
Abstract
Iron bioavailability is often limited especially in calcareous soils. Trichoderma harzianum strongly improves plant iron uptake and growth in calcareous soils. However, little is known about the mechanisms by which T. harzianum mobilizes iron in calcareous soils. Here, the model strain T. harzianum NJAU4742 and a synthetic microbial community (SynCom) was used to show that the efficacy of T. harzianum in enhancing plant iron nutrition in calcareous soils depends on the soil microbiome. Enhanced iron-mobilization functions of the SynCom were observed in the presence of T. harzianum NJAU4742. Concurrently, T. harzianum NJAU4742 improved the iron-mobilization capacity of the SynCom by enriching strains that are able to do so. Finally, Chryseobacterium populi was identified as a key driver of iron mobilization, while their synergistic colonization further enhances this process. This study unveils a pivotal mechanism by which T. harzianum NJAU4742-mediated re-structuring of the soil microbiome and ameliorates plant iron nutrition.
Similar content being viewed by others


Introduction
Iron plays a critical role in various cellular processes and is an essential mineral nutrient for plant growth1. However, despite its abundance in the earth’s crust, iron bioavailability is often limited in most soils. It forms highly insoluble oxide and hydroxide minerals, which are unavailable for plant growth2, in neutral and alkaline soil. The rhizosphere microbiome has the potential to assist plants in the acquisition of iron from soils3,4,5,6. Several microbial isolates have been identified that significantly enhance plant iron nutrition7,8,9 and plants can enhance iron uptake also by recruiting siderophore-secreting bacteria to their roots. These findings underline the important roles of the rhizosphere microbiome in the acquisition of iron by plants.
In agriculture, beneficial microbes hold great potential as inoculants to sustainably enhance plant growth and crop yields10,11. However, the composition and functioning of microbiomes can be influenced by microbial inoculants, either directly through their secreted metabolites or indirectly via their impact on plant physiology and associated microbial interactions5,12,13,14. Trichoderma spp. are among the most-widely used beneficial microbes15,16. Although Trichoderma growth and spore production are reduced under alkaline conditions17,18,19 and Trichoderma spp. favor neutral pH conditions17,18, plant growth promotion effects of Trichoderma spp. are also found under alkaline conditions. For instance, inoculation with Trichoderma spp. increased the iron concentration of wheat, cucumber, and white lupin on an iron-deficient calcareous medium20,21,22. Also in saline-alkali soil, Trichoderma spp. were found to affect soil available nutrition, pH, and bacterial community composition while improving the growth of medicago and maize23,24. However, the mechanisms by which Trichoderma spp. promote plant growth under alkaline conditions are poorly understood.
As a result of niche overlap, Trichoderma spp. are frequently in competition with other soil fungi, especially under stress condition25,26,27. The interactions of Trichoderma spp. with bacteria, however, appear to be more synergistic and often enhance plant development. In soils with neutral pH, Trichoderma spp. are thought to promote plant growth by regulating the structure of the rhizosphere microbiome structure16,28,29,30. It is, therefore, not unlikely that also under alkaline condition, modulation of the rhizosphere microbiome is important for the plant growth-promoting effects of Trichoderma spp. The intricate and dynamic interactions in the soil microbiome, however, bring big challenges for the study of plant-microbiome relationships31,32. To overcome this, researchers now use synthetic microbial communities (SynCom) to mimic rhizosphere microbiome under controlled and reproducible conditions32,33,34. Here we investigated interactions between T. harzianum NJAU4742 and soil microbiome in improving plant iron nutrition in calcareous soil using the SynCom, which simulated the soil microbiome.
Results
Synergic effect of Trichoderma harzianum NJAU4742 and SynCom on promoting plant iron accumulation and iron availability in calcareous soil
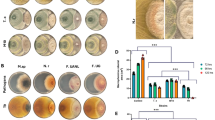
To investigate potential synergic effects of T. harzianum NJAU4742 and the rhizosphere microbiome on iron availability in the rhizosphere and plant iron accumulation, a pot experiment was conducted using natural soil and gamma-irradiated soil, the latter treated to eliminate the native microbiome (Fig. 1A-C). Pots with both soil treatments were either inoculated with T. harzianum NJAU4742 spores or left uninoculated, and subsequently planted with tomato seedlings. After 25 days of inoculation, iron accumulation in the plants (F3,20 = 19.62, P < 0.05) and iron availability in rhizosphere soil (F3,20 = 30.79, P < 0.05) was significantly increased in pots inoculated with T. harzianum NJAU4742, but this increase was not apparent in pots from which the native microbiome was eliminated (Fig. 1B and C). Moreover, iron accumulation was significantly higher in plants growing in soil with the native microbiome, than when plants had grown in sterilized soil (Fig. 1B). These data suggest illustrates T. harzianum NJAU4742 and rhizosphere microbiome synergistically improved iron availability in the rhizosphere and iron uptake by the plants.

The effects of the interaction between T. harzianum NJAU742 and the rhizosphere microbiome on plant growth (A), plant iron accumulation (B), and available iron of rhizosphere in calcareous soils (C). Simplified schematic of the experimental workflow (D, Image created with BioRender.com, with permission). Effect of Trichoderma and SynCom on tomato plant biomass (E), total iron accumulation in plants (F), and available iron content in the rhizosphere soil (G) under different inoculation treatments in the pot experiment. The changes in soil available iron content under different inoculation treatments in microcosm experiment (H). The inoculation treatments used in the study included T. harzianum NJAU4742 alone (T), SynCom alone (SynCom), and a combination of T. harzianum NJAU4742 and SynCom (T+SynCom). The bars in panels B–E indicate the mean ± standard deviation (s.d.) based on six independent biological replicates (shown as grey dots over the bars). Different lowercase letters above each bar indicate significant differences based on Analysis of Variance (ANOVA) followed by Tukey’s HSD test (P < 0.05).
In order to further investigate interactions between T. harzianum NJAU4742 and the rhizosphere microbiome, we created a SynCom representative of tomato rhizosphere (Fig. 1D and Supplementary Figure 1). We isolated bacteria from the soil and selected 98 strains that were previously detected to form a SynCom comprising 4 phyla, 7 classes, 13 orders, 23 families, and 40 genera. These strains comprise culturable fraction of the most abundant taxonomies in the tomato rhizosphere. Similar to the effects described above, neither T. harzianum NJAU4742 alone nor the SynCom alone had significant effect on plant dry weight and iron accumulation in the plant (Fig. 1E and F). Co-inoculation of T. harzianum NJAU4742 and the SynCom, however, significantly increased tomato plant dry weight (F3,20 = 10.78, P < 0.05, Fig. 1E), iron accumulation in plant (F3,20 = 8.894, P < 0.05, Fig. 1F), as well as the amount of available iron in rhizosphere soil (F3,20 = 47.57, P < 0.05, Fig. 1G).
Simultaneously, we incubated gnotobiotic calcareous soil with the same treatments as the pot experiment in closed microcosms, but without plants. Also there, co-inoculation of T. harzianum NJAU4742 and the SynCom significantly increased the soil iron availablity (F3,20 = 7.827, P < 0.05, Fig. 1H). Together, these findings indicate that synergistic interactions directly between T. harzianum NJAU4742 and the rhizosphere microbiome enhance iron availability in calcareous soils and iron uptake by plants.
Exudates of T. harzianum enhance SynCom iron mobilization
To investigate the effects of T. harzianum NJAU4742 on iron mobilization by the rhizosphere microbiome, we cultured T. harzianum NJAU4742 in iron-limited medium. Subsequently, we incubated the SynCom with insoluble Fe₂O₃ in either filter-sterilized Trichoderma growth medium (Tf+SynCom) or sterile water (SynCom) in microtiter plates. We found that the SynCom solubilized iron in the water (SynCom, Fig. 2A), but that the Trichoderma filtrate further increased the iron-mobilization ability of the SynCom (Tf+SynCom, F3,20 = 2117, P < 0.05, Fig. 2A). We then measured siderophore production at the end of this experiment (without Fe₂O₃), by filter sterilizing all treatments, mixing the cell-free filtrates with Chrome Azurol S (CAS) solution, and measuring OD at 630 nm. Strain T. harzianum NJAU4742 had produced siderophores in the iron-limited medium. The SynCom, however, produced more siderophores (SynCom, Fig. 2B), and SynCom siderophore production was further increased by the Trichoderma filtrate (Tf+SynCom, F3,20 = 8614, P < 0.05, Fig. 2B). Although Trichoderma filtrate (Tf) had pH of 7.2, the addition of Trichoderma filtrate to the SynCom (pH 6.85) reduced pH significantly (Tf+SynCom, F3,20 = 889.6, P < 0.05, Fig. 2C) to pH 6.41. These results suggest that the SynCom drives iron mobilization, but that this is enhanced exudates secreted by T. harzianum NJAU4742.

Trichoderma iron-limited filtrate effects on iron-mobilization ability in microtiter plate experiment (A). Effects of Trichoderma iron-limited filtrate on siderophore production (B) and pH (C) of SynCom. Principal component analysis (PCA) for 18 organic acids under different inoculation treatments in microtiter plate experiment (D). The solid line shows there are significant differences between SynCom and Tf+SynCom. Effect of Trichoderma iron-limited filtrate on oxalic acid (E) and α-ketoglutaric acid (F) of SynCom. The inoculation treatments used in the study included Tf, SynCom, and Tf+SynCom. Different lowercase letters above each bar represent significant differences based on analysis of variance (ANOVA) followed by Tukey’s HSD test (P < 0.05).
To investigate the changes in pH value in cell-free filtrates of the SynCom, we used Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) and measured the relative concentration of 18 organic acids. Principal component analysis (PCA) showed that 64.84% of the variation in organic acid levels was affected by the addition of Trichoderma filtrate, significantly (Adonis: R2 = 0.297, P < 0.05) affecting organic acid composition Fig. 2D). Two of the 18 organic acids, oxalic acid and α-ketoglutaric acid, significantly increased when the SynCom was incubated in presence of Trichoderma filtrates (F3,20 = 2696, F3,20 = 83.77, P < 0.05, Fig. 2E and F), whereas the sterile Trichoderma filtrate contained negligible amounts of these acids (Tf, Fig. 2E and F). Simultaneously, four organic acids significantly decreased (Supplementary Figure 2). Further analysis revealed that pH was negatively correlated to iron content of the cell-free filtrate and siderophore production (Supplementary Figure 3). The levels of α-ketoglutaric acid and oxalic acid showed a significant negative correlation with pH (Supplementary Figure 3), while iron content of the filtrates was significantly and positively correlated to siderophore production, content of α-ketoglutaric acid and oxalic acid (Supplementary Figure 3). These results underline that T. harzianum NJAU4742 enhances iron mobilization by SynCom and suggest that it stimulates the SynCom to produce siderophores, oxalic acid and α-ketoglutaric acid under iron-limited conditions.
T. harzianum NJAU4742 selected species of SynCom related to iron mobilization
We then assessed how T. harzianum NJAU4742 affected SynCom microbial community structure by sequencing 16S amplicons of SynCom DNA from the soil microcosm and microtiter plate experiments described above (Figs. 1 and 2). We found that neither T. harzianum NJAU4742 nor its filtrate affected richness of the SynCom (P > 0.05, Supplementary Figure 4A and C). Co-inoculation of T. harzianum NJAU4742 with the SynCom significantly changed SynCom composition in microcosms without plants (Supplementary Figure 4B). In the microtiter plate experiment, Trichoderma filtrate significantly affected beta diversity of the SynCom (Supplementary Figure 4D). Constrained analysis of principal coordinates (CAP) explained 72.68% and 81.41% of the variation in microcosm and microplate data, respectively, (Fig. 3A and C). Notably, iron content of the medium was positively associated with the community structure of SynCom in presence of Trichoderma filtrate in both microcosm and microtiter plate experiments (P < 0.05, Fig. 3A and C, Supplementary Figure 5). In contrast, the pH showed significantly negatively association with the community structure of SynCom in presence of Trichoderma filtrate in a microtiter plate (P < 0.05, Fig. 3C). These findings suggest that the community composition of SynCom related to iron mobilization is affected by T. harzianum NJAU4742.

Constrained analysis of principal coordinates (CAP) for the correlation between iron content and SynCom structure in microcosm (A) and microtiter plate experiment (C). Only significant factors are shown in the CAP figures, and the arrows indicate the lengths and angles between explanatory and response variables, reflecting their correlations. Difference analysis of SynCom under Trichoderma in microcosm treatment (B) and Trichoderma iron-limited filtrate in microtiter plate treatment (D). Correlation between Trichoderma iron-limited filtrate treatment effect on the relative change of single bacteria abundance and iron-mobilization characteristics of strains (E). Effects of Trichoderma iron-limited filtrate on the growth, pH, siderophore production, and iron-mobilization ability of 24 single strains (F). Venn network analysis for key isolates both enriched in microcosm and microtiter plate treatment and significantly affected by Trichoderma iron-limited filtrate (G). The inoculation treatments used in the study included SynCom alone (SynCom), a combination of T. harzianum NJAU4742 and SynCom (T+SynCom), and a combination of Trichoderma iron-limited filtrate and SynCom (Tf+SynCom).
We then identified SynCom members that were significantly enriched in response to T. harzianum NJAU4742. In the microcosm experiment, the relative abundance of 19 species significantly increased, while 18 species significantly decreased in response to T. harzianum NJAU4742 (P < 0.05, Fig. 3B and Supplementary Table 2). Among the enriched species, five—Burkholderia cepacia, Enterobacter cloacae, Pseudarthrobacter oxydans, Chryseobacterium rhizoplanae, and Flavobacterium tangerine—exhibited relative abundances exceeding 1% (Supplementary Figure 4E and Supplementary Table 2). Similarly, the addition of Trichoderma filtrate led to significant increases in the relative abundance of 24 species and decreases in 17 species (P < 0.05, Fig. 3D and Supplementary Table 3). Notably, five species—Klebsiella michiganensis, Sphingobacterium mizutaii, Bacillus megaterium, Ochrobactrum pseudogrignonense, and Klebsiella variicola—achieved relative abundances above 1% (Supplementary Figure 4F and Supplementary Table 3). The relative abundance of 24 enriched SymCom member was positively correlated to iron content of the medium in the microtiter plate experiment (R² = 0.41, P < 0.01, Fig. 3E). This suggests that these enriched SynCom species improve iron mobilization.
Subsequently, we tested the effect of Trichoderma filtrate on the 24 SynCom members that were significantly enriched in the microtiter plate experiment one-by-one. Of the 24 strains tested individually, the growth of 16 single strains was affected by Trichoderma filtrate, 14 strains significantly changed the pH of the medium in presence of Trichoderma filtrate, 13 strains exhibited enhanced siderophore production in presence of Trichoderma filtrate and 6 strains by themselves increased the amount of mobilized iron in presence of Trichoderma filtrate (P < 0.05, Fig. 3F). These results indicated that T. harzianum NJAU4742 directly affects the individual member of the SynCom.
In the soil microcosm experiment, 19 SynCom members increased in presence of T. harzianum NJAU4742, whereas 24 SynCom members with Trichoderma filtrate in the microtiter plates. Notably, five SynCom members—Delftia acidovorans, Chryseobacterium populi, Ochrobactrum pseudogrignonense, Paenibacillus cellulositrophicus, and Enterobacter cloacae—were consistently enriched across both experimental systems (Fig. 3G). Of these 5 strains, only C. populi showed a high response to Trichoderma filtrate that affected growth, lowering of pH, production of siderophores and mobilization of iron (Fig. 3G).
Synergic effects of C. populi and T. harzianum on iron mobilization
To further validate the synergistic effects between C. populi and T. harzianum NJAU4742, we conducted two complementary experiments (plate and pot assays) to examine the response of C. populi to T. harzianum NJAU4742. In the microtiter plate experiment, C. populi exposed to Trichoderma filtrate (Tf) exhibited significant increases compared to the control treatment in OD600 (F₁,₁₀ = 242.8, P < 0.05), iron content (F₁,₁₀ = 286.8, P < 0.05), siderophore production (F₁,₁₀ = 397.9, P < 0.05), α-ketoglutaric acid content (F₁,₁₀ = 104.4, P < 0.05), and oxalic acid content (F₁,₁₀ = 74.41, P < 0.05), alongside a decrease in pH (F₁,₁₀ = 1314, P < 0.05) (Fig. 4A).

Effects of Trichoderma iron-limited filtrate (Tf) on the growth and iron mobilization function of C. populi (A). The effects of C. populi inoculation and co-inoculation with Trichoderma (T) on tomato plant biomass, total plant iron accumulation, and rhizosphere soil available iron content (B). Different lowercase letters above each bar represent significant differences based on analysis of variance (ANOVA) followed by Tukey’s HSD test (P < 0.05).
Finally, we assessed iron mobilization and plant growth promotion in a pot experiment where tomato plants were inoculated with C. populi and T. harzianum NJAU4742 individually or together. Compared to the control and T. harzianum NJAU4742 (T) treatments, inoculation with C. populi significantly increased plant dry weight (F₃,₈ = 96.72, P < 0.05), iron accumulation (F₃,₈ = 733.8, P < 0.05), available iron (F₃,₈ = 389.2, P < 0.05), and decreased rhizosphere pH (F₃,₈ = 164.2, P < 0.05) (Fig. 4B). Furthermore, co-inoculation with T. harzianum NJAU4742 enhanced these effects, consistent with findings from the microtiter plate experiment.
To further investigate whether C. populi provides reciprocal benefits to T. harzianum NJAU4742, we cultured T. harzianum NJAU4742 using C. populi iron-limited filtrate (CHRYf). The results showed that CHRYf significantly promoted the growth of T. harzianum NJAU4742 (Supplementary Figure 6), suggesting that T. harzianum and C. populi synergistically promote the growth of each other.
Discussion
In this study, we reveal the relationship between changes in the rhizosphere microbiome structure mediated by T. harzianum NJAU 4742 and plant iron nutrition, highlighting a pivotal mechanism underlying the enhancement of rhizosphere microbiome capacity to improve plant iron uptake.
Trichoderma typically promotes plant growth and nutrition in soils, mostly in the soils with neutral or acidic pH35, but its effectiveness is limited under alkaline conditions due to its preference for an acidic environment17,18. In our study, T. harzianum NJAU 4742 did significantly increase plant iron nutrition in calcareous soil, but could not do so in soil from which the native microbiome was eliminated. This is line with a previous study showing that soil fungi are considered to be a key factor affecting bacterial community assembly processes26. It suggested that soil bacterial communities can be activated by Trichoderma36, and that this can affect the functional composition of bacterial communities or change their activity37. We used a SynCom to mimic the natural soil microbiome and found that the iron-mobilizing ability of both the natural microbiome and SynCom were activated by T. harzianum NJAU 4742. In this activation, microbial communication may play an important role in promoting the abundance of specific microbial taxa. It is known that many signaling molecules play a vital role in the direct communication between microorganisms38,39,40, suggesting that metabolites of Trichoderma might play a role in activating the microbiome. It will be interesting to find out whether Trichoderma can consistently affect distinct microbiomes in the same way, e.g. by using multiple SynComs composed of root-associated bacteria from different plant species. Moreover, SynCom can also involve multiple fungal species to investigate broader fungal-bacterial interactions in relation to plant iron accumulation.
Our findings suggest that T. harzianum NJAU4742 secretes metabolites that enhance the iron-mobilization function of SynCom, as many studies have reported that fungi can enhance certain bacterial functions, such as phosphorus solubilization41, disease control ability30, carbon source utilization42, and pollutant degradation43. Iron deficiency may serve as a crucial signal influencing the cooperative relationship between Trichoderma and the microbiome, in a way similar to how organic phosphorus affects the interaction between arbuscular mycorrhizal fungi and phosphate-solubilizing bacteria44. A previous study showed that fungal species modulated the availability of iron to bacterial species through the provision of fungal siderophores, such as ferrichrome and coprogen45. The fungal siderophores are absorbed by bacteria and promote the growth of bacteria in iron-limited condition, suggesting fungi can also play a direct role in iron mobilization. However, Trichoderma’s effect on iron mobilization appeared largely indirect and mediated though the microbiome. Overall, our results suggest that the ability of Trichoderma to enhance the iron-mobilization by the rhizosphere microbiome in iron-limited conditions are an essential part of Trichoderma’s ability to promote plant growth in calcareous soils.
Whereas microbiome community composition strongly influences microbiome function, soil fungi are recognized as drivers of bacterial community assembly processes26. Previous studies have shown that Trichoderma can affect the abundance, composition and functioning of soil microbiomes. Trichoderma was shown to directly or indirectly contribute to phosphorus solubilization and shape the rhizosphere microbial community, thereby enhancing crop yield46. However, Trichoderma can also inhibit plant pathogenic microorganisms and lead to an enrichment of microbial taxa that are noted for plant growth-promoting properties28. In our study, T. harzianum NJAU4742 significantly affects the alpha and beta diversity of SynCom, suggesting the Trichoderma-driven changes in microbiome composition improved iron mobilization by the microbiome. Our SynCom experiments identified isolates that were promoted by Trichoderma filtrate, whose ability on mobilizing iron were in line with the changes of their abundance in microtiter plates, suggesting that the compositional changes effected by Trichoderma enhanced the iron mobilization function of the microbiome. Whereas the Trichoderma-driven enrichment of bacteria with a high capacity to mobilize iron appears to be the primary mechanism enhancing SynCom functioning, the specific metabolites from T. harzianum NJAU 4742 regulating SynCom assembly remain unknown.
Identifying these core isolates are essential for comprehending the assembly processes and stability of microbial communities47. We found there are only five isolates both enriched in microcosm and microtiter plate experiments, which showed that there are isolates with functional redundancy in the SynCom related to culturing environment, provided a basis for maintaining the SynCom functions to adapt to different environments48. Despite similar functions exhibited by microbial communities, differences in community structure may arise due to microbial functional redundancy, suggesting that distinct structures might not directly impact community functions49. Based on the response of isolates to the Trichoderma iron-limited filtrate, only one isolate (C. populi) was ultimately selected from five isolates. Among them, D. acidovorans, O. pseudogrignonense, and E. cloacae only have part functions that significantly respond to Trichoderma regulation, while Paenibacillus cellulositrophicus has no functions that directly respond to Trichoderma regulation, which indicated the enrichment of these isolates may have other functions beyond simply participating in iron mobilization.
We confirmed that C. populi has the capacity to mobilize iron, which is consistent with our hypothesis. Previous studies found Chryseobacterium spp. significantly enhanced plant growth and iron nutrition in tomatoes50, sunflowers51, and rice52. In this light, we previously characterized of chryseochelin, an interesting siderophore produced by Chryseobacterium spp. In this study, we found the siderophore production of C. populi was significantly increased by T. harzianum NJAU 4742, which may be an important way for Chrysobacterium to mobilize insoluble iron. Moreover, we found T. harzianum and C. populi together significantly decreased pH of the rhizosphere, even though the T. harzianum NJAU4742 iron-limited filtrate was alkaline53, which means the pH reduction is associated with plant root. Simultaneously, we noticed a new cue that the cell-free filtrates of C. populi can promote the growth of Trichoderma, showing that the Trichoderma-enriched C. populi in turn also facilitate the growth of T. harzianum (Supplementary Figure 6). Previous studies have discovered that bacteria protect fungi in biotic and abiotic stress from stress damage54,55. Mycoavidus can protect Mortierella verticillata NRRL 6337 from nematode attack through highly cytotoxic macrolactones54 and Paraburkholderia edwinii can protect Aspergillus from phenazine-1-carboxylic acid by sequestering it55. Similarly, in our study, the protective function on T. harzianum exerted through C. populi. However, Chrysobacterium spp. have also been reported to have antifungal effect in several studies56. Overall, the specific mechanisms of interaction between T. harzianum NJAU 4742 and C. populi require further study.
In this study, we determined that the efficacy of T. harzianum NJAU 4742 in enhancing plant iron nutrition in calcareous soil depends on the soil microbiome. T. harzianum NJAU4742 enhanced the iron-mobilization function of a SynCom by boosting the SynCom’s capacity to secrete siderophores, secrete organic acids, and decrease pH under iron-limited conditions. Furthermore, we demonstrated that T. harzianum NJAU 4742 shapes microbiome composition by enriching specific functional taxa, including the isolate C. populi, and that T. harzianum NJAU 4742, with this isolate can synergistically promote plant growth. These insights collectively contribute novel perspectives and empirically grounded insights that hold the potential to address the pervasive challenge of iron deficiency in plants cultivated in calcareous soil.
Materials and methods
Synergic effect of Trichoderma harzianum NJAU4742 and microbiome on promoting plant iron accumulation and iron availability in calcareous soil
Cultivation of T. harzianum NJAU4742
The T. harzianum NJAU4742 strain is a model benifical fungi (Genbank accession GCA_002022785.1). Spore suspension of T. harzianum NJAU4742 stored in −80 °C freezer was plated on potato dextrose agar (PDA) plate for recovery. Strain T. harzianum NJAU4742 was cultured in the dark at 28 °C for 2 d. Spores were induced under light after fresh mycelium was transferred to a new PDA plate using a sterile puncher and cultured in the dark at 28 °C for 3 d. The spores were scraped off with 5 mL sterile water and filtered with four-layer sterile gauze. The spore density was later calculated by a hemocytometer, and adjusted to 107 spore mL−1 with sterile water53.
Construction of synthetic microbial community
In order to further quantify this synergic effect in a more standardized and repeatable way, we isolated bacteria from the soil and selected 98 strains to construct a SynCom. These strains comprise culturable fraction of the most abundant taxonomies in the tomato rhizosphere57. This SynCom comprised 4 phyla, 7 classes, 13 orders, 23 families, and 40 genera (Supplementary Figure 1).
In order to recover the bacterial culture, every single strain was incubated with tryptone soya agar57 (TSA, containing 15 g L−1 tryptone, 5 g L−1 soytone, 5 g L−1 sodium chloride, and 20 g L−1 agar, pH 7.0) at 30°C for 24 h. Single colony of each strain was transferred to 200 μL tryptone soya broth (TSB, liquid TSA) in a 96-well microtiter plate, with agitation of 170 r.p.m. at 30 °C for 16 h. The time allows the bacteria reach to stationary phase. We measured the optical density of each stain (OD600) with SpectraMax M5 microplate reader (Molecular Devices, USA), and adjusted the optical density to 1.0 with sterile water. Each single strain was mixed in equal proportions to construct the SynCom58,59.
Tomato seeds were surface sterilized in 75% ethyl alcohol for 1 min, 3% NaClO for 5 min, and rinsed in sterile distilled water five times, then placed on sterile filter paper for 5 d at 28°C to accelerate germination. The germinated seeds were sown on a sterile seedling medium (121 °C, 0.1 MPa, 2 h) and transferred to controlled glasshouse (ambient temperature variation ranging from 24 °C to 28 °C; 70% relative humidity; 16/8 h photoperiod with 200 μmol m−2 s−1 photosynthetic photon flux density, PPFD).
The calcareous soil was collected in Jingtai county, Lanzhou, Gansu (36°43′N, 103°33′E, Supplementary Table 1), which is tomato growing soil. This soil contained low concentration of iron due to relatively high pH. We used untreated soil (with natural soil microbiome) and sterilized soil (without natural soil microbiome) to investigate the synergic effect of soil microbiome and T. harzianum NJAU 4742 on plant iron accumulation. Untreated soil is natural calcareous soil with microbiome, while sterilized soil was prepared by subjecting natural calcareous soil to gamma radiation (50 kGy 60Co gamma-radiation). Air-dried soils were filtered through a 20-mesh sieve. Tomato seedlings with 3 leaves were transferred to the pots with 250 g either sterilized or untreated soil, and put in controlled glasshouse. We set up four treatments: 1) sterilized calcareous soil without T. harzianum NJAU4742, 2) sterilized calcareous soil with T. harzianum NJAU4742 (106 spore g−1 soil), 3) untreated calcareous soil without T. harzianum NJAU4742, 4) untreated calcareous soil with T. harzianum NJAU4742. T. harzianum NJAU4742 inoculation: 250 g sterilized or untreated calcareous soil inoculated with 10 mL spore suspensions (7.5 mL sterile water mixed with 2.5 mL 108 spores mL−1 suspensions) of T. harzianum NJAU4742. There were 6 replicates (6 tomato plants) for each treatment, and 24 pots in total. Tomato plants were sampled after 25 d to measure iron accumulation. The rhizosphere soil was also collected to measure available iron content.
The pot experiment of interaction between T. harzianum NJAU4742 and SynCom, the seedlings, and sterilized calcareous soil are the same as above. A total of 4 treatments were set up in this pot experiment (Fig. 1D): a. complete sterilized control (control); b. inoculation of T. harzianum NJAU4742 alone (T, 106 spore g−1 soil); c. inoculation of SynCom alone (SynCom, 107 CFU g−1 soil); d. co-inoculation of T. harzianum NJAU4742 and SynCom (T+SynCom, the final concentration was same as above). One seedling was used per pot and each treatment was replicated six times, for a total of 24 pots. Plants were watered with 10 mL of sterilized water regularly, and the greenhouse was kept clean to avoid introducing other soil microorganisms. Plants were harvested after 25 d to measure dry weight and iron accumulation. The rhizosphere soil was collected using sterile centrifuge tubes and naturally air-dried to measure available iron and pH.
In order to further determine interaction between T. harzianum NJAU4742 and the SynCom is not driven by plant, we used soil microcosms system (without plant) by using sterilized calcareous soil with the SynCom, the same materials as the pot experiment (Fig. 1D). In the microcosm, 100 g dried sterilized calcareous soil was placed in a 340 mL sterilized soil culture bottle, and weighed the bottles with soil recorded as original weight. The soil culture bottle was stabilized at room temperature for 2 w, and sterile distilled water was added to the stabilized sterilized soil to balance the water content as 40% of the maximum field water holding capacity every 2 d last 4 w60. All bottles were tested for bacterial and fungal contamination, and only completely sterile bottles were retained for the microcosm experiment. A total of four treatments were set up the same as pot the experiment in this experiment: a. Complete sterilized control (control); b. inoculation of T. harzianum NJAU4742 alone (T, 106 spore g−1 soil); c. inoculation of SynCom alone (SynCom, 107 CFU g−1 soil); d. co-inoculation of T. harzianum NJAU4742 and SynCom (T+SynCom, the final concentration was same as above). Each treatment had 6 replicates, a total of 24 bottles. The bottle was placed in a dark growth chamber with a constant temperature at 25 °C, and balanced water every 2 d until harvest. After 3 w, the soil per bottle was mixed well and collected, some of which were quickly frozen in liquid nitrogen and stored in an −80 °C refrigerator for amplicon sequencing, and the other part was naturally air-dried to measure available iron, manganese, copper, zinc, calcium and magnesium content and pH.
The leaf, stem, and root samples were fastly dried at 105 °C for 30 min and slowly dried at 75 °C until constant weight, and then ground into a powder using a tissue lyser (TL2010S, DHS Life Science & Technology, China). The iron of samples was extracted by mixed acid method (nitric acid:perchloric acid = 4:1) and measured by the Inductively Coupled Plasma-Optical Emission spectroscopy (ICP-OES, Agilent 710, USA). The iron accumulation was calculated as the concentration of iron content × biomass.
In order to measure iron, manganese, copper, and zinc content, we mixed 12.5 g air-dried soil with 25 mL DTPA extractant (0.005 M CaCl2, 0.1 M CaCl2, pH 7.3), shaked at 180 r.p.m. for 2 h, and determined the content by using Inductively Coupled Plasma-Optical Emission spectroscopy (ICP-OES, Agilent 710, USA)9. Calcium and magnesium content was measured as follows: 2.5 g air-dried soil was mixed with 25 mL 1 M neutral ammonium acetate solution (use ammonia water to adjust to pH 6.97), shaked for 30 min, determined by ICP-OES. Ten g soil was mixed with 25 mL of CO2-free deionized water (boiled for 5 min, sealed with plastic wrap, and cooled to room temperature), shaked well, and measured by FE22-Standard pH meter (Mettler Toledo, Switzerland).
Exudates of T. harzianum enhance SynCom iron mobilization
In order to obtain T. harzianum NJAU4742 iron-limited filtrate, we inoculated 2.5 mL T. harzianum NJAU4742 spores suspension (107 spores mL-1) into 250 mL iron-limited LNM medium (NH4Cl 0.25 g L−1, NaCl 0.125 g L−1, NaH2PO4·2H2O 0.5905 g L−1, Na2HPO4·12H2O 2.427 g L−1, KH2PO4 0.75 mg L−1, CaCl2·2H2O 0.147 mg L−1, MgSO4·7H2O 4.92 mg L−1, glucose 2 g L−1, casamino acids 3 g L−1, pH 6.8, Coolaber, Beijing, China), cultured at 28 °C at 170 r.p.m. for 5 d, then centrifuged at 10000 r.p.m. with 4 °C for 20 min, filtered (0.22 μm) and stored in −80 °C refrigerator.
The microtiter plate experiment: 1.35 mL 10% iron-limited LNM medium was placed into 24-well plate with 1.5 mg sterile Fe2O3 powder in each well as a basic medium for the microtiter plate experiment. A total of four treatments were set up in the microtiter plate experiment: a. added 0.15 mL sterile deionized water (control); b. added 0.15 mL T. harzianum NJAU4742 iron-limited filtrate (Tf); c. added 15 μL SynCom (OD600 = 1.0) and 0.15 mL sterile deionized water (SynCom); d. added 0.15 mL T. harzianum NJAU4742 iron-limited filtrate and 15 μL SynCom (Tf+SynCom). We washed all microbial suspension with 0.85% NaCl three times at 500 r.p.m. for 5 min, before culturing in LNM medium. Each treatment was replicated 6 times, and shaked plate at 170 r.p.m. and 30 °C for 12 h. Then pH, organic acid, and iron content of each well were measured, and amplicon sequencing was conducted. The method for pH, organic acid, iron content, and growth of single isolate was the same as above (added 15 μL isolate suspension (OD600 = 1.0) and 0.15 mL sterile deionized water or 0.15 mL T. harzianum NJAU4742 iron-limited filtrate).
Due to the direct impact of iron content on the siderophore production, in order to observe changes more intuitively in siderophore production of SynCom and isolates, we removed the Fe2O3 powder from the medium and cultured SynCom and isolates the same as above, then measured the siderophore production. Siderophore production was assayed using a modified version of the universal chemical assay developed by Schwyn and Neilands61. Bacterial samples were centrifuged at 5000 r.p.m. for 20 min, filtered (0.22 μm). Then, 100 µL cell-free filtrate or uninoculated medium control were added to 100 μl CAS assay solution in a 96-well microtiter plate, mixed with CAS in equal proportions, incubated at room temperature for 1 h. The OD630 of the cell-free filtrates (A) and uninoculated medium control (Ar) were then measured using a plate reader (SpectraMax M5) at room temperature. The formula for calculating the relative content of siderophores: SU = 1 - A / Ar.
The iron content of Syncom and isolates: bacterial samples were centrifuged at 15,000 r.p.m. at 4 °C for 10 min and filtered (0.22 uM) to retain filtrate. The filtrate was digested with mixed acid (nitric acid:perchloric acid = 4:1, 120 °C for 30 minutes, 150 °C for 30 minutes, and 190 °C for acid removal) and determined by ICP-OES.
Organic acids of SynCom and Chryseobacterium populi62: bacterial samples were centrifuged at 15000 r.p.m. at 4 °C for 10 min and filtered to retain filtrate. 0.10 mL filtrate samples were taken and placed in a 2 mL centrifuge tube. Added 100 μL 50% acetonitrile water, 100 μL 3-NPH (200 mM) and 100 μL EDC (120 mM; The solution containing 6% pyridine (2:1:1:1, v/v/v/v) was mixed for 1 min (total volume 400 μL), and the reaction was performed at 40 °C for 1 h, during which the shock was performed once in 5 min. After the reaction was completed, the reaction was centrifugated at 4 °C for 15 min at 12000 r.p.m. The filtrate was filtered through a 0.22 μM filter membrane, the filtrate was then applied to LC-MS/MS analysis and diluted 100 times with 50% acetonitrile water for LC-MS/MS analysis. LC-MS/MS analyses were performed using a Waters Acquity UPLC (Waters, USA) with an Acquity UPLC HSS T3 (1.8 µm, 2.1 mm × 100 mm) coupled to AB SCIEX 5500 Qtrap-MS (AB SCIEX, USA)63. The mobile phase consisted of water with 0.01% formate and Acetonitrile with 0.01% formate. The column and auto-sampler temperature were set at 30 °C and 4 °C respectively, and the injection volume was 1 μL. The raw data was integrated by MultiQuant, and the content of each compound was then calculated with the help of standard curve. 20 mg of standard product was dissolved in 1 mL 50% acetonitrile solution; Removed the standard and the prepared 3-NPH (200 mM) and EDC (120 mM) solution (containing 6% pyridine) (2:1:1, v/v/v) respectively; vortexed for 1 min and (total volume 1 mL); Reacted at 40 °C for 1 h, and shocked once in 5 min; Standard series solution was diluted with 50% acetonitrile. Drawed the standard curve according to the standard solutions with diverse concentrations and their peak areas used in the calculation of the concentration of organic acids in samples.
The growth of single isolates: 2 μL single isolate suspension was taken into 96-well plate with 178 μL 10% iron-limited LNM medium in each well, the treatment was added with 20 μL T. harzianum NJAU4742 iron-limited filtrate and the control was added with 20 μL sterile deionized water, three replicates for each treatment and control. The microtiter plate was cultured at 170 r.p.m. and 30 °C for 12 h, and measured OD600 using a SpectraMax M5 microplate reader (Molecular Devices, USA)57,64.
T. harzianum NJAU4742 selected species of SynCom related to iron mobilization
Microbial DNA of each sample was extracted using the HiPure Soil DNA Kits (Magen, Guangzhou, China) according to the manufacturer’s protocols. The 16S rDNA V3-V4 hypervariable regions of the bacterial 16S rRNA gene were amplified by PCR (95 °C for 5 min, followed by 30 cycles at 95 °C for 1 min, 60 °C for 1 min, and 72 °C for 1 min and a final extension at 72 °C for 7 min) using the primer pair 341 F (5′-CCTACGGGNGGCWGCAG-3′) and 806 R (5′-GGACTACHVGGGTATCTAAT-3′). 50 μL mixture containing 10 μL of 5 × Q5@ Reaction Buffer, 10 μL of 5 × Q5@ High GC Enhancer, 1.5 μL of 2.5 mM dNTPs, 1.5 μL of each primer (10 μM), 0.2 μL of Q5@ High-Fidelity DNA Polymerase (New England Biolabs, USA), and 50 ng of template DNA. Amplicons were extracted from 2% agarose gels and purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, U.S.) according to the manufacturer’s instructions and quantified using ABI StepOnePlus Real-Time PCR System (Life Technologies, Foster City, USA). Purified amplicons were pooled in equimolar and paired-end sequenced (PE250) on an Illumina platform at Guangzhou Genedenovo Biological Technology Co., Ltd. The sequence data were processed following the EasyAmplicon pipeline65. Briefly, primers and barcodes are cut, and then low-quality reads are filtered out to get clean amplicons using Vsearch. And, clean amplicons are denoised into amplicon sequence variants (ASVs) using Unoise3. Finally, the clean amplicons will be mapped to the SynCom 16S rDNA sequence identified ASVs to generate a feature table. Then, we converted the feature table into a species table to match our SynCom isolates.
Synergic effects of C. populi and T. harzianum on iron mobilization
The sterilized soil and tomato seeds preparation were similar to those of the previous pot experiment. C. populi were grown on TSA plate for 2 d at 28 °C. Bacterial suspensions were then prepared by scraping colonies into TSB medium, and shaked at 170 r.p.m. for 16 h at 28 °C. The optical density of C. populi was then adjusted to 1.0 to obtain bacterial suspension. A total of four treatments were set up, including completely sterilized treatment (control), T. harzianum NJAU4742 alone (T), C. populi alone (C. populi), and co-treatment with T. harzianum NJAU4742 and C. populi (T + C. populi). Four replicates were set up for each treatment, and harvested after 25 d.
C. populi suspension (2.5 mL) was taken into 250 mL iron-limited LNM medium, and cultured at 28 °C at 170 r.p.m. for 12 h. The iron-limited filtrate of C. populi (CHRYf) was obtained by centrifuged at 10000 r.p.m. at 4 °C for 20 min, and stored in -80 °C refrigerator after filtered with sterile 0.22 μm filters. Then, 2 μL T. harzianum NJAU4742 spore suspension and 20 μL of the iron-limited filtrate of C. populi were inoculated into a 96-well microtiter plate with 178 μL 10% iron-limited LNM medium in each well. A control treatment was settled as 20 μL sterile deionized water replacing CHRYf. The microtiter plate was cultured at 28 °C. Growth of T. harzianum NJAU4742 was measured (OD750) at 48 h and 72 h, respectively66.
Statistical Analysis
All statistical analyses were performed with R statistical software (version 4.0.3), implemented within the RStudio graphical user interface (RStudio Team, 2023). Part figures were performed online with EVenn67 and BioRender (https://www.biorender.com). Analysis of variance (ANOVA), Tukey’s HSD test, and independent sample t-test (P < 0.05) were performed with multcomp68 package and amplicon65 package. The alpha diversity (richness) was calculated with the ‘diversity’ function of vegan package. The principal coordinate analysis (PCoA) was performed with the ‘pcoa’ functions of the ape package and ‘vegdist’ functions of the vegan package (https://github.com/vegandevs/vegan) based on the species level abundance table, and visualized with the ggplot2 package. Permutational multivariate analysis of variance (PERMANOVA) and analysis of similarities (Anosim) was performed with the ‘adonis2’ and ‘anosim’ function of the vegan package. Constrained analysis of principal coordinates (CAP) was performed with the ‘decorana’, ‘cmdscale’, and ‘envfit’ functions of the vegan and ggplot2 packages. Differential abundance testing was performed with ‘edgeR’ package. Correlation analysis was performed with the ‘stat_cor’ functions of ggpubr package.
Data availability
All sequencing data generated for this manuscript is available from the NCBI SRA under accession PRJNA1100393 and Genbank accession PQ722364.
References
Jeong, J. & Guerinot, M.L. Homing in on iron homeostasis in plants. Trends Plant Sci. 14, 280–285 (2009).
Cuerinot, M.L. & Yi, Y. Iron: Nutritious, Noxious, and Not Readily Available. Plant Physiol. 104, 815–820 (1994).
Kappler, A. et al. An evolving view on biogeochemical cycling of iron. Nat. Rev. Microbiol. 19, 360–374 (2021).
Adeleke, B.S. et al. Rhizosphere Microbiomes Mediating Abiotic Stress Mitigation for Improved Plant Nutrition. Ecologies 5, 375–401 (2024).
Chaudhary, A., et al. Exploring Beneficial Microbes and Their Multifaceted Applications: An Overview. Microb. Inoculants: Appl. Sustain. Agric. 2024, 1–28 (2024).
Datta, S., Hamim, I., Jaiswal, D.K. & Sungthong, R. Sustainable agriculture. BMC Plant Biol. 23, 588 (2023).
Rahimi, S.et al. The role of plant growth-promoting rhizobacteria (PGPR) in improving iron acquisition by altering physiological and molecular responses in quince seedlings. Plant Physiol. Biochem. 155, 406–415 (2020).
Harbort, C.J. et al. Root-Secreted Coumarins and the Microbiota Interact to Improve Iron Nutrition in Arabidopsis. Cell Host Microbe 28, 825–837 (2020).
Kariman, K., Scanlan, C., Boitt, G. & Rengel, Z. Feremycorrhizal symbiosis confers growth and nutritional benefits to mycorrhizal and non-mycorrhizal crops. Soil Biol. Biochem. 151, 108060 (2020).
Berg, G., Kusstatscher, P., Abdelfattah, A., Cernava, T. & Smalla, K. Microbiome Modulation—Toward a Better Understanding of Plant Microbiome Response to Microbial Inoculants. Front. Microbiol. 12, 650610 (2021).
Poppeliers, S.W., Sánchez-Gil, J.J. & De Jonge, R. Microbes to support plant health: understanding bioinoculant success in complex conditions. Curr. Opin. Microbiol. 73, 102286 (2023).
Eltlbany, N. et al. Enhanced tomato plant growth in soil under reduced P supply through microbial inoculants and microbiome shifts. FEMS Microbiol. Ecol. 95, fiz124 (2019).
Finkel, O.M., Castrillo, G., Herrera Paredes, S., Salas González, I. & Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Microbiol. 38, 155–163 (2017).
Krishna, R. et al. Impact of Plant Growth-Promoting Microorganism (PGPM) Consortium on Biochemical Properties and Yields of Tomato Under Drought Stress. Life 14, 1333 (2024).
Harman, G.E., Howell, C.R., Viterbo, A., Chet, I. & Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56 (2004).
Hang, X., et al. Trichoderma-amended biofertilizer stimulates soil resident Aspergillus population for joint plant growth promotion. npj Biofilms Microbio. 8, 57 (2022).
Steyaert, J.M. et al. Yield and cold storage of Trichoderma conidia is influenced by substrate pH and storage temperature. J. Basic Microbiol. 57, 419–427 (2017).
Steyaert, J.M., Weld, R.J. & Stewart, A. Ambient pH intrinsically influences Trichoderma conidiation and colony morphology. Fungal Biol. 114, 198–208 (2010).
Chi, Z. et al. Intrinsic enzyme-like activity of magnetite particles is enhanced by cultivation with Trichoderma guizhouense. Environ. Microbiol. 23, 893–907 (2021).
García-López, A.M., Avilés, M. & Delgado, A. Iron availability thresholds for the inoculation of cucumber with Trichoderma asperellum T34. J. Plant Nutr. Soil Sci. 176, 867–875 (2013).
De Santiago, A., Quintero, J.M., Avilés, M. & Delgado, A. Effect of Trichoderma asperellum strain T34 on iron nutrition in white lupin. Soil Biol. Biochem. 41, 2453–2459 (2009).
De Santiago, A., Quintero, J.M., Avilés, M. & Delgado, A. Effect of Trichoderma asperellum strain T34 on iron, copper, manganese, and zinc uptake by wheat grown on a calcareous medium. Plant Soil 342, 97–104 (2011).
Zhang, F., Xu, X., Wang, G., Wu, B. & Xiao, Y. Medicago sativa and soil microbiome responses to Trichoderma as a biofertilizer in alkaline-saline soils. Appl. Soil Ecol. 153, 103573 (2020).
Fu, J., Xiao, Y., Wang, Y., Liu, Z. & Yang, K. Trichoderma affects the physiochemical characteristics and bacterial community composition of saline–alkaline maize rhizosphere soils in the cold-region of Heilongjiang Province. Plant Soil 436, 211–227 (2019).
Poveda, J. & Eugui, D. Combined use of Trichoderma and beneficial bacteria (mainly Bacillus and Pseudomonas): Development of microbial synergistic bio-inoculants in sustainable agriculture. Biol. Control. 176, 105100 (2022).
Jiao, S., et al. Linking soil fungi to bacterial community assembly in arid ecosystems. iMeta 1, e2 (2022).
Xiao, D. et al. Network analysis reveals bacterial and fungal keystone taxa involved in straw and soil organic matter mineralization. Appl. Soil Ecol. 173, 104395 (2022).
Li, H. et al. Microbiomes across root compartments are shaped by inoculation with a fungal biological control agent. Appl. Soil Ecol. 170, 104230 (2022).
Zhang, L. et al. Trichoderma spp. promotes ginseng biomass by influencing the soil microbial community. Front. Microbiol. 15, 1283492 (2024).
Tao, C. et al. Additive fungal interactions drive biocontrol of Fusarium wilt disease. N. Phytol. 238, 1198–1214 (2023).
Niu, B., Paulson, J.N., Zheng, X. & Kolter, R. Simplified and representative bacterial community of maize roots. Proc. Natl. Acad. Sci. USA. 114, E2450–E2459 (2017).
Emmenegger, B., et al. Identifying microbiota community patterns important for plant protection using synthetic communities and machine learning. Nat. Commun. 14, 7983 (2023).
Qu, Q. et al. Rhizosphere Microbiome Assembly and Its Impact on Plant Growth. J. Agric. Food Chem. 68, 5024–5038 (2020).
Vogel, C.M., Potthoff, D.B., Schäfer, M., Barandun, N. & Vorholt, J.A. Protective role of the Arabidopsis leaf microbiota against a bacterial pathogen. Nat. Microbiol. 6, 1537–1548 (2021).
Tyśkiewicz, R., Nowak, A., Ozimek, E. & Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 23, 2329 (2022).
Zhang, Y. et al. Pre-colonization of PGPR triggers rhizosphere microbiota succession associated with crop yield enhancement. Plant Soil 439, 553–567 (2019).
Santoyo, G. How plants recruit their microbiome? New insights into beneficial interactions. J. Adv. Res. 40, 45–58 (2022).
Del Carmen Orozco-Mosqueda, M. et al. Arthrobacter agilis UMCV2 induces iron acquisition in Medicago truncatula (strategy I plant) in vitro via dimethylhexadecylamine emission. Plant Soil 362, 51–66 (2013).
Hartmann, A. & Schikora, A. Quorum Sensing of Bacteria and Trans-Kingdom Interactions of N-Acyl Homoserine Lactones with Eukaryotes. J. Chem. Ecol. 38, 704–713 (2012).
Abdalla, A.K. et al. Exopolysaccharides as Antimicrobial Agents: Mechanism and Spectrum of Activity. Front. Microbiol. 12, 664395 (2021).
Zhou, J., Zhang, L., Feng, G. & George, T.S. Arbuscular mycorrhizal fungi have a greater role than root hairs of maize for priming the rhizosphere microbial community and enhancing rhizosphere organic P mineralization. Soil Biol. Biochem. 171, 108713 (2022).
Wang, C. & Kuzyakov, Y. Mechanisms and implications of bacterial–fungal competition for soil resources. ISME J. 18, wrae073 (2024).
Xiao, L. et al. Biodegradation and adsorption of benzo[a]pyrene by fungi-bacterial coculture. Ecotoxicol. Environ. Saf. 283, 116811 (2024).
Jiang, F., Zhang, L, Zhou, J., George, T.S.& Feng, G.Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. N. Phytol. 230, 304–315 (2021).
Pierce, E.C. et al. Bacterial–fungal interactions revealed by genome-wide analysis of bacterial mutant fitness. Nat. Microbiol. 6, 87–102 (2020).
Qiao, C. et al. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Appl. Soil Ecol. 142, 136–146 (2019).
Shade, A. & Handelsman, J. Beyond the Venn diagram: the hunt for a core microbiome: The hunt for a core microbiome. Environ. Microbiol. 14, 4–12 (2012).
Nguyen, J., Lara-Gutiérrez, J. & Stocker, R. Environmental fluctuations and their effects on microbial communities, populations and individuals. FEMS Microbiol. Rev. 45, fuaa068 (2021).
Louca, S. et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2, 936–943 (2018).
Radzki, W. et al. Bacterial siderophores efficiently provide iron to iron-starved tomato plants in hydroponics culture. Antonie van. Leeuwenhoek 104, 321–330 (2013).
Ambrosini, A. et al. Screening of plant growth promoting Rhizobacteria isolated from sunflower (Helianthus annuus L.). Plant Soil 356, 245–264 (2012).
De Souza, R., Meyer, J., Schoenfeld, R., Da Costa, P.B. & Passaglia, L.M.P. Characterization of plant growth-promoting bacteria associated with rice cropped in iron-stressed soils. Ann. Microbiol. 65, 951–964 (2015).
Miao, Y., et al. Proteomic analysis reflects an environmental alkalinization-coupled pH-dependent mechanism of regulating lignocellulases in Trichoderma guizhouense NJAU4742. Biotechnol. Biofuels 13, 6 (2020).
Büttner, H. et al. Bacterial endosymbionts protect beneficial soil fungus from nematode attack. Proc. Natl. Acad. Sci. USA. 118, e2110669118 (2021).
Dahlstrom, KM & Newman, DK Soil bacteria protect fungi from phenazines by acting as toxin sponges. Curr. Biol. 32, 275–288 (2022).
Gandhi Pragash, M., Badri Narayanan, K., Ravindra Naik, P. & Sakthivel, N. Characterization of Chryseobacterium aquaticum Strain PUPC1 Producing a Novel Antifungal Protease from Rice Rhizosphere Soil. J. Microbiol. Biotechnol. 19, 99–107 (2009).
Gu, S. et al. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. Nat. Microbiol. 5, 1002–1010 (2020).
Finkel, O.M. et al. A single bacterial genus maintains root growth in a complex microbiome. Nature 587, 103–108 (2020).
Schmitz, L. et al. Synthetic bacterial community derived from a desert rhizosphere confers salt stress resilience to tomato in the presence of a soil microbiome. ISME J. 16, 1907–1920 (2022).
Xun, W., et al. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 9, 35 (2021).
Schwyn, B. & Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56 (1987).
Dunn, W.B. et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 6, 1060–1083 (2011).
Ding, W. et al. Comparative physiological and metabolomic analyses of the hyper-accumulation of astaxanthin and lipids in Haematococcus pluvialis upon treatment with butylated hydroxyanisole. Bioresour. Technol. 292, 122002 (2019).
Gu, S. et al. Siderophore-Mediated Interactions Determine the Disease Suppressiveness of Microbial Consortia. mSystems 5, e00811–e00819 (2020).
Liu, Y. et al. EasyAmplicon: An easy-to-use, open-source, reproducible, and community-based pipeline for amplicon data analysis in microbiome research. iMeta 2, e83 (2023).
Cruz-Magalhães, V. et al. The NADPH Oxidases Nox1 and Nox2 Differentially Regulate Volatile Organic Compounds, Fungistatic Activity, Plant Growth Promotion and Nutrient Assimilation in Trichoderma atroviride. Front. Microbiol. 9, 3271 (2019).
Chen, T., Zhang, H., Liu, Y., Liu, Y.-X. & Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 48, 863–866 (2021).
Hothorn, T., Bretz, F. & Westfall, P. Simultaneous Inference in General Parametric Models. Biometr. J. 50, 346–363 (2008).
Acknowledgements
This research was funded by the National Natural Science Foundation of China (42477125 and 42107140), Guangxi Science and Technology Program (GuiKe AA24010003), Technology Innovation Special Fund of Jiangsu Province for Carbon Dioxide Emission Peaking and Carbon Neutrality (BE2022423), the Jiangsu Agricultural Science and Technology Innovation Fund (CX(22)1004), China Agriculture Research System (CARS-28-10).
Author information
Authors and Affiliations
Contributions
Y.S., C.D., Z.W., and Q.S. designed the experiments. Y.S., H.P., S.L., and L.Z. performed the experiments. Y.S. and S.G. analyzed the data. Y.S., S.G., R.B. and T.Y. wrote and revised the paper with editorial input from all other authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Shao, Y., Gu, S., Peng, H. et al. Synergic interactions between Trichoderma and the soil microbiomes improve plant iron availability and growth. npj Biofilms Microbiomes 11, 56 (2025). https://doi.org/10.1038/s41522-025-00684-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41522-025-00684-z
