upc2 upregulates the expression of genes involved in ergosterol biosynthesis and iron acquisition, underscoring the importance of these processes for hypoxic fitness15. Despite these insights, the genetic and regulatory networks underlying adaptation to hypoxic conditions, especially to severe hypoxic conditions, remain poorly understood.
High-throughput functional genomic screening of mutant libraries is a powerful approach to identify genes important for C. albicans fitness and virulence in host-relevant conditions16,17. The Gene Replacement and Conditional Expression (GRACE) collection is a C. albicans mutant library consisting of over 3200 strains18,19. Each strain carries a replacement of one of the two alleles of a gene with a selectable marker flanked by unique molecular barcodes, while the other allele is placed under the control of a tetracycline-repressible promoter18,19. Previous applications of this library have uncovered genes critical for fitness and pathogenesis under diverse host-relevant conditions, such as febrile temperature, serum exposure, and nutrient limitation16,17.
In this study, we screened the GRACE18,19 collection for genes differentially required for normoxic and extreme-hypoxic fitness. We identified that ERG251, one of the two sterol C4-methyl oxidase genes encoded by C. albicans, is specifically important for ergosterol production and fitness under extreme hypoxia. Loss of ERG251 compromises C. albicans virulence and colonization in mice. Through random mutagenesis, we identified critical amino acid residues governing Erg251 function in low oxygen environments. Finally, we leveraged pharmacological inhibitors of sterol C4-methyl oxidases to establish Erg251 as a potential therapeutic target. Our study provides a systematic analysis of genes essential for C. albicans survival under extreme hypoxic conditions, revealing molecular targets that offer promising avenues for antifungal development.
Results
Identification of C. albicans genes differentially required for normoxic and extreme-hypoxic fitness
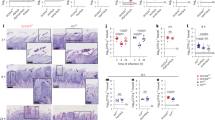
Previous studies utilizing the GRACE collection have identified a core set of essential genes in C. albicans that are critical for growth in standard laboratory conditions18,19,20. To investigate the genes important for C. albicans growth under extreme hypoxic conditions (EHC), the expanded GRACE library (GRACEv1 and GRACEv2) was spotted onto doxycycline(DOX)-supplemented agar medium and incubated in an air-tight chamber filled with gas mixture lacking an obvious source of oxygen (10% CO2, 10% H2, and ~80% N2). Growth of the macro-colonies were independently evaluated by three researchers to assign a “defect score” (DS), ranging from 0 (wild-type-like growth) to 4 (no growth) as a semi-quantitative measure of fitness. 366 strains (representing 363 genes) were identified as hits, showing an average EHC defective score (DSEHC) ≥ 2.3 (Fig. 1a). A focused re-examination validated 274 hits ( ~ 75% validation rate), among which 229 strains exhibited an overall DSEHC score ≥3 (average score from primary screen and validation), representing the core set of genes essential for EHC fitness. As a counter-test, aerobic fitness of hits was evaluated using the same scoring method (DSAC). The vast majority of genes (226/229) also played an important role for fitness in aerobic conditions (AC), as evidenced by having DSAC > 1 or the notable growth defect in the spot dilution assays (Supplementary Fig. 1a). Only ERG251 (sterol C-4 methyl oxidase), PDC11 (pyruvate decarboxylase), and ENT2 (epsin) were specifically essential for growth in EHC. To validate the conditional essentiality of the three genes, we generated homozygous deletion mutants in a prototrophic wild-type background (SC5314). Under EHC, growth of the erg251, pdc11, and ent2 deletion mutants was arrested on synthetic defined minimal medium (SD), mirroring the corresponding GRACE strains on DOX-supplemented medium (Fig. 1b and Supplementary Fig. 1a). The ent2 deletion mutant showed partially restored EHC growth on rich medium (YPD), while the growth of the erg251 and pdc11 deletion mutants remained blocked regardless of nutrient availability. Homozygous deletion of PDC11 appeared more detrimental to AC fitness compared to transcriptional depletion of the gene or deletion of ERG251 or ENT2. As a pyruvate decarboxylase, Pdc11 likely supports EHC growth by functioning with alcohol dehydrogenase (Adh) to enable fermentation21. When mitochondrial respiration is stalled by insufficient oxygen, the fermentation pathway re-oxidizes NADH, which allows glycolysis to continue and fuel the cell (Fig. 1c). Deletion of the main ethanol-generating Adh in C. albicans, Adh1 (encoded by C5_05050W)22,23, phenocopied the pdc11 mutant in blocking EHC growth. Using the adh1 mutant as a proxy, we confirmed that growth inhibition correlated with significant decreases in ATP levels and ethanol production (Fig. 1d).

a Schematic of the screening and analysis pipelines of GRACE libraries18,19. DSAC or DSEHC cut-offs for each step are provided in the “Methods”. Genes specifically required for EHC (extreme hypoxic condition) or AC (aerobic condition) fitness are highlighted in green or purple, respectively. b Growth of homozygous deletion strains were compared with wild-type (WT) parent by a spot dilution assay on YPD and SD agar. Homozygous gene deletion is denoted by italicized gene name in lower-case through this study. c Schematic of fermentation pathway and spot dilution assay showing adh1 and pdc11 mutants are blocked in EHC growth. d WT and adh1 mutant strains grown in liquid SD medium under EHC were sampled at indicated time points. Whole cell ATP levels and medium ethanol concentration were normalized to biomass determined by optical density at 600 nm (OD). LCD Limit of confident detection. e Spot dilution assay of hit strains. An overnight culture of each strain grown in SD ± 0.05 μg mL−1 DOX were serially diluted and spotted on SD agar ± 20 μg mL−1 DOX, respectively. Media were supplemented with 20 mg L−1 histidine. f AC and EHC growth of mutant strains was compared as described in (b). g, h, DOX treatment depletes tetO-driven HSF1 expression under EHC. Overnight cultures were grown in YPD ± 0.1 μg mL−1 DOX were respectively plated on YPD ± 20 μg mL−1 DOX agar and grown for 24 h under EHC before analyzed by (g) RT-qPCR or (h) anti-TAP immunoblotting. In (h), loading of the non-DOX-treated sample was titrated. The blot was probed for Hsp90 to show that Hsp90 was not depleted with Hsf1. CBS Coomassie Blue Staining for loading control. d, g show mean ± standard deviation (SD) of biological triplicates. Unpaired t-tests were performed and two-tailed p-values reported.
We next sought to identify genes that were specifically important for aerobic fitness but dispensable for growth in EHC. To do so, we leveraged a previous dataset that assessed the importance of GRACE strains for growth under AC18. These strains were re-tested under medium and temperature conditions used in the current study in both AC and EHC (Fig. 1a). Through this approach, we identified 348 genes that were required for aerobic growth (average DSAC ≥ 3 in three independent experiments). Among them, 44 genes appeared to play a less critical role in EHC growth, as their GRACE strains showed DSEHC ≤ 1 (Fig. 1a). Further validation using spot dilution assays on DOX-containing medium identified 13 strains that exhibited consistently better EHC growth than the other candidates (Fig. 1e and Supplementary Fig. 1a). These 13 genes are involved in diverse functions, including iron-sulfur cluster assembly, protein import into the mitochondria, and RNA processing. We generated homozygous deletion mutants for TRR1 and ACO1, while attempts to delete ISU1, HSF1, C3_07170C, NMD3, and CDC12 were not successful. Both trr1 and aco1 homozygous deletion mutants grew under EHC but not under AC (Fig. 1f), confirming our observations from the corresponding GRACE strains (Fig. 1e and Supplementary Fig. 1a). TRR1 encodes thioredoxin reductase, a key enzyme in redox homeostasis and reactive oxygen species detoxification24, while Aco1, the C. albicans aconitase, is a crucial tricarboxylic acid cycle enzyme. Lack of essentiality for these two genes under EHC conditions aligns with the fact that EHC is a less oxidative environment and that C. albicans shifts to fermentative metabolism under this condition. Of note, rich medium partially restored the growth of the aco1 deletion mutant under AC conditions (Fig. 1f), suggesting that its inability to grow on SD is due to a metabolic bottleneck.
Difficulty in obtaining homozygous deletion mutants for other candidate genes led us to test if their expression was efficiently depleted by DOX under EHC. HSF1, a gene with five failed deletion attempts, was used as an example to evaluate the extent of transcript and protein depletion achieved by the DOX-repressible system under EHC. We utilized a strain where one allele of HSF1 was deleted, and the other was C-terminally TAP-tagged and placed under the control of a tetO-promoter (tetO-HSF1-TAP/∆)25. Similar to the GRACE strain, growth of tetO-HSF1-TAP/∆ was only inhibited in AC, not in EHC (Supplementary Fig. 1b). Under EHC, DOX treatment decreased HSF1 RNA in the tetO-HSF1-TAP/∆ from ~800% to ~10% of the native HSF1 level, which was corroborated with depletion of the Hsf1-TAP protein (Fig. 1g, h). Based on these observations, we hypothesized that compared to AC, EHC may lower the demand for certain gene products such that even minimal expression from the tetO promoter is sufficient to support growth.
Overall, our screens identified several genes that were selectively important for C. albicans fitness in either AC or EHC due to their roles in governing metabolic adaptation and stress response signaling upon changes in oxygen availability. The remainder of the study will focus on a molecular-level dissection of the link between ERG251 and EHC fitness in C. albicans.
Erg251 is required for sterol C4-methyl oxidation under EHC
C. albicans ERG251 encodes a sterol C4-methyl oxidase, which plays an integral role in the ergosterol biosynthesis pathway. Coordinating with Erg26 and Erg27, these enzymes catalyze the oxygen-dependent removal of two C4-methyl groups from 4,4-dimethyl zymosterol (Fig. 2a)26,27,28. Despite the reported functional variation between the ERG251 A and B alleles for other traits29, either allele fully restored EHC growth of the erg251-deletion mutant (Fig. 2b and Supplementary Fig. 2a). Growth of the mutant could also be restored by reducing the severity of the hypoxic environment (Fig. 2c), confirming that the defect is contingent on oxygen level. erg251 mutant cells exposed to EHC showed decreased colony formation when moved back to the permissive AC condition, indicative of progressive loss of viability under EHC (Supplementary Fig. 2b). This stands in stark contrast to the wild-type parent, where the entire population survived switches between AC and EHC in either direction (Supplementary Fig. 2c). To test if Erg251 supports EHC growth by functioning as a sterol C4-methyl oxidase, we used the tetO promoter to drive conditional expression from both alleles (tetO-ERG251) and assessed the impact of gene depletion on sterol synthesis using Gas Chromatography-Mass Spectrometry. Without DOX treatment, the tetO-ERG251 strain was fully competent in ergosterol production in both AC and EHC (Fig. 2d). Addition of DOX to deplete ERG251 under EHC led to a significant reduction in ergosterol and, concurrently, a retention of C-4 methyl group(s). C4-dimethyl sterols such as lanosterol, eburicol, 4,4-dimethylzymosterol, as well as gramisterol, an alternative C4-monomethyl product previously found in erg251-deletion background29,30, showed significant accumulation upon 24-h DOX treatment under EHC (Fig. 2d bottom and Supplementary Fig. 2f). Extended exposure to DOX did not drastically alter the accumulation profile (Supplementary Fig. 2g). In stark contrast, ergosterol synthesis was insensitive to ERG251 depletion under AC (Fig. 2d top). Given that C4-methyl oxidase activity is essential in diverse fungi31,32,33,34, we postulated the ability of C. albicans to grow and produce ergosterol under AC in the absence of ERG251 is likely supported by ERG25, the only other C4-methyl oxidase encoded in the C. albicans genome. Indeed, depletion of ERG25 in an erg251-deletion strain inhibited growth (Fig. 2e) and C4 demethylation (Fig. 2f) in AC, resulting in a sterol profile resembling that observed with ERG251 depletion under EHC. Notably, deletion of ERG25 did not impair growth under EHC (Fig. 2g), reinforcing that Erg251 is the primary C4-methyl oxidase when oxygen availability is low.

a Schematic of ergosterol biosynthesis. b ERG251 alleles (A or B) were expressed from the native promoter. Gene re-introduction and promoter replacement were homozygous unless otherwise specified. c Spotting assay showing O2-level contingent growth of erg251 on SD. d Sterol profiles of strains grown on SD agar ± 20 µg mL−1 DOX analyzed by GC-MS after 8-h AC or 24-h EHC incubation. ns not significant (p-values: 0.88; 0.80; 0.42) e The tetO-promoter was used to drive ERG25 in WT (ERG251) and erg251 backgrounds. AC growth was assessed on YPD ± 20 µg mL−1 DOX. f Sterol profiles of the erg251 tetO-ERG25 strain after 24-h EHC growth on SD agar ± 20 µg mL−1 DOX. g Spotting assay showing ERG25 is dispensable for fitness. h Expression of ERG251 was assessed after 6-h of AC or EHC growth on YPD. i Immunoblotting comparing expression of Erg251-GFP and Erg25-GFP from their native promoter after 9-h EHC growth on YPD. Loading of the Erg251-GFP sample was titrated. *: adjusted image for visualizing Erg25-GFP signal; Coomassie Blue Staining (CBS). j Sequence and position of putative Upc2 binding sites. Highlighted “CG” were changed to “AT” in the mutant promoter (pupc2_mut.) and expression measured after 6-h EHC growth on YPD. Analyses in this panel were based on ERG251(B). k–m ERG25(-GFP) expression driven by the ERG251 promoter (p251) or tetO promoter was compared to ERG251(-GFP) using RT-qPCR (k) and immunoblot (l). Growth was tested under different O2 levels on YPD via spotting assay (m). P-values comparing ERG25 expression are reported in (k). n The erg251-deletion strains re-introduced with p251-driven ERG251 or ERG25 were grown for 24 h on SD agar under EHC. Sterol profiles were analyzed by GC-MS. o Phylogenetic tree of fungal sterol C4-methyl oxidases. p Sterol C4-methyl oxidase genes were expressed from the native C. albicans ERG251 promotor (p251) in an erg251 tetO-ERG25 background (“Parent”). Cell growth was assessed on YPD + 20 µg mL−1 DOX. q Independent C. auris transformants (#1, #2) were tested to assess growth via spotting assays. r Transcript levels were measured after 7-h AC or 9-h EHC growth on YPD. s Graphic summary. d (AC part), f, h, j, k, and r show mean ± SD of biological triplicates examined by unpaired t-tests (two-tailed p-values reported); d (EHC part) and n show biological quadruplicates (mean ± SD) examined by Mann–Whitney tests (two-tailed p-values reported).
To elucidate why ERG251 is the primary functional sterol C4-methyl oxidase in EHC, we compared the expression of ERG251 and ERG25 under low and high oxygen conditions. In contrast to cells grown on fully aerated agar, which exhibited higher ERG25 expression, cells grown under EHC selectively transactivated ERG251 (Fig. 2h). Immunoblot analyses of functional C-terminal GFP tagged versions of the two proteins revealed an even greater difference in protein levels under EHC (Fig. 2i and Supplementary Fig. 2d, e). The transcriptional induction of ERG251 was at least in part dependent on Upc2, a master regulator of ergosterol biosynthesis genes35, as disruption of the five Upc2 binding consensus sequences (“TCGNNTA”) in the ERG251 promoter significantly reduced EHC-dependent induction (Fig. 2j). To test if overexpression renders ERG25 competent in supporting EHC growth, we placed ERG25 (or the GFP-tagged version) under the control of the ERG251 promoter. Despite the drastic increase in ERG25 transcript and protein levels, this chimeric construct only partially restored EHC growth of the erg251 deletion strain (Fig. 2k–m), consistent with the unremedied C4-methyl retention in this strain (Fig. 2n). A similar growth phenotype was observed when ERG25 overexpression was driven by an unrepressed tetO promoter (Fig. 2k and m). Overall, our results suggest ERG251 is induced in a Upc2-dependent manner in EHC and possesses more adaptive enzymatic properties to support EHC growth and ergosterol synthesis under low oxygen conditions.
ERG251 orthologs from diverse Candida spp. show conserved advantages under EHC
Conserved syntenic homologs of ERG251 and ERG25 are present in other pathogenic Candida species such as C. dubliniensis, C. parapsilosis, and C. tropicalis (Fig. 2o). To evaluate their respective roles in supporting EHC growth, we introduced ERG251 or ERG25 from other fungal species, driven by the C. albicans ERG251 promoter, into the C. albicans erg251 tetO-ERG25 strain (hereafter referred to as “Parent”). When the endogenous ERG25 was depleted by DOX, ERG251 or ERG25 from C. dubliniensis, C. parapsilosis, and C. tropicalis as the sole C4-methyl oxidase supported robust growth in AC (Fig. 2p). When tested under EHC, all three ERG251 orthologs consistently supported better growth than the ERG25 orthologs from the same origin species. In contrast to the three closer relatives of C. albicans, Nakaseomyces glabratus (formerly Candida glabrata) and the model yeast Saccharomyces cerevisiae encode ERG25 as their only sterol C4-methyl oxidase. Despite the ERG25 designation, the two genes have greater sequence similarity to ERG251 orthologs (Fig. 2o), and are capable of supporting growth in both AC and EHC when expressed in C. albicans (Fig. 2p). Finally, functional comparison of the ERG251 and ERG25 orthologs of the emerging pathogen Candidozyma auris was hindered because the latter functioned poorly in the heterologous expression system as evidenced by the inability to support growth of the “Parent” under AC (Fig. 2p). Therefore, we generated and characterized erg251 and erg25 deletion mutants of C. auris to investigate the importance of these enzymes in supporting growth in different oxygen environments. Similar to the C. albicans homologs, C. auris ERG25 was nonessential in both AC and EHC, whereas C. auris ERG251 was essential in low oxygen environments (Fig. 2q). The increased dependency of C. auris fitness on ERG251 in EHC correlated with its induction under this condition (Fig. 2r), suggesting that C. auris also applies transcriptional regulation of ERG251 to adapt to growth environments (Fig. 2s). Taken together, our results support a conserved role of ERG251 orthologs in adaption to EHC in diverse human fungal pathogens.
ERG251 supports C. albicans pathogenesis
Adaptation to hypoxic conditions is crucial for C. albicans colonization and its ability to cause invasive infections. To assess if ERG251 is required for C. albicans pathogenesis, we revisited the role of ERG251 in biofilm formation, a key virulence trait of C. albicans30,36. A previous study discovered deletion of ERG251 abolished biofilm growth and speculated a role of the hypoxic condition in causing this phenotype30. This reported biofilm defect was recapitulated using our independently generated erg251 homozygous deletion mutant and was rescued by the re-introduction of the wild-type ERG251 allele (Fig. 3a). In addition, biofilm formation could be restored by reducing the growth medium by half to improve oxygenation (Fig. 3a). To further interrogate whether Erg251 was critical for biofilm formation or simply growth in poorly oxygenated liquid medium, we cultured C. albicans sterol C4-methyl oxidase mutants in YPD under various aeration conditions. Robust planktonic growth of the erg251 mutant required a combination of low culture volume and agitation to maximize oxygen availability, highlighting insufficient aeration cripples erg251 proliferation in liquid culture (Fig. 3b). Next, we assessed the role of ERG251 in C. albicans virulence using a mouse model of invasive candidiasis. Female BALB/c mice were retro-orbitally infected with either wild-type (SC5314), erg251 homozygous mutant, or complemented strains. In the absence of ERG251 mice showed significantly delayed death compared to mice infected with the wild-type or complemented controls (Fig. 3c), similar to what was noted in a previous study29. The slower disease progression of the mutant-infected mice correlated with lower fungal burden in the kidney and liver at early stages of infection, as quantified by colony forming units (CFUs) recovered from tissue homogenates and further supported by histological staining with an anti-Candida antibody (Fig. 3d, e). Aligning with the lower fungal burden, anti-CD45 staining revealed lower immune infiltration in the mutant-infected kidneys. Detection of the filaments in the erg251 mutant by Periodic Acid-Schiff staining (Fig. 3f) suggests that morphogenesis, an essential virulence trait, is not fully blocked in the absence of ERG251, consistent with the eventual progression of infection to a fatal outcome with the mutant strain.

a Biofilm growth of indicated strains induced in 100 μL or 200 μL medium in 96-well plates was measured by metabolic activity using an XTT reduction assay. Data showed mean ± SD of biological triplicates. ns not significant (p-values: 0.45; 0.15; 0.18) b 100 µL or 200 µL of YPD culture of each strain was grown in 96-well plates with or without agitation (~ 170 rpm). Experiments were repeated twice and gave similar results. Data present mean ± SD of technical octuplicates of one experiment. The WT and erg25 curves were respectively nudged +0.04 and +0.02 units along the y-axis for visualization. c–e Immunocompetent BALB/c female mice were infected with the indicated C. albicans strains by retro-orbital injection. Survival was monitored for four weeks (c; n = 8). Fungal burden (CFU per gram tissue) in kidney and liver was assessed three days post-infection (d; n = 10). Contiguous kidney sections (three days post-infection) were analyzed by immunohistochemical staining using α-Candida and α-CD45 antibodies (e). Tissues were visualized by H&E. ns in (c): not significant (p-values: 0.16; 0.20) f Sections from the kidneys in (e) were analyzed by Periodic Acid-Schiff (PAS) staining. Yellow arrowheads indicate filaments. g The erg251-deletion mutant and the A-allele-complemented strain were introduced as a 1:1 mixed inoculum to female BALB/c mice (n = 4). Relative strain abundance in the feces was monitored every 5 days for 20 days using qPCR. Lines indicate medians. Statistical tests performed: unpaired t-tests (a; two-tailed p-values reported); Mantel–Cox tests (c); Mann–Whitney tests (d and g; two-tailed p-values reported).
C. albicans is capable of colonizing various sites of the gastrointestinal tract, including regions where oxygen is extremely limited. To characterize the role of ERG251 in C. albicans commensalism, we compared the erg251 deletion mutant with a complemented strain through a competition assay in a mouse model of stable gastrointestinal colonization37. The two strains were introduced at ~1:1 ratio into mice, and relative strain abundance in the feces was monitored every five days for three weeks. The erg251 deletion mutant was rapidly outcompeted by the complemented strain and showed persistent depletion over the course of the experiment (Fig. 3g). Overall, these results highlight the vital role of ERG251 in both virulence and colonization of C. albicans in mouse models.
Random mutagenesis screen identifies mutations disabling Erg251 function under EHC
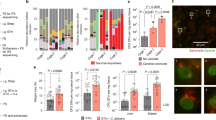
Given the critical role of Erg251 in governing C. albicans pathogenesis and commensalism in a mammalian host, we wanted to define distinct features of Erg251 that enable its adaptation to limited oxygen. To do so, we performed a random mutagenesis using error-prone PCR and introduced our mutant library into the C. albicans erg251 tetO-ERG25 “Parent”, the same background used for expressing heterologous sterol C4-methyl oxidases. Transformants were first selected on medium supplemented with both nourseothricin and DOX for integration of AC-permissive alleles (Fig. 4a). Colonies were then assessed for fitness in EHC through replica plating. Colonies with robust AC growth but poor EHC growth, indicative of preserved Erg251 function in AC but compromised function in EHC, were selected and named as BinGo (Bad in chamber, Good outside chamber) (Fig. 4a). These ERG251 alleles were cloned and sequenced to identify candidate mutations. We identified a total of twenty-six BinGo homozygous mutants that had integration of the same allele at both ERG251 loci. Reintroducing the ERG251 BinGo alleles into the “Parent” strain faithfully recapitulated the phenotype of the original isolates (Supplementary Fig. 3a). With the exception of erg251-1, which had a non-synonymous substitution of K93E, all other alleles contained multiple missense mutations or truncations (Fig. 4b). By individually reverting >60% of the mutations back to the wild-type sequence, we identified at least one mutation for each allele that made major contributions to the BinGo phenotype (Fig. 4b and Supplementary Fig. 3b).

a Strategy to identify erg251 BinGo mutants. Error-prone PCR pool of ERG251 fused with a NAT resistance marker was introduced into the “Parent” strain. Colonies that grew robustly on YPD + NAT + DOX under AC, but poorly under EHC were selected b Missense mutations on the prioritized BinGo alleles. Magenta, light green, or black, respectively, color-code major, minor, or insignificant improvement of EHC fitness when the indicated mutation was back-mutated. Grey: untested. c A predicted model of Erg251 suggests locational enrichment of mutations making major contributions to BinGo phenotypes. Sites highlighted magenta in (b) are marked. Yellow: transmembrane motifs predicted by TMAlphaFold (TM1 to TM5). Yellow dots count the number of times for major contributing mutations identified at a given residue. d Mutants sufficient to drive a notable BinGo phenotype and their predicted spatial position relative to Fe2. AC and EHC growth of WT and mutant strains were tested on YPD + 20 µg mL−1 DOX. e Targeted mutagenesis identifies additional BinGo substitutions. Each mutant was expressed in the “Parent” background and growth on YPD + 20 µg mL−1 DOX was assessed under both AC and EHC. f Spot dilution of erg25-deletion strains expressing mutant (homozygous substitution) or WT ERG251 on YPD under AC and EHC. AC growth was tested at both 37 °C and 30 °C to determine temperature-dependent defects. GC-MS analyses confirm comparable ergosterol content in WT and mutant strains. ns not significant (p-values:0.29; 0.11) g The native promoter-driven WT ERG251 or BinGo mutants was expressed in a background where one copy of ERG251 was deleted and the other controlled by a tetO promoter (tetO-ERG251/Δ). Strains were grown on SD + 20 µg mL−1 DOX under EHC before collection for sterol profiling by GC-MS. Detailed information for (b, d, and e) are provided in Supplementary Figs. 3b and 5a, b. Sterol contents in (f and g) show mean ± SD of biological triplicates. Unpaired t-tests were performed and two-tailed p-values reported. Exact p-values in (g) were provided in Source Data.
No solved structure of any C4-methyl oxidase has been reported to date. To propose a 3D map of the BinGo-contributing mutations, we generated a model of iron-bound Erg251 using AlphaFold3 and the transmembrane domain prediction database TmAlphaFold38,39. This model predicts that Erg251 is an integral membrane protein with five transmembrane helixes and a diiron center in which two iron atoms (annotated as Fe1 and Fe2) are coordinated by five and four histidine residues, respectively (Fig. 4c and Supplementary Fig. 4a). The overall topology and the structure of the diiron center resembled the solved structures of other members in the same superfamily: mammalian stearoyl-Coenzyme A desaturase 1 (SCD1; PDB: 6WF2; Supplementary Fig. 4b) and yeast sphingolipid α-hydroxylase (Scs7; PDB:4ZR1)40,41. In support of the structural model, mutating any of the predicted iron-interacting histidine residues to leucine resulted in loss of function (Supplementary Fig. 4c). According to this model, mutations making major contributions to the BinGo phenotype are enriched in the cytoplasmic region near Fe2, but less often seen in the membrane-embedded region or the ER-facing region of the protein (Fig. 4c). When individual BinGo-contributing mutations were introduced into the wild-type ERG251 allele, >80% of them showed limited or no impact on EHC growth, suggesting that the phenotype of most BinGo alleles are composite effects from multiple mutations (Supplementary Fig. 5a). However, substitutions K93E, C108F, S248F, G251S, or Y252N alone were sufficient to specifically impair growth in EHC (Fig. 4d and Supplementary Fig. 5a). On the predicted Erg251 model, all five sites are located in the hotspot near Fe2 (Fig. 4c, d). To diversify changes at these sites, we made additional substitutions varying in size and polarity. Some changes were selected to mimic the changes identified by the screen (e.g., K93D vs K93E and S248L vs S248F), while others were selected to investigate properties divergent from the screen hit (e.g., K93F vs K93E and Y252A vs Y252N). Substitutions detrimental to Erg251 function under both AC and EHC conditions were identified as well as BinGo substitutions for K93, S248, and Y252, but not for G251, C108 (Fig. 4e and Supplementary Fig. 5b). We also generated additional substitutions for S250, a non-hit residue in the screen but proximal to the hotspot, however we did not identify any substitutions specifically impairing fitness under EHC (Fig. 4e and Supplementary Fig. 5b). None of the mutants were haplo-insufficient for AC growth, but reducing gene dosage from two alleles to one exacerbated fitness defects under EHC (Supplementary Fig. 5a). Of note, no mutations from this expansion caused AC-specific fitness detriments, indicating that Erg251 function under EHC is more vulnerable to structural perturbation.
BinGo mutations of ERG251 reduce C-4 methyl oxidation under EHC and C. albicans pathogenesis in mice
The Erg251K93E and Erg251Y252A substitutions were chosen to represent BinGo mutants identified by random and targeted mutagenesis for further characterization. Both mutants were recessive for the EHC growth defect and could be stably expressed under these conditions (Supplementary Fig. 5c, d). To further assess the functionality of Erg251K93E and Erg251Y252A, we deleted ERG25 in each strain. Left as the sole sterol C4-methyl oxidase gene, each mutant remained sufficient for normal growth and ergosterol production in AC (Fig. 4f). In contrast, when each mutant allele was expressed in heterozygously with a tetO-driven wild-type ERG251, addition of DOX to deplete the wild-type allele led to accumulation of C-4 methyl sterols (Fig. 4g), highlighting that K93 and Y252 are necessary for proper sterol production in low oxygen environments. To assess whether these specific amino acids were also necessary for C. albicans virulence, we tested the mutants for in vitro biofilm growth and in vivo fitness. We observed strains with Erg251K93E and Erg251Y252A phenocopied the erg251 homozygous deletion mutant, causing significant reductions in in vitro biofilm formation and decreased competitive fitness in kidney infection and gastrointestinal tract colonization (Fig. 5a–c). Overall, recapitulation of the erg251 deletion phenotypes with the EHC-sensitive BinGo mutants suggests that adaptation to hypoxic environments in the host is crucial for C. albicans virulence and colonization, and this process is highly dependent on Erg251.

a Biofilm growth was induced in 200 μL of medium in 96-well plates. Results present mean ± SD of XTT reads from biological triplicates. b, c Mice were infected with ~1:1 mixed inoculum of barcoded strains expressing WT ERG251 or one of the BinGo mutants. In the systemic infection model (b; n = 4), relative kidney fungal burden of each strain was assessed three days post-infection by qPCR. In the commensalism model (c; n = 5), relative strain abundance in the feces was monitored every 5 days for 25 days. d Ferrous (Fe2+) supplementation restores EHC growth to BinGo mutants. Spot dilution assays were performed under EHC. e, f Strains expressing WT ERG251 or the BinGo mutants as the sole source of sterol C4-methyl oxidase were tested via disk diffusion assay against cobalt on Hinton-Mueller agar (HMA) and BPS on ascorbic acid supplemented HMA (e). Annular radii (AR; in millimeter) of the cobalt inhibition zones were measured from biological quintuplicates (f; medians marked). Sensitivity to BPS was compared visually. Mutants showed clear and consistent hypersensitivity in multiple experiments and representative images shown in (e). g erg251-deletion strains re-introduced with either the native ERG251 or p251-driven ERG25 were tested for EHC growth with or without FeSO4 supplementation. h, i Strains expressing ERG251 or ERG25 from the ERG251 promoter as the sole source of sterol C4-methyl oxidase were tested in the same format as the assay in (e). AR of the cobalt inhibition zones from biological quadruplicates were plotted with the medians marked. ns not significant (p-value: 0.11). No noticeable differences in sensitivity to BPS were observed in multiple tests. Statistical tests performed: unpaired t-tests (a; two-tailed p-values reported); Mann–Whitney tests (b, c, f, and i); two-tailed p-values reported.
BinGo substitutions K93E and Y252A may sensitize Erg251 to low oxygen via destabilizing Fe2
Previous studies identified the four-histidine-coordinated Fe atom (Fe2) as a major liability of purified SCD1, with loss of Fe2 during the catalytic cycle resulting in enzyme inactivation42. Given that all identified BinGo point mutations showed spatial proximity to Fe2, we hypothesized that these mutations may impair Erg251 function by destabilizing Fe2. We tested this hypothesis by examining if increasing iron availability could rescue the BinGo mutants under EHC. Low oxygen allows for supplementing ferrous at supra-physiological concentrations without causing toxicity. As hypothesized, ferrous supplementation improved growth of the erg251K93E and erg251Y252A mutants in low oxygen conditions in a dose-dependent manner (Fig. 5d). Similar rescuing effects were observed for the other three BinGo mutants tested: erg251Y252N, erg251C108F, and erg251K93F (Fig. 5d). In line with these results, when erg251K93E and erg251Y252A were the only sterol C4-methyl oxidase in the cell, mutants were hypersensitive to iron deprivation. In disk diffusion assays performed under AC, iron-competing or chelating agents, cobalt and bathophenanthroline disulfonate (BPS), produced larger inhibition zones on the mutant strains than on the wild-type (Fig. 5e, f). Collectively, our genetic evidence supports that the BinGo mutations interfere with Fe binding, which becomes devastating for Erg251 function under oxygen- or iron-deprived conditions.
In parallel, we assessed the impact of increasing iron availability on Erg25 function under EHC. In contrast to the growth-restoring effects observed in the BinGo mutants, Fe supplementation under EHC did not rescue growth of the erg251 deletion mutant (Fig. 5d) or improve the partial growth observed with the ERG25 overexpression strain (Fig. 5g). In addition, the ERG25 overexpression strain was not hypersensitive to cobalt or BPS, but showed slight hyposensitivity to low cobalt concentrations compared to the strain expressing ERG251 (Fig. 5h, i). Therefore, unlike the BinGo mutants, Erg25 functionality is insensitive to alteration in iron availability.
Targeting sterol C4-methyl oxidases presents a promising therapeutic approach to combat C. albicans infections
The pivotal role of ERG251 in C. albicans pathogenesis suggests this sterol C4-methyl oxidase may serve as a promising therapeutic target for antifungal development. Among the known fungal sterol C4-methyl oxidase inhibitors43,44,45, the depsipeptide PF1163A exhibits potent bioactivity against a broad spectrum of human fungal pathogens and demonstrates synergy with azoles in vitro43,46. We performed a comparative study to assess the respective roles of ERG251 and ERG25 in governing susceptibility to this inhibitor. Through disk diffusion assays, we observed that while PF1163A inhibited growth of the wild-type strain and erg251 homozygous deletion mutant to a similar extent, it was significantly more potent against the erg25 deletion mutant (Fig. 6a). To understand this differential sensitivity, we compared expression levels of ERG251 and ERG25 following PF1163A exposure. Transcript levels of ERG251 rapidly increased through Upc2-dependent transactivation, becoming the predominant sterol C4-methyl oxidase (Fig. 6b and Supplementary Fig. 6a). Overexpression of ERG25 from the inducible ERG251 promoter conferred resistance to PF1163A (Fig. 6c). Similar reductions in PF1163A susceptibility were observed in strains expressing ERG25 from C. dubliniensis, C. tropicalis, or C. parapsilosis, but not in those expressing the ERG251 orthologs (Fig. 6d). Overall, these results suggest that Erg251 in multiple Candida species is more susceptible to PF1163A, while Erg25 showed conserved greater tolerance to this inhibitor.

a Representative disk diffusion assay results of PF1163A (structure shown) against WT, erg251, and erg25 strains. Quantified size of inhibition zones was shown by the AR in millimeter (lines: medians of biological quadruplicates). ns not significant (p-values: 0.6; 0.89; 0.2) b Gene expression was measured from log-phase cultures treated with DMSO or 20 μM PF1163A for 75 min. c Disk diffusion assay with the same layout as in (a). d, e Sensitivity of Candida ERG251 and ERG25 orthologs to PF1163A and 1181-0519. C. albicans strains expressing p251-driven fungal ERG251 or ERG25 as the sole source of sterol C4-methyl oxidase were tested for susceptibility to 10.5 nmol PF1163A (d) or 50 nmol 1181-0519 (e) in disk diffusion assays. f SC5314 was tested in a dual-disk diffusion assay with PF1163A and 1181-0519 at the indicated dose. The experiment was repeated three times with comparable results. g Strains were grown in 200 µL of RPMI + 10% FBS at 37 °C for 24 h and treated with DMSO or PF1163A. Metabolic activity of biofilm was measured using by XTT reduction. h Co-culture of C. albicans and luciferized human HEK293T (HEK293T-fLuc) cells were treated with DMSO or PF1163A for 24 h. Fungal burden was determined by OD and HEK293T-fLuc survival by luminescence. i J774A.1 cells infected with C. albicans were treated with DMSO or PF1163A. Macrophage death stained by propidium iodide (PI) was quantified normalized to DMSO controls. Macrophage Survival = 1- normalized PI area. j C. albicans infected C. elegans (AU37, glp-4 sek-1) was treated with either PF1163A or DMSO and survival monitored for 96 h. ns not significant (p-value: 0.68). b, d, and e show mean ± SD of biological triplicates. g–i show mean ± SD of biological triplicates and curves fit by nonlinear regression. Statistical tests used: Mann–Whitney tests (a, d, and e); two-tailed p-values reported), unpaired student t-tests (b; two-tailed p-values reported), and Mantel–Cox tests (j). In (d, e, g–i), SDs were not shown if smaller than the limit for proper display.
To assess whether Candida sterol methyl oxidases also show differential sensitivity to other inhibitors, we turned to 1181-0519, a hydrazone-containing scaffold that is structurally-divergent from PF1163A44. Though two to fourfold less active against C. albicans compared to PF1163A (Supplementary Fig. 6b), 1181-0519 treatment of strains expressing either ERG25 or ERG251 from multiple Candida species resulted in less differential zones of inhibition in disc diffusion assays (Fig. 6e). The ability of 1181-0519 to inhibit both enzymes to a similar extent suggests a potential synergy with this compound and PF1163A. Indeed, spotting the individual compounds on discs in close proximity selectively abolished C. albicans growth between the two discs where both compounds had diffused into the agar, suggesting the potential of leveraging inhibitor cocktails to ablate sterol C4-methyl oxidase activity (Fig. 6f).
Finally, to demonstrate that targeting sterol C4-methyl oxidase is a promising antifungal strategy, we assessed if PF1163A treatment could impair C. albicans virulence traits. As predicted, PF1163A inhibited biofilm formation in vitro at a concentration as low as 1 μM (Fig. 6g). The efficacy of PF1163A was also tested in two C. albicans co-culture models: luciferase-expressing HEK293T cells and J774A.1 (J7) macrophages. In the first assay, PF1163A inhibited C. albicans growth and recused human cell viability at 2 μM, a concentration that conferred undetectable mammalian cytotoxicity (Fig. 6h and Supplementary Fig. 6c). In co-culture with J7 macrophages, PF1163A prevented macrophage killing by fungal cells (Fig. 6i). Unfortunately, the metabolic instability of PF1163A precluded testing in a mouse infection model (Supplementary Fig. 6d). Therefore, we assessed PF1163A efficacy in an alternative nematode model of fungal infection. At 20 µM, a concentration that showed no host toxicity, PF1163A significantly improved survival of C. albicans-infected Caenorhabditis elegans (Fig. 6j), supporting the potential of sterol C4-methyl oxidase inhibition as a therapeutic approach for combating C. albicans infections.
Discussion
The growing threat of antifungal resistance, coupled with a limited therapeutic arsenal highlights the urgent need to expand our understanding of fungal biology to uncover additional promising antifungal targets. In this study, we utilized a functional genomic screening strategy designed to mimic the physiologically relevant extreme hypoxic condition to uncover environment-specific vulnerabilities of C. albicans. In addition to the genetic signature for metabolic plasticity our initial functional genomic screens revealed, we uncovered how C. albicans tailors sterol synthesis to adapt to low-oxygen environments, specifically through the function of the sterol C4-methyl oxidase, Erg251. When oxygen is scarce, C. albicans exclusively relies on Erg251 to complete sterol C4-methyl oxidation, the most oxygen-consuming step of ergosterol biosynthesis. Deletion of ERG251 or specifically disabling its function via the identified “BinGo” mutations decreased C. albicans virulence and colonization in mice, highlighting that adaptability to hypoxia plays a central role in C. albicans pathogenesis and commensalism.
The remarkable metabolic robustness of C. albicans under EHC is dependent on efficient and balanced utilization of precious oxygen molecules for the oxygen-dependent synthesis of essential building blocks, including unsaturated fatty acids, heme, and sterols. In other species, including S. cerevisiae and Aspergillus fumigatus, insufficient sterol C4-methyl oxidase activity leads to the accumulation of the enzyme substrates, 4,4-dimethyl zymosterol or 4,4-dimethyl fecosterol34,47. In contrast, disabling C4-methyl oxidation in C. albicans lead to the significant accumulation of the Erg11 substrate lanosterol, suggesting that C-14 and C-4 demethylation are coordinated in this pathogen. The negative feedback regulation on Erg11 appeared to require severe impairment of C4-methyl oxidase activity as less severe hypoxic conditions, which allow partial function of Erg25, did not lead to an accumulation of lanosterol in an erg251 mutant30. In Schizosacchromyces pombe and Cryptococcus neoformans, C4-methyl sterols directly regulate sterol synthesis by conveying signals to the sterol-sensing SREBP orthologs48,49. Whether certain C4-methyl sterol(s) have direct impacts on Erg11 function in C. albicans remains to be elucidated. Consistent with previous findings29,30, impairment of C. albicans C4-methyl oxidase activity also leads to the accumulation of gramisterol. This C4-monomethyl sterol, likely produced by Erg6 and Erg2 from 4α-methyl zymosterol, increases to a greater extent than 4,4-dimethylzymosterol, suggesting that the second C4-methyl group is less favorable for oxidation. Coincidently, wild-type C. albicans grown under EHC showed a small buildup of 4α-methyl zymosterol (Fig. 2d), further supporting that the removal of the second C4-methyl group could be a rate-limiting step of sterol biosynthesis in sub-optimal environments.
Sterol C4-methyl oxidases belong to the membrane-integrated fatty acid hydroxylase super family, which share a diiron center fully coordinated with histidine residues. In the solved structures for SCD1 and Scs740,41, such an architecture features a long distance (> 6.2 Å) between the diiron, corroborating the computational model-derived distance (5.4 Å) between Erg251’s Fe1 and Fe2. The identification and characterization of BinGo mutants provided valuable insights for sterol C4-methyl oxidase reactivity, particularly highlighting the role of the four histidine-coordinated Fe atom (Fe2). Previous work using purified SCD1 discovered that Fe2 is exchangeable with environmental iron and prone to loss during catalysis42. For Erg251, Fe2-perturbing mutations at K93 and Y252 selectively abolished activity under EHC. One plausible model aligning with both observations is that during catalysis, an intermediate phase requires Erg251 to transiently adopt a Fe2-destabilizing conformation, and completion of this phase requires oxygen engagement. Limited oxygen availability makes Fe2 stall longer in such a vulnerable state and in turn, become more susceptible to destabilizing mutations. Such a model is supported by the fact that high concentrations of ferrous ion in medium restores growth of the BinGo mutants in EHC. In contrast, Erg25 does not show improved function upon iron supplementation, suggesting that its poor activity under EHC is not attributable to diiron center instability. Possible mechanisms underlying Erg251’s superior function under low oxygen environments includes higher affinity to oxygen. On the other hand, sterol C4-methyl oxidation is a highly coordinated process, which requires sterol C4-methyl oxidases to interact with the upstream (cytochrome b5 as an electron carrier) and downstream (Erg26 and Erg27) components27,34,50,51. Compared to Erg25, Erg251 might function better in this complex under EHC. The two possible mechanisms are not mutually exclusive.
Among the C. albicans ergosterol synthetic enzymes, sterol C4-methyl oxidase is uniquely encoded by paralogs. Compared to ERG251, ERG25 seems less versatile in function or responsive to Upc2-mediated transactivation. However, sequentially and functionally conserved ERG25 orthologs were found not only in the close relatives of C. albicans, but also in more divergent species such as C. auris. From an evolutionary perspective, the selective advantage to have two “flavors” of sterol C4-methyl oxidases remain unclear. However, the observation that Erg25 was hyposensitive to the Penicillium product PF1163A compared with Erg251 hints that this enzyme might outperform Erg251 upon challenge by inhibitory xenobiotics generated by other microbes. Our heterologous expression assays also suggest that the only C4-methyl oxidase encoded by S. cerevisiae retains activity under EHC. As a “Crabtree effect” positive yeast, S. cerevisiae metabolizes sugar through alcoholic fermentation even in the presence of plentiful oxygen, which was thought to be an adaptation to oxygen-limited environments during evolution52. The property of S. cerevisiae Erg25 appears to align with this speculated evolutionary path.
The ergosterol biosynthesis pathway has been the central focus in antifungal drug development. Azoles, which inhibit 14α-demethylase (Erg11), have served as the standard of care in clinic for decades. However, the rise of resistance has prompted interest in exploring alternative targets within the pathway. Using PF1163A as a probe and a nematode infection model, we showed that the sterol C4-methyl oxidases can be a promising antifungal target. The preferential inhibition of PF1163A on Erg251 orthologs, along with its synergy with 1181-0519, not only provides evidence that the two Candida sterol C4-methyl oxidases can exhibit differential susceptibility to pharmacological inhibition but also underscores inhibition of both Erg251 and Erg25 as a versatile strategy to enhance compound efficacy. Overall, functional insights and genetic resources from our study pave the way for an integrated platform for assessing sterol C4-methyl oxidase inhibition strategy for antifungal development.
Methods
Strains and culture conditions
All strains were archived in 25% glycerol at −80 °C for long term storage. For experiments, strains were freshly streaked on YPD agar plates and kept at room temperature without refrigeration for no longer than a week. The pdc11 and adh1 homozygous deletion mutants were maintained on YPGalactose for better growth. YP-based medium is composed of 1% yeast extract, 2% peptone, and 2% glucose or galactose. The basic recipe for each liter of synthetic defined minimal medium (SD) is 1.7 g yeast nitrogen source (without ammonium sulfate), 1 g monosodium glutamate hydrate (MSG), and 20 g glucose. SD medium used to grow GRACE strains was with 1% glucose and supplemented with 20 mg L−1 histidine. For sterol profile analyses, ammonium sulfate (5 g L−1) instead of MSG was used as the nitrogen source to minimize filamentous growth and in turn, facilitate cell harvest. Solid medium contained 1.5 to 2% agar. DOX (Bio Basic) was added at the indicated concentrations from a 50 mg mL−1 aqueous stock. EHC in this study is referred to as the condition in an AS-580 chamber (Anaerobe Systems) running under oxygen-free gas mixture (10% CO2, 10% H2, and 80% N2) at 37 °C. The less stringent hypoxic conditions (0.5–2.5% O2) were achieved using the nBIONIX pouch system (Bulldog Bio). For iron supplementation under EHC, 100× ferrous sulfate solution (20, 80, 160 mM; Bioshop) was made in deoxygenated water and spread on 30 mL YPD plates.
Arrayed growth screen of the GRACE collection under AC and EHC conditions
To identify genes required EHC fitness, individual strains were pinned into 96-well plates containing liquid SD medium with 0.5 μg mL−1 of DOX (stock in H2O, Bio Basics) and grown overnight at 30 °C. Overnight cultures were 1:20 diluted in sterile H2O and spotted onto SD agar supplemented with 20 μg mL−1 of DOX. Plates were incubated under EHC at 37 °C for five days and growth was independently scored by three researchers using a 0 to 4 scale (0: wild-type-like growth; 4: no growth; 1–3: increasing degrees of growth defects). The average score was denoted as “DSEHC_0.” Strains showing DSEHC_0 ≥ 2.3 were re-arrayed and re-tested for EHC fitness (as validation) and AC fitness (as the counter-test) on DOX-supplemented medium. The GRACE strains for the genes that were found important for AC fitness in a previous study18 (score >1 on DOX-containing plates) were tested in parallel. This experiment was repeated in biological triplicate, resulting in three determinations of DSEHC (DSEHC_1; DSEHC_2; DSEHC_3) and DSAC (DSAC_1; DSAC_2; DSAC_3) for each strain. Cut-offs for the left arm in Fig. 1a: Screen: DSEHC_0 ≥ 2.3; Validation: mean (DSEHC_1, DSEHC_2, DSEHC_3) ≥ 2.3; Prioritization: mean (DSEHC_0, DSEHC_1, DSEHC_2, DSEHC_3) ≥ 3; AC fitness filter: mean (DSAC_1; DSAC_2; DSAC_3) ≤ 1. Cut-offs for the right arm in Fig. 1a: Fitness test at 37 °C: mean (DSAC_1; DSAC_2; DSAC_3) ≥ 3; EHC fitness filter: mean (DSEHC_1, DSEHC_2, DSEHC_3) ≤ 1. All DS scores are provided in Supplementary Data 1 and 2.
Strain construction
Names and genotypes of all strains used in this study were provided in detail in Supplementary Data 3. The panel(s) where each strain was used was also annotated. For C. albicans, homozygous genetic modification was achieved using a transient CRISPR method53 with standard lithium-PEG (polyethylene glycol) heat-shock transformation method. To generate the trr1 and aco1 homozygous deletion mutants, cells were plated on YPD plates after heat shock and incubated under EHC for 24 h before replicated to YPD + Nourseothricin (NAT; 150 μg mL−1; WERNER BioAgents) plate for selection. All strains generated in this study and additional details regarding strain and plasmid construction are available upon request.
Spot dilution assay and viability test
To assess fungal growth, overnight cultures were diluted to OD600 of 0.2, followed by a serial tenfold dilution. 4 μL of each dilution was spotted onto the indicated agar medium. Plates were imaged after 32–48 h incubation under AC or 72–96 h incubation under EHC depending on the type of medium. To assess viability of erg251 mutant cells after EHC exposure, overnight cultures in YPD were diluted and plated on twenty YPD agar plates at ~250 CFUs (colony formation units) per plate. Four plates were incubated at 37 °C under AC and total CFUs were counted as the starting reference (set to 100%). The remaining sixteen plates were incubated under EHC. A set of four were retrieved at each indicated time point and incubated under AC. CFUs were determined after 30 h and normalized to the starting reference. WT cells survival after AC-to-EHC and EHC-to-AC-switch was similarly assessed. WT cells from an AC (or EHC) culture were plated on YPD agar. CFU counts after 3-day incubation under EHC (or after 2-day incubation under AC) were compared with the CFU of the starting reference.
Measurement of ATP level and ethanol production
Overnight cultures of SC5314 and the adh1 mutant in YPGalactose were washed with SD medium (2% glucose; ammonium sulfate as nitrogen source) and brought into the air-tight chamber, where they were diluted 1:10 in deoxygenated SD and incubated on a rotator in 1.2 mL aliquots. At each timepoint, one aliquot was taken out of the chamber for the following analyses: (1) ATP measurement: 100 μL culture was added to 1 mL extraction buffer (10% trichloroacetate (TCA), 4 mM EDTA) and incubated on a rotator for ~ 3 min. After centrifugation at 16,000 × g for 30 s, the supernatant was diluted 1:100 in TE (100 mM Tris-HC pH 7.8, 1 mM EDTA) and ATP was measured using the BacTiter-glo (Promega) kit in a 50 μL reaction. Luminescence was read in a 384-well flat white plate using a Tecan Spark (controlled by Spark Control) or F200 Pro (controlled by Magellan 7.2) plate reader. ATP solutions with known concentrations were used to generate a standard curve. (2) Ethanol production54: 750 μL of spent medium or SD medium containing known concentrations of ethanol was extracted by equal volume of Tri-n-butyl phosphate for 20 min at 30 °C with vigorous shaking. After separation by centrifugation, 400 μL of the top layer was mixed with equal volume of potassium dichromate solution (3% in 5 M sulfuric acid) and the reaction was left for 20 min at 30 °C with shaking. At this point, 250 μL of the bottom layer was transferred to a 96-well clear plate to determine absorbance at 595 nm. The limit of confident detection of ethanol is ~ 0.0125% using this method. (3) OD600 measurement. OD600 of 200 μL of a 1:10 diluted culture was measured in a 96-well plate using SpectraMax plate reader (Molecular Devices). Both ATP level and ethanol production were normalized to OD600.
Quantitative reverse transcription PCR (RT-qPCR)
ERG25 and ERG251 transcript levels with and without PF1163A treatment (20 μM for 75 min) were assessed in log-phase culture grown in YPD. In other experiments measuring gene expression, cells were collected from agar surface by scraping from the corresponding solid medium using a cell scraper. Media used for culturing and harvesting under EHC were deoxygenated for at least 12 h. Cell pellets were flash-frozen in liquid nitrogen and stored at −80 °C. RNA extraction and reverse transcription were performed as previously described55. 4 µL of 1:80 diluted cDNA was used to assess relative gene expression in a 10 µL SYBR green quantitative PCR reaction (SsoAdvanced Universal SYBR Green Supermix; Bio-Rad) with ACT1 level as the internal reference Data were collected using Bio-Rad CFX384 with CFX Manager. Sequences of the qPCR primers used in this study are provided in Supplementary Table 1.
Whole-cell lysate preparation and immunoblotting
Cells were harvested in the same method as for collecting samples for RT-qPCR. Whole cell lysates prepared using a TCA-precipitation-based method56 were resolved on 10% precast SDS-gels (Invitrogen) and transferred to polyvinylidene fluoride membrane. Blots were probed using anti-GFP (11814460001; Roche; 1:3000), anti-Hsp9057 (1:7000), or anti-TAP (CAB1001; Invitrogen; 1:3000) primary antibody and the proper HRP-conjugated secondary antibody (goat anti-mouse HRP (1706516; Bio-Rad; 1:7500); goat anti-rabbit HRP (611-1302; Rockland; 1:7500)). Blots were developed using ClarityTM ECL substrate (Bio-Rad) and after chemiluminescence imaging, stained with Coomassie R-250 (3% in 40% methanol/10% acetic acid) to confirm proper loading.
Disk diffusion assay
Disk diffusion assays were performed on Mueller–Hinton agar (Sigma-Aldrich; 38 g L−1; 25 mL in 10 cm regular petri dish) supplemented with 2% glucose at 35 °C by following the Clinical and Laboratory Standard Institute guidelines58. C. albicans cells were applied to agar surface by streaking twice from a ~ 2 × 106 cells mL−1 inoculum using a cotton swab. Compounds were spotted on 6 mm precut filter paper (Cytiva) using the following stocks: 0.125, 0.25, 0.5, 1 mg mL−1 PF1163A (in DMSO; Cayman); 0.625, 3.1, 12.5, 50, 200 mM 1181-0519 (in DMSO; synthesized for this study); 0.0625, 0.25, 1 M aqueous CoCl2 (Sigma-Aldrich); 25, 50, 100 mM aqueous bathophenanthrolinedisulfonic acid (BPS; Sigma-Aldrich). Plates for testing sensitivity to BPS were supplemented with 1 mM ascorbic acid (Bioshop) by spreading 250 μL of 100 mM fresh aqueous stock using glass beads. The sizes of inhibition zones were measured in Antibiogramj59 to calculate the annular radius.
Sterol profiling by GC-MS
Sample collection
Overnight cultures in SD broth were appropriately diluted and spread to SD agar (with ammonium sulphate as nitrogen source) made in 15 cm petri dishes. DOX (20 µg mL−1) was added when specified. Plates were incubated at 37 °C under AC or EHC for the indicated duration and cells harvested from the agar surface by scraping in SD with or without 20 µg mL−1 DOX broth using a cell scraper. After centrifugation, wet weight of each sample was determined in a 1.5 mL tube with known weight. Centrifugation and weighing were done in a fast pace to minimize reoxygenation of the EHC samples. Pellets (usually in a range of 100–300 mg) were flash-frozen in liquid nitrogen and stored at −80 °C.
Extraction
Unsaponifiable lipids were extracted as reported previously45. Cell pellets were resuspended in 3 ml extraction solution (25% KOH w/v, 64% methanol v/v) and transferred to borosilicate glass tubes. 30 µl of 2 mg mL−1 cholesterol (Bioshop) chloroform solution was added as an internal reference. Samples were incubated in an 80 °C water bath for one hour and vortexed every 20 min. After the samples were cooled to room temperature, 1 ml water and 3 ml chloroform were added. Each sample was extracted by vortexing (3 times 10 s) and then centrifuged at 1500 × g for 5 min. The bottom chloroform phases were transferred to glass vials by Pasteur pipettes. Chloroform was removed in Genevac (EZ-2 Series SP Scientific; extreme low boiling point mode) and samples were stored at −20 °C until analysis.
Derivatization
Dried lipid extracts were re-dissolved in chloroform with sonication, then split into two equal aliquots. Each aliquot was evaporated to dryness under a stream of nitrogen (1 mL/min). For derivatization, 20 µL pyridine and 30 µL N-methyl-N-(trimethylsilyl)trifluoroacetamide (Thermo Scientific, TS-48910) were added to each aliquot, followed by incubation at 37 °C for 30 min. After derivatization, 20 µl from each sample was transferred to a GC auto-sampler vial. 1 µL of sample was injected into the gas chromatography-mass spectrometry (GC-MS) for analysis. Each sample was analyzed in duplicate.
GC-MS and quantification
Derivatized sterol samples were analyzed using an Agilent GC/MS system (HP7890/HP5975) equipped with a DB-5MS column (30 m × 0.25 mm ID, 0.25 μm film thickness; Agilent 122-5532) and a 5 m S21 intermediate polarity fused silica guard column (0.53 mm ID). The injection was performed at 200 °C into a heated interface, with helium as the carrier gas at a constant flow rate of 1 mL/min. The oven temperature was initially held at 50 °C for 2.5 min, then ramped to 70 °C at 7.5 °C/min, followed by a further increase to 310 °C at 5 °C/min, where it was held for 1 min. The MS detector was activated at 6 min, and mass spectra were recorded in SIM mode at 0.9 scans/s across a range of 50–700 m/z. Biological triplicates or quadruplicates were analyzed in technical duplicates. Sterol species were identified by comparison with authentic standards or, when unavailable, by spectral matching against the NIST database. Peak areas were normalized to the internal standard (cholesterol) and the sample dry weight. Data analysis was performed using Agilent MassHunter Quantitative Analysis software.
Biofilm assay
The C. albicans biofilm formation assay was performed as previously described30, with slight modifications. Briefly, strains were grown overnight in YPD at 30 °C, then washed with phosphate-buffered saline (PBS) three times and resuspended in RPMI medium (10.4 g L−1 RPMI powder with L-glutamine (Gibco), 165 mM MOPS, 10% HI-FBS, pH 7.4) at 106 cells mL−1. 100 µL of cell suspension was added to each well of a 96-well plate containing 100 µL of pre-warmed RPMI medium. Cells were incubated at 37 °C with mild shaking (60 rpm) to allow adherence. After 90 min, each well was washed twice with PBS to remove non-adherent cells. 100 µL or 200 µL of fresh medium was added, and cells were incubated at 37 °C with mild shaking. After 24 h, non-adherent cells were washed away with PBS and biofilm metabolic activity was measured using the XTT reduction assay. 90 μL of XTT at 1 mg mL−1 and 10 μL of phenazine methosulfate at 0.32 mg mL−1 were added to each well, followed by incubation at 37 °C with mild shaking for 15 min. Supernatant was transferred to a fresh plate and absorbance was measured at 490 nm using a spectrophotometer. To test the efficacy of PF1163A in preventing biofilm formation, the compound (or same volume of DMSO vehicle control) was added at each indicated concentration to the growth medium after the adhesion stage.
Mice experiments
All procedures involving animals were approved by the Institutional Animal Care and Use Committee at the University of California San Francisco and were carried out according to the National Institute of Health guidelines for the ethical treatment of animals. Experiments were performed with 8- to 10-week-old female BALB/c mice from Charles River Laboratories, Hollister facility, room H48 (Strain code 028). Animals were provided with fresh water and chow ad libitum. Housing was performed under a 12:12 light: dark cycle in a well-ventilated room (10–15 air changes per hour) with stabilized temperature (68–79 degrees Fahrenheit) and humidity (30–70%).
Systemic candidiasis model in mice
Mice (n = 8) were infected with 1 × 105 CFU with wild-type (SC5314), the erg251-deletion mutant (CaLC9269), ERG251-A (CaLC9286), or ERG251-B complemented strains (CaLC9289) via the retrobulbar sinus. Infected animals were assessed at least once daily for signs of significant morbidity (decreased motor activity, respiratory distress, or a body condition score of 2 or less). To minimize suffering and guarantee humane euthanasia, animals that displayed any of these criteria were immediately euthanized by two independent procedures (administration of CO2, followed by cervical dislocation or bilateral thoracotomy).
Determination of fungal burden in organs
Mice (n = 10) were infected with 1 × 105 CFU with wild-type (SC5314), erg251-deletion mutant (CaLC9269), ERG251-A (CaLC9286), or ERG251-B complemented strains (CaLC9289) via the retrobulbar sinus. The kidney and liver were collected on day 3 post-infection. Organs were weighted and homogenized in 1 mL of saline solution (0.9%). Serial dilutions were performed and plated on Sabouraud agar. CFUs were counted after two days of growth at 30 °C and were normalized by the organ’s weight. To assess the impacts of BinGo mutants on virulence, relative fitness in the kidney was determined by calculating the competitive index as described previously60. Mice (n = 5) were infected with 1 × 105 CFU with a mixture of barcoded strains expressing wild-type ERG251 (CaLC10578) and erg251K93E (CaLC10581) or wild-type and erg251Y252A (CaLC10584) in a 1:1 competition via the retrobulbar sinus. Kidneys were recovered on day 3 post-infection and plated on Sabouraud agar. Colonies were washed from plates with sterile water and collected by centrifugation. Cell pellets were stored at −80 °C prior to isolation of genomic DNA. Genomic DNA from plated inocula and samples recovered from kidneys were isolated as previously described60. Strain abundance was determined by qPCR (SYBR Green; Bio-Rad), using strain-specific primers (Supplementary Table 1) and a Roche LightCycler 480 instrument. The competitive Index (CI) was determined as previously described60 using abundance fold change in the recovered (R) sample relative to the inoculum (I). To do this, concentration (Con.) of the WT and the mutant strain were determined by referring to a standard curve generated by the respective gDNA. The logarithmic form of R/I ratio for a given strain (x; WT or mutant) was calculated using the following formula: log2[Con.R_x/(Con.R_WT+Con.R_mut)]-log2[Con.I_x/(Con.I_WT+Con.I_mut)].
Commensalism model in mice
The mouse model of C. albicans commensalism was performed as previously described60. Female BALB/c mice (8–10-week-olds) were singly housed and treated with penicillin 1500 un mL−1 and streptomycin 2 mg mL−1 in drinking water starting 7 days before gavage and continuously for the remainder of the experiment. Feces were plated on Sabouraud agar to monitor for fungal growth. Mice were infected with 108 CFUs of a 1:1 mixture of the competition pair in 0.9% saline by gavage. C. albicans was recovered from the inoculum and host feces every 5 days for 3 weeks, and strain quantification was performed by qPCR. Competitive fitness was presented by log2 (R/I) values.
Histological analyses
Kidneys from mice infected with the indicated C. albicans strains were collected on Day 3 post-infection. Formalin-fixed kidneys were bisected in the sagittal orientation and paraffin-embedded using standard methods. Serial sections (5 µm) at multiple levels were stained by routine H&E and PAS methods. For immunohistochemistry, α-Candida (1:1000, Abcam ab53891) and α-CD45 (1:1200, BioLegend 103102) antibodies were used followed by detection with diaminobenzidine (ImmPACT DAB Substrate Kit, Peroxidase-Biolynx, Vector Laboratories, SK-4105). Slides were imaged at 40× magnification bright field (3DH Pannoramic Slide Scanner; the Imaging Facility at The Hospital for Sick Children, Toronto) and processed in SlideViewer.
Random mutagenesis of ERG251
To generate a mutant library, ERG251 was amplified ~20 fold using the GeneMorph II Random Mutagenesis Kit (Agilent). The library was fused to a nourseothricin resistance marker in tandem with a downstream homologous region. The final product was transformed into the “Parent” (erg251 tetO-ERG25) with DNA fragments expressing Cas9 and sgRNA targeting the erg251-deletion loci. Transformants were grown at 30 °C for two days on YPD + DOX (20 µg mL−1) + NAT (150 µg mL−1) and replica-plated onto two YPD + DOX + NAT plates to search for BinGo transformants.
Co-culture assays
HEK293T cells (ATCC; CRL-3216) expressing firefly luciferase (HEK293T-fLuc) were grown in RPMI medium (Gibco) supplemented with 10% heat-inactivated fetal bovine serum (HI-FBS; Gibco 16000044). Experiments were performed in cells within 10 passages post-recovery from stocks stored in liquid nitrogen. HEK293T-fLuc cells were seeded at 1 × 105 cells mL−1 (20 µL) in black-walled, clear-bottom 384-well plates (Corning 3764). Following overnight incubation at 37 °C with 5% CO2, continuous infusion, overnight C. albicans (SC5314) cultures were washed with PBS three times and added to wells seeded with 20 µL HEK293T-fLuc cells at a concentration of 2.5 × 104 cells mL−1. Twofold dilutions of test compound were added. 10 µM caspofungin was also added to wells as a control that kills fungal cells. Control plates containing monocultures of HEK293T-fLuc cells were prepared in the similar way. All assay plates were incubated for 24 h at 37 °C with 5% CO2. After fungal growth was determined by measuring optical density at 600 nm (OD600; with DMSO-treated wells as reference for maximal fungal growth), mammalian cell viability was quantified using Steady-Glo Luciferase Assay. 10 µL Steady-Glo Luciferase Assay reagent was added to each assay well, followed by incubation at room temperature for 10 min. Luminescence was measured after incubation at room temperature using a TECAN Spark microplate reader. Data were normalized to the signal obtained from the 10 µM caspofungin-treated control (reference for maximal HEK293T-fLuc growth). Data present mean and SD of three independent experiments and each of experiment was performed in technical triplicate.
Macrophage killing assay
Mouse monocyte-macrophage lineage J774A.1 cells (J7; ATCC; TIB-67) were maintained in RPMI + 10% HI-FBS. Experiments were performed in cells within 10 passages post-recovery from stocks stored in liquid nitrogen. To quantify macrophage death in co-culture with C. albicans, J7 cells were seeded at 4 × 105 cells mL−1 (100 µL) in clear-bottom 96-well plates. Cell culture was grown in RPMI medium supplemented with 3% HI-FBS for 18 h at 37 °C with 5% CO2. On the following day, overnight culture of wild-type C. albicans (SC5314) was washed three times with PBS and diluted to 8 × 105 cells mL−1 in RPMI medium supplemented with 3% HI-FBS and 2 µg mL−1 of propidium iodide (PI, Sigma, P417). 100 µL of fungal cell suspension was added to wells seeded with J7 cells. Co-cultures were treated with PF1163A in a twofold serial dilution with equal volume of DMSO as vehicle controls. Co-cultures were incubated for 16 h at 37 °C with 5% CO2 and imaged using the IncuCyte S3 Live-Cell Analysis System. Red area was quantified using the IncuCyte Base Analysis Software. Data were normalized to DMSO controls. Data present the mean and SD of three independent experiments and each of experiment was performed in technical triplicate.
Metabolic stability assays (Analiza Inc.)
Mouse liver microsomal stability assays were performed by Analiza Inc. following the same procedures as in Puumala et al. 61.
Construction of phylogenetic tree for ERG251 and ERG25 orthologs
The protein sequences of the fungal sterol C4-methyl oxidases were downloaded from Candida Genome Database or Yeast Genome Database (Supplementary Data 4). The tree was generated with Maximum Likelihood method with 1000 bootstrap replicates and LG + G amino acid substitution model in Mega 1162.
Synthesis of 1181-0519
-
(1).
N-(2-oxopropyl)acetamide: Synthesis of the known compound followed the previously reported literature63. In brief, the reactants glycine (7.500 g, 0.1000 mol), pyridine (47.4 g, 48.5 mL, 0.599 mol), and acetic anhydride (118.4 g, 109.6 mL, 1.159 mol) were added to a 500 mL round bottom flask and heated under reflux with stirring for 6 h. The excess pyridine and acetic anhydride were removed by distillation. Water (200 mL) was added to the residue and boiled for 30 min followed by distillation to remove the remaining water and acetic acid. Purification was achieved with a gradient of 0–2% methanol in dichloromethane to give the product as a pale-yellow oil consistent with literature characterization64. Yield: 3.57 g, (31%). Rf: 0.24 (2% methanol in dichloromethane) 1H NMR (600 MHz, DMSO-d6) δ 8.11 (s, NH), 3.87 (d, J = 5.6 Hz, 2H), 2.06 (s, 3H), 1.86 (s, 3H) (Supplementary Data 5). NMR was performed using Bruker Avance NEO 600 (14.1 Tesla Ultrashield Plus; iBBO Z-gradient) and analyzed by Topspin 4.5.
-
(2).
N-(2-(2-(4-nitrophenyl)hydrazineylidene)propyl)acetamide (1181-0519): 4-nitrophenylhydrazine hydrochloride (0.176 g, 0.928 mmol) and N-(2-oxopropyl)acetamide (0.100 g, 0.867 mmol) were dissolved into DMSO (5 mL) and stirred at room temperature for 4 h. The reaction was quenched by water (20 mL) and the precipitate was vacuum filtered to yield the product as an orange powder. Purity determined using HPLC (LC: Agilent 1290 Infinity II; UV: 1290 VWD (G71148); column: InfinityLab Poroshell 120 EC-C18, 1.9 µm, 2.1 × 50 mm):>98% pure (Rt = 2.28 min). Yield: 0.070 g, (32%). 1HNMR (DMSO-d6, 600 MHz) δ 9.89 (s, NH), 8.12 (t, J = 5.3 Hz, NH), 8.09 (apparent doublet, J = 9.3 Hz, 2H), 7.20 (apparent doublet, J = 9.0 Hz, 2H), 3.88 (d, J = 5.8 Hz, 2H), 1.88 (s, 3H), 1.87 (s, 3H). 13C NMR (DMSO-d6, 151 MHz) δ 169.85, 152.06, 149.11, 138.57, 126.35, 111.94, 45.51, 23.02, 14.60. 15N NMR (60.8 MHz, DMSO-d6) δ 370.27, 309.58, 143.89, 112.99. HRMS (Thermo Fisher Scientific DFS (Double Focusing) Mass Spectrometer; analyzed by Thermo Xcalibur 2.2) for C11H14O3N4 [M + H] + calcd. 251.1099, found 251.1094 (Supplementary Data 5).
C. elegans infection model
C. elegans strain AU37 (glp-4(bn2); sek-1(km4)) was obtained from the Caenorhabditis Genetics Center (University of Minnesota) and was maintained and propagated at 15 °C on nematode growth medium (NGM) seeded with Escherichia coli (OP50), as previously described65. C. elegans infection assays were performed following a liquid-based protocol adapted from previous studies66,67,68. In brief, a 5 mL culture of C. albicans strain (SC5314) was incubated overnight in YPD at 30 °C with shaking (220 rpm). 50 μL of the saturated overnight culture was deposited and spread over the surface of 10 cm brain heart infusion (BHI) agar plates supplemented with 45 μg mL−1 kanamycin, 100 μg mL−1 streptomycin, and 100 μg mL−1 ampicillin. Dried agar plates were incubated overnight at 30 °C prior to use in infection assays. Synchronized C. elegans L1s obtained from an embryo preparation69 were plated onto 10 cm NGM agar plates (~ 1500 worms/plate) seeded with OP50 and incubated for 72 h at 25 °C to allow worm development to adulthood. Adult worms were washed off plates into 50 mL conical tubes with ~10 mL of M9 buffer and filtered through 30 µm cell strainers (pluriStrainer®) to separate adults from any unsynchronized larvae. Retained adult worms were subsequently washed off filter into 15 mL conical tubes with ~ 10 mL of M9 buffer. Conical tubes were centrifuged at 9 × g for 1 min to gently pellet worms, buffer was removed via aspiration, and worms were washed twice with ~10 mL of fresh M9 buffer. Worm suspensions were counted using a stereomicroscope and ~1500 worms were pipetted onto either previously prepared BHI agar plates seeded with C. albicans (SC5314) or NGM plates seeded with E. coli (OP50; uninfected control). Plates were incubated for 3 h at 25 °C to allow C. elegans to forage and consume C. albicans (or E. coli) from agar surface. Following the infection period, worms were washed off agar plates into 15 mL conical tubes with ~10 mL of M9 buffer. Conical tubes were centrifuged at 9 × g for 1 min to gently pellet worms, buffer/microbial suspension was removed via aspiration, and worms were washed an additional 5 times with ~10 mL of fresh M9 buffer to remove as much unconsumed C. albicans as possible. Washed worms (~ 1500) were then transferred onto 10 cm unseeded NGM agar plates (supplemented with 45 μg mL−1 kanamycin, 100 μg mL−1 streptomycin, and 100 μg mL−1 ampicillin) and allowed to crawl around for 1 h to remove remaining traces of C. albicans (SC5314) or E. coli (OP50) from their cuticle prior to use. Chemical treatment was setup in 48-well flat-bottom culture plates, where 100 μL of a 20% BHI-M9 solution (supplemented with 45 μg mL−1 kanamycin, 100 μg mL−1 streptomycin, and 100 μg mL−1 ampicillin) containing either dissolved PF1163A or DMSO solvent (1% v/v) was distributed to desired wells. ~50 adult worms from either C. albicans or E. coli (uninfected control)-exposed populations were then sterilely transferred to corresponding wells for each chemical treatment using a platinum wire pick. Following the addition of worms, assay plates were sealed with parafilm and placed in a plastic container lined with wet paper towel to prevent evaporation. Containers were incubated for 4 days at 25 °C with shaking (160 rpm). Worm survival was scored manually for each treatment using a stereomicroscope at 0, 16, 24, 48, 72, and 96-h timepoints. Kaplan–Meier survival analysis was performed in GraphPad Prism 10. Infection experiments were performed in biological triplicates.
Statistics and reproducibility
At least three biological repeats were performed for quantitative assays. Qualitative experiments, such as spotting growth assays, were repeated at least twice or using additional transformants. Highly similar results were obtained and representative images shown. t-tests (paired or unpaired), Mann–Whitney tests, and Mantel–Cox tests were performed in GraphPad Prism 10 to examine the significance of difference. No statistical method was used to predetermine sample size. No data were excluded from the analyses. The experiments were not randomized. Except scoring the GRACE screen results without genetic annotation, the investigators were not blinded to allocation during experiments and outcome assessment.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Raw data of this study are available in Source Data or as Supplementary Data. Source data are provided with this paper.
References
Denning, D. W. Global incidence and mortality of severe fungal disease. Lancet Infect. Dis. https://doi.org/10.1016/S1473-3099(23)00692-8 (2024).
Brown, G. D., Denning, D. W. & Levitz, S. M. Tackling human fungal infections. Science 336, 647–647 (2012).
Sprague, J. L., Kasper, L. & Hube, B. From intestinal colonization to systemic infections: Candida albicans translocationand dissemination. Gut Microbes 14, 2154548 (2022).
Lee, Y., Robbins, N. & Cowen, L. E. Molecular mechanisms governing antifungal drug resistance. npj Antimicrob. Resist. 1, 5 (2023).
Sekeresova Kralova, J. et al. Competitive fungal commensalism mitigates candidiasis pathology. J. Exp. Med. 221, e20231686 (2024).
Zheng, L., Kelly, C. J. & Colgan, S. P. Physiologic hypoxia and oxygen homeostasis in the healthy intestine. A review in the theme: cellular responses to hypoxia. Am. J. Physiol. Cell Physiol. 309, C350 (2015).
Burgain, A., Tebbji, F., Khemiri, I. & Sellam, A. Metabolic reprogramming in the opportunistic yeast Candida albicans in response to hypoxia. mSphere 5, e00913-19 (2020).
Pradhan, A. et al. Hypoxia promotes immune evasion by triggering β-glucan masking on the Candida albicans cell surface via mitochondrial and cAMP-protein kinase A signaling. mBio 9, e01318-18 (2018).
Mulhern, S. M., Logue, M. E. & Butler, G. Candida albicans transcription factor Ace2 regulates metabolism and is required for filamentation in hypoxic conditions. Eukaryot. Cell 5, 2001–2013 (2006).
Burgain, A. et al. A novel genetic circuitry governing hypoxic metabolic flexibility, commensalism and virulence in the fungal pathogen Candida albicans. PLoS Pathog. 15, e1007823 (2019).
Dumitru, R., Hornby, J. M. & Nickerson, K. W. Defined anaerobic growth medium for studying Candida albicans basic biology and resistance to eight antifungal drugs. Antimicrob. Agents Chemother. 48, 2350 (2004).
Desai, P. R., van Wijlick, L., Kurtz, D., Juchimiuk, M. & Ernst, J. F. Hypoxia and temperature regulated morphogenesis in Candida albicans. PLoS Genet. 11, e1005447 (2015).
Bonhomme, J. et al. Contribution of the glycolytic flux and hypoxia adaptation to efficient biofilm formation by Candida albicans. Mol. Microbiol. 80, 995–1013 (2011).
Henry, M., Burgain, A., Tebbji, F. & Sellam, A. Transcriptional control of hypoxic hyphal growth in the fungal pathogen Candida albicans. Front. Cell Infect. Microbiol. 11, 770478 (2022).
Synnott, J. M., Guida, A., Mulhern-Haughey, S., Higgins, D. G. & Butler, G. Regulation of the hypoxic response in Candida albicans. Eukaryot. Cell 9, 1734 (2010).
Lash, E. et al. Ent2 governs morphogenesis and virulence in part through regulation of the Cdc42 signaling cascade in the fungal pathogen Candida albicans. mBio 14, e0343422 (2023).
Xiong, E. H. et al. Functional genomic analysis of genes important for Candida albicans fitness in diverse environmental conditions. Cell Rep. 43, 114601 (2024).
Fu, C. et al. Leveraging machine learning essentiality predictions and chemogenomic interactions to identify antifungal targets. Nat. Commun. 12, 6497 (2021).
Roemer, T. et al. Large-scale essential gene identification in Candida albicans and applications to antifungal drug discovery. Mol. Microbiol 50, 167–181 (2003).
Omeara, T. R. et al. Global analysis of fungal morphology exposes mechanisms of host cell escape. Nat. Commun. 6, 1–10 (2015).
Setiadi, E. R., Doedt, T., Cottier, F., Noffz, C. & Ernst, J. F. Transcriptional response of Candida albicans to hypoxia: linkage of oxygen sensing and Efg1p-regulatory networks. J. Mol. Biol. 361, 399–411 (2006).
Song, Y. et al. ADH1 promotes Candida albicans pathogenicity by stimulating oxidative phosphorylation. Int. J. Med. Microbiol. 309, 151330 (2019).
Mukherjee, P. K. et al. Alcohol dehydrogenase restricts the ability of the pathogen Candida albicans to form a biofilm on catheter surfaces through an ethanol-based mechanism. Infect. Immun. 74, 3804 (2006).
Da, A. et al. Thioredoxin regulates multiple hydrogen peroxide-induced signaling pathways in Candida albicans. Mol. Cell Biol. 30, 4550 (2010).
Veri, A. O. et al. Tuning Hsf1 levels drives distinct fungal morphogenetic programs with depletion impairing Hsp90 function and overexpression expanding the target space. PLoS Genet. 14, e1007270 (2018).
Jordá, T. & Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes 11, 795 (2020).
Mo, C., Valachovic, M., Randall, S. K., Nickels, J. T. & Bard, M. Protein-protein interactions among C-4 demethylation enzymes involved in yeast sterol biosynthesis. Proc. Natl. Acad. Sci. USA 99, 9739–9744 (2002).
Darnet, S. & Rahier, A. Enzymological properties of sterol-C4-methyl-oxidase of yeast sterol biosynthesis. Biochim. Biophys Acta (BBA) - Mol. Cell Biol. Lipids 1633, 106–117 (2003).
Zhou, X. et al. Erg251 has complex and pleiotropic effects on sterol composition, azole susceptibility, filamentation, and stress response phenotypes. PLoS Pathog. 20, e1012389 (2024).
Xiong, L. et al. Biofilm-associated metabolism via ERG251 in Candida albicans. PLoS Pathog. 20, e1012225 (2024).
Li, L. & Kaplan, J. Characterization of yeast methyl sterol oxidase (ERG25) and identification of a human homologue. J. Biol. Chem. 271, 16927–16933 (1996).
Okamoto, M. et al. Erg25 controls host-cholesterol uptake mediated by Aus1p-associated sterol-rich membrane domains in Candida glabrata. Front. Cell Dev. Biol. 10, 820675 (2022).
Matha, A. R., Xie, X., Maier, R. J. & Lin, X. Nickel tolerance is channeled through C-4 methyl sterol oxidase Erg25 in the sterol biosynthesis pathway. PLoS Genet. 20, e1011413 (2024).
Bard, M. et al. Cloning and characterization of ERG25, the Saccharomyces cerevisiae gene encoding C-4 sterol methyl oxidase. Proc. Natl. Acad. Sci. USA 93, 186–190 (1996).
MacPherson, S. et al. Candida albicans zinc cluster protein Upc2p confers resistance to antifungal drugs and is an activator of ergosterol biosynthetic genes. Antimicrob. Agents Chemother. 49, 1745–1752 (2005).
Finkel, J. S. & Mitchell, A. P. Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 9, 109–118 (2010).
Witchley, J. N. et al. Candida albicans morphogenesis programs control the balance between gut commensalism and invasive infection. Cell Host Microbe 25, 432 (2019).
Abramson, J. et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630, 493–500 (2024).
Dobson, L. et al. TmAlphaFold database: membrane localization and evaluation of AlphaFold2 predicted alpha-helical transmembrane protein structures. Nucleic Acids Res. 51, D517–D522 (2023).
Zhu, G., Koszelak-Rosenblum, M., Connelly, S. M., Dumont, M. E. & Malkowski, M. G. The crystal structure of an integral membrane fatty acid α-hydroxylase. J. Biol. Chem. 290, 29820–29833 (2015).
Shen, J., Wu, G., Tsai, A. L. & Zhou, M. Structure and mechanism of a unique diiron center in mammalian stearoyl-CoA desaturase. J. Mol. Biol. 432, 5152–5161 (2020).
Shen, J., Wu, G., Pierce, B. S., Tsai, A. L. & Zhou, M. Free ferrous ions sustain activity of mammalian stearoyl-CoA desaturase-1. J. Biol. Chem. 299, 104897 (2023).
Nose, H. et al. PF1163A, a novel antifungal agent, inhibit ergosterol biosynthesis at C-4 sterol methyl oxidase. J. Antibiotics 55, 969–974 (2002).
Lee, A. Y. et al. Mapping the cellular response to small molecules using chemogenomic fitness signatures. Science 344, 208 (2014).
Kim, S. H. et al. Inhibiting C-4 methyl sterol oxidase with novel diazaborines to target fungal plant pathogens. ACS Chem. Biol. 17, 1343–1350 (2022).
Okabe, M., Sugita, T., Kinoshita, K. & Koyama, K. Macrolides from a marine-derived fungus, Penicillium meleagrinum var. viridiflavum, showing synergistic effects with fluconazole against azole-resistant Candida albicans. J. Nat. Prod. 79, 1208–1212 (2016).
Blosser, S. J., Merriman, B., Grahl, N., Chung, D. & Cramer, R. A. Two C4-sterol methyl oxidases (Erg25) catalyse ergosterol intermediate demethylation and impact environmental stress adaptation in Aspergillus fumigatus. Microbiology 160, 2492–2506 (2014).
Hughes, A. L., Lee, C. Y. S., Bien, C. M. & Espenshade, P. J. 4-methyl sterols regulate fission yeast SREBP-scap under low oxygen and cell stress. J. Biol. Chem. 282, 24388–24396 (2007).
Chang, Y. C., Bien, C. M., Lee, H., Espenshade, P. J. & Kwon-Chung, K. J. Sre1p, a regulator of oxygen sensing and sterol homeostasis, is required for virulence in Cryptococcus neoformans. Mol. Microbiol. 64, 614–629 (2007).
Gaylor, J. L. & Mason, H. S. Investigation of the component reactions of oxidative sterol demethylation EVIDENCE AGAINST PARTICIPATION OF CYTOCHROME P-450. J. Biol. Chem. 243, 4966–4972 (1968).
Rahier, A., Smith, M. & Taton, M. The role of cytochrome b5in 4α-methyl-oxidation and C5(6) desaturation of plant sterol precursors. Biochem. Biophys. Res. Commun. 236, 434–437 (1997).
Dashko, S., Zhou, N., Compagno, C. & Piškur, J. Why, when, and how did yeast evolve alcoholic fermentation?. FEMS Yeast Res. 14, 826–832 (2014).
Min, K., Ichikawa, Y., Woolford, C. A. & Mitchell, A. P. Candida albicans gene deletion with a transient CRISPR-Cas9 system. mSphere 1, e00130-16 (2016).
Seo, H. B. et al. Measurement of ethanol concentration using solvent extraction and dichromate oxidation and its application to bioethanol production process. J. Ind. Microbiol. Biotechnol. 36, 285–292 (2009).
Zhang, A., Liu, Z. & Myers, L. C. Differential regulation of white-opaque switching by individual subunits of Candida albicans mediator. Eukaryot. Cell 12, 1293–1304 (2013).
Liu, Z. & Myers, L. C. Mediator tail module is required for Tac1-activated CDR1 expression and azole resistance in Candida albicans. Antimicrob. Agents Chemother. 61, e01342-17 (2017).
Larsen, B. et al. Isolation and partial characterization of Hsp90 from Candida albicans. Ann. Clin. Lab. Sci. 33, 86–93 (2003).
James S. et al. CLSI M100 Performance Standards for Antimicrobial Susceptibility Testing (Clinical and Laboratory Standards Institute, 2025).
Alonso, C. A. et al. Antibiogramj: a tool for analysing images from disk diffusion tests. Comput Methods Prog. Biomed. 143, 159–169 (2017).
Noble, S. M., French, S., Kohn, L. A., Chen, V. & Johnson, A. D. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 42, 590–598 (2010).
Puumala, E. et al. Structure-guided optimization of small molecules targeting Yck2 as a strategy to combat Candida albicans. Nat. Commun. 16, 1–16 (2025).
Tamura, K., Stecher, G. & Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. https://doi.org/10.1093/molbev/msab120.
Wiley, R. H. & Borum, O. H. Conversion of α-amino acids to acylamido ketones and oxazoles. J. Am. Chem. Soc. 70, 2005–2006 (1948).
Yuan, C. & Chen, D. Studies on organophosphorus compounds 55. A new and facile synthetic route to 1-Alkyl(Aryl)-2-amino-1-hydroxyalkylphosphonic acids. Synth. (Stuttg.) 1992, 531–532 (1992).
Stiernagle, T. Maintenance of C. elegans. WormBook 1–11 https://doi.org/10.1895/WORMBOOK.1.101.1 (2006).
Kim, G. H., Rosiana, S., Kirienko, N. V. & Shapiro, R. S. A simple nematode infection model for studying Candida albicans pathogenesis. Curr. Protoc Microbiol. 59, e114 (2020).
Breger, J. et al. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 3, 0168–0178 (2007).
Iyer, K. R. et al. Identification of triazenyl indoles as inhibitors of fungal fatty acid biosynthesis with broad-spectrum activity. Cell Chem. Biol. 30, 795–810.e8 (2023).
Burns, A. R. et al. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat. Protoc. 1, 1906–1914 (2006).
Acknowledgements
We thank Merck and Genome Canada for making the original GRACE mutant collections available. We thank all past and current Cowen lab members and Dr. Peter Stogios for helpful discussions. We acknowledge Drs. Vivian Bradaschia and Karuna Kapoor at The Centre for Phenogenomics Pathology Core and Dr. Kimberly Lau at the Imaging Facility at The Hospital of Sick Children for their assistance with histological analyses. B.Y. is supported by a Canadian Institute of Health Research Canada Graduate Scholarship-Doctoral award (199307). L.E.C. is supported by a National Institutes of Health (NIH) R01 grant (1R01AI127375) and a Canadian Institutes of Health Research Foundation grant (FDN-154288), and is a Canada Research Chair (Tier 1) in Microbial Genomics & Infectious Disease and co-Director of the CIFAR Fungal Kingdom: Threats & Opportunities program. D.D. and D.P. are supported by Brock Start-up Funds. The authors thank the Brock-Niagara Validation, Prototyping and Manufacturing Institute (VPMI) at Brock University for their support and access to the LC-MS system.
Author information
Authors and Affiliations
Contributions
Z.L., B.Y., N.R., and L.E.C. conceptualized the study. L.E.C. and D.D. obtained funding and provided resources for the study. G.D.W. and S.M.N. provided resources for this study. Z.L., B.Y., P.B., X.C., D.P., T.D., S.H., C.F., and L.S.A. conducted investigations. Z.L., B.Y., P.B., X.C., C.F., and T.D. performed analysis and visualization. N.R., D.D., G.D.W., S.M.N., and L.E.C. supervised the study. Z.L., B.Y., N.R., and L.E.C. wrote the draft manuscript with P.B., X.C., D.P., T.D., and D.D.’s contribution to the method section. All authors reviewed and edited the final manuscript. P.B. and X.C. contributed equally to this work.
Corresponding author
Ethics declarations
Competing interests
L.E.C. is a co-founder and shareholder in Bright Angel Therapeutics, a platform company for the development of novel antifungal therapeutics. L.E.C. is a Science Advisor for Kapoose Creek, a company that harnesses the therapeutic potential of fungi. All other authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Liu, Z., Yiu, B., Basso, P. et al. Sterol-C4-methyl-oxidase Erg251 governs Candida albicans hypoxic growth, commensalism and virulence. Nat Commun 16, 11479 (2025). https://doi.org/10.1038/s41467-025-66482-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-66482-5
