
Abstract
The gut microbiota significantly influences cancer immunotherapy efficacy by modulating immune responses, remodeling the tumor microenvironment (TME), and producing key metabolites. Strategies such as FMT, probiotics, and dietary interventions show promise in enhancing responses to ICIs and ACTs while reducing immune-related adverse events (irAEs). This review summarizes clinical and preclinical findings and discusses microbiota-based interventions and future directions for precision immunotherapy.
Similar content being viewed by others
Introduction
In 2001, the concept of “microbial community” was first introduced, which mainly consists of 40 trillion microorganisms, including bacteria, fungi, archaea, and viruses, with more than 100 times as many genes as the human genome1,2,3,4. Among them, the gut microbiota is particularly significant, colonizing the colon and comprising over 97% of the total microbial population2,5. Given that it contains up to 60-70% of peripheral immune cells and interacts with a substantial number of microbiota, the gut is regarded as the largest peripheral immune organ in the body6,7. These microorganisms help maintain intestinal homeostasis through metabolic and immune-mediated pathways, influencing key physiological processes such as metabolism, inflammation, immunity, and neurology8,9,10. For instance, microbial metabolites such as short-chain fatty acids (SCFAs) and bile acids (BAs) play crucial roles in shaping both innate and adaptive immune responses11,12,13.
Disruptions in gut microbial composition have been linked to cancer development through multiple mechanisms, including genomic instability, alterations in the tumor microenvironment, and modulation of immune evasion strategies14,15. Malignant tumors remain the second leading cause of death globally, and while conventional treatments like chemotherapy, radiotherapy, and surgery have improved patient outcomes, they are often limited by drug resistance, recurrence, and adverse side effects. In recent years, immunotherapies—such as immune checkpoint inhibitors (ICIs) and adoptive cell transfer therapies (ACTs)—have revolutionized cancer treatment16,17. However, responses to these therapies vary significantly among patients, with some exhibiting primary or acquired resistance, underscoring the need for novel adjuvant approaches18,19,20.
In recent years, the influence of the gut microbiota on tumorigenesis and therapeutic response has been gradually uncovered with the help of next-generation sequencing (NGS) and third-generation sequencing technologies21,22,23. Specific microbial compositions not only differentiate cancer patients from healthy individuals but also correlate with immunotherapy outcomes, making them potential biomarkers for predicting treatment efficacy24,25. Strategies to modulate gut microbiota—such as fecal microbiota transplantation (FMT), probiotics, prebiotics, and dietary interventions—have shown promise in enhancing immunotherapy response and mitigating treatment-related toxicity26,27,28,29,30. Additionally, tumor-associated microbiota within the tumor microenvironment (TME) may influence therapeutic outcomes by modulating inflammation, promoting immune cell infiltration, and enhancing drug-induced immunogenic cell death31,32,33. These findings have opened new avenues for microbiome-based precision oncology.
Despite these advancements, the mechanisms underlying interactions between the gut microbiota and tumor immunity remain incompletely understood, and research on microbiota-driven immunotherapy optimization is still in its early stages. A deeper understanding of the molecular and systems biology functions of gut microbes is critical for identifying novel therapeutic targets and improving clinical applications. In this review, we examine the role of the gut microbiota in shaping immunotherapy efficacy, focusing on ICIs and ACTs, and exploring its impact on treatment-related toxicity. We also discuss emerging microbiota-targeted interventions and personalized therapeutic strategies, highlighting key studies that have advanced the field. Finally, we address current challenges and future perspectives in leveraging gut microbiota for cancer immunotherapy.
The evolving role of gut microbiota in cancer immunotherapy: a historical perspective
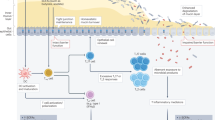
Immunotherapies, particularly ICIs and ACTs—including T-cell receptor-engineered T cells (TCR-T) and chimeric antigen receptor T cells (CAR-T)—have transformed cancer treatment and significantly improved patient outcomes34,35,36. These therapies work by activating or modulating the immune system to enhance anti-tumor responses. However, their efficacy varies widely across cancer types and patient populations, with key challenges such as antigen escape, limited transport, and insufficient tumor infiltration affecting treatment success37,38,39,40,41. Additionally, tumor mutation burden (TMB)—the total number of somatic mutations per coding region of a tumor genome—has emerged as a significant biomarker predicting immunotherapy response. Tumors with higher TMB, such as melanoma and non-small-cell lung cancer (NSCLC), tend to generate more neoantigens, thereby enhancing tumor immunogenicity and responsiveness to ICIs42. Conversely, cancers with lower TMB, such as prostate and pancreatic cancer, often exhibit reduced neoantigen formation, resulting in limited immune recognition and decreased efficacy of immunotherapies43. Emerging clinical studies have identified a strong correlation between gut microbiota composition and immunotherapy outcomes in cancer patients (Fig. 1 and Table 1)44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89. Fecal sample analyses suggest that variations in gut microbial communities may influence treatment efficacy through underlying biological mechanisms. These studies demonstrate the utility of microbiota interventions in clinical practice. However, given the complexity of the gut microbiota and significant individual heterogeneity, translation of microbiota-based insights into clinical practice remains limited. Further investigation is needed to fully elucidate these interactions and harness the gut microbiota to optimize immunotherapy strategies.

ACT, adoptive cell therapy; PD-L1, programmed cell death ligand 1; CTLA-4, cytotoxic T lymphocyte-associated antigen-4; ICI, immune checkpoint inhibitor; Rs, responders; NRs, non-responders; TME, tumor microenvironment; CAR-T, chimeric antigen receptor T-cell therapy; NGS, next generation sequencing. Early mouse studies (2007–2013) demonstrated that gut microbiota can enhance anti-tumor immune responses, whereas antibiotic treatment attenuates the efficacy of ACT. In 2015, research first revealed a direct relationship between gut microbiota and ICI response, identifying specific bacterial species such as Bifidobacterium and Bacteroides fragilis as promoters of ICI efficacy. Subsequent preclinical and clinical studies in 2018 explored the potential of gut microbiota and its metabolites as predictive markers for the effectiveness of ICIs and ACTs. Clinical breakthroughs between 2020 and 2021 further demonstrated that fecal microbiota transplantation (FMT) could enhance ICI response, while gut microbial metabolites were found to improve the activity of CAR-T cells. More recent retrospective and prospective studies (2022–2023) have established correlations between gut microbiome composition and CAR-T cell therapy efficacy, leveraging next-generation sequencing (NGS) and machine learning algorithms for predictive analysis. In 2024–2025, researchers focus on multi-omics and metabolic mechanisms underlying gut microbiota interactions with immunotherapy.
Gut microbiota and ICIs: modulation, response and therapeutic potential
ICIs work by enhancing the body’s immune response against tumors, primarily by blocking inhibitory immune checkpoints such as programmed cell death protein 1 (PD-1), its ligand PD-L1, and cytotoxic T lymphocyte-associated antigen-4 (CTLA-4). Currently, ICIs have been widely used in the treatment of a variety of cancers. Apart from mismatch repair deficient (dMMR) colorectal cancer, which is consistently responsive to ICIs, in general, only 20-40% of patients (including the most responsive tumor types such as melanoma, lung cancer and renal cell carcinoma) are susceptible to ICIs45,46. Early preclinical research suggested that gut microbiota plays a key role in modulating anti-tumor immune responses90,91,92. In 2015, Sivan et al. first reported that commensal bacteria influence both spontaneous immune responses to tumors and the effectiveness of ICIs93. They demonstrated that oral administration of Bifidobacterium enhanced the anti-tumor efficacy of PD-L1 inhibitors by promoting dendritic cell (DC) maturation and increasing tumor-specific CD8 + T cell activity. Another study that same year showed that antibiotic treatment could reduce the effectiveness of ICIs, while supplementation with Bacteroides fragilis in melanoma models restored anti-CTLA-4 efficacy by stimulating Th1 cell activation in tumor-draining lymph nodes and enhancing intra-tumoral dendritic cell maturation94.
By 2018, clinical trials confirmed that gut microbiota composition and diversity could predict ICI response25,45. Patients with non-small cell lung cancer (NSCLC) and renal cell carcinoma (RCC) who had higher gut microbial diversity responded better to anti-PD-1 therapy25. In metastatic melanoma, responders exhibited increased levels of Bifidobacterium longum, Collinsella aerofaciens, and Enterococcus faecium, and fecal microbiota transplantation (FMT) from these patients into germ-free mice improved anti-PD-L1 therapy outcomes45. Retrospective studies further linked antibiotic use to reduced ICI efficacy and lower survival rates in patients with advanced solid tumors, reinforcing the role of gut microbiota in shaping anti-tumor immune responses95,96. Prospective studies have since demonstrated significant correlations between microbiome composition and ICI outcomes in melanoma, NSCLC, and hepatocellular carcinoma (HCC)46,47,48,49. Notably, researchers showed that FMT from ICI responders, combined with anti-PD-1 therapy, could overcome resistance in patients with refractory melanoma55,74. The first phase I FMT clinical trial further highlighted the role of gut microbiota in modulating responses to anti-PD-1 therapy54. Researchers then focused on multi-omics and metabolite mechanisms. Zhu et al. characterized five response-associated gut phenotypes through multi-omics analysis, in which the metabolite phenylethyl fluoride was negatively associated with response and shown to attenuate anti-PD-1 efficacy in vivo, elucidating the interplay between gut microbiota, gut metabolome, and immunotherapy response97.
Despite concerted efforts by researchers to combine diverse cohorts and expand sample sizes for studying microbiota associated with immunotherapy63, identifying consistent beneficial microbial markers remains challenging. This difficulty stems from multiple confounding factors, including individual and environmental variability, differences in clinical trial design, and methodological inconsistencies. Moreover, gut microbiota linked to response varies by ICI type—bacteria associated with anti-PD-1 and anti-PD-L1 efficacy may differ from those influencing anti-CTLA-4 response, reflecting distinct mechanisms of immune modulation54,93,98. For example, Gunjur et al. suggest that future development of gut microbiota diagnostics or therapeutics should be tailored to ICB treatment protocols rather than to cancer type. Tumor type further influences microbiome interactions, resulting in variability among patients99,100,101. For example, in advanced melanoma, high levels of Bifidobacterium, Coriobacteriaceae, Ruminococcaceae, and Lachnospiraceae correlate with favorable anti-PD-1 responses25,45,46,47,62. In contrast, NSCLC, RCC, and HCC patients with greater abundance of Akkermansia muciniphila exhibit improved anti-PD-1 efficacy25,102. Interestingly, Akkermansia muciniphila has also been linked to resistance in antibiotic-treated patients, suggesting a dual role in ICI response103. Most studies to date have relied on metagenomic sequencing of fecal samples to identify microbial species associated with ICI response. However, a comprehensive analysis at the subspecies or strain level has yet to be conducted. Recent studies have revealed that two branches of the same bacterial subspecies do not play identical roles in tumorigenesis and progression, which underscore the necessity for finer-grained studies to elucidate the precise interactions between the microbiome and ICI, with the objective of optimizing the effects of immunotherapy104.
Gut microbiota and ACTs: influence on efficacy and toxicity
ACTs, refer to collecting and modifying a patient’s immune cells in vitro to enhance their tumor-killing ability before reinfusion into the body, include T-cell receptor (TCR) therapy, chimeric antigen receptor (CAR) T-cell therapy, natural killer (NK) cell therapy, and tumor-infiltrating lymphocyte (TIL) therapy. Many studies have shown that ACTs, particularly CAR-T cell therapy, has demonstrated significant efficacy in hematologic malignancies and melanoma105,106.
The influence of gut microbiota on ACT efficacy was first discovered in 2007 when researchers found that the broad-spectrum antibiotic ciprofloxacin reduced the therapeutic effectiveness of ACT in mice. This effect was reversed by supplementing bacterial lipopolysaccharide (LPS), which activates the Toll-like receptor (TLR)-4 pathway107. Subsequent studies have provided further indirect evidence linking gut microbiota to ACT outcomes. In 2017, a study showed that vancomycin treatment, which increased Proteobacteria while depleting Firmicutes and Bacteroidetes, enhanced ACT efficacy by promoting CD8α + DCs and upregulating interleukin (IL)-12. In contrast, antibiotics such as neomycin and metronidazole, which deplete Gram-negative aerobic and anaerobic bacteria, respectively, impaired ACT efficacy108. This suggests that gut microbiota modulation may improve ACT effectiveness. In 2018, it was shown that gut microbiota-mediated bile acid metabolism increased CXCR6 + NKT cells in the liver, contributing to the anti-tumor activity of HCC109. In the same year, researchers found that SCFA, another key microbial metabolite, could strengthen anti-tumor immunity of ACT by inhibiting histone deacetylase to increase the cytotoxicity of CD8+ T cells110. Since then, researchers have gradually focused on the effects of gut microbial metabolites on the efficacy of ACT and demonstrated these findings111,112,113.
During this time, as ACT therapies evolved, several retrospective studies found that gut microbiota were associated with ACT toxicity and had potential as biomarkers. One study analyzing plasma samples from colorectal cancer (CRC) patients treated with ACT and chemotherapy found that responders had significantly higher blood levels of Bifidobacterium, Lactobacillus, and Enterococcus, indicating that the blood microbiome may serve as a biomarker for immunotherapy response60. In 2022, Lauren et al. found that patients treated with piperacillin/tazobactam, meropenem, or imipenem/cilastatin (P-I-M) antibiotics within four weeks before CD19-targeted CAR-T therapy had lower survival rates and higher neurotoxicity risks. Sequencing data identified Ruminococcus, Bacteroides, and Faecalibacterium as key microbial taxa associated with CAR-T efficacy, with bacterial metabolic pathways such as peptidoglycan synthesis and pentose phosphate metabolism emerging as potential biomarkers for response monitoring70. In the same year, Melody et al. found that the composition of the gut microbiota appeared to change at different stages of CAR-T therapy. In multiple myeloma (MM) patients, microbial diversity and abundance varied between those in complete remission (CR) and partial remission (PR), with Bifidobacterium, Prevotella, Sutterella, and Collinsella showing significant fluctuations. Moreover, patients who developed severe cytokine release syndrome (CRS) exhibited higher levels of Bifidobacterium, Leuconostoc, Stenotrophomonas, and Staphylococcus, suggesting a potential role of gut microbes in CAR-T-associated toxicities71. Targeted microbiota modulation could offer a strategy to mitigate these side effects.
Additionally, gut microbes can shape TME by promoting chemokine secretion, recruiting tumor-infiltrating T cells, and improving patient prognosis114. Notably, intratumoral injection of Escherichia coli MG1655 has been shown to normalize tumor vasculature and reprogram tumor-associated macrophages (TAMs) to the M1 phenotype, leading to increased chemokine CCL5 production and enhanced T cell infiltration of ACTs115. This approach, which combined bacterial therapy targeting the interior of solid tumors with ACT targeting the periphery of tumors, successfully eradicated early melanoma and slowed pancreatic tumor progression. Current research on the interaction between gut microbiota and ACTs is still in its early stages, with most studies focused on hematologic malignancies. The role of gut microbiota in ACTs efficacy for solid tumors remains unexplored, and the dominant bacterial species and underlying mechanisms require further investigation. In the future, interventions such as microbiota transplantation or metabolite supplementation before T-cell activation could enhance the effectiveness of ACTs and reduce treatment-related toxicity.
Gut microbiota and CpG-ODN immunotherapy: modulation and clinical implications
Pattern recognition receptors (PRRs) play a crucial role in initiating and sustaining innate immunity. Among them, TLRs, nucleotide-binding oligomeric domain-like receptors (NLRs), and C-type lectin receptors (CLRs) are key mediators116. Innate immune cells such as DCs and NK cells express PRRs to detect pathogen-associated molecular patterns, bridging innate and adaptive immune responses. CpG oligodeoxynucleotides (CpG-ODNs), synthetic immunomodulatory agents, enhance immune responses by activating TLR9, thereby stimulating both cellular and humoral immunity117,118.
Studies have demonstrated the gut microbiota’s essential role in CpG-ODN-based immunotherapy. Guiducci et al. found that CpG-ODN treatment in tumors rapidly shifts macrophages from an immunosuppressive M2 phenotype to a pro-inflammatory M1 state, producing high levels of TNF and IL-12, leading to tumor necrosis. However, this effect was absent in sterile or antibiotic-treated mice, suggesting that gut microbiota–derived LPS activate TLR4, priming myeloid cells in tumors to respond to TLR9 stimulation119. Similarly, Iida et al. reported that disrupting the microbiota impaired tumor responses to CpG-ODNs and platinum-based chemotherapy120. Their findings linked specific bacterial species to TNF production, where Gram-negative Ruminococcus (e.g., A. shahii) enhanced TNF expression, while Gram-positive Lactobacillus species (e.g., L. murinum, L. intestinalis, L. fermentum) were negatively associated. Notably, restoring A. shahii in antibiotic-treated mice reinstated TNF production, underscoring the microbiota’s role in mediating immune responses. The influence of gut microbiota on CpG-ODN immunotherapy has been further validated in clinical trials. A phase II study in melanoma patients receiving CpG-ODN combined with ICIs revealed that responders had an enriched population of Gram-negative Bacteroidaceae, Enterobacteriaceae, and Firmicutes, reinforcing the microbiota’s role in modulating bone marrow-derived immune functions within the TME85.
Although the precise mechanisms linking gut microbiota to tumor immunotherapy remain under investigation, emerging research suggests that distinct microbial subtypes influence clinical outcomes, with shared genetic traits modulating host immune responses62. Future studies should focus on identifying key bacterial strains and their metabolites to develop targeted microbiota-based strategies for enhancing immunotherapy efficacy.
Gut microbiota and cancer immunotherapy: mechanisms of immune modulation
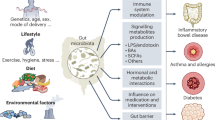
As mentioned earlier, gut microbes can modulate immunotherapy responses, and the mechanisms by which gut microbiota modulate anti-tumor immune responses also have important implications, with roles involving the immunomodulation of the body, including the immune system and the TME, by the microbiota, either directly or through its metabolites, and by exerting interactions with immunotherapy (Fig. 2).

DC, dendritic cell; MHC-I, major histocompatibility complex class I; MHC-II, major histocompatibility complex class II; TCR, T cell receptor; PRR, pattern recognition receptor; NK, natural killer; CTL, cytotoxic T lymphocyte. Within the gut, the gut microbiota plays a crucial role in maintaining the intestinal barrier, protecting against pathogens, and shaping immune responses. Certain bacterial species, such as Bifidobacterium and Bacteroides fragilis, can interacting with DCs and activate CD8 + T cells and Th1 cells. Additionally, invasive microbiota and microbiota-derived metabolites—including lipopolysaccharides (LPS) and peptidoglycans—activate PRRs, promoting macrophage polarization toward the pro-inflammatory M1 phenotype and enhancing T cell activation. Specific microbial strains, such as Lactobacillus plantarum and Bifidobacterium, also stimulate NK cell activation, reinforcing antitumor immunity. Systemically, these microbiota-driven immune responses extend beyond the gut, increasing CTL infiltration into distant tumor sites, particularly within the tumor microenvironment (TME). Notably, Bifidobacterium, Clostridiales, and certain microbial metabolites enhance CD8 + T cell infiltration into tumor tissues, strengthening the immune response against cancer. Furthermore, gut microbiota-derived antigens share similar epitopes with tumor-associated antigens, promoting cross-reactivity and amplifying tumor-targeting immune responses. Conversely, some microbial metabolites may inhibit T cell recognition of tumor epitopes, potentially dampening anti-tumor immunity.
Shaping innate and adaptive immunity
Growing evidence indicates that gut microbiota plays a crucial role in regulating innate immunity by influencing immune signaling and maintaining gut barrier integrity. The microbiota shapes the development and function of key innate immune cells, including DCs and macrophages. Disruptions in microbial balance weaken the intestinal mucosal barrier, allowing microbial products such as lipopolysaccharides (LPS), peptidoglycan, and flagellin to enter the bloodstream. These microbial components activate pattern recognition receptors (PRRs), such as TLRs and NLRs, triggering proinflammatory cytokine production, DC maturation, and NK cell activation, all of which contribute to immune responses and inflammation121. Additionally, specific gut bacteria activate PRRs within gut-associated lymphoid tissue (GALT) and mesenteric lymph nodes, further stimulating DCs and enhancing immune function122. For example, Bifidobacterium bifidum and Bifidobacterium fragilis promote innate immunity by interacting with DCs and macrophages through their metabolites and stimulating the release of cytokines such as IL-12 and TNF-α92,93,123. Gut microbiota also influence innate immune signaling through NLRs. Studies suggest that commensal bacteria help regulate intestinal inflammation by interacting with these receptors124,125,126. In addition, lactobacillus plantarum can enhance the expression of natural cytotoxicity receptors (NCRs), activating NK cells and reinforcing innate immunity127. Furthermore, the microbiota plays a role in shaping the phenotype and function of innate lymphoid cells (ILCs), particularly ILC3s, further underscoring its importance in immune regulation128. For instance, Gut microbiota metabolites can directly modulate ILC3 functions, influencing cytokine production, tissue repair, and inflammation regulation129. These interactions subsequently alter the immune microenvironment, which can enhance or impair responses to cancer immunotherapy. Recent studies highlight that microbial-derived aryl hydrocarbon receptor (AhR) ligands enhance ILC3 functionality, promoting anti-tumor immunity and improving immunotherapeutic outcomes130.
Beyond innate immunity, gut microbiota contributes to the development of adaptive immune responses, particularly through molecular mimicry and thus stimulation of T cell activation and differentiation, which in turn influence systemic immune homeostasis131,132,133. Studies suggest that gut microbes regulate CD4 + T cell subtypes, including Th1, Th2, Th17, and regulatory T cell (Treg)134,135,136. Germ-free (GF) mice, for instance, exhibit an imbalance skewed toward a Th2-dominant response137. Certain bacterial species, such as Bacteroides fragilis, produce polysaccharide A (PSA), which engage TLR 2/1 heterodimers and Dectin-1, triggering downstream phosphoinositide 3-kinase signaling138. This cascade leads to the production of the immunomodulatory cytokine IL-10 by CD4+ Tregs, contributing to immune homeostasis. The gut microbiota also influences intestinal CD8 + T cell function, which extends to the regulation of peripheral immune cells139,140,141,142. Moreover, the gut microbiota has been shown to play a dual role in cancer progression by mediating B cell responses and thus modulating immune responses. On the one hand, B-cell responses that are indirectly induced by specific microbiota (e.g., Bifidobacterium bifidum, Ackermannia spp.) through activation of DC and CD8 + T cells may promote antitumor immunity through the production of tumor-reactive antibodies, the enhancement of antigen presentation, and the promotion of cytotoxic T-cell responses143,144. Conversely, under certain conditions, certain intestinal flora (e.g., Mycobacterium spp.) may attenuate T-cell-mediated antitumor responses by inducing regulatory B-cell differentiation and secreting immunosuppressive factors such as IL-10 and TGF-β143. The specific microbial communities and their metabolites play a pivotal role in determining whether B cells exert tumor-promoting or tumor-suppressive effects.
Disruptions in microbiota-T cell interactions can create a proinflammatory environment within and beyond the gastrointestinal tract, including the TME. These insights highlight the intricate crosstalk between the gut microbiota and host immunity, offering potential therapeutic avenues to enhance immunotherapy efficacy.
Harnessing metabolites for the regulation of anti-tumor immunity
Gut microbiota influence both systemic and tumor-specific immune responses through their metabolites—small molecules that diffuse from the gut and modulate the immune system145. These microbial metabolites can be broadly categorized based on their origin and synthesis: (1) metabolites derived from dietary components, such as SCFAs, tryptophan metabolites, and inosine; (2) host-produced metabolites modified by gut microbiota, such as secondary bile acids; and (3) metabolites synthesized directly by gut microbiota, including branched-chain amino acids (BCAAs) and vitamins146.
SCFAs, such as butyrate and propionate, play a key role in immune regulation. Bacteroidetes, Lachnospiraceae, and Roseburia produce propionic acid via the succinate or acrylic acid pathways, while Firmicutes species, including Ruminococcus and Clostridium, generate butyric acid147,148. These SCFAs are absorbed in the colon and influence T cell differentiation through G-protein-coupled receptor signaling and histone deacetylase inhibition149,150. Similarly, tryptophan metabolism by gut microbiota yields various bioactive compounds, such as indoles, 5-hydroxytryptamine, and kynurenine, which exert bidirectional effects on immune responses151. For example, Lactobacillus reuteri metabolizes tryptophan in the TME to produce indole-3-aldehydes, which enhance CD8+ T cell activity and inhibit melanoma growth via AhR signaling152. However, in pancreatic ductal adenocarcinoma (PDAC) models, Lactobacillus reuteri-derived tryptophan metabolites activate TAMs, promoting an immunosuppressive phenotype153. These findings highlight the need for tumor-specific intervention strategies. Other microbial metabolites also modulate immune responses. Inosine, a purine metabolite produced by Akkermansia muciniphila and Bifidobacterium longum, enhances antitumor immunity by inhibiting UBA6 expression in tumor cells, thereby increasing tumor immunogenicity154. Additionally, inosine interacts with adenosine 2A receptors on T lymphocytes, leading to IL-12 receptor β2 activation and IFN-γ upregulation, further strengthening immune responses155.
Gut microbiota-derived lipid metabolites, particularly sphingolipids such as ceramides and sphingosine-1-phosphate (S1P), also significantly modulate immune responses and influence cancer immunotherapy efficacy156,157,158. Sphingolipids not only serve as structural components of cellular membranes but also act as signaling molecules that regulate cell behavior and immune cell communication. Specific gut microbes, including Bacteroidetes, produce sphingolipids that shape host immune responses157,159. These microbiota-derived sphingolipids can either enhance or impair the efficacy of immunotherapies by modulating immune cell trafficking, proliferation, and activation. For instance, S1P has been shown to regulate lymphocyte egress from lymphoid tissues, influencing their availability at tumor sites, thus directly impacting immunotherapy outcomes160. The immunomodulatory roles of these lipid metabolites underscore the importance of exploring microbial lipid metabolism further to develop targeted strategies enhancing the effectiveness of cancer immunotherapies.
Bile acids, primarily converted by gut microbiota in the cecum and colon, also influence immune function. Species within Firmicutes, particularly Clostridium, convert primary bile acids into secondary bile acids such as lithocholic acid (LCA) and deoxycholic acid (DCA)161. These metabolites regulate T cell differentiation and macrophage polarization, with secondary bile acids notably suppressing Th17 cell function162,163. However, their effects on tumor immunity are complex. For example, secondary bile acids from Clostridium scindens impair the tumor-suppressive function of NKT cells109. In contrast, LCA inhibits breast cancer cell growth by activating TGR5 and constitutive androstane receptors, leading to oxidative stress, suppression of epithelial-mesenchymal transition (EMT), and reduced angiogenesis and metastasis164. Gut microbiota also produce BCAAs and vitamins that influence anti-tumor immune response. Bacteria degrade proteins and unabsorbed amino acids to synthesize BCAAs such as leucine, isoleucine, and valine, which promote effector T cell activation and proliferation through mTORC1 signaling165. Meanwhile, Firmicutes (such as Lactobacillus and Enterococcus) and Bacteroides synthesize B vitamins. Bacteroides fragilis, Prevotella, and Ruminococcus lactis produce vitamin B3, which binds to GPR109A receptors in myeloid cells, inhibiting NF-κB signaling, reducing immunosuppressive myeloid cell polarization and enhancing CD8 + T cell cytotoxicity166,167. Likewise, vitamin B6, synthesized by Lactobacillus acidophilus and Bifidobacterium bifidum, boosts T lymphocyte proliferation and promotes antitumor immunity by stimulating T cell activity168,169.
These findings underscore the critical role of microbial metabolites in shaping antitumor immune responses. However, their bidirectional effects and individual variability present challenges in therapeutic applications. To maximize their potential, future research must integrate multi-omics data and develop personalized strategies to optimize tumor immunotherapy with minimal toxicity.
Synergizing ICI to enhance antitumor efficacy and managing risks
As previously mentioned, the gut microbiota has the capacity to augment the efficacy of immunotherapy by modulating host immunity. However, several studies have demonstrated that the gut microbiota can also interact with immunotherapy to further enhance its efficacy. Vétizou et al. found that Bacteroides thetaiotaomicron and Bacteroides fragilis stimulate TLR4- and IL-12- dependent T helper 1 (Th1) responses, enhancing the effects of CTLA-4 blockade94. Other studies have shown that Firmicutes and Actinobacteria activate mucosa-associated innate T cells and CD56 + CD8+ T cells in peripheral blood, increasing the expression of human leukocyte antigen (HLA) class II genes, CD74, and granzyme K (GZMK) in CD8+ T cells at tumor sites, thereby improving anti-PD-1 responses55. Certain bacterial species also facilitate immune cell infiltration into tumors, further amplifying the effects of immunotherapy. Enterococcus hirae migrates from the small intestine to secondary lymphoid organs, increasing the intratumoral CD8 + T/Treg ratio, while Bacteroides intestinihominis accumulates in the colon and stimulates IFN-γ-producing γδ T cells, improving cyclophosphamide efficacy in advanced lung and ovarian cancer patients115,170. Similarly, Akkermansia and high-fiber diets have been shown to induce tumor-infiltrating NK cell-DC interactions through stimulator of interferon genes (STING) signaling, leading to type I interferon (IFN-I) production and enhanced ICI efficacy171. Additionally, Bifidobacterium preferentially accumulates in tumors and promotes CD47-based immunotherapy in a STING- and interferon-dependent manner172. Beyond direct immune modulation, the commensal microbiota may also activate tumor antigen-specific T cell responses by modulating the immunogenicity of tumor cells. Microbiota can enhance ICI efficacy by providing tumor cross-antigens that stimulate tumor-specific T cell responses131,173,174. Fluckiger et al. found that a tail-length tapering protein (TMP) from an Enterococcus phage closely resembles the proteasomal subunit β-4, triggering TMP-specific CD8+ T cell responses during cyclophosphamide or anti-PD-1 therapy173. Similarly, Bessell’s team identified an antigenic epitope (SVYRYYGL) in Bifidobacterium breve that mimics the tumor epitope (SIYRYYGL), enabling cross-reactive T cell responses that suppress tumor growth174. Moreover, bacterial-specific peptides have been identified in HLA molecules from glioblastoma tissues and tumor cell lines, suggesting a direct link between gut microbiota and tumor antigen presentation175.
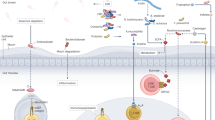
While gut microbiota enhance immunotherapy, they also play a dual role in modulating immunotherapy-related toxic responses, particularly immune-associated adverse events (irAEs), either by promoting immune homeostasis and treatment tolerance or by exacerbating adverse effects through immune dysregulation (Fig. 3). These toxicities vary by therapy type—anti-CTLA-4 therapy is often linked to colitis and pituitary inflammation, anti-PD-1 therapy to thyroid dysfunction and pneumonitis, and CAR-T therapy to cytokine release syndrome (CRS) and neurotoxicity176,177,178. A key mechanism underlying immunotherapy-related toxicity is intestinal barrier disruption. Studies show that SCFA-rich diets can significantly reduce gastrointestinal toxicity induced by immunotherapy179,180. Specific microbial taxa, such as Akkermansia muciniphila and members of the Ruminococcaceae family, have been associated with high-fiber and omega-3-rich diets, promoting both optimal antitumor immune responses and reduced irAEs by supporting mucus turnover and epithelial integrity64. Certain bacterial populations also confer resistance to irAEs. For instance, increased Bacteroidetes abundance has been linked to reduced colitis risk in anti-CTLA-4 therapy, and Bacteroides vulgatus and Bacteroides dorei were found to correlate negatively with irAE development in metastatic melanoma patients receiving anti-CTLA-4 and anti-PD-1 treatment44,181. Furthermore, FMT from healthy donors has been shown to restore microbial diversity, increase Treg proportions in colonic mucosa, and mitigate ICI-induced colitis182.

LPS, lipopolysaccharide; SCFA, short chain fatty acids; FMT, fecal microbiota transplantation. With respect to the amplification of immunotherapy toxicity, the presence of certain bacterial species, including Lactobacillus intestinalis, Streptococcus, and E. faecalis, along with other Firmicutes, disrupt the intestinal barrier, leading to elevated levels of LPS and promotion of Th17 cell activation, which in turn, promotes an increase in interleukin-17 (IL-17) secretion and inflammatory response. Conversely, microbiota exemplified by Akkermansia muciniphila, Lactobacillus, Bifidobacterium fragilis, and Bifidobacterium bifidum, as well as FMT from healthy donors, which collectively enhanced Treg activity, promoted SCFA production, and upregulated anti-inflammatory cytokines such as IL-10 and TGF-β, thereby reducing immunotherapy toxicity.
Conversely, gut microbiota imbalances can amplify immunotherapy toxicity by fostering pro-inflammatory conditions. For example, a high abundance of Bacteroides intestinalis has been associated with colitis and upregulation of mucosal IL-1β in melanoma patients treated with anti-CTLA-4 and anti-PD-1 therapy56. Additionally, studies in mice have shown that antibiotic treatment can reduce colitis, CRS, and liver injury caused by immunoagonist therapy, suggesting a microbiota-driven mechanism183. However, antibiotic use has also been linked to increased neurotoxicity in patients receiving CD19-targeted CAR-T therapy71. Interestingly, baseline gut microbiota composition may predict both treatment efficacy and toxicity. In metastatic melanoma patients treated with anti-CTLA-4, an abundance of Faecalibacterium and other Firmicutes was associated with improved treatment responses but higher irAEs, whereas Bacteroidetes correlated with poorer responses and lower irAE incidence—an effect termed the “efficacy-toxicity coupling effect”184.
These findings underscore the complex and individualized nature of microbiota-mediated immune regulation in immunotherapy. However, the mechanisms of the gut microbiota are complex and individualized, and further studies are needed to elucidate the precise pathways of the gut microbiota before translating its regulatory role into new strategies in clinical treatment.
The intratumoral microbiome: a critical regulator of TME and immunotherapy responses
The TME constitutes a sophisticated ecosystem that includes tumor cells, stromal cells, immune cells, and an intricate molecular and physical landscape. A critical yet often overlooked component of this ecosystem is the tumor-associated microbiota, consisting of bacteria, fungi, archaea, and viruses, which are either intrinsically present at the tumor site or translocated into tumors via various physiological routes152,185,186. Recent research has underscored the pivotal roles these microbial communities play in tumorigenesis, progression, metastasis, and response to cancer therapies187.
The composition and diversity of the intratumoral microbiota are variable across cancer types, reflecting distinct environmental niches and tumor-specific physiological conditions. For instance, breast cancer tissues exhibit richer microbial diversity compared to other cancers, predominantly harboring Proteobacteria and Firmicutes188. In gastrointestinal cancers, a notable enrichment of Fusobacterium nucleatum (Fn) has been documented189,190,191,192. In gastric cancer, Fn recruits tumor-associated neutrophils via IL17/NF-κB signaling, driving their polarization toward immunosuppressive phenotypes and increasing PD-L1 expression. Paradoxically, Fn-infected tumors exhibit increased sensitivity to anti-PD-L1 therapy, indicating dual roles in immune modulation189. Similarly, in CRC, Fn secretes the Fap2 protein, which interacts with TIGIT receptors on NK cells, thereby inhibiting their activity, recruiting myeloid-derived suppressor cells (MDSCs), and enhancing tumor cell proliferation while impairing antitumor immunity190. Moreover, Fusobacterium nucleatum has been found to colonize hepatic metastases via hematogenous dissemination, subsequently promoting tumor cell invasiveness through activation of the TLR4/MyD88 signaling axis191. In NSCLC, butyrate-producing bacteria, notably Roseburia, correlate with disease recurrence and metastasis. Circulating microbial DNA signatures enriched with butyrate producers have also emerged as predictive biomarkers for recurrence, underscoring the clinical relevance of microbial metabolites192.
Microbial influence on tumor progression also extends to other cancer types. In squamous cell carcinoma, Staphylococcus species release enterotoxin B, which enhances the expression of CD4 + STAT5 and HDAC-1 in T cells, subsequently increasing IL-9 secretion and promoting apoptosis of carcinoma cells193. Additionally, microbial metabolites significantly impact tumor development through mechanisms such as induction of DNA damage and activation of oncogenic pathways194. Escherichia coli, for example, produces colibactin, leading to DNA methylation changes and double-strand DNA breaks. Similarly, Bacteroides fragilis generates reactive oxygen species (ROS), thereby inducing genomic instability and facilitating tumorigenesis195,196.
The intratumor microbiota is not only affected by the dynamics of the TME, but also acts as a feedback regulator of its morphology and function. Hypoxia, low pH, and metabolic stress in the TME may be more favorable for the growth of both parthenogenetic and purely anaerobic bacteria197. Bifidobacterium, for example, migrates to colonize and enrich CRC tumors198. The microbiota, in turn, affects the TME through metabolic and immune interactions. Numerous studies have demonstrated that specific gut microbiota and their metabolites influence the TME by promoting the formation and maturation of tertiary lymphoid structures (TLS). TLS serve as critical hubs for local immune activation, facilitating the recruitment, organization, and activation of immune cells, notably antigen-presenting cells, T cells, and B cells, thus coordinating robust anti-tumor immunity. For example, in CRC mouse models, the gut commensal bacterium H. hepaticus has been shown to drive TLS formation by inducing CD4+ T cell differentiation into follicular helper T (Tfh) cells, thereby enhancing immune cell infiltration within the TME199. Additionally, microbial metabolites such as SCFAs and trimethylamine N-oxide can reshape the TME to support TLS maturation through activation of the STING signaling pathway and enhancement of T and NK cell activity200. Furthermore, other microbial-derived metabolites—including indole derivatives, hydrogen sulfide, and bile acids—modulate tumor cell metabolism and immune homeostasis201. The dynamic evolution of the gut microbiome is intimately associated with mechanisms of immune escape in tumors. Specific Clostridium spp. produce metabolites that induce the differentiation of Tregs while inhibiting the establishment of immunosuppression in the TME202. Moreover, D-Lactate produced by Lactobacillus is capable of converting M2-type TAMs to M1-type, thereby remodeling the TME and enhancing anti-tumor immune responses203.
Given these insights, targeting tumor-associated microbiota and their metabolites emerges as a promising therapeutic strategy. Synthetic microbiome interventions, inhibitors against specific tumor-associated bacteria, or microbiome-based metabolite modulation could effectively disrupt oncogenic microbial interactions, thereby enhancing immune responsiveness and treatment efficacy. Future research should thus prioritize comprehensive characterization of the intratumoral microbiome across diverse cancer types, unraveling the precise molecular mechanisms underlying microbial-tumor-immune interactions. This approach holds significant potential for optimizing personalized cancer immunotherapy strategies, ultimately improving patient outcomes across multiple malignancies.
Strategies for gut microbiota intervention combined with immunotherapy
The composition and diversity of the gut microbiota are closely linked to the efficacy of cancer therapies. Beyond merely understanding these associations, researchers are now exploring ways to modulate the microbiota to enhance immunotherapy effectiveness and reduce toxicity. Strategies for microbiota-targeted interventions combined with immunotherapy include antibiotics, FMT, probiotics, dietary and prebiotic-based approaches, and engineered microbiota therapies (Fig. 4). A multitude of pertinent clinical trials are currently ongoing, and a comprehensive review of studies pertaining to these therapeutic strategies from recent years is provided below.

FMT, fecal microbiota transplantation. Gut microbiota modulation through different approaches, including antibiotics, FMT, probiotics, diet and prebiotics, and other strategies, has been shown to influence immunotherapy outcomes. Selective antibiotics, such as vancomycin and tigecycline, can be beneficial, whereas broad-spectrum antibiotics may negatively impact treatment response. FMT, administered via oral capsules, colonoscopy, or gastroscopy, facilitates the transfer of beneficial gut microbiota from immunotherapy-responsive donors to patients. The use of probiotics, including Lactobacillus, Bifidobacterium, Clostridium butyricum, and Akkermansia, supports a favorable gut microbial composition. Diet and prebiotics, such as inulin gel, pectin, high-fiber diets, fasting-mimicking diets, and ketogenic diets, contribute to microbiota modulation and immune system support. Additionally, other strategies, including engineered microorganisms, microbial peptides, and microbial metabolites, represent emerging therapeutic approaches. These interventions collectively contribute to optimizing gut microbiota composition, thereby promoting immune responses and enhancing immunotherapy efficacy.
Antibiotics
The utilization of antibiotics in the management of patients with malignant tumors is predominantly driven by postoperative prophylactic infections, treatment-related infections, and the management of comorbidities. However, these interventions inevitably alter the gut microbiota, potentially affecting immunotherapy outcomes204,205,206. Preclinical and clinical studies suggest that antibiotic use, particularly broad-spectrum antibiotics, may impair ICI efficacy (Table 2)95,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228. In mouse models of NSCLC and melanoma, antibiotics such as vancomycin, ampicillin, metronidazole, and neomycin disrupted IL-17-producing γδT17 cells, promoting tumor metastasis229. Clinical studies have similarly reported reduced progression-free survival (PFS) and overall survival (OS) in patients with advanced RCC and NSCLC who received antibiotics prior to ICI therapy230. The results of a retrospective analysis and two clinical studies related to ICI for NSCLC showed that both antibiotics and another microbiologically modifiable proton pump inhibitor significantly shortened PFS and OS in patients treated with atalizumab231. These findings suggest a pivotal role for the antibiotic-influenced gut microbiota in ICI treatment.
The impact of antibiotics on tumor immunotherapy is also related to the duration of patient exposure to antibiotics. A multicenter prospective study reported that antibiotic treatment given prior to, but not at the time of, ICI treatment were associated with poorer response and lower OS118. Conversely, another study suggested that antibiotic use within 30 days before or after ICI initiation correlated with improved benefit from immunotherapy213. The divergent outcomes observed in the two studies may be attributable to factors such as the timing of antibiotic administration, the specific nature of the tumor, and individual variations. A meta-analysis further highlighted the impact of antibiotic timing, showing that patients who avoided antibiotics within 42 days before ICI therapy had significantly longer OS than those who received antibiotics within 60 days prior to treatment, which is consistent with previous findings, as the microbiota returned to near-baseline levels within 42 days after administration of an antibiotic cocktail regimen (ABX) for 4 days to healthy adults208,232. The aforementioned studies underscore the pivotal role of the temporal aspect of antibiotic administration in the context of immunotherapy. Additionally, broad-spectrum antibiotics appear to have a more detrimental impact on OS compared to narrow-spectrum antibiotics226,233.
Despite concerns about their negative impact on immunotherapy, certain antibiotics may enhance treatment efficacy when used selectively. Vancomycin, which targets gram-positive bacteria such as Clostridium difficile, has been shown to promote NKT cell migration and IFN-γ production, thereby inhibiting primary and metastatic HCC growth in mice109,234. A clinical trial (NCT03785210) is currently evaluating the efficacy of vancomycin in combination with ICIs for HCC treatment. Antibiotics may also counteract tumor-associated microbial activities that impair therapy. For example, some Clostridium species synthesize androgens, reducing the effectiveness of androgen deprivation therapy (ADT) in prostate cancer235. Vancomycin-containing antibiotic regimens may enhance ADT efficacy by eliminating these bacteria236. Similarly, tigecycline has demonstrated synergistic effects with B-cell lymphoma-2 inhibitors, suggesting potential therapeutic applications in diffuse large B-cell lymphoma237. Beyond systemic effects, localized antibiotic interventions may be particularly beneficial in tumors with a high microbial burden. The pancreas, due to its anatomical connection with the gut, harbors a substantial microbial load linked to pancreatic cancer progression and immunosuppression238,239. ABX has been shown to disrupt these microbial communities, inhibits pancreatic ductal adenocarcinoma invasion, and promotes ICI outcome by increasing the differentiation of M1 macrophages and Th1 CD4 + T cells, activating CD8 + T cells, and upregulating the expression of PD-1 on effector T cells240. Additionally, ABX can reduce bacterial translocation to the liver and intestine, thereby inhibiting CRC metastasis241.
While antibiotics offer potential benefits in modulating tumor-associated microbiota, their application in cancer treatment must be carefully tailored. Factors such as the patient’s immune status, tumor genetics, TME, and antibiotic resistance should be considered. Therefore, treatment regimens should be tailored to the specific condition of the patient and can be based on pre-treatment sequencing of the gut microbiome and targeted formulation design approaches to enhance treatment efficacy while reducing disruption of the gut microbiota by broad-spectrum antibiotics.
FMT
FMT refers to the reconstitution of a new intestinal flora by transplanting functional flora from healthy human feces into a patient’s gastrointestinal tract, either orally or via colonoscopy or gastroscopy. Originally developed to treat refractory Clostridium difficile infections, FMT has since been explored for other gastrointestinal disorders such as Crohn’s disease and ulcerative colitis242,243,244. Its potential in oncology has emerged from preclinical studies demonstrating that germ-free mice receiving fecal transplants from immunotherapy-responsive patients exhibit stronger anti-tumor immune responses compared to those receiving transplants from non-responders46,94.
Currently, clinical trials investigating FMT in cancer immunotherapy are in early stages (Table 3)244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275. Two independent trials (NCT03353402 and NCT03341143) first demonstrated that FMT could overcome resistance to ICI therapy in metastatic melanoma26,55. In NCT03353402, 10 patients with PD-1–refractory melanoma underwent FMT followed by renewed anti-PD-1 therapy, resulting in tumor reduction in three patients, including two partial responses (PRs) and one complete response (CR)26. Similarly, NCT03341143 assessed FMT combined with pembrolizumab in 15 patients, yielding two PRs, one overall response (OR), and three cases of stable disease (SD) lasting over a year55. These studies also highlighted favorable immune changes, including increased CD8 + T-cell activation and reduced IL-8–expressing myeloid cells. Microbiome analysis revealed enrichment of beneficial bacteria (Bifidobacterium longum, Lachnospiraceae, Ruminococcaceae) and a decline in potentially detrimental species such as Bacteroides. Additionally, microbial diversity changes correlated with increased levels of primary and secondary bile acids and benzoic acid metabolites. A subsequent FMT study (NCT03772899) differed from the two previous studies. The present study was distinctive in that its FMT donor was from a healthy individual. Although its conclusions are subject to further validation due to the trial’s small sample size and lack of a control group, the preliminary results suggest that the use of a healthy donor is also a safe alternative. Concomitantly, this prompts further inquiry into the optimal FMT donor: a cancer survivor with heightened sensitivity to immunotherapy, or a non-malignant individual devoid of a medical history74.
Beyond tumor response, FMT is also being explored for managing irAEs, particularly in reducing steroid-refractory colitis. Trials such as NCT03819296, NCT04038619, NCT04721041, and NCT04163289 are assessing its potential to mitigate irAEs. Early studies suggest FMT is effective in treating steroid-resistant gastrointestinal graft-versus-host disease (GvHD) following hematopoietic stem cell transplantation (HSCT), with patients showing significant clinical remission276,277. Similarly, in ICI-induced colitis, case reports and small trials indicate that FMT can restore gut homeostasis, with notable increases in beneficial microbes such as Akkermansia, Blautia, and Bifidobacterium182. A study by Halsey et al. reported that 92% of 12 patients with refractory immune-mediated colitis achieved clinical remission post-FMT, accompanied by increased gut microbial diversity and enrichment of Collinsella and Bifidobacterium278. These findings, supported by numerous ongoing and completed trials, reinforce the therapeutic potential of FMT in malignancies279,280,281,282. Despite these promising outcomes, concerns remain regarding the long-term safety of FMT. In 2019, two patients developed Escherichia coli bacteremia, with one fatality, following FMT from the same donor283. This finding prompted the Food and Drug Administration (FDA) to issue a safety warning regarding the risk of infection with FMT therapy. A retrospective study further revealed that 9% of screened donors carried multidrug-resistant organisms, highlighting the need for stringent donor screening protocols, particularly for immunocompromised patients284.
However, not all PD-1–refractory patients respond to renewed therapy after FMT, likely due to factors such as immunosuppression, donor-recipient microbiota incompatibility, or suboptimal FMT delivery methods. Additionally, there are many off-target microbiota components in donor feces that are transplanted into recipients that have unknown effects. Future research should focus on refining microbiota transfer protocols, identifying optimal donor-recipient pairings, and integrating FMT with complementary approaches to enhance antitumor efficacy.
Probiotics
The concept of probiotics, first introduced by Metchnikoff, refers to live microorganisms that confer health benefits when administered in sufficient quantities285,286. Preclinical studies indicate that supplementing specific gut microbes, such as Lactobacillus, Bifidobacterium, Enterococcus, Faecalibacterium, and Ruminococcaceae, can enhance tumor immunotherapy287,288,289,290,291. For instance, Bifidobacterium pseudolongum produces inosine, which activates the T cell adenosine A2A signaling pathway, improving immunotherapeutic efficacy176. Similarly, Bifidobacterium bifidum K57 supplementation has been shown to boost the immune response against the TME by enhancing CD4+ and CD8 + T cell activation and increasing IFN-γ and IL-2 secretion, thereby synergizing with anti-PD-1 therapy to reduce tumor burden290.
Advancements in NGS have identified Akkermansia as another probiotic that promotes immunotherapy292,293. In Peyer’s patches, Akkermansia induces an increase in Tfh cells, while its outer membrane protein, Amuc, triggers adaptive immune responses by activating the TLR2 signaling pathway and decreasing immunosuppressive Treg cell levels294,295. In addition, commensal bacterial strains contribute to immunotherapy by maintaining gut homeostasis. Tanoue et al. isolated 11 bacterial strains from healthy donors that induced IFN-γ production by CD8 + T cells, thereby enhancing ICI efficacy in murine tumor models296. While some Clostridium species are linked to colon carcinogenesis, others, such as Roseburia intestinalis, Eubacterium hallii, Faecalibacterium prausnitzii, and Anaerostipes caccae, have demonstrated tumor-suppressive properties by activating and infiltrating intratumoral CD8+ T cells, effectively reducing tumor growth297. These studies suggest that the interaction between the gut microbiota and the tumor and immune system may involve more complex processes.
Early clinical trials have explored probiotics to modulate the gut microbiota and enhance antitumor immunity in cancer patients. Probiotic strains with anti-inflammatory and mucosal-protective properties, such as Lactobacillus and Bifidobacterium, have been used to mitigate chemotherapy- and radiotherapy-induced toxicity298,299,300. Consoli et al. reported that probiotic treatment significantly reduced IL-1β, IL-10, and IL-23A mRNA levels in colonic mucosa, leading to a lower incidence of postoperative infections301. A clinical trial in CRC patients found that supplementation with Bifidobacterium lactis and Lactobacillus increased butyrate-producing bacteria, particularly Faecalibacterium and Clostridiales spp., in tumor, mucosal, and fecal microbiota, further supporting the role of probiotics in enhancing antitumor responses302. Several ongoing clinical trials are evaluating the safety and efficacy of probiotics in combination with ICI therapy (Table 4)303,304,305,306,307,308,309,310,311,312,313,314,315,316,317. MRx0518, a flagellin-producing probiotic, activates TLR5 and NF-κB signaling pathways to exert antitumor effects318. A phase 1/2 trial combining MRx0518 with pembrolizumab met its primary efficacy endpoint in RCC patients, though the pharmaceutical company later withdrew from development due to financial constraints319. Retrospective analyses have shown that Clostridium butyricum CBM588 significantly PFS and OS in NSCLC patients receiving anti-PD-1 therapy, even among those who also received antibiotics320. In a phase I clinical trial (NCT03829111), CBM588 significantly extended PFS in metastatic RCC patients, with a response rate of 58% when combined with an ICI, compared to 20% with ICI alone27. Additionally, CBM588 improved ICI efficacy in NSCLC patients treated with proton pump inhibitors (PPIs)321. Another promising probiotic, Bifidobacterium bifidum EDP1503, enhances the CD8 + T cell-to-Treg cell ratio, and preliminary results from a phase I clinical trial (NCT03775850) suggest it is safe and well tolerated in combination with pembrolizumab322.
Despite their demonstrated benefits in non-neoplastic diseases, probiotic-based cancer therapies remain underexplored, with only a limited number of clinical trials available for validation323,324. The heterogeneity of commercial probiotics and the lack of standardized quality control raise concerns about their integration with immunotherapy. Indeed, studies have demonstrated that patients who consume sufficient dietary fiber and do not utilize probiotics tend to exhibit optimal efficacy with immunotherapy28. Moreover, recent studies suggest that specific gut microbes may enhance anti-tumor immune responses by influencing TLS within the TME, which are closely linked to patient responses to ICI therapy200,325,326. These emerging insights highlight the potential for developing targeted microbiota-based interventions to optimize cancer immunotherapy.
Diet and prebiotics
Beyond direct modulation of the gut microbiota, dietary interventions can significantly influence microbial composition, bacterial metabolites, and immune responses327,328,329. Preclinical studies suggest that dietary strategies such as caloric restriction, intermittent fasting, high-fiber diets, ketogenic diets, and targeted micronutrient supplementation can enhance immunotherapy by reshaping gut microbiota composition330,331,332,333,334,335. For example, dietary restriction in mice promotes the accumulation of memory T cells in the bone marrow, strengthening immune responses against infections and tumors336. Consistent with the previous study, during chronic or intermittent fiber deficiency, the gut microbiota utilizes host-secreted mucus glycoproteins as a nutrient source, leading to a disruption of the colonic mucosal barrier and inducing a decrease in Bifidobacterium abundance and an increase in Mucococcus abundance28,337. Consequently, a poor response to anti-PD-1 therapy was observed in mice fed a low-fiber diet28. A ketogenic diet and its primary metabolite, 3-hydroxybutyrate, have been shown to enhance anti-PD-1 therapy by preventing PD-L1 upregulation in bone marrow cells and promoting the expansion of CXCR3+ T cells328. In addition, a serine/glycine-free diet has been shown to slow CRC growth while promoting the accumulation of cytotoxic T cells to enhance anti-tumor immunity. And blocking the PD-1/PD-L1 pathway further enhances the effects of this dietary approach, which highlight the potential of combining the serine/glycine-free diet with immunotherapy338.
Clinical studies further support the role of diet in immunotherapy outcomes. An analysis by Wastyk et al. found that high-fiber and fermented food diets influenced gut microbiota and immune function in distinct ways: high-fiber diets increased carbohydrate-active enzymes involved in glycan metabolism with moderate effects on microbial diversity, while fermented diets enhanced microbial diversity and reduced inflammatory markers339. A prospective study of melanoma patients undergoing ICI therapy across multiple cohorts found that higher dietary fiber intake correlated with better clinical outcomes, particularly in patients with greater bacterial diversity65. Additionally, dietary fiber produces SCFAs, such as butyrate, which can directly activate tumor-fighting CD8+ T cells and enhance anti-PD-1 therapy109,339,340. However, conflicting evidence suggests that elevated SCFA levels in peripheral blood may limit anti-CTLA-4 efficacy, indicating that the role of metabolites in antitumor immunity may vary across tissues341. Therefore, precision in dietary interventions is crucial for optimizing their therapeutic potential.
Prebiotics, a more targeted dietary approach, are chemically defined non-digestible fibers—primarily oligosaccharides and polysaccharides such as inulin and pectin—that selectively promote the growth of beneficial gut microbes342,343,344. Prebiotics improve epithelial integrity, enhance mucosal barrier function, and activate innate immune cells, contributing to improved immunotherapy outcomes345,346,347,348. Han et al. demonstrated that an oral inulin gel, designed to release in the colon, effectively modulated gut microbiota, promoted SCFA metabolism, induced systemic memory T cell responses, and strengthened anti-PD-1 therapy349. Meanwhile, pectin has been shown to enhance ICI efficacy through multiple mechanisms and it improves anti-PD-1 responses in colorectal cancer by increasing butyrate-producing gut flora and modulating T cell infiltration into the TME350. Additionally, pectin has been linked to increased abundance of mucophilic bacilli, which produce cyclic AMP and activate STING signaling, resulting in type I interferon-mediated immunity229.
Other prebiotics, such as polysaccharides derived from medicinal plants, have also shown promise. Ginseng polysaccharides have been found to enhance anti-PD-1 therapy by remodeling gut metabolite composition, altering tryptophan metabolism, and suppressing Treg cell responses351. Similarly, polyphenol-rich Kamagra supplementation in mice increased gut microbial diversity and boosted beneficial bacteria such as Ruminococcaceae, Oscillospiraceae, and Akkermansia muciniphila352. These changes improved the CD8+ T/Treg cell ratio in the TME and reversed ICI resistance. Unlike whole-food dietary interventions, these extracted or modified prebiotics allow for more precise modulation of gut microbiota and offer potential for targeted immunotherapy enhancements. Building on these findings, a number of clinical trials of dietary or prebiotic interventions for tumor immunotherapy are underway (Table 5)353,354,355,356,357,358,359,360,361,362,363,364,365,366,367. We look forward to the results of these clinical trials.
Other strategies such as engineered microorganisms
Advancements in synthetic biology have enabled the genetic modification of bacteria to selectively target tumors. Engineered strains such as Salmonella, Escherichia coli, and Bifidobacterium can colonize the TME, exert immunogenic effects, and locally release therapeutic agents to enhance immune-mediated tumor clearance368,369,370,371. For instance, L-arginine plays a crucial role in T cell metabolism, supporting their survival and anti-tumor activity372. The engineered probiotic Escherichia coli Nissle 1917 has been designed to colonize tumors and convert ammonia, a metabolic byproduct, into L-arginine, thereby promoting CD4 + T and CD8 + T cell infiltration and enhancing the efficacy of anti-PD-L1 therapy373. Another engineered strain, SYNB1891, derived from E. coli Nissle, stimulates STING activation in antigen-presenting cells (APCs), triggering innate immune pathways that contribute to anti-tumor immunity374. Although a clinical trial (NCT04167137) evaluating SYNB1891 in combination with atezolizumab for advanced solid tumors was paused due to funding constraints, its potential remains promising. Chowdhury et al. engineered a non-pathogenic E. coli strain to target lysis and release anti-CD47 nanoantibodies in the tumor microenvironment.CD47 is overexpressed in a variety of human cancers, and inhibition of its function promotes tumor phagocytosis by immune cells. The engineered bacteria showed significant anti-tumor effects in a mouse model, promoting the activation of tumor-infiltrating T cells, stimulating rapid tumor regression and inhibiting metastasis, thereby prolonging the survival of the mice375.
Recent studies have further expanded the use of engineered microorganisms in tumor immunotherapy. Engineered E. coli Nissle 1917 strains have been developed to deliver tumor-specific neoantigens, and stimulate a robust anti-tumor immune response by activating DCs, NK cells, and tumor-infiltrating T cells. This targeted approach enhances immunotherapy specificity while minimizing toxicity to normal tissues376. Other strategies include using engineered E. coli to deliver an IL-18 mutant, which significantly boosts CD8 + T cell and NK cell activity, suppresses tumor growth, and enhances the tumor-targeting abilities of CAR-NK cells377. Several engineered bacterial strains, such as ADXS11-001, Ty21a, JNJ-64041809, and VXM01, have also demonstrated potential in immunotherapy by leveraging tumor-specific antigen presentation378,379,380,381,382,383. In addition, Chen et al. genetically engineered the skin-dwelling bacterium Staphylococcus epidermidis to express tumor antigens, thereby eliciting T-cell responses against melanoma. Following colonization, these engineered bacteria induce tumor-specific T cells that mature, enter systemic circulation, and infiltrate both local and metastatic tumor sites which exert cytotoxic effects and suppress tumor growth384. Notably, recent studies have identified that microbiota, such as Bifidobacterium and Lactobacillus derive vesicles—such as outer membrane vesicles (OMVs) and extracellular vesicles (EVs)—that have the ability to deliver immunomodulatory molecules, antigens, or adjuvants to immune cells385,386,387. For example, Lactobacillus-derived EVs can inhibit colon tumor formation by modulating the SIRT5/p53 axis and thereby inhibiting colon tumor formation. These studies further expand the scope of microbiota-based oncology interventions.
Beyond engineered bacteria, other microbiota-based approaches are being explored to improve immunotherapy. Phage therapy, particularly CRISPR-Cas3 engineered phages, can modulate the gut microbiota, immune system, and TME388,389,390,391,392. Microbial metabolites, such as valproic acid (VPA), function as histone deacetylase (HDAC) inhibitors, influencing immune signaling pathways and promoting tumor cell apoptosis393. Clinical trials assessing VPA in combination with immune checkpoint inhibitors (NCT02446431, NCT02624128, NCT01106872) are underway. Small-molecule peptides derived from the microbiota also hold promise394. EO2401, a microbial peptide that mimics tumor-associated antigens (TSAs), has been shown to activate memory T cells and elicit a strong immune response395. In the ROSALIE Phase 1/2 trial, EO2401 combined with nivolumab and bevacizumab in glioblastoma patients led to a median survival of 14.5 months and an 18-month survival rate of 43.1%, with favorable tolerability396.
Furthermore, the field is moving towards the development of synthetic consortia with defined immunomodulatory properties and the implementation of longitudinal studies linking microbiota dynamics to the durability of therapeutic responses. By tracking temporal changes in the microbiome and immune parameters, researchers can identify predictive biomarkers of long-term response and resistance397. Meanwhile, advances in synthetic biology enable the rational design of microbial communities or synthetic constructs that integrate multiple immunoregulatory mechanisms, thus providing a multi-faceted and precise approach to cancer immunotherapy. These studies underscore the potential of microbiota-based interventions in cancer treatment. Engineered microorganisms not only provide precise modulation of the TME but may also offer systemic protection against metastasis. While challenges remain, continued research and technological advancements are expected to position microbiota-based therapies as a key component of future cancer treatment strategies.
Challenges and future directions
Understanding the relationship between gut microbiota and immunotherapy remains complex. The microbiota influences immune responses through various mechanisms such as metabolite production and inflammation regulation, but these interactions are bidirectional—some microbial communities enhance immunity, while others suppress it. Moreover, variations in TMEs further complicate microbiota-driven immune modulation398. Many current findings are derived from animal models, yet differences in microbial composition and immune function between mice and humans limit their clinical relevance399,400. The absence of standardized research methodologies also contributes to inconsistent data, making it difficult to draw definitive conclusions. Critically, this field faces additional limitations. First, technical variability in microbiome sequencing methods—such as differences in 16S rRNA gene regions targeted, DNA extraction protocols, and sequencing platforms—can result in substantial discrepancies between studies401,402. Second, inter-trial inconsistencies, including patient selection, sample handling, and data processing, further undermine reproducibility403. FMT also suffers from significant ‘donor effects,’ where outcomes depend strongly on the specific microbiota composition of the donor, complicating interpretation and clinical standardization404. Moreover, host genetics can profoundly shape microbial composition and immune responses, yet is often underappreciated in current research designs405,406. Addressing these methodological and biological confounders is essential for the field’s progress.
Despite its promise, microbiota modulation faces significant clinical hurdles. Many studies have small sample sizes, limiting statistical power and generalizability. Standardization is another major challenge, as microbiota composition varies widely among individuals due to factors such as diet, medication use, and environmental exposure. Additionally, the lack of uniform protocols for microbiome analysis affects reproducibility. Safety concerns further complicate clinical translation—approaches like FMT may not reliably restore beneficial bacteria, and unintended microbial shifts could lead to adverse effects407. Addressing these issues will be critical for integrating microbiota-based strategies into clinical practice.
Advancements in genomics, metabolomics, and microbiomics are driving more precise microbiota research408. Non-invasive techniques, such as microbial DNA analysis from blood samples, hold potential for early cancer detection and treatment monitoring409. Emerging tools, including artificial gut models and ingestible biosensors, will enhance real-time microbiota analysis410,411. Multi-omics approaches combining microbiome, immune, and metabolic data may identify key microbial factors influencing immunotherapy responses, paving the way for targeted interventions. Notably, recent studies have revealed a significant influence of the gut microbiota on vaccine efficacy, particularly highlighting the roles of Bifidobacterium and Bacteroides412. These bacteria can interact with DCs, which are pivotal for initiating anti-tumor immune responses. Modulating the gut microbiota through dietary interventions or targeted microbial supplementation may enhance the host’s ability to recognize and respond to tumor antigens introduced by vaccines. For example, Jing et al. demonstrated that the synergistic administration of Lactobacillus rhamnosus GG and jujube powder markedly improved the efficacy of whole-cell cancer vaccines in the MC38 murine cancer model, primarily by increasing the proportion of CD8 + IFNγ + T cells413. Moreover, vaccine efficacy has been found to correlate with gut microbiota composition414. Therefore, integrating gut microbiota modulation with cancer vaccination represents a promising therapeutic strategy. As research progresses, personalized cancer immunotherapy may become a reality. Patients with an imbalanced microbiome might benefit from interventions, such as strain-specific probiotics, engineered bacterial therapies, or dietary modifications, while those with a favorable microbiota profile associated with positive immune responses may require only minor adjustments to optimize treatment outcomes. Beyond the gut, microbial communities in the oral cavity, skin, and TME may also influence immune responses, broadening the scope of personalized therapeutic strategies415,416,417.
Achieving clinical translation will require interdisciplinary collaboration among immunologists, microbiologists, oncologists, and data scientists. Large-scale, multi-center studies and standardized data collection methods are essential for generating clinically relevant insights. Artificial intelligence (AI) and machine learning can improve patient stratification and help identify microbiota-driven mechanisms that influence treatment outcomes. By integrating expertise across multiple fields, microbiota research can play a central role in advancing precision medicine for cancer treatment.
Conclusion
In summary, the gut microbiota plays a pivotal role in modulating cancer immunotherapy, influencing treatment efficacy, patient response, and therapy-associated toxicity. Recent advances in sequencing technologies and microbiota-targeted interventions, such as FMT, probiotics, and dietary modifications, have opened new avenues for enhancing immunotherapy outcomes. However, significant challenges remain in translating these findings into clinical practice, primarily due to inter-individual variability and the complexity of host-microbiota interactions. Future research should focus on integrating multi-omics approaches, AI, and precision medicine to establish causal relationships between microbiota composition and immunotherapy response. Additionally, exploring the mechanisms by which microbial metabolites influence tumor immune microenvironments will be crucial for developing novel therapeutic strategies. By harnessing the potential of the gut microbiota, clinicians may be able to personalize immunotherapy regimens, improve treatment efficacy, and overcome drug resistance. Ultimately, continued interdisciplinary research will be essential to fully unlock the clinical potential of microbiota-targeted interventions, paving the way for more effective and individualized cancer treatments.
Data availability
No datasets were generated or analysed during the current study.
References
Lederberg, J. & McCray, A. T. ‘Ome Sweet’ Omics—a genealogical treasury of words. Scientist 15, 8–8 (2001).
Sender, R., Fuchs, S. & Milo, R. Are we really vastly outnumbered? revisiting the ratio of bacterial to host cells in humans. Cell 164, 337–340 (2016).
Gilbert, J. A. et al. Current understanding of the human microbiome. Nat. Med. 24, 392–400 (2018).
Martinez-Guryn, K., Leone, V. & Chang, E. B. Regional diversity of the gastrointestinal microbiome. Cell Host Microbe 26, 314–324 (2019).
Sender, R., Fuchs, S. & Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 14, e1002533 (2016).
Maynard, C. L., Elson, C. O. & Hatton, R. D. Reciprocal interactions of the intestinal microbiota and immune system. Nature 489, 231–241 (2012).
Belkaid, Y. & Harrison, O. J. Homeostatic Immunity and the Microbiota. Immunity 46, 562–576 (2017).
Jugder, B. E., Kamareddine, L. & Watnick, P. I. Microbiota-derived acetate activates intestinal innate immunity via the Tip60 histone acetyltransferase complex. Immunity 54, 1683–1697.e3 (2021).
Zheng, D., Liwinski, T. & Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 30, 492–506 (2020).
Lin, K., Zhu, L. & Yang, L. Gut and obesity/metabolic disease: Focus on microbiota metabolites. MedComm 3, e171 (2022).
Oliphant, K. & Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: major fermentation by-products and their impact on host health. Microbiome 7, 91 (2019).
Chen, Y., Wang, Z., Chen, X., Chao, G. & Zhang, S. The role of gut microbiota and metabolites in regulating the immune response in drug-induced enteritis. J. Appl. Microbiol. 134, lxad032 (2023).
Visekruna, A. & Luu, M. The role of short-chain fatty acids and bile acids in intestinal and liver function, inflammation, and carcinogenesis. Front. Cell Dev. Biol. 9, 703218 (2021).
Ansaldo, E., Farley, T. K. & Belkaid, Y. Control of immunity by the microbiota. Annu. Rev. Immunol. 39, 449–479 (2021).
Dzutsev, A. et al. Microbes and cancer. Annu. Rev. Immunol. 35, 199–228 (2017).
Zhang, Y. & Zhang, Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol. Immunol. 17, 807–821 (2020).
Sharma, P., Hu-Lieskovan, S., Wargo, J. A. & Ribas, A. Primary, adaptive, and acquired resistance to cancer immunotherapy. Cell 168, 707–723 (2017).
Pan, C. et al. Next-generation immuno-oncology agents: current momentum shifts in cancer immunotherapy. J. Hematol. Oncol. 13, 29 (2020).
Wang, M. et al. Clinical observation of the efficacy of PD-1/PD-L1 inhibitors in the treatment of patients with advanced solid tumors. Immun. Inflamm. Dis. 9, 1584–1595 (2021).
Hegde, P. S. & Chen, D. S. Top 10 challenges in cancer immunotherapy. Immunity 52, 17–35 (2020).
Cullin, N., Antunes, C. A., Straussman, R., Stein-Thoeringer, C. K. & Elinav, E. Microbiome and cancer. Cancer Cell 39, 1317–1341 (2021).
Fehervari, Z. Microbiota shape tumor immunity. Nat. Immunol. 22, 1469 (2021).
Consortium I. T. P.-C. A. O. W. G. Pan-cancer analysis of whole genomes. Nature 578, 82–93 (2020).
Yonekura, S. et al. Cancer Induces a Stress Ileopathy Depending on β-Adrenergic Receptors and Promoting Dysbiosis that Contributes to Carcinogenesis. Cancer Discov. 12, 1128–1151 (2022).
Routy, B. et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359, 91–97 (2018).
Baruch, E. N. et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science 371, 602–609 (2021).
Dizman, N. et al. Nivolumab plus ipilimumab with or without live bacterial supplementation in metastatic renal cell carcinoma: a randomized phase 1 trial. Nat. Med. 28, 704–712 (2022).
Spencer, C. N. et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 374, 1632–1640 (2021).
Liu, Q. et al. Role of the gut microbiota in tumorigenesis and treatment. Theranostics 14, 2304–2328 (2024).
Blake, S. J., Wolf, Y., Boursi, B. & Lynn, D. J. Role of the microbiota in response to and recovery from cancer therapy. Nat. Rev. Immunol. 24, 308–325 (2024).
Wang, M., Yu, F. & Li, P. Intratumor microbiota in cancer pathogenesis and immunity: from mechanisms of action to therapeutic opportunities. Front. Immunol. 14, 1269054 (2023).
Ji, H. et al. Intratumoural microbiota: from theory to clinical application. Cell Commun. Signal 21, 164 (2023).
Cao, Y. et al. Intratumoural microbiota: a new frontier in cancer development and therapy. Signal Transduct. Target Ther. 9, 15 (2024).
Schmid, P. et al. Atezolizumab plus nab-paclitaxel as first-line treatment for unresectable, locally advanced or metastatic triple-negative breast cancer (IMpassion130): updated efficacy results from a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 21, 44–59 (2020).
He, K., et al. Safety and clinical activity of target-preserving anti-CTLA-4 antibody ONC-392 as monotherapy in NSCLC patients who progressed on PD(L)1-targeted immunotherapy. J. Clin. Oncol. 41, 9024–9024 (2023).
Zhang, P., Zhang, G. & Wan, X. Challenges and new technologies in adoptive cell therapy. J. Hematol. Oncol. 16, 97 (2023).
Ventola, C. L. Cancer immunotherapy, Part 3: challenges and future trends. P T 42, 514–521 (2017).
Zaretsky, J. M. et al. Mutations associated with acquired resistance to PD-1 blockade in melanoma. N. Engl. J. Med. 375, 819–829 (2016).
Sade-Feldman, M. et al. Resistance to checkpoint blockade therapy through inactivation of antigen presentation. Nat. Commun. 8, 1136–1136 (2017).
Majzner, R. G. & Mackall, C. L. Tumor antigen escape from CAR T-cell therapy. Cancer Discov. 8, 1219–1226 (2018).
Starzer, A. M., Preusser, M. & Berghoff, A. S. Immune escape mechanisms and therapeutic approaches in cancer: the cancer-immunity cycle. Ther. Adv. Med Oncol. 14, 17588359221096219 (2022).
Cancer Genome Atlas Research Network Comprehensive molecular profiling of lung adenocarcinoma. Nature 511, 543–550 (2014).
Chalmers, Z. R. et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med. 9, 34 (2017).
Dubin, K. et al. Intestinal microbiome analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat. Commun. 7, 10391 (2016).
Gopalakrishnan, V. et al. Gut microbiome modulates response to anti- PD-1 immunotherapy in melanoma patients. Science 359, 97–103 (2018).
Matson, V. et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 359, 104–108 (2018).
Peters, B. A. et al. Relating the gut metagenome and metatranscriptome to immunotherapy responses in melanoma patients. Genome Med. 11, 61 (2019).
Katayama, Y. et al. The role of the gut microbiome on the efficacy of immune checkpoint inhibitors in Japanese responder patients with advanced non-small cell lung cancer. Transl. Lung Cancer Res. 8, 847–853 (2019).
Jin, Y. et al. The diversity of gut microbiome is associated with favorable responses to anti-programmed death 1 immunotherapy in Chinese patients with NSCLC. J. Thorac. Oncol. 14, 1378–1389 (2019).
Zheng, Y. et al. Gut microbiome affects the response to anti-PD-1 immunotherapy in patients with hepatocellular carcinoma. J. Immunother. Cancer 7, 193 (2019).
Limeta, A. et al. Meta-analysis of the gut microbiota in predicting response to cancer immunotherapy in metastatic melanoma. JCI Insight 5, e140940 (2020).
Vernocchi, P. et al. Network analysis of gut microbiome and metabolome to discover microbiota-linked biomarkers in patients affected by non-small cell lung cancer. Int J. Mol. Sci. 21, 8730 (2020).
Peng, Z. et al. The gut microbiome is associated with clinical response to anti-PD-1/PD-L1 immunotherapy in gastrointestinal cancer. Cancer Immunol. Res. 8, 1251–1261 (2020).
Salgia, N. J. et al. Stool microbiome profiling of patients with metastatic renal cell carcinoma receiving anti-PD-1 immune checkpoint inhibitors. Eur. Urol. 78, 498–502 (2020).
Davar, D. et al. Fecal microbiota transplant overcomes resistance to anti-PD-1 therapy in melanoma patients. Science 371, 595–602 (2021).
Andrews, M. C. et al. Gut microbiota signatures are associated with toxicity to combined CTLA-4 and PD-1 blockade. Nat. Med. 27, 1432–1441 (2021).
Takada, K. et al. Clinical impact of probiotics on the efficacy of anti-PD-1monotherapy in patients with non-small cell lung cancer: a multicenter retrospective survival analysis study with inverse probability of treatment weighting. Int. J. Cancer 149, 473–482 (2021).
Chau, J. et al. Prospective correlation between the patient microbiome with response to and development of immune-mediated adverse effects to immunotherapy in lung cancer. BMC Cancer 21, 808 (2021).
Mao, J. et al. Gut microbiome is associated with the clinical response to anti-PD-1 based immunotherapy in hepatobiliary cancers. J. Immunother. Cancer 9, e003334 (2021).
Chung, M. W. et al. Gut microbiome composition can predict the response to nivolumab in advanced hepatocellular carcinoma patients. World J. Gastroenterol. 27, 7340–7349 (2021).
Yang, D. et al. Blood microbiota diversity determines response of advanced colorectal cancer to chemotherapy combined with adoptive T cell immunotherapy. Oncoimmunology 10, 1976953 (2021).
Yin, H. et al. The commensal consortium of the gut microbiome is associated with favorable responses to anti-programmed death protein 1 (PD-1) therapy in thoracic neoplasms. Cancer Biol. Med. 18, 1040 (2021).
Lee, K. A. et al. Cross-cohort gut microbiome associations with immune checkpoint inhibitor response in advanced melanoma. Nat. Med. 28, 535–544 (2022).
McCulloch, J. A. et al. Intestinal microbiota signatures of clinical response and immune-related adverse events in melanoma patients treated with anti-PD-1. Nat. Med. 28, 545–556 (2022).
Simpson, R. C. et al. Diet-driven microbial ecology underpins associations between cancer immunotherapy outcomes and the gut microbiome. Nat. Med. 28, 2344–2352 (2022).
Newsome, R. C. et al. Interaction of bacterial genera associated with therapeutic response to immune checkpoint PD-1 blockade in a United States cohort. Genome Med. 14, 35 (2022).
Chu, S. et al. Airway Fusobacterium is associated with poor response to immunotherapy in lung cancer. Onco Targets Ther. 15, 201–213 (2022).
Lee, P. C. et al. Gut microbiota and metabolites associate with outcomes of immune checkpoint inhibitor-treated unresectable hepatocellular carcinoma. J. Immunother. Cancer 10, e004779 (2022).
Che, H. et al. Association of Helicobacter pylori infection with survival outcomes in advanced gastric cancer patients treated with immune checkpoint inhibitors. BMC Cancer 22, 904 (2022).
Peiffer, L. B. et al. Composition of gastrointestinal microbiota in association with treatment response in individuals with metastatic castrate resistant prostate cancer progressing on enzalutamide and initiating treatment with anti-PD-1 (pembrolizumab). Neoplasia 32, 100822 (2022).
Smith, M. et al. Gut microbiome correlates of response and toxicity following anti-CD19 CAR T cell therapy. Nat. Med. 28, 713–723 (2022).
Hu, Y. et al. CAR-T cell therapy-related cytokine release syndrome and therapeutic response is modulated by the gut microbiome in hematologic malignancies. Nat. Commun. 13, 5313 (2022).
Xu, L. et al. Genomic and microbial factors affect the prognosis of anti-pd-1 immunotherapy in nasopharyngeal carcinoma. Front. Oncol. 12, 953884 (2022).
Routy, B. et al. Fecal microbiota transplantation plus anti-PD-1 immunotherapy in advanced melanoma: a phase I trial. Nat. Med. 29, 2121–2132 (2023).
Dora, D. et al. Non-small cell lung cancer patients treated with Anti-PD1 immunotherapy show distinct microbial signatures and metabolic pathways according to progression-free survival and PD-L1 status. Oncoimmunology 12, 2204746 (2023).
Haberman, Y. et al. Gut microbial signature in lung cancer patients highlights specific taxa as predictors for durable clinical benefit. Sci. Rep. 13, 2007 (2023).
Han, Z. et al. The gut microbiome affects response of treatments in HER2-negative advanced gastric cancer. Clin. Transl. Med. 13, e1312 (2023). 2023.
Zhao, W. et al. Fecal microbiota transplantation plus tislelizumab and fruquintinib in refractory microsatellite stable metastatic colorectal cancer: an open-label, single-arm, phase II trial (RENMIN-215). EClinicalMedicine 66, 102315 (2023).
Stein-Thoeringer, C. K. et al. A non-antibiotic-disrupted gut microbiome is associated with clinical responses to CD19-CAR-T cell cancer immunotherapy. Nat. Med. 29, 906–916 (2023).
Björk, J. R. et al. Longitudinal gut microbiome changes in immune checkpoint blockade-treated advanced melanoma. Nat. Med. 30, 785–796 (2024).
Sun, L. et al. Gut microbiota and metabolites associated with immunotherapy efficacy in extensive-stage small cell lung cancer: a pilot study. J. Thorac. Dis. 16, 6936–6954 (2024).
Sitthideatphaiboon, P. et al. Dietary pattern and the corresponding gut microbiome in response to immunotherapy in Thai patients with advanced non-small cell lung cancer (NSCLC). Sci. Rep. 14, 27791 (2024).
Yang, Z. et al. Gut microbiome model predicts response to neoadjuvant immunotherapy plus chemoradiotherapy in rectal cancer. Med 5, 1293–1306.e4 (2024).
Zhang, M. et al. A gut microbiota rheostat forecasts responsiveness to PD-L1 and VEGF blockade in mesothelioma. Nat. Commun. 15, 7187 (2024).
Rosario, S. R. et al. Integrative multi-omics analysis uncovers tumor-immune-gut axis influencing immunotherapy outcomes in ovarian cancer. Nat. Commun. 15, 10609 (2024).
Davar, D. et al. Neoadjuvant vidutolimod and nivolumab in high-risk resectable melanoma: a prospective phase II trial. Cancer Cell 42, 1898–1918.e12 (2024).
Gao, W. et al. Predictable regulation of gut microbiome in immunotherapeutic efficacy of gastric cancer. Genes Immun. 26, 1–8 (2024).
Macandog, A. D. G. et al. Longitudinal analysis of the gut microbiota during anti-PD-1 therapy reveals stable microbial features of response in melanoma patients. Cell Host Microbe 32, 2004–2018.e9 (2025).
Yu, Z. et al. Lachnoclostridium intestinal flora is associated with immunotherapy efficacy in nasopharyngeal carcinoma. Head. Neck 47, 269–281 (2025).
Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450 (2013).
Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569–573 (2013).
Viaud, S. et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 342, 971–976 (2013).
Sivan, A. et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 350, 1084–1089 (2015).
Vétizou, M. et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 350, 1079–1084 (2015).
Pinato, D. J. et al. Association of prior antibiotic treatment with survival and response to immune checkpoint inhibitor therapy in patients with cancer. JAMA Oncol. 5, 1774–1778 (2019).
Elkrief, A. et al. Antibiotics are associated with decreased progression-free survival of advanced melanoma patients treated with immune checkpoint inhibitors. Oncoimmunology 8, e1568812 (2019).
Zhu, X. et al. Interplay between gut microbial communities and metabolites modulates pan-cancer immunotherapy responses. Cell Metab. S1550-4131(24)00495-9 (2025).
Gunjur, A. et al. A gut microbial signature for combination immune checkpoint blockade across cancer types. Nat. Med. 30, 797–809 (2024).
Byrd, A. L. et al. Gut microbiome stability and dynamics in healthy donors and patients with non-gastrointestinal cancers. J. Exp. Med 218, e20200606 (2021).
Kartal, E. et al. A faecal microbiota signature with high specificity for pancreatic cancer. Gut 71, 1359–1372 (2022).
Terrisse, S. et al. Intestinal microbiota influences clinical outcome and side effects of early breast cancer treatment. Cell Death Differ. 28, 2778–2796 (2021).
He, D. et al. Response to PD-1-based immunotherapy for non- small cell lung cancer altered by gut microbiota. Oncol. Ther. 9, 647–657 (2021).
Derosa, L. et al. Intestinal Akkermansia muciniphila predicts clinical response to PD-1 blockade in patients with advanced non- small-cell lung cancer. Nat. Med. 28, 315–324 (2022).
Zepeda-Rivera, M. et al. A distinct Fusobacterium nucleatum clade dominates the colorectal cancer niche. Nature 628, 424–432 (2024).
Baulu, E., Gardet, C., Chuvin, N. & Depil, S. TCR-engineered T cell therapy in solid tumors: State of the art and perspectives. Sci. Adv. 9, eadf3700 (2023).
Uslu, U., Castelli, S. & June, C. H. CAR T cell combination therapies to treat cancer. Cancer Cell 42, 1319–1325 (2024).
Paulos, C. M. et al. Microbial translocation augments the function of adoptively transferred self/tumor-specific CD8+ T cells via TLR4 signaling. J. Clin. Invest. 117, 2197–2204 (2007).
Uribe-Herranz, M. et al. Gut microbiota modulates adoptive cell therapy via CD8α dendritic cells and IL-12. JCI Insight 3, e94952 (2018).
Ma, C. et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 360, eaan5931 (2018).
Luu, M. et al. Regulation of the effector function of CD8+ T cells by gut microbiota-derived metabolite butyrate. Sci. Rep. 8, 14430 (2018).
He, Y. et al. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8+ T cell immunity. Cell Metab. 33, 988–1000.e7 (2021).
Luu, M. et al. Microbial short-chain fatty acids modulate CD8+ T cell responses and improve adoptive immunotherapy for cancer. Nat. Commun. 12, 4077 (2021).
Asokan, S., Cullin, N., Stein-Thoeringer, C. K. & Elinav, E. CAR-T cell therapy and the gut microbiota. Cancers (Basel) 15, 794 (2023).
Cremonesi, E. et al. Gut microbiota modulate T cell trafficking into human colorectal cancer. Gut 67, 1984–1994 (2018).
Zhu, C. et al. Remolding the tumor microenvironment by bacteria augments adoptive T cell therapy in advanced-stage solid tumors. Signal Transduct. Target Ther. 9, 307 (2024).
Pulendran, B., Arunachalam, P. S. & O’Hagan, D. T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 20, 454–475 (2021).
Shirota, H. & Klinman, D. M. Recent progress concerning CpG DNA and its use as a vaccine adjuvant. Expert Rev. Vaccines 13, 299–312 (2014).
Ribas, A. et al. Overcoming PD-1 blockade resistance with CpG-A toll-like receptor 9 agonist vidutolimod in patients with metastatic melanoma. Cancer Discov. 11, 2998–3007 (2021).
Guiducci, C. et al. Redirecting in vivo elicited tumor infiltrating macrophages and dendritic cells towards tumor rejection. Cancer Res. 65, 3437–3446 (2005).
Iida, N. et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 342, 967–970 (2013).
Levy, M., Kolodziejczyk, A. A., Thaiss, C. A. & Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 17, 219–232 (2017).
Rescigno, M. et al. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2, 361–367 (2001).
Deng, H. et al. A novel strain of Bacteroides fragilis enhances phagocytosis and polarises M1 macrophages. Sci. Rep. 6, 29401 (2016).
Ramanan, D., Tang, M. S., Bowcutt, R., Loke, P. & Cadwell, K. Bacterial sensor Nod2 prevents inflammation of the small intestine by restricting the expansion of the commensal Bacteroides vulgatus. Immunity 41, 311–324 (2014).
Birchenough, G. M., Nystrom, E. E., Johansson, M. E. & Hansson, G. C. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science 352, 1535–1542 (2016).
Castro-Dopico, T. et al. Anti-commensal IgG drives intestinal inflammation and Type 17 immunity in ulcerative colitis. Immunity 50, 1099–1114.e10 (2019).
Qiu, Y. et al. Lactobacillus plantarum enhanced IL-22 production in Natural Killer (NK) cells that protect the integrity of intestinal epithelial cell barrier damaged by enterotoxigenic Escherichia coli. Int J. Mol. Sci. 18, 2409 (2017).
Guo, X. et al. Innate lymphoid cells control early colonization resistance against intestinal pathogens through ID2-dependent regulation of the microbiota. Immunity 42, 731–743 (2015).
Hu, C. et al. Gut microbiota-derived short-chain fatty acids regulate group 3 innate lymphoid cells in HCC. Hepatology 77, 48–64 (2023).
Polonio, C. M. et al. 2025. The aryl hydrocarbon receptor: a rehabilitated target for therapeutic immune modulation. Nat Rev Drug Discov (2025). Online ahead of print.
Balachandran, V. P. et al. Identification of unique neoantigen qualities in long-term survivors of pancreatic cancer. Nature 551, 512–516 (2017).
Davar, D. & Zarour, H. M. Facts and hopes for gut microbiota interventions in cancer immunotherapy. Clin. Cancer Res 28, 4370–4384 (2022). 2022.
Kalaora, S. et al. Identification of bacteria-derived HLA-bound peptides in melanoma. Nature 592, 138–143 (2021).
Sanchez, J. F., Maknojia, A. A. & King, K. Y. Blood and guts: how the intestinal microbiome shapes hematopoiesis and treatment of hematologic disease. Blood 143, 1689–1701 (2024).
Atarashi, K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341 (2011).
Round, J. L. et al. The toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 332, 974–977 (2011).
Qian, L. J. et al. Early-life gut microbial colonization shapes Th1/Th2 balance in asthma model in BALB/c mice. BMC Microbiol. 17, 135 (2017).
Erturk-Hasdemir, D. et al. Symbionts exploit complex signaling to educate the immune system. Proc. Natl. Acad. Sci. USA 116, 26157–26166 (2019).
Liu, Q., Sun, Z. & Chen, L. Memory T cells: strategies for optimizing tumor immunotherapy. Protein Cell 11, 549–564 (2020).
Lu, Y. et al. Gut microbiota influence immunotherapy responses: mechanisms and therapeutic strategies. J. Hematol. Oncol. 15, 47 (2022).
Fujiwara, D. et al. Systemic control of plasmacytoid dendritic cells by CD8+ T cells and commensal microbiota. J. Immunol. 180, 5843–5852 (2008).
Wei, B. et al. Commensal microbiota and CD8+ T cells shape the formation of invariant NKT cells. J. Immunol. 184, 1218–1226 (2010).
Zhu, R. et al. Gut microbiota: influence on carcinogenesis and modulation strategies by drug delivery systems to improve cancer therapy. Adv. Sci. (Weinh.) 8, 2003542 (2021).
Pei, B., Peng, S., Huang, C. & Zhou, F. Bifidobacterium modulation of tumor immunotherapy and its mechanism. Cancer Immunol. Immunother. 73, 94 (2024).
Krautkramer, K. A., Fan, J. & Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 19, 77–94 (2021).
Yang, W. & Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol. Immunol. 18, 866–877 (2021).
Zhang, T., Yang, Y., Liang, Y., Jiao, X. & Zhao, C. Beneficial effect of intestinal fermentation of natural polysaccharides. Nutrients 10, 1055 (2018).
Viretta, L., Oldfield, D. & Sali, A. Inflammatory bowel diseases and the efficacy of probiotics as functional foods. Front Biosci. (Elite Ed.) 16, 13 (2024).
Sun, J. et al. Butyrate as a promising therapeutic target in cancer: From pathogenesis to clinic. Int J. Oncol. 64, 44 (2024).
Gomes, A. P. et al. Altered propionate metabolism contributes to tumour progression and aggressiveness. Nat. Metab. 4, 435–443 (2022).
Xue, C. et al. Tryptophan metabolism in health and disease. Cell Metab. 35, 1304–1326 (2023).
Bender, M. J. et al. Dietary tryptophan metabolite released by intratumoral Lactobacillus reuteri facilitates immune checkpoint inhibitor treatment. Cell 186, 1846–1862.e26 (2023).
Hezaveh, K. et al. Tryptophan-derived microbial metabolites activate the aryl hydrocarbon receptor in tumor-associated macrophages to suppress anti-tumor immunity. Immunity 55, 324–340.e8 (2022).
Zhang, L. et al. Inhibition of UBA6 by inosine augments tumour immunogenicity and responses. Nat. Commun. 13, 5413 (2022).
Mager, L. F. et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 369, 1481–1489 (2020).
An, D. et al. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 156, 123–133 (2014).
Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 18, 33–50 (2018).
Lee, M., Lee, S. Y. & Bae, Y. S. Functional roles of sphingolipids in immunity and their implication in disease. Exp. Mol. Med. 55, 1110–1130 (2023).
Johnson, E. L. et al. Sphingolipids produced by gut bacteria enter host metabolic pathways impacting ceramide levels. Nat. Commun. 11, 2471 (2020).
Chakraborty, P. et al. Pro-Survival Lipid Sphingosine-1-Phosphate Metabolically Programs T Cells to Limit Anti-tumor Activity. Cell Rep. 28, 1879–1893.e7 (2019).
Grüner, N. & Mattner, J. Bile Acids and Microbiota: Multifaceted and Versatile Regulators of the Liver-Gut Axis. Int J. Mol. Sci. 22, 1397 (2021).
Paik, D. et al. Human gut bacteria produce TauEta17-modulating bile acid metabolites. Nature 603, 907–912 (2022).
Li, W. et al. A bacterial bile acid metabolite modulates Treg activity through the nuclear hormone receptor NR4A1. Cell Host Microbe 29, 1366–1377.e9 (2021).
Režen, T. et al. The role of bile acids in carcinogenesis. Cell Mol. Life Sci. 79, 243 (2022).
Yin, Q., Ni, J. J. & Ying, J. E. Potential mechanisms and targeting strategies of the gut microbiota in antitumor immunity and immunotherapy. Immun. Inflamm. Dis. 12, e1263 (2024).
Uebanso, T., Shimohata, T., Mawatari, K. & Takahashi, A. Functional Roles of B-Vitamins in the Gut and Gut Microbiome. Mol. Nutr. Food Res 64, e2000426 (2020).
Yang, Y. et al. Dietary vitamin B3 supplementation induces the antitumor immunity against liver cancer via biased GPR109A signaling in myeloid cell. Cell Rep. Med. 5, 101718 (2024).
Hossain, K. S., Amarasena, S. & Mayengbam, S. B Vitamins and Their Roles in Gut Health. Microorganisms 10, 1168 (2022).
Sujol, G., Docquier, A., Boulahtouf, A., Castet-Nicolas, A. & Cavaillès, V. Vitamin B6 and cancer: from clinical data to molecularly mechanisms. Bull. Cancer 98, 1201–1208 (2011).
Daillere, R. et al. Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity 45, 931–943 (2016).
Lam, K. C. et al. Microbiota triggers STING-type I IFN-dependent monocyte reprogramming of the tumor microenvironment. Cell 184, 5338–56.e21 (2021).
Wang, T. et al. Inosine is an alternative carbon source for CD8(+)-T-cell function under glucose restriction. Nat. Metab. 2, 635–647 (2020).
Fluckiger, A. et al. Cross-reactivity between tumor MHC class I-restricted antigens and an enterococcal bacteriophage. Science 369, 936–942 (2020).
Bessell, C. A. et al. Commensal bacteria stimulate antitumor responses via T cell cross-reactivity. JCI insight 5, e135597 (2020).
Naghavian, R. et al. Microbial peptides activate tumour-infiltrating lymphocytes in glioblastoma. Nature 617, 807–817 (2023).
Martins, F. et al. Adverse effects of immune-checkpoint inhibitors: epidemiology, management and surveillance. Nat. Rev. Clin. Oncol. 16, 563–580 (2019).
Baxi, S., Yang, A. & Gennarelli, R. L. Immune-related adverse events for anti-PD-1 and anti-PD-L1 drugs: systematic review and meta-analysis. BMJ 360, k793 (2018).
Wang, Z., Guo, Y. & Han, W. Current status and perspectives of chimeric antigen receptor modified T cells for cancer treatment. Protein Cell 8, 896–925 (2017).
Liu, Y. et al. Intestinal microbiota and its effect on vaccine-induced immune amplification and tolerance. Vaccines (Basel) 12, 868 (2024).
Zheng, Y. et al. Effect of a probiotic formula on gastrointestinal health, immune responses and metabolic health in adults with functional constipation or functional diarrhea. Front. Nutr. 10, 1196625 (2023).
Usyk, M. et al. Bacteroides vulgatus and Bacteroides dorei predict immune-related adverse events in immune checkpoint blockade treatment of metastatic melanoma. Genome Med. 13, 160 (2021).
Wang, Y. et al. Fecal microbiota transplantation for refractory immune checkpoint inhibitor-associated colitis. Nat. Med. 24, 1804–1808 (2018).
Blake, S. J. et al. The immunotoxicity, but not anti-tumor efficacy, of anti-CD40 and anti-CD137 immunotherapies is dependent on the gut microbiota. Cell Rep. Med. 2, 100464 (2021).
Chaput, N. et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 28, 1368–1379 (2017).
Nejman, D. et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 368, 973–980 (2020).
Choi Y, et al. Immune checkpoint blockade induces gut microbiota translocation that augments extraintestinal antitumor immunity. Sci Immunol. 8, eabo2003 (2023).
Ferrari, V. & Rescigno, M. The intratumoral microbiota: friend or foe?. Trends Cancer 9, 472–479 (2023).
Xu, J. et al. Research progress on the impact of intratumoral microbiota on the immune microenvironment of malignant tumors and its role in immunotherapy. Front. Immunol. 15, 1389446 (2024).
Zhang, T. et al. Intratumoral Fusobacterium nucleatum recruits tumor-associated neutrophils to promote gastric cancer progression and immune evasion. Cancer Res. 85, 1819–1841 (2025). 2025.
Gur, C. et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 42, 344–355 (2015).
Chen, S. et al. Fusobacterium nucleatum reduces METTL3-mediated m6A modification and contributes to colorectal cancer metastasis. Nat. Commun. 13, 1248 (2022).
Ma, Y. et al. Intratumor microbiome-derived butyrate promotes lung cancer metastasis. Cell Rep. Med. 5, 101488 (2024).
Miao, B. P. et al. Inhibition of squamous cancer growth in a mouse model by Staphylococcal enterotoxin B-triggered Th9 cell expansion. Cell Mol. Immunol. 14, 371–379 (2017).
Sun, J., Chen, F. & Wu, G. Potential effects of gut microbiota on host cancers: focus on immunity, DNA damage, cellular pathways, and anticancer therapy. ISME J. 17, 1535–1551 (2023).
Wilson, R. M. et al. The human gut bacterial genotoxin colibactin alkylates DNA. Science 363, eaar7785 (2019).
Goodwin, A. C. et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc. Natl Acad. Sci. USA 108, 15354–15359 (2011).
Geller, L. T. et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 357, 1156–1160 (2017).
Wyatt, M. & Greathouse, K. L. Targeting Dietary and Microbial Tryptophan-Indole Metabolism as Therapeutic Approaches to Colon Cancer. Nutrients 13, 1189 (2021).
Overacre-Delgoffe, A. E. et al. Microbiota-specific T follicular helper cells drive tertiary lymphoid structures and anti-tumor immunity against colorectal cancer. Immunity 54, 2812–2824.e4 (2021).
Yang, L., Li, A., Wang, Y. & Zhang, Y. Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy. Signal Transduct. Target Ther. 8, 35 (2023).
Shi, Y. et al. Intra-tumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling. J. Exp. Med. 217, e20192282 (2020).
Dwivedi, M., Kumar, P., Laddha, N. C. & Kemp, E. H. Induction of regulatory T cells: A role for probiotics and prebiotics to suppress autoimmunity. Autoimmun. Rev. 15, 379–392 (2016).
Han, S. et al. d-lactate modulates M2 tumor-associated macrophages and remodels immunosuppressive tumor microenvironment for hepatocellular carcinoma. Sci. Adv. 9, eadg2697 (2023).
Takeuchi, T., Nakanishi, Y. & Ohno, H. Microbial metabolites and gut immunology. Annu. Rev. Immunol. 42, 153–178 (2024).
Domzaridou, E. et al. The impact of oral antibiotics prior to cancer diagnosis on overall patient survival: findings from an english population-based cohort study. Curr. Oncol. 30, 8434–8443 (2023).
Kim, H. et al. The effect of antibiotics on the clinical outcomes of patients with solid cancers undergoing immune checkpoint inhibitor treatment: a retrospective study. BMC Cancer 19, 1100 (2019).
Huang, X. Z. et al. Antibiotic use and the efficacy of immune checkpoint inhibitors in cancer patients: a pooled analysis of 2740 cancer patients. Oncoimmunology 8, e1665973 (2019).
Wilson, B. E. et al. The effect of antibiotics on clinical outcomes in immune-checkpoint blockade: a systematic review and meta-analysis of observational studies. Cancer Immunol. Immunother. 69, 343–354 (2020).
Yang, M. et al. Antibiotic administration shortly before or after immunotherapy initiation is correlated with poor prognosis in solid cancer patients: an up-to-date systematic review and meta-analysis. Int. Immunopharmacol. 88, 106876 (2020).
Cren, P. Y. et al. Is the survival of patients treated with ipilimumab affected by antibiotics? An analysis of 1585 patients from the French National hospital discharge summary database (PMSI). Oncoimmunology 9, 1846914 (2020).
Kulkarni, A. A. et al. Comparative analysis of antibiotic exposure association with clinical outcomes of chemotherapy versus immunotherapy across three tumour types. ESMO Open 5, e000803 (2020).
Lalani, A. A. et al. Effect of antibiotic use on outcomes with systemic therapies in metastatic renal cell carcinoma. Eur. Urol. Oncol. 3, 372–381 (2020).
Fessas, P. et al. Early antibiotic exposure is not detrimental to therapeutic effect from immunotherapy in hepatocellular carcinoma. Liver Cancer 10, 583–592 (2021).
Kostine, M. et al. Baseline co-medications may alter the anti-tumoural effect of checkpoint inhibitors as well as the risk of immune-related adverse events. Eur. J. Cancer 157, 474–484 (2021).
Wu, Q., Liu, J., Wu, S. & Xie, X. The impact of antibiotics on efficacy of immune checkpoint inhibitors in malignancies: a study based on 44 cohorts. Int Immunopharmacol. 92, 107303 (2021).
Mohiuddin, J. J. et al. Association of antibiotic exposure with survival and toxicity in patients with melanoma receiving immunotherapy. J. Natl Cancer Inst. 113, 162–170 (2021).
Jing, Y. et al. Association of antibiotic treatment with immune-related adverse events in patients with cancer receiving immunotherapy. J. Immunother. Cancer 10, e003779 (2022).
Nyein, A. F. et al. Effect of prior antibiotic or chemotherapy treatment on immunotherapy response in non-small cell lung cancer. BMC Cancer 22, 101 (2022).
Zhao, L. et al. Association of blood biochemical indexes and antibiotic exposure with severe immune-related adverse events in patients with advanced cancers receiving PD-1 inhibitors. J. Immunother. 45, 210–216 (2022).
Poizeau, F. et al. The association between antibiotic use and outcome among metastatic melanoma patients receiving immunotherapy. J. Natl Cancer Inst. 114, 686–694 (2022).
Pressner, S., Heiland, M., Preissner, R., Wirth, M. & Wollenberg, B. Antibiotics significantly decrease the survival of head and neck carcinoma patients with immunotherapy: a real-world analysis of more than 3000 cases. Cancers (Basel) 15, 2342 (2023).
Pinato, D. J. et al. Association between antibiotics and adverse oncological outcomes in patients receiving targeted or immune-based therapy for hepatocellular carcinoma. JHEP Rep. 5, 100747 (2023).
Kulkarini, A. A. et al. Association of antibiotic exposure with residual cancer burden in HER2-negative early stage breast cancer. NPJ Breast Cancer 10, 24 (2024).
Glitza, I. C. et al. Randomized placebo-controlled, biomarker-stratified phase ib microbiome modulation in melanoma: impact of antibiotic preconditioning on microbiome and immunity. Cancer Discov. 14, 1161–1175 (2024).
Elkrief, A. et al. Antibiotics are associated with worse outcomes in lung cancer patients treated with chemotherapy and immunotherapy. NPJ Precis Oncol. 8, 143 (2024).
Prasad, R. et al. Antibiotic-induced loss of gut microbiome metabolic output correlates with clinical responses to CAR T-cell therapy. Blood 145, 823–839 (2024).
Takada, K., Takamori, S., Shimokawa, M., Pinato, D. & Cortellini, A. Prior antibiotics, proton pump inhibitors, and probiotics in patients with extensive stage small cell lung cancer treated with immune checkpoint blockade: a post-hoc analysis of the phase I/III IMpower 133 trial. Int J. Cancer 156, 914–919 (2025).
Bonnefin, C. et al. Antibiotics use decreases survival in cutaneous squamous cell carcinoma patients receiving immune checkpoint inhibitors. Eur. J. Cancer 217, 115223 (2025).
Cheng, M. et al. Microbiota modulate tumoral immune surveillance in lung through a γδT17 immune cell-dependent mechanism. Cancer Res. 74, 4030–4041 (2014).
Derosa et al. Negative association of antibiotics on clinical activity of immune checkpoint inhibitors in patients with advanced renal cell and non-small-cell lung cancer. Ann. Oncol. 29, 1437–1444 (2018).
Chalabi, M. et al. Efficacy of chemotherapy and atezolizumab in patients with non-small-cell lung cancer receiving antibiotics and proton pump inhibitors: pooled post hoc analyses of the OAK and POPLAR trials. Ann. Oncol. 31, 525–531 (2020).
Palleja, A. et al. Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 3, 1255–1265 (2018).
Ahmed, J. et al. Use of broad-spectrum antibiotics impacts outcome in patients treated with immune checkpoint inhibitors. Oncoimmunology 7, e1507670 (2018).
Cai, J., Sun, L. & Gonzalez, F. J. Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe 30, 289–300 (2022).
Pernigoni, N. et al. Commensal bacteria promote endocrine resistance in prostate cancer through androgen biosynthesis. Science 374, 216–224 (2021).
Terrisse, S. et al. Immune system and intestinal microbiota determine efficacy of androgen deprivation therapy against prostate cancer. J. Immunother. Cancer 10, e004191 (2022).
Ravà, M. et al. Therapeutic synergy between tigecycline and venetoclax in a preclinical model of MYC/BCL2 double-hit B cell lymphoma. Sci. Transl. Med. 10, eaan8723 (2018).
Pushalkar, S. et al. The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov. 8, 403–416 (2018).
Sethi, V., Vitiello, G. A., Saxena, D., Miller, G. & Dudeja, V. The role of the microbiome in immunologic development and its implication for pancreatic cancer immunotherapy. Gastroenterology 156, 2097–2115.e2 (2019).
Vitiello, G. A., Cohen, D. J. & Miller, G. Harnessing the microbiome for pancreatic cancer immunotherapy. Trends Cancer 5, 670–676 (2019). 2019.
Bertocchi, A. et al. Gut vascular barrier impairment leads to intestinal bacteria dissemination and colorectal cancer metastasis to liver. Cancer Cell 39, 708–724.e11 (2021).
Schwan, A., Sjölin, S., Trottestam, U. & Aronsson, B. Relapsing Clostridium difficile enterocolitis cured by rectal infusion of normal faeces. Scand. J. Infect. Dis. 16, 211–215 (1984).
Pai, N., Popov, J., Hill, L. & Hartung, E. Protocol for a double-blind, randomised, placebo-controlled pilot study for assessing the feasibility and efficacy of faecal microbiota transplant in a paediatric Crohn’s disease population: PediCRaFT Trial. BMJ Open 9, e030120 (2019).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/ NCT06218602 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT03353402 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT03772899 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT05251389 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT03341143 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04577729 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06623461 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04521075 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04951583 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT05008861 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06403111 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05502913 (Accessed 15 February 2025).
US National Library of MediciAccessed 15ne. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04924374 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04056026 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06486220 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05750030 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05690048 Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06405113 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04130763 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04038619 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04729322 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT05279677 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04163289 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04038619 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04758507 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04116775 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04883762 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT04521075 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/ct2/show/NCT05286294 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03819296 (Accessed 15 February 2025).
Chinese Clinical Trial Register. https://www.chictr.org.cn/historyversionpub.aspx?regno=ChiCTR2100042292 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05273255 (Accessed 15 February 2025).
Schluter, J. et al. The gut microbiota is associated with immune cell dynamics in humans. Nature 588, 303–307 (2020).
Zhao, Y. et al. Safety and efficacy of fecal microbiota transplantation for grade IV steroid refractory GI-GvHD patients: interim results from FMT2017002 trial. Front Immunol. 12, 678476 (2021).
Halesy, T. M. et al. Microbiome alteration via fecal microbiota transplantation is effective for refractory immune checkpoint inhibitor-induced colitis. Sci. Transl. Med. 15, eabq4006 (2023).
Helmink, B. A., Khan, M. A. W., Hermann, A., Gopalakrishnan, V. & Wargo, J. A. The microbiome, cancer, and cancer therapy. Nat. Med. 25, 377–88 (2019).
Bajaj, J. S. et al. Microbial functional change is linked with clinical outcomes after capsular fecal transplant in cirrhosis. JCI Insight 4, e133410 (2019).
Ianiro, G. et al. Faecal microbiota transplantation for the treatment of diarrhoea induced by tyrosine-kinase inhibitors in patients with metastatic renal cell carcinoma. Nat. Commun. 11, 4333 (2020).
Hartstra, A. V. et al. Infusion of donor feces affects the gut-brain axis in humans with metabolic syndrome. Mol. Metab. 42, 101076 (2020).
DeFilipp, Z. et al. Drug-resistant E. coli bacteremia transmitted by fecal microbiota transplant. N. Engl. J. Med. 381, 2043–2050 (2019).
Vendrik, K. E. W. et al. Periodic screening of donor faeces with a quarantine period to prevent transmission of multidrug-resistant organisms during faecal microbiota transplantation: a retrospective cohort study. Lancet Infect. Dis. 21, 711–721 (2021).
Podolsky, S. H. Metchnikoff and the microbiome. Lancet 380, 1810–1811 (2012).
Hill, C. et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514 (2014).
Rong, Y. et al. Reactivity toward Bifidobacterium longum and Enterococcus hirae demonstrate robust CD8( + ) T cell response and better prognosis in HBV-related hepatocellular carcinoma. Exp. Cell Res. 358, 352–359 (2017).
Lee, B. et al. Modulation of the gut microbiota alters the tumour-suppressive efficacy of Tim-3 pathway blockade in a bacterial species- and host factor-dependent manner. Microorganisms 8, 1395 (2020).
Zhuo, Q. et al. Lysates of Lactobacillus acidophilus combined with CTLA-4-blocking antibodies enhance antitumor immunity in a mouse colon cancer model. Sci. Rep. 9, 20128 (2019).
Lee, S. H. et al. Bifidobacterium bifidum strains synergize with immune checkpoint inhibitors to reduce tumour burden in mice. Nat. Microbiol. 6, 277–288 (2021).
Li, Q. et al. Streptococcus thermophilus inhibits colorectal tumorigenesis through secreting beta-galactosidase. Gastroenterology 160, 1179–1193.e14 (2021).
Corb Aron, R. A. et al. Recognizing the benefits of pre-/probiotics in metabolic syndrome and type 2 diabetes mellitus considering the influence of Akkermansia muciniphila as a key gut bacterium. Microorganisms 9, 618 (2021).
Ansaldo, E. et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science 364, 1179–1184 (2019).
Lordan, C., Thapa, D., Ross, R. P. & Cotter, P. D. Potential for enriching next-generation health-promoting gut bacteria through prebiotics and other dietary components. Gut Microbes 11, 1–20 (2020).
Shi, L. et al. Combining IL-2-based immunotherapy with commensal probiotics produces enhanced anti-tumor immune response and tumor clearance. J. Immunother. Cancer 8, e000973 (2020).
Tanoue, T. et al. A defined commensal consortium elicits CD8 T cells and anti-cancer immunity. Nature 565, 600–605 (2019).
Montalban-Arques, A. et al. Commensal Clostridiales strains mediate effective anti-cancer immune response against solid tumors. Cell Host Microbe 29, 1573–1588.e7 (2021).
Roy, S. & Trichieri, G. Microbiota: a key orchestrator of cancer therapy. Nat. Rev. Cancer 17, 271–285 (2017).
Das, M. Probiotics for chemoradiotherapy-induced oral mucositis. Lancet Oncol. 20, E14 (2019).
Jiang, C. et al. A randomized, double-blind, placebo-controlled trial of probiotics to reduce the severity of oral mucositis induced by chemoradiotherapy for patients with nasopharyngeal carcinoma. Cancer 125, 1081–1090 (2019).
Consoli, M. L. et al. Randomized clinical trial: impact of oral administration of Saccharomyces boulardii on gene expression of intestinal cytokines in patients undergoing colon resection. JPEN J. Parenter. Enter. Nutr. 40, 1114–1121 (2016).
Hibberd, A. A. et al. Intestinal microbiota is altered in patients with colon cancer and modified by probiotic intervention. BMJ Open Gastroenterol. 4, e000145 (2017).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04988841 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03595683 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04699721 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04909034 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05094167 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06428422 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06768931 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05032014 (Accessed 15 February 15, 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05122546 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05220124 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03686202 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03775850 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03637803 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04208958 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06508034 (Accessed 15 February 2025).
Lauté-Caly, D. L. et al. The flagellin of candidate live biotherapeutic Enterococcus gallinarum MRx0518 is a potent immunostimulant. Sci. Rep. 9, 801 (2019).
Available online: https://www.biospace.com/4d-pharma-announces-positive-interim-results-from-the-phase-i-ii-study-of-the-combination-of-mrx0518-and-keytruda-pembrolizumab-for-the-treatment-of-renal-cell-carcinoma (Accessed 15 February 2025)
Tomita, Y. et al. Association of probiotic Clostridium butyricum therapy with survival and response to immune checkpoint blockade in patients with lung cancer. Cancer Immunol. Res. 8, 1236–1242 (2020).
Tomita, Y. et al. Clostridium butyricum therapy restores the decreased efficacy of immune checkpoint blockade in lung cancer patients receiving proton pump inhibitors. Oncoimmunology 11, 2081010 (2022).
McHale, D. et al. Oral delivery of a single microbial strain, EDP1503, induces anti-tumor responses via gut-mediated activation of both innate and adaptive immunity. Ann. Oncol. 31, S195 (2020).
Feuerstadt, P. et al. SER-109, an oral microbiome therapy for recurrent Clostridioides difficile infection. N. Engl. J. Med. 386, 220–229 (2022).
Snigdha, S. et al. Probiotics: Potential novel therapeutics for microbiota-gut-brain axis dysfunction across gender and lifespan. Pharm. Ther. 231, 107978 (2022).
Cabrita, R. et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature 577, 561–565 (2022).
Ruddle, N. H. Tertiary Lymphoid Structures and Immunotherapy: Challenges and Opportunities. Methods Mol. Biol. 2864, 299–312 (2025).
Carmody, R. N. et al. Diet dominates host genotype in shaping the murine gut microbiota. Cell Host Microbe 17, 72–84 (2015).
Le Chatelier, E. et al. Richness of human gut microbiome correlates with metabolic markers. Nature 500, 541–546 (2013).
Wastyk, H. C. et al. Gut-microbiota-targeted diets modulate human immune status. Cell 184, 4137–4153.e14 (2021).
Buque, A. et al. Immunoprophylactic and immunotherapeutic control of hormone receptor-positive breast cancer. Nat. Commun. 11, 3819 (2020).
Ferrere, G. et al. Ketogenic diet and ketone bodies enhance the anticancer effects of PD-1 blockade. JCI Insight 6, e145207 (2021).
Levesque, S. et al. A synergistic triad of chemotherapy, immune checkpoint inhibitors, and caloric restriction mimetics eradicates tumors in mice. Oncoimmunology 8, e1657375 (2019).
Pietrocola, F. et al. Caloric restriction mimetics enhance anticancer immunosurveillance. Cancer Cell 30, 147–160 (2016).
Pomatto-Watson, L. C. D. et al. Daily caloric restriction limits tumor growth more effectively than caloric cycling regardless of dietary composition. Nat. Commun. 12, 6201 (2021).
Wang, Y. et al. NAD+ supplement potentiates tumor-killing function by rescuing defective TUB-mediated NAMPT transcription in tumor-infiltrated T cells. Cell Rep. 36, 109516 (2021).
Collins, N. et al. The bone marrow protects and optimizes immunological memory during dietary restriction. Cell 178, 1088–1101.e15 (2019).
Desai, M. S. et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 167, 1339–1353.e21 (2016).
Tong, H. et al. Dual impacts of serine/glycine-free diet in enhancing antitumor immunity and promoting evasion via PD-L1 lactylation. Cell Metab. 36, 2493–2510.e9 (2024).
Koh, A., De Vadder, F., Kovatcheva-Datchary, P. & Bäckhed, F. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 165, 1332–1345 (2016).
Morrison, D. J. & Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7, 189–200 (2016).
Coutzac, C. et al. Systemic short chain fatty acids limit antitumor effect of CTLA-4 blockade in hosts with cancer. Nat. Commun. 11, 2168 (2020).
Sanders, M. E., Merenstein, D. J., Reid, G., Gibson, G. R. & Rastall, R. A. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 16, 605–616 (2019).
Gibson, G. R. et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 14, 491–502 (2017).
Becerril-Alarcon, Y. et al. Inulin supplementation reduces systolic blood pressure in women with breast cancer undergoing neoadjuvant chemotherapy. Cardiovasc Ther. 2019, 5707150 (2019).
Beukema, M., Faas, M. M. & de Vos, P. The effects of different dietary fiber pectin structures on the gastrointestinal immune barrier: impact via gut microbiota and direct effects on immune cells. Exp. Mol. Med. 52, 1364–1376 (2020).
Le Bastard, Q. et al. The effects of inulin on gut microbial composition: a systematic review of evidence from human studies. Eur. J. Clin. Microbiol. Infect. Dis. 39, 403–413 (2020).
Panebianco, C., Andriulli, A. & Pazienza, V. Pharmacomicrobiomics: exploiting the drug-microbiota interactions in anticancer therapies. Microbiome 6, 92 (2018).
Zheng, D. W. et al. Prebiotics-encapsulated probiotic spores regulate gut microbiota and suppress colon cancer. Adv. Mater. 32, e2004529 (2020).
Han, K. et al. Generation of systemic anti-tumour immunity via the in situ modulation of the gut microbiome by an orally administered inulin gel. Nat. Biomed. Eng. 5, 1377–1388 (2021).
Zhang, S. L. et al. Pectin supplement significantly enhanced the anti- PD-1 efficacy in tumor- bearing mice humanized with gut microbiota from patients with colorectal cancer. Theranostics 11, 4155–4170 (2021).
Huang, J. et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 71, 734–745 (2022).
Messaoudene, M. et al. A natural polyphenol exerts antitumor activity and circumvents anti-PD-1 resistance through effects on the gut microbiota. Cancer Discov. 12, 1070–1087 (2022).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06466434 (Accessed 15 February 2025).
Faria, S. et al. Diet and Immune Effects Trial (DIET)- a randomized, double-blinded dietary intervention study in patients with melanoma receiving immunotherapy. BMC Cancer 24, 1493 (2024).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06298734 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06236360 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT04866810 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03709147 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT03700437 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06671613 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05763992 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06610097 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05083416 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06391099 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05356182 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT05832606 (Accessed 15 February 2025).
US National Library of Medicine. ClinicalTrials.gov https://clinicaltrials.gov/study/NCT06438588 (Accessed 15 February 2025).
Liang, K. et al. Genetically engineered Salmonella typhimurium: recent advances in cancer therapy. Cancer Lett. 448, 168–181 (2019).
Chang, Y. et al. Dihydromyricetin attenuates Escherichia coli lipopolysaccharide-induced ileum injury in chickens by inhibiting NLRP3 inflammasome and TLR4/NF-κB signalling pathway. Vet. Res. 51, 72 (2020).
Taniguchi, S., Shimatani, Y. & Fujimori, M. Tumor-Targeting therapy using gene-engineered anaerobic-nonpathogenic Bifidobacterium longum. Methods Mol. Biol. 1409, 49–60 (2016).
Gurbatri, C. R., Arpaia, N. & Danino, T. Engineering bacteria as interactive cancer therapies. Science 378, 858–864 (2022).
Geiger, R. et al. L-Arginine modulates T cell metabolism and enhances survival and anti-tumor activity. Cell 167, 829–42.e13 (2016).
Canale, F. P. et al. Metabolic modulation of tumours with engineered bacteria for immunotherapy. Nature 598, 662–666 (2021).
Leventhal, D. S. et al. Immunotherapy with engineered bacteria by targeting the STING pathway for anti-tumor immunity. Nat. Commun. 11, 2739 (2020).
Chowdhury, S. et al. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med 25, 1057–1063 (2019).
Redenti, A. et al. Probiotic neoantigen delivery vectors for precision cancer immunotherapy. Nature 635, 453–461 (2024).
Yang, S. et al. Non-pathogenic E. coli displaying decoy-resistant IL18 mutein boosts anti-tumor and CAR NK cell responses. Nat Biotechnol. (2024) Online ahead of print.
Eng, C. et al. A phase II study of axalimogene filolisbac for patients with previously treated, unresectable, persistent/recurrent loco-regional or metastatic anal cancer. Oncotarget 11, 1334–1343 (2020).
Galicia-Carmona, T. et al. ADXS11-001 LM-LLO as specific immunotherapy in cervical cancer. Hum. Vaccin. Immunother. 17, 2617–2625 (2021).
Pennington, S. H. et al. Nonspecific effects of oral vaccination with live-attenuated Salmonella Typhi strain Ty21a. Sci. Adv. 5, eaau6849 (2019).
Domingos-Pereira, S. et al. Preclinical efficacy and safety of the Ty21a vaccine strain for intravesical immunotherapy of non-muscle-invasive bladder cancer. Oncoimmunology 6, e1265720 (2017).
Drake, C. G. et al. Safety and preliminary immunogenicity of JNJ-64041809, a live-attenuated, double-deleted Listeria monocytogenes-based immunotherapy, in metastatic castration-resistant prostate cancer. Prostate Cancer Prostatic Dis. 25, 219–228 (2022).
Schmitz-Winnenthal, F. H. et al. A phase 1 trial extension to assess immunologic efficacy and safety of prime-boost vaccination with VXM01, an oral T cell vaccine against VEGFR2, in patients with advanced pancreatic cancer. Oncoimmunology 7, e1303584 (2018).
Chen, Y. E. et al. Engineered skin bacteria induce antitumor T cell responses against melanoma. Science 380, 203–210 (2023).
Preet, R. et al. Gut commensal Bifidobacterium-derived extracellular vesicles modulate the therapeutic effects of anti-PD-1 in lung cancer. Nat. Commun. 16, 3500 (2025).
Liu, X. Z. et al. Bioengineered Bacterial Membrane Vesicles with Multifunctional Nanoparticles as a Versatile Platform for Cancer Immunotherapy. ACS Appl. Mater. Interfaces 15, 3744–3759 (2023).
Zhang, J. et al. Antitumorigenic potential of Lactobacillus-derived extracellular vesicles: p53 succinylation and glycolytic reprogramming in intestinal epithelial cells via SIRT5 modulation. Cell Biol. Toxicol. 40, 66 (2024).
Hsu, B. B. et al. Dynamic modulation of the gut microbiota and metabolome by bacteriophages in a mouse model. Cell Host Microbe 25, 803–814.e5 (2019).
Selle, K. et al. In vivo targeting of clostridioides difficile using phage-delivered CRISPR-Cas3 antimicrobials. mBio 11, e00019–e00020 (2020).
Voorhees, P. J., Cruz-Teran, C., Edelstein, J. & Lai, S. K. Challenges & opportunities for phage-based in situ microbiome engineering in the gut. J. Control Release 326, 106–119 (2020).
Zheng, D. W. et al. Phage-guided modulation of the gut microbiota of mouse models of colorectal cancer augments their responses to chemotherapy. Nat. Biomed. Eng. 3, 717–728 (2019).
Cieplak, T., Soffer, N., Sulakvelidze, A. & Nielsen, D. S. A bacteriophage cocktail targeting Escherichia coli reduces E. coli in simulated gut conditions, while preserving a non-targeted representative commensal normal microbiota. Gut Microbes 9, 391–399 (2018).
Lipska, K., Gumieniczek, A. & Filip, A. A. Anticonvulsant valproic acid and other short-chain fatty acids as novel anticancer therapeutics: Possibilities and challenges. Acta Pharm. 70, 291–301 (2020).
Chen, P. B. et al. Directed remodeling of the mouse gut microbiome inhibits the development of atherosclerosis. Nat. Biotechnol. 38, 1288–1297 (2020).
Reardon, D. et al. EO2401 microbiome derived therapeutic vaccine + nivolumab, with/without standard continuous, or low-dose symptom directed, bevacizumab, in recurrent glioblastoma: phase 1–2 EOGBM1–18/ROSALIE study. J. Immunother. Cancer 10, A1–A1603 (2022).
Available online: https://www.onclive.com/view/eo2401-plus-nivolumab-and-bevacizumab-shows-activity-in-progressive-recurrent-glioblastoma (Accessed 27 February 2025)
Zhang, S. et al. Tumor microbiome: roles in tumor initiation, progression, and therapy. Mol. Biomed. 6, 9 (2025).
Yousefi, Y., Baines, K. J. & Maleki Vareki, S. Microbiome bacterial influencers of host immunity and response to immunotherapy. Cell Rep. Med 5, 101487 (2024).
Mrochen, D. M. et al. Staphylococcus aureus host tropism and its implications for murine infection models. Int J. Mol. Sci. 21, 7061 (2020).
Christen, U. & Hintermann, E. Animal models for autoimmune hepatitis: are current models good enough?. Front Immunol. 13, 898615 (2022).
Cani, P. D. Human gut microbiome: hopes, threats and promises. Gut 67, 1716–1725 (2018).
Allali, I. et al. A comparison of sequencing platforms and bioinformatics pipelines for compositional analysis of the gut microbiome. BMC Microbiol. 17, 194 (2017).
Lozupone, C. A. et al. Meta-analyses of studies of the human microbiota. Genome Res. 23, 1704–1714 (2013).
Available online: https://asm.org/articles/2024/february/fecal-microbiota-transplants-past-present-future (Accessed 27 February 2025)
Xu, F. et al. The interplay between host genetics and the gut microbiome reveals common and distinct microbiome features for complex human diseases. Microbiome 8, 145 (2020).
Qin, Y. et al. Combined effects of host genetics and diet on human gut microbiota and incident disease in a single population cohort. Nat. Genet. 54, 134–142 (2022).
Ianiro, G. et al. Variability of strain engraftment and predictability of microbiome composition after fecal microbiota transplantation across different diseases. Nat. Med 28, 1913–1923 (2022).
Kato, S., Subbiah, V. & Kurzrock, R. Counterpoint: successes in the pursuit of precision medicine: biomarkers take credit. J. Natl Compr. Canc Netw. 15, 863–866 (2017).
Bronkhorst, A. J., Ungerer, V. & Holdenrieder, S. The emerging role of cell-free DNA as a molecular marker for cancer management. Biomol. Detect Quantif. 17, 100087 (2019).
Rehan, M. et al. Smart capsules for sensing and sampling the gut: status, challenges and prospects. Gut 73, 186–202 (2023).
Lucchetti, M. et al. Integration of multiple flexible electrodes for real-time detection of barrier formation with spatial resolution in a gut-on-chip system. Microsyst. Nanoeng. 10, 18 (2024).
Jordan, A., Carding, S. R. & Hall, L. J. The early-life gut microbiome and vaccine efficacy. Lancet Microbe 3, e787–e794 (2022).
Jing, N. et al. Enhancing therapeutic effects of murine cancer vaccine by reshaping gut microbiota with Lactobacillus rhamnosus GG and jujube powder. Front. Immunol. 14, 1195075 (2023).
de Jong, S. E., Olin, A. & Pulendran, B. The impact of the microbiome on immunity to vaccination in humans. Cell Host Microbe 28, 169–179 (2020).
Issrani, R., Reddy, J., Dabah, T. H. E. M. & Prabhu, N. Role of oral microbiota in carcinogenesis: a short review. J. Cancer Prev. 27, 16–21 (2022).
Choudhry, H. The microbiome and its implications in cancer immunotherapy. Molecules 26, 206 (2021).
Lau, H. C. H., Zhang, X. & Yu, J. Gut microbiota and immune alteration in cancer development: implication for immunotherapy. eGastroenterology 1, e100007 (2023).
Acknowledgements
This research received no external funding.
Author information
Authors and Affiliations
Contributions
Conceptualization, W.L. & K.Z., Software & figure, Y.L.; Validation, H.Z. & Q.L.; Writing-Original Draft Preparation, W.L. & K.Z. & Y.L.; Writing-Review & Editing, K.Z.; Visualization, H.Z. & Q.L.; Supervision, H.Z. & Q.L.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lei, W., Zhou, K., Lei, Y. et al. Gut microbiota shapes cancer immunotherapy responses. npj Biofilms Microbiomes 11, 143 (2025). https://doi.org/10.1038/s41522-025-00786-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41522-025-00786-8
This article is cited by
-
Gut microbiota drives cancer evolution and therapy resistance
Molecular Cancer (2026)
-
Recent advances and future perspectives in multidisciplinary research on osteosarcoma
Holistic Integrative Oncology (2026)
-
Gut microbial diversity and compositional shifts are associated with differential tumor growth rates in mice
BMC Microbiology (2025)
-
Engineering bacteria for enhanced tumor therapy: from surface modification to synthetic genetic circuits
Journal of Hematology & Oncology (2025)
-
Gut microbial metabolites in cancer immunomodulation
Molecular Cancer (2025)
