
Abstract
The plant circadian clock drives temporal differences in susceptibility to pathogens. We investigated the role of TIMING OF CAB EXPRESSION 1 (TOC1) in the regulation of defence against Botrytis cinerea in Arabidopsis. The temporal variation in susceptibility to B. cinerea observed in wild-type Arabidopsis was abolished in TOC1-ox and toc1-2 plants under both diurnal and constant light conditions. In addition, TOC1-ox plants were more susceptible than Col-0 following inoculation at dawn, while inoculation at night led to enhanced resistance in toc1-2 plants versus C24 plants, suggesting that TOC1 is a negative regulator of immunity. RNA-seq analysis showed that the genes mis-regulated in toc1-2 plants had significant enrichment for terms related to biotic stress, an overrepresentation of G-box elements in their promoters and included genes encoding key transcription factors (TFs) involved in defence against necrotrophic pathogens. Chromatin immunoprecipitation-qPCR showed that TOC1 occupies G-box containing regions of the defence TFs ERF4, ORA47, ORA59 and WRKY33 in a pathogen-responsive and MYC2-dependent manner. We suggest that the phased TOC1 occupancy of defence gene promoters contributes to the gating of plant immunity against necrotrophic pathogens, while the MYC2-dependent release of TOC1 in response to pathogen detection allows plants to mount an acute immune response.
Similar content being viewed by others


Introduction
Plant immune responses against pathogens are tightly regulated at multiple levels as they are energetically costly, can negatively impact plant growth and often involve mutually antagonistic signalling pathways, depending on the pathogen type. The plant circadian clock restricts elongation and growth to specific phases of the day1. It also appears to gate defence responses to biotrophic and necrotrophic pathogens, such that a maximal response occurs at the time of day when they are most likely to be encountered or plants are most vulnerable2,3,4,5,6. It has been proposed that the growth inhibition observed during immune responses to pathogen attack is not solely due to resource competition between growth and defence, but also to conflicting activation of hormone signalling pathways which are also subject to circadian co-ordination7. The fungal clock has also been shown to regulate the virulence of the plant pathogen Botrytis cinerea, reflecting an interplay between the circadian systems of host and pathogen8.
The circadian oscillator consists of three interlocking transcription-translation feedback loops: the core, morning and evening loops. The core loop consists of two morning expressed MYB-related transcription factors (TFs) CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY), and the evening-phased TIMING OF CAB EXPRESSION 1 (TOC1) also known as PSEUDO RESPONSE REGULATOR 1 (PRR1)9,10,11. CCA1 and LHY repress the expression of TOC1 during the day by binding to the evening element motif (AAATATCT) present in its promoter12. At dusk, CCA1 and LHY levels decline and TOC1 levels increase, repressing the expression of CCA1 and LHY in the evening by binding to their promoters13,14,15. TOC1 regulates target genes by binding to DNA via the TOC1 morning element (T1ME, TGTG) which is present in both the morning element (ME, GTGTGG) and hormone upregulated at dawn (HUD, CATGTG) motifs, and also via a G-box (CACGTG) containing motif13,14.
Plants respond to pathogen attack with a two-layered innate immune system16,17. The first layer is mediated by pattern recognition receptors (PRRs) at the plasma membrane which recognise conserved pathogen-associated molecular patterns (PAMPs), including flagellin and chitin, to activate PAMP-triggered immunity (PTI). To allow successful colonisation of the host plant, pathogens have evolved virulence factors including effectors which are secreted into the host cells where they target a range of host proteins including PRRs to suppress PTI18,19. Notably effectors from oomycete and viral pathogens that target the clock protein CCA1 HIKING EXPEDITION (CHE) have recently been identified20,21. Plants in turn have evolved intracellular nucleotide-binding leucine-rich repeat receptors which directly or indirectly detect effectors and activate effector-triggered immunity (ETI). Although mediated by different receptors, PTI and ETI trigger many overlapping responses suggesting that they are interconnected, and it has been proposed that ETI acts as an amplification module that is reliant on the PTI machinery22.
An output common to PTI and ETI is the production of defence hormones, most notably salicylic acid (SA), jasmonic acid (JA) and ethylene (ET) which play critical roles in host transcriptional reprogramming23. ET and JA activate signalling pathways leading to resistance against necrotrophic pathogens, while SA mediates responses to biotrophic pathogens24. The crosstalk between the JA/ET and SA signalling pathways is broadly characterised by reciprocal antagonism, although synergistic interactions have also been reported25. Plant pathogens have evolved virulence mechanisms to take advantage of this antagonism; for example the JA-isoleucine (JA-Ile) analogue coronatine secreted by Pseudomonas syringae that suppresses SA-mediated defence responses including stomatal closure in response to PAMP detection26,27.
JA plays a critical role in defence against both necrotrophic pathogens and herbivores. Its biosynthesis and signalling show circadian oscillations, with peak JA accumulation occurring during the subjective day in Arabidopsis28,29. These fluctuations drive rhythmic expression of JA-responsive genes and align with diurnal patterns of pathogen and herbivore activity30. While the expression of JA-responsive genes may be rhythmically induced because of oscillations in JA accumulation and signalling, some JA-responsive genes are directly regulated by DNA-binding core clock proteins. Thus, besides their role in the clock, core clock proteins also regulate genes involved in stress-responsive output pathways by binding to regulatory motifs present in their promoters13,14,28.
As clock TFs are rhythmically expressed, their presence and activity at specific times of the day may contribute to the temporal expression of defence genes4,14,31. For example, LUX ARRHYTHMO, a component of the evening complex, has been implicated in both SA and JA-mediated immune responses and has been shown to bind to the promoters of EDS1 and JAZ55. The clock component TIME FOR COFFEE acts as a negative regulator of JA signalling by interacting with MYC2, a key TF in JA signalling, and promoting its degradation32. MYC TFs bind to G-boxes and related variants (CACATG, CACGCG, and CACGAG) in the promoters of their target genes. In the absence of JA, MYC TFs are bound by JA ZIM-domain (JAZ) proteins, which recruit the general repressor TOPLESS through Novel Interactor of JAZ (NINJA)33,34. However, in the presence of JA-Ile, JAZ proteins interact with CORONATINE INSENSITIVE1 (COI1) and are targeted for proteasomal degradation, resulting in the removal of the repressor complex and allowing activation of gene transcription by MYC proteins. In Arabidopsis, the paralogs MYC2, MYC3 and MYC4 function redundantly to regulate temporal immunity against the necrotrophic pathogen B. cinerea35.
Transcriptome profiling showed that around half of the TF-encoding genes that responded differentially to B. cinerea inoculation at subjective dawn versus night exhibit rhythmic expression, and that these genes are strongly enriched for direct targets of TOC1, PRR5 and PRR73. ChIP-seq analysis demonstrated that TOC1 binds to the promoters of TIC, JAZ1, JAZ5 and JAZ9, as well as the promoters of other genes involved in JA/ET signalling including EIN2, EBF1, ERF5, LOX3 and ORA4714. However, whether TOC1 directly regulates immune responses against B. cinerea is unclear. TOC1 has DNA-binding activity and occupies the G-box motif in promoters that is recognised by the MYC TFs. Given this overlap in promoter motif recognition, we hypothesised that TOC1 and MYC2 might co-regulate a subset of defence genes, providing a mechanism for circadian modulation of JA-dependent immunity.
We tested this by first comparing defence responses to the necrotrophic pathogen B. cinerea in TOC1 mis-expressing plants and wild-type plants and then used RNA-seq to identify defence genes potentially regulated by TOC1. We next examined TOC1 binding to G-box-containing promoters of selected defence-related TFs that are known MYC2 targets and assessed whether pathogen challenge alters TOC1 or MYC2 occupancy at these sites. Finally, we tested whether the absence of either TOC1 or MYC2 affected the binding of the other TF at target gene promoters.
Results
Mis-expression of TOC1 abolishes temporal variation in immunity and results in altered susceptibility to B. cinerea under both LL and LD conditions
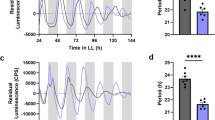
To determine whether TOC1 is required for temporal variation in resistance to B. cinerea we examined the susceptibility profile of plants mis-expressing this gene under constant light (LL) conditions. The toc1-2 null mutant (C24 background), which has a shortened period of 22 h under LL, and the TOC1 over-expressor (TOC1-ox, Col-0 background), which displays arrhythmic circadian outputs under LL, were inoculated with B. cinerea spores at subjective dawn (corresponding to circadian time 24, CT24) and subjective night (CT42), the times of minimal and maximal susceptibility to B. cinerea in wild-type Arabidopsis3,10,36.
As expected, both wild-type ecotypes displayed significant time-of-day dependent variation in mean lesion size following pathogen challenge (Fig. 1A); lesion sizes on Col-0 leaves were significantly smaller 72 hpi at CT24 versus CT42, as were those on C24 leaves. In contrast, neither the TOC1-ox nor toc1-2 lines showed a significant difference in lesion size between leaves inoculated at CT24 versus CT42 (Fig. 1A, Supplementary Fig. 1A). Lesions on TOC1-ox leaves inoculated at CT24 were significantly larger than those on Col-0 leaves (Fig. 1A), while average lesion sizes on TOC1-ox leaves after inoculation at both CT24 and CT42 were not significantly different to those on Col-0 leaves infected at CT42. In contrast, lesion sizes at both time points on toc1-2 leaves were comparable to those observed on wild-type C24 leaves following infection at CT24 (Fig. 1A, Supplementary Fig. 1A).

Detached leaves from four-week-old plants grown under a 16 h light and 8 h dark cycle were inoculated with B. cinerea spores (A) under constant light conditions at subjective dawn (CT24) or subjective night (CT42) and (B) under diurnal conditions at dawn (ZT0) or night (ZT18) and lesion size measured at 72 hpi. Data shown are mean values ± SEM, with the number of leaves (from independent plants) inoculated indicated within the bars. Under LL (A) time of inoculation (F = 11.6, p = 0.0009, η2 = 0.02), plant genotype (F = 180.8, p < 0.0001, η2 = 0.78) and the interaction between these factors (F = 4.1, p 0.008, η2 = 0.02) had a significant effect on lesion size in a two-way ANOVA. Under LD (B) time of inoculation (F = 24.6, p < 0.0001, η2 = 0.05), plant genotype (F = 87.3, p < 0.0001, η2 = 0.54) and the interaction between these factors (F = 3.3, p 0.021, η2 = 0.02) had a significant effect on lesion size in a two-way ANOVA. Mean lesion sizes with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. Results shown are from one experiment representative of two (A) or three (B) independent experiments.
The loss of temporal variation in susceptibility to B. cinerea observed in toc1-2 was unexpected as the clock is functional in this line under LL conditions, albeit with a shorter period of 22 h10. This suggests that TOC1 might have a role in regulating immunity output pathways in addition to its function as a core component of the circadian clock. To test this we repeated these experiments under LD conditions, when both toc1-2 and TOC1-ox are rhythmic with a period of approximately 24 h37,38 to mitigate the effects of broader circadian mis-regulation in these lines. Again, Col-0 and C24 wild-type plants displayed temporal variation in susceptibility to B. cinerea that was not observed in either TOC1-ox or toc1-2 (Fig. 1B, Supplementary Fig. 1B). Lesion areas were significantly larger on TOC1-ox versus Col-0 leaves at both ZT0 and ZT18, while irrespective of the time of inoculation the lesions on toc1-2 leaves were not significantly different from those on C24 leaves inoculated at ZT0 (Fig. 1B, Supplementary Fig. 1B). Collectively, these experiments indicate that TOC1 is required for temporal variation in immunity to B. cinerea in Arabidopsis under both LL and LD conditions. Furthermore, constitutive expression of TOC1 appears to lock plants into the phenotype observed in wild-type plants inoculated at ZT18/CT42 (i.e. maximal susceptibility), while loss of TOC1 locks plants into the phenotype observed in wild-type plants inoculated at ZT0/CT24 (i.e. maximal resistance). This is consistent with TOC1 acting as a repressor of immunity against B. cinerea.
RNA-seq analysis of toc1-2 mutants provides additional evidence that TOC1 is a negative regulator of immunity
To determine whether TOC1 is indeed acting as a negative regulator of immunity we performed RNA-seq analysis on uninfected and B. cinerea-challenged toc1-2 and C24 plants. To minimise the potential for false positives due to differences in the phase of gene expression between wild-type and toc1-2 plants in LL, we carried out this experiment in LD conditions, where there is minimal difference in the phase of gene expression in toc1-238. Plants were grown under standard 16 h light/8 h dark conditions for four weeks and detached leaves inoculated with either B. cinerea spores or grape juice (mock control) at dawn (ZT0). RNA-seq analysis was performed on samples harvested at ZT18 and ZT22, times at which TOC1 expression is maximal in wild-type plants and by which point a substantial transcriptional response to B. cinerea infection can be expected in Arabidopsis39,40. Two strategies were used to investigate whether TOC1 might regulate defence genes in Arabidopsis. In the first, the transcriptome profiles of uninfected C24 and toc1-2 plants were compared to look for evidence of regulation of defence genes by TOC1 in the absence of pathogen attack. In the second, the transcriptional response that occurs in response to B. cinerea challenge was compared in C24 versus toc1-2 plants.
In the absence of fungal challenge, there were 252 and 1370 differentially expressed genes (DEGs) between C24 and toc1-2 plants at ZT18 and ZT22 respectively, with an overlap of 103 genes. The majority of these DEGs were upregulated in the toc1-2 mutant; 197/252 (78%) at ZT18 and 860/1370 (63%) at ZT22 (Supplementary Data 1), in line with view that TOC1 generally acts as a transcriptional repressor. This proportion is similar to the 69% of DEGs that were upregulated in the toc1-101 mutant versus Col-0 at ZT12 when grown under a 12 h light/12 h dark cycle41. While these studies are not directly comparable due to the use of different mutants, genetic backgrounds, light-dark cycles and sampling times we note that 70/197 genes upregulated in toc1-2 at ZT18 (2 h into the 8 h dark period) were also upregulated in toc1-101 at ZT12 (start of the 12 h dark period)41.
To explore the potential biological functions of these DEGs, we combined the DEGs from ZT18 and ZT22 and performed GO enrichment analysis on the resulting 969 upregulated and 550 downregulated sets of genes. DEGs upregulated in toc1-2 plants were enriched for GO terms associated with abiotic stimuli such as light and water responses, as well as terms related to photosynthesis, circadian rhythm and photomorphogenesis. Notably, terms related to biotic stress were also enriched in the DEG gene sets (Fig. 2A). The GO terms “defence response” and “response to bacterium” were enriched in both up- and downregulated genes, while “response to fungus” and “wounding” were enriched in the down-regulated genes only. There was also enrichment for terms relating to phytohormone signalling, with “hormone related signalling” and “response to ET” enriched in the upregulated genes and “response to JA” in the downregulated genes. In total 307 DEGs (including 22 TFs) were annotated with GO terms relating to biotic stress/defence responses, of which 173 were upregulated in toc1-2 plants and 134 downregulated (Supplementary Data 2).

A Gene ontology enrichment analysis of genes mis-regulated in unchallenged toc1-2 versus C24 plants. GO terms are ranked in descending order by p-value (-log10) as determined by Fisher’s exact test with Bonferroni correction. B Venn diagrams indicating number of genes differentially expressed (DE) in response to Botrytis cinerea in toc1-2 and C24 plants at 18 and 22 hpi. Genes DE in C24 but not toc1-2 plants (indicated by white text) were classified as TOC1-modulated B. cinerea responsive genes. C Scatter plot showing log2 expression values of TOC1-modulated B. cinerea responsive genes in C24 plants infected with B. cinerea versus mock control (C24 inf/mock, X-axis) against those from unchallenged toc1-2 versus C24 plants (toc1-2/C24 mock, Y-axis). Pearsons R2 values are 0.66 (18 hpi) and 0.59 (22 hpi) respectively.
TOC1 binds directly to the T1ME found within the ME and HUD regulatory motifs13. ChIP-seq experiments have also identified a G-box related motif as a TOC1 binding site, and TOC1 occupies a G-box in the CCA1 promoter14,42. We looked for enrichment of these motifs in the upstream regions of the 307 DEGs annotated with biotic stress/defence GO terms to determine whether they are likely to be direct targets of TOC1. The G-box, ME and HUD motifs were all significantly overrepresented in the promoters of the 173 genes up-regulated in toc1-2 versus C24 in the absence of pathogen challenge, while no significant enrichment was observed in the 134 genes down-regulated in toc1-2 (Supplementary Table 1). Potential TOC1 binding motifs in the promoters of TFs mis-regulated in unchallenged toc1-2 plants are shown in Supplementary Table 2.
We interrogated the 307 DEGs annotated with GO terms relating to biotic stress/defence responses to explore the potential role of TOC1 in regulating immunity to B. cinerea. Genes upregulated in unchallenged toc1-2 plants include known key components of plant immune responses to necrotrophic pathogens (Supplementary Data 2). These include the ET- and JA-responsive TFs ETHYLENE INSENSITIVE 3 (EIN3), its homolog EIN3-like 1 (EIL1) and the AP2-ET response factor (ERF) TFs ERF4, ERF11, ERF72 and ORA59. Quantitative PCR analysis confirmed the increased expression of EIN3, EIL1 and ORA59 in toc1-2 plants and demonstrated that they were correspondingly downregulated in the TOC1-ox line (Supplementary Fig. 2). In addition, two genes encoding JA biosynthetic enzymes (OPC-8:CoA LIGASE and OXOPHYTODIENOATE-REDUCTASE 3) and one catalysing the synthesis of the inositol-phosphate 8 co-factor of the ASK1-COI1-JAZ co-receptor complex (ARABIDOPSIS HOMOLOG OF YEAST VIP1) are upregulated in toc1-2 plants. We also observed upregulation of the salicylic acid receptor NPR1 and of MKK9 which stabilises EIN3 in response to ET43.
Finally, we compared the transcriptional response to B. cinerea infection of toc1-2 and C24 plants. At 18 hpi, 584 and 516 genes were differentially expressed in response to B. cinerea in C24 and toc1-2 plants respectively, rising to 2008 and 3870 at 22 hpi (Fig. 2B, Supplementary Data 3). We classified DEGs as TOC1-modulated B. cinerea responsive genes if they showed a significant response to this pathogen in C24 but not toc1-2 plants; 295 (18 hpi) and 653 (22 hpi) DEGs met this criterion (Fig. 2B, Supplementary Fig. 3, Supplementary Data 4). There was also evidence for significant enrichment of known TOC1-binding motifs in this set of genes, with the G-box and ME enriched in the promoters of genes upregulated in response to B. cinerea, and the G-box and HUD motifs enriched in the promoters of downregulated genes (Supplementary Table 3). Note that there are two ways for a gene to appear in the TOC1-modulated B. cinerea responsive gene lists. The first is that the gene is already mis-regulated in uninfected toc1-2 plants and little or no further change in expression is observed in response to pathogen attack. The alternative is that TOC1 is required either for the induction or repression of the gene in response to B. cinerea infection. To determine the proportion of genes that fall into these two categories, we plotted the log2-fold change (log2-FC) values from B. cinerea versus mock inoculation in the C24 background against the log2-FC values from mock inoculated toc1-2 versus C24 plants for the 295 (18 hpi) and 653 (22 hpi) DEGs (Fig. 2C). A strong positive correlation was observed between these values at both 18 hpi (slope = 0.51, R2 = 0.66) and 22 hpi (slope = 0.47, R2 = 0.59), indicating that the majority of the TOC1-modulated B. cinerea-responsive genes showed no change in expression in toc1-2 following fungal challenge because they were already mis-regulated.
TOC1 occupancy of the G-box is pathogen responsive and modulated by MYC2
Having established that TOC1 is a negative regulator of immunity against necrotrophic pathogens, we then used ChIP-qPCR to determine whether it can bind directly to the promoters of defence TFs. Of the known TOC1-binding motifs, the G-box is the most interesting in the context of immunity as this motif is also bound by the MYC TFs which are key regulators of JA-mediated signalling, which could allow crosstalk between the clock and JA signalling. We therefore selected promoter regions of four defence TFs, ERF4, ORA47, ORA59 and WRKY33, containing G-box motifs for ChIP-qPCR analysis. The promoter regions analysed contain a single canonical G-box (CACGTG), except for the ORA47 amplicon which contains two, and no G-box variants (Supplementary Fig. 4). All four genes have previously been identified as potential targets of TOC1 and MYC2 and are also DE in response to B. cinerea infection14,40,44. While ORA59 and ERF4 were significantly upregulated in toc1-2 in our RNA-seq experiment, WKRY33 and ORA47 were not. However quantitative PCR analysis of WRKY33 and ORA47 in plants mis-expressing TOC1 confirmed that they are potential TOC1 target genes (Fig. 3), though WRKY33 appears to be positively regulated by TOC1 in contrast to ERF4, ORA47 and ORA59.

Detached leaves from four-week-old plants grown under a 16 h light and 8 h dark cycle were inoculated at dawn with Botrytis cinerea spore suspension or mock control. Leaf tissue was harvested at 18 hpi and relative expression of ORA47 and WRKY33 determined by qPCR with normalisation to the geometric mean of ACTIN2 and MON1 expression. Pathogen challenge (F = 101.6, p < 0.0001, η2 = 0.56) and plant genotype (F = 19.3, p < 0.0001, η2 = 0.32) had a significant effect on ORA47 expression in a two-way ANOVA. Pathogen challenge (F = 68.5, p < 0.0001, η2 = 0.47) and plant genotype (F = 18.7, p < 0.0001, η2 = 0.39) had a significant effect on WRKY33 expression in a two-way ANOVA. Mean expression values with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. The data presented are mean values ± SEM from three biological replicates. This experiment was performed once.
ChIP was performed at ZT18 on two-week-old plants expressing YFP-tagged TOC1 (TMG) or MYC2 (YPet) fusion proteins under the control of their native promoter 18 hpi with B. cinerea spores or grape juice mock control at dawn. To determine whether the binding of one of these TFs is influenced by the binding of the second, we generated plant lines expressing MYC2-YPet in a toc1-2 null background (YPet toc1) and TOC1-YFP in a myc2-2 null background (TMG myc2) and performed ChIP on mock and B. cinerea inoculated plants. As a positive control, we measured the occupancy of TOC1 at a known TOC1-binding site in the CCA1 promoter, a canonical G-box 193 bp upstream of the transcriptional start site (Supplementary Fig. 5)14,42. This experiment was performed at ZT0 (when TOC1 absent) and ZT15 (TOC1 present), and the expected enrichment was observed at ZT15 (Supplementary Fig. 5).
ChIP qPCR analyses indicated that in the absence of pathogen challenge TOC1 binds the G-box containing promoter regions of all four TFs in TMG plants, with significantly higher percentage input values observed for these DNA regions versus the exonic sites from the same genes (Fig. 4A–D). A marked decrease in TOC1 occupancy of target promoters was observed in response to B. cinerea infection. Significantly lower percentage input values were observed in infected versus mock TMG plants for all four TFs, and with the exception of ERF4, these were not significantly different from exonic control values (Fig. 4A–D). This was not due to degradation of TOC1 after B. cinerea infection as western blot analysis revealed no apparent difference in TOC1-YFP levels between uninfected and infected TMG plants (Supplementary Fig. 6).

Two-week old TMG and TMG myc2 seedlings were inoculated at ZT0 with B. cinerea spore suspension or mock control. A–D Seedlings were cross-linked at ZT18 and chromatin immunoprecipitated with an anti-GFP antibody. ChIP-qPCR analysis of TOC1 occupancy of a G-box containing region of the promoter and an exonic region of each TOC1-target gene was performed (position relative to transcription start site is indicated). Values shown are mean percentage input ± SEM from three independent experiments (two for ERF4 and ORA47 exonic regions). Mean values with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. No antibody control values and results of two-way ANOVA for each target gene are presented in Supplementary Tables 4 and 5, respectively. E–H Seedlings were harvested at ZT18 and relative expression of each target gene was determined by qPCR with normalization to the geometric mean of ACTIN2 and MON1 expression. The data presented are mean expression values ± SEM from three biological replicates. Mean values with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. Results of two-way ANOVA for each target gene are presented in Supplementary Table 5.
The results obtained in TMG myc2 plants were strikingly different. TOC1 occupancy of TF promoters was significantly lower in mock challenged TMG myc2 versus TMG plants, and the B. cinerea-responsive reduction in promoter occupancy by TOC1 was completely abolished in the TMG myc2 plants, with no significant difference in percentage input values observed between mock and infected plants except for ERF4 where TOC1 occupancy was now significantly higher in infected plants (Fig. 4A–D). To determine how these changes in TOC1 occupancy correlated with target gene expression level, we performed qPCR on mock and B. cinerea infected TMG and TMG myc2 plants. The mRNA levels of all four TFs were significantly higher in infected TMG plants versus mock controls (Fig. 4E–H). The loss of MYC2 had no effect on target gene expression in uninfected plants, but following infection was associated with significantly lower mRNA levels of ERF4, ORA47 and ORA59, and higher levels of WRKY33 in TMG myc2 versus TMG plants (Fig. 4E–H).
ChIP qPCR analysis also demonstrated enrichment of MYC2 binding at the G-box containing promoter regions versus exonic control regions (Fig. 5A–D). In contrast to TOC1 this was not responsive to pathogen challenge, except for ORA59 where percentage input values were significantly lower in infected versus uninfected YPet plants. As with TOC1-YFP, western blot analysis revealed no apparent change in MYC-YPet protein levels in infected plants (Supplementary Fig. 7). The loss of TOC1 had relatively minor effects on MYC2 binding to the promoters of the four defence TFs. No significant difference in MYC2 occupancy of the WRKY33 and ORA47 promoters was observed in either infected or uninfected YPet toc1 versus YPet plants, while reduced MYC2 occupancy of the ERF4 (uninfected and infected) and ORA59 (uninfected only) promoters was observed in YPet toc1 plants (Fig. 5A–D). Quantitative PCR analysis of mRNA levels revealed that the loss of TOC1 in YPet toc1 plants was associated with increased mRNA levels of ERF4 and ORA59 in the absence of pathogen challenge, and lower levels of WRKY33 in pathogen challenged plants relative to the YPet control (Fig. 5E–H).

Two-week old MYC2-YPet (YPet) and MYC2-YPet toc1 (YPet toc1) seedlings were inoculated at ZT0 with B. cinerea spore suspension or mock control. A–D Seedlings were cross-linked at ZT18 and chromatin immunoprecipitated with an anti-GFP antibody. ChIP-qPCR analysis of MYC2 occupancy of a G-box containing region of the promoter and an exonic region of each target gene was performed (position relative to transcription start site is indicated). Values shown are mean percentage input ± SEM from four independent experiments (two for ERF4 and ORA47 exonic regions). Mean values with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. No antibody control values and results of two-way ANOVA for each target gene are presented in Supplementary Tables 6 and 7, respectively. E–H Seedlings were harvested at ZT18 and relative expression of each target gene was determined by qPCR with normalization to the geometric mean of ACTIN2 and MON1 expression. The data presented are mean expression values ± SEM from three biological replicates. Mean values with different letters are significantly different (p < 0.05) as determined by Fisher LSD post-hoc test. Results of two-way ANOVA for each target gene are presented in Supplementary Table 7.
Finally, to explore the relationship between TOC1 and defence TF expression over the course of the day we analysed previously published RNA-seq data generated from 12 day old seedlings harvested every 3 h under LL conditions45. TOC1 mRNA levels peaked at 13.5 h (early in the subjective night), while the acrophase of ORA47, ORA59 and WRKY33 expression was markedly offset from that of TOC1, occurring at 9 h, as was that of ERF4, peaking at 18 h (Supplementary Fig. 8). This offset in peak mRNA levels was also evident in the translatome - the population of mRNAs bound by ribosomes - and thus likely persists at the protein level (Supplementary Fig. 8).
Discussion
The circadian clock allows plants to anticipate changes in the environment and optimise responses according to the time of the day. This includes attack by pathogens where the clock modulates immune responses in Arabidopsis such that the magnitude of the response varies with time of day. It has been well established that a functional circadian clock is required for optimal defence as arrhythmic mutants display both loss of temporal variation in susceptibility and increased susceptibility to pathogen challenge2,3,5,6. However, the mechanism by which the clock regulates immunity remains largely unknown. One possible mechanism is that the clock TFs themselves may directly regulate defence gene expression. The observation that both CCA1 and LUX bind directly to the promoters of defence genes supports this hypothesis5,46. Here we show that the core clock TF TOC1 functions as a negative regulator of immunity to necrotrophic pathogens by binding directly to the promoters of key defence TFs and that pathogen challenge results in the removal of TOC1 via a MYC2-dependent mechanism allowing an acute immune response to be activated.
Botrytis cinerea infection assays performed on TOC1-ox and toc1-2 plants showed that the temporal variation in susceptibility to this pathogen observed in wild-type Arabidopsis was abolished under LL conditions (Fig. 1A). While the loss of temporal variation could be attributed to the absence of a functional clock in TOC1-ox plants under LL conditions, the toc1-2 mutant maintains other rhythmic outputs under LL, albeit with a shortened period of 22 h10. Furthermore, this loss of temporal variation in TOC1 mis-expressing plants persisted under LD conditions, when both toc1-2 and TOC1-ox plants display rhythms with a 24 h period37,38. These observations suggest that TOC1 can function in plant immunity output pathways as well as within the core oscillator feedback loops. TOC1 levels peak at the end of the day, which coincides with the time at which immune responses to necrotrophic pathogens are weakest3. The constitutive expression of TOC1 had a more pronounced effect on susceptibility following inoculation at (subjective) dawn, when TOC1 would not normally be expressed. In contrast the toc1-2 mutation conferred enhanced resistance at (subjective) night when TOC1 expression would normally peak (Fig. 1A, B), suggesting that it results from the alleviation of TOC1-mediated repression of immunity during the night period.
RNA-seq analysis of toc1-2 plants provided further evidence that TOC1 is a regulator of immunity. Genes mis-expressed in unchallenged toc1-2 plants were enriched for GO terms related to biotic stress (Fig. 2A). This included 22 TFs, many of which have previously been linked to JA/ET-mediated immunity, including the key regulators EIN3, EIL1 and ORA5947,48,49. In addition, a comparison of the transcriptional response induced by B. cinerea infection of toc1-2 versus C24 plants identified a set of 295 genes at 18 hpi and 653 genes are 22 hpi that are pathogen-responsive in C24 but not in toc1-2 plants (Fig. 2B). Rather than requiring TOC1 for up- or downregulation in response to pathogen challenge, most of these genes appear to be mis-regulated in unchallenged toc1-2 plants (Fig. 2C).
Quantitative PCR analysis performed on TOC1 mis-expressing plants confirmed the constitutive induction of EIN3, EIL1 and ORA59 in toc1-2 plants, and demonstrated that they are also repressed in TOC1-ox plants (Supplementary Fig. 2). While EIN3 and EIL1 do not contain putative TOC1 binding sites in their promoters, 12 of the other TFs mis-expressed in toc1-2 plants do (Supplementary Table 2). These include MYB51 which is a positive regulator of camalexin biosynthesis, ERF4 which modulates ET signalling, ERF6 a positive regulator of defence against B. cinerea and ORA5948,50,51,52. Notably, ERF4 and ORA59 both contain a G-box in their promoters which is the motif recognised by the MYC TFs.
The binding of multiple TFs to the same promoter element in a gene creates opportunities for crosstalk and signal integration between different signalling pathways that can promote robustness in the immune response while also driving specificity25. Binding of both TOC1 and the MYC TFs to the G-box of defence TFs represents a potential site of crosstalk between the circadian clock and JA signalling, particularly as MYC2 and TOC1 exhibit the same phase of rhythmic expression, both peaking at the end of the day32,35. We selected the G-box containing promoter regions of four TFs previously identified as potential targets of TOC1 and MYC2 and with well characterised roles in JA/ET-mediated immunity to investigate the potential for such crosstalk. Both ORA59 and ERF4 act as integrators of JA and ET signalling in response to necrotrophic pathogen attack, but function as transcriptional activator and repressor respectively48,52,53. Plants over-expressing ORA59 display enhanced resistance to B. cinerea, while ORA59-silenced lines display enhanced susceptibility48. The opposite pattern of resistance to necrotrophic pathogens is observed in plants mis-expressing ERF453. The increased susceptibility of wrky33 mutants to B. cinerea has been linked to loss of WRKY33-mediated repression of JAZ gene expression and a failure to repress SA-mediated signalling during infection, which together lead to repression of JA-mediated immune responses. ORA47 binds directly to the promoters of several JA biosynthetic enzymes to drive JA production54,55.
ChIP-qPCR demonstrated that both TOC1 and MYC2 occupy G-box containing regions of the ERF4, ORA47, ORA59 and WRKY33 promoters in unchallenged plants at ZT18, the end of the light period (Figs. 4 and 5). TOC1 occupancy of these promoters was highly responsive to pathogen challenge and was largely abolished in B. cinerea infected plants. In contrast, with the exception of a modest decrease at the ORA59 promoter, MYC2 occupancy was unchanged (Figs. 4 and 5). Both TOC1 binding in unchallenged plants and the release of TOC1 upon infection appear to be MYC2-dependent; reduced TOC1 binding was observed in uninfected TMG myc2 plants and the attenuation of TOC1 binding observed in B. cinerea infected TMG plants was completely abolished (Fig. 4A–D). It is important to note that TMG was generated in the Col-0 background, so both TMG and the TMG myc2 line are homozygous for the wild-type TOC1 allele36. Therefore, any difference in apparent promoter occupancy cannot be due to changes in the level of wild-type TOC1 protein.
In contrast, binding of MYC2 to the promoters of these defence TFs was much less dependent on the presence of TOC1. The loss of TOC1 had no effect on MYC2 binding to the WRKY33 or ORA47 promoters but was associated with reduced binding to the ORA59 promoter in unchallenged plants, and to the ERF4 promoter in both unchallenged and infected plants (Fig. 5A–D). The increased occupancy by TOC1 in pathogen challenged TMG myc2 plants correlated with reduced mRNA levels of ERF4, ORA47 and ORA59 (Fig. 4E–H) suggesting that the release of TOC1 relieves transcriptional repression of these genes. This is in line with their increased expression in unchallenged toc1-2 plants (Fig. 3, Supplementary Fig. 2) and YPet toc1 plants (Fig. 5E–H). In contrast, WRKY33 mRNA levels were significantly higher in pathogen challenged TMG myc2 versus TMG control plants, and lower in infected YPet toc1 plants versus YPet controls. Together with the increased WRKY33 mRNA levels observed in TOC1-ox plants and reduced levels in toc1-2 mutants (Fig. 3), these data suggest that TOC1 is an activator of WKRY33 expression. However, it is nonetheless the case that WRKY33 mRNA levels increase in response to B. cinerea infection, when TOC1 occupation of the WRKY33 promoter is abolished (Fig. 4D, H) and thus the precise nature of TOC1 regulation of WRKY33 expression remains unclear.
Our data indicate that the loss of MYC2 results in decreased TOC1 occupancy at the G-box containing region of the promoters of these defence TFs in unchallenged plants, but increased occupancy in B. cinerea infected plants. It therefore appears that MYC2 might modulate TOC1 occupancy in an infection-dependent manner. How might this occur? Firstly, it is unknown whether TOC1 binds directly to the G-box DNA sequence or whether it does so indirectly via other DNA-binding proteins. While the CCT domain of TOC1 allows it to bind directly to DNA, TOC1 can also interact indirectly with target gene promoters through interactions with other DNA-binding proteins, including CHE, and the G-box binding phytochrome interacting factors (PIFs)13,15,56.
One possibility is that TOC1 does not bind directly to the G-box but instead is recruited to target gene promoters by MYC2. In the absence of a JA-Ile signal, MYC2 is bound to the G-box in a complex with JAZ proteins which recruit the co-repressors NINJA and TOPLESS57. JA-Ile binding to COI1 leads to ubiquitination of JAZ proteins relieving this repression. While it has not been demonstrated, it is possible that the removal of the JAZ proteins leads to a conformational change in MYC2. If so, one hypothesis for the mechanism of MYC2 modulation of TOC1 occupancy of defence gene promoters is as follows: in unchallenged plants inactive MYC2 bound to the G-box recruits TOC1 to defence gene promoters resulting in TOC1-mediated repression of target genes such as ERF4, ORA47 and ORA59 (Fig. 6A). This repression might occur via the formation of NUCLEAR FACTOR (NF)-YB/NF-YC/TOC1 trimeric complexes, regulated by TOC1 phosphorylation status, which can recruit HISTONE DEACETYLASE 15 to repress target gene expression58,59.

In uninfected plants, MYC2 is kept in an inactive state by the JAZ repressor complex. A TOC1 is recruited to the G-box (orange) by direct interaction with inactive MYC2, repressing defence gene expression. Detection of B. cinerea leads to COI1-mediated degradation of JAZ repressors. MYC2 undergoes a conformational change, releasing TOC1 and abolishing TOC1-mediated repression. B Inactive MYC2 promotes direct binding of TOC1 to the G-box. Active MYC2 displaces TOC1 from the G-box or recruits co-activators (X) which block TOC1 binding.
Pathogen challenge leads to an increase in JA-Ile levels resulting in the COI1-mediated degradation of JAZ proteins and a conformational change in MYC2 that leads to the release of TOC1 from the G-box, relieving the repression of target genes (Fig. 6A). The observation that TOC1 occupancy of defence gene promoters is not completely abolished in TMG myc2 plants may reflect the functional redundancy of MYC2 with MYC3 and MYC4 in immunity, which also display rhythmic expression, with slightly delayed acrophases relative to MYC235,60,61. Alternatively, TOC1 may bind directly to the G-box sequence, which is enhanced by inactive MYC2 and inhibited by active MYC2 or co-activators recruited to the promoter following the release of JAZ repression (Fig. 6B). Determining the nature of the interaction between TOC1 and MYC2, and whether this is modulated by infection status, will be critical to understand the molecular basis of the crosstalk between the circadian clock and JA signalling.
This model may seem at odds with the prevailing hypothesis that MYC2 is a negative regulator of immunity against necrotrophic pathogens62. This is based on the reduced susceptibility to B. cinerea and Plectosphaerella cucumerina observed in myc2 mutants and the increased expression of defence marker genes such as PR4 and PDF1.2 following exogenous application of MeJA32,63,64. However, in contrast to the results obtained when applying MeJA, PDF1.2 induction following infection with B. cinerea is actually reduced in myc2 versus Col-0 plants and MYC2 over-expressor (MYC2-ox) plants display enhanced resistance to B. cinerea35,64. Increased resistance to the biotrophic pathogen Pseudomonas syringae has also been reported in both myc2 and MYC2-ox plants65. Together these observations suggest that MYC2 may play a more nuanced role in immunity than is currently appreciated and highlight the need for caution when extrapolating the regulatory function of TFs in immunity from exogenous application of plant hormones.
TOC1 is one of five PSEUDO RESPONSE REGULATOR proteins in Arabidopsis. ChIP-seq analysis has demonstrated that the target genes of PRR5, PRR7 and PRR9 are also enriched for the G-box motif and, like TOC1, formation of a heterotrimeric complex with NF-YB and NF-YC at a CCACA motif within the LHY promoter has been demonstrated in vitro for all three PRR proteins42,66,67,68. Furthermore, protein-protein interactions between TOC1 and other PRRs occur, with the TOC1-PRR3 interaction promoting TOC1 stability by preventing ZEITLUPE-mediated degradation of TOC1, while the TOC1-PRR5 interaction enhances the nuclear accumulation of TOC1, although its effect on TOC1 chromatin binding is unclear58,69,70. The sequential expression of PRR9, PRR7, PRR5 and TOC1 restricts CCA1 and LHY expression to the early morning71. It is conceivable that defence gene expression is also restricted by the sequential expression of the PRR genes and the roles of PRR5, PRR7 and PRR9 in temporal regulation of immunity should be investigated.
In summary, we have demonstrated that the core clock TF TOC1 contributes to the regulation of immunity against B. cinerea in Arabidopsis, occupying the G-box containing region of defence TFs in a pathogen-responsive and MYC2-dependent manner. We suggest that TOC1 occupancy of defence gene promoters contributes to the gating of the plant immune response to necrotrophic pathogens as follows. Oscillation of TOC1 protein levels over a 24 h cycle, peaking at the end of the day helps delimit the phase of defence gene expression. This regulation requires MYC2 which exhibits the same rhythmic expression pattern and acrophase as TOC1, helping to define the window of TOC1-mediated regulation of immunity35. In response to pathogen detection plants must mount an acute immune response that overrides this phased defence gene expression, which is accomplished via a MYC2-dependent release of TOC1 from defence TF promoters. Finally, we note a recent report that flagellin-triggered ROS production and FRK1 expression are suppressed in TOC1-ox plants, which also display enhanced susceptibility to P. syringae DC3000 infection at subjective dawn, indicating that the role of TOC1 in regulating immunity is not restricted to defence against necrotrophic pathogens72.
Methods
Arabidopsis genotypes
All lines used in this study were in the Col-0 background, except for toc1-2 which is in C2410. The TOC1-ox, TMG (TOC1 minigene; pTOC1:TOC1-YFP) and MYC2-YPet (pMYC2:MYC2-YPet-3xHA) and myc2-2 lines have been described previously14,35,36,73. The TMG myc2-2 line was generated by crossing TMG and myc2-2 plants and the MYC2-YPet toc1-2 line by crossing MYC2-YPet and toc1-2 plants. Plants that were homozygous for the YFP transgene and toc1-2 or myc2-2 mutation were identified in the F3 generation and the expression of the TOC1-YFP or MYC2-YPet fusion proteins confirmed by Western blot analysis (Supplementary Fig. 9).
Plant growth conditions
Plants were grown either on a 1:1 mixture of Jiffy 7 peat pellets (Jiffy international AS) and vermiculite, or on half-strength MS plates without sucrose, in controlled growth chambers (PGC-6L, Percival Scientific) and entrained for two to four weeks under long day conditions of 16 h light 8 h dark (80 to 120 μmol m-2 s-1 cool white fluorescent light produced by Philips F17T8/TL81 lamps) at 22 °C74.
Botrytis cinerea infection assays
Botrytis cinerea (pepper isolate) was sub-cultured on peach halves at 25 °C in the dark for 14 d prior to collection of infectious spores. Susceptibility of Arabidopsis to B. cinerea at each time of infection under LD or LL conditions was determined by inoculating detached leaves (with petioles embedded in 1% (w/v) agar) with 10 μL of half-strength grape juice containing 5 × 104 spores mL-175. Necrotic lesions were imaged 72 h post inoculation (hpi) and lesion area determined using ImageJ software (http://rsbweb.nih.gov/ij/).
RNA-Seq analysis
Detached leaves from four-week-old C24 or toc1-2 plants were inoculated with B. cinerea spore suspension (5 × 104 spores mL-1) or half-strength grape juice at ZT0 (six 10 μL drops per leaf) and harvested at ZT18 or ZT22. Total RNA was extracted using Trizol (Thermo Fisher Scientific) and then further purified using the RNEasy cleanup kit (Qiagen), with on-column DNAse treatment. Two biological replicates per plant genotype and time post-infection combination were used in this experiment, each consisting of total RNA pooled from single detached leaves from six different plants.
RNA-Seq libraries were prepared using a TruSeq Stranded mRNA Library Prep kit (Illumina) and 150 bp paired-end reads were generated on the Illumina Novaseq 6000 sequencing platform at Sequentia Biotech (Barcelona, Spain). Sequencing reads were trimmed using Trimmomatic to remove adapter and low quality bases; a minimum quality of 25 and a minimum read length of 35 bp was set. On average 31.4 M reads were produced per sample and only a small fraction was lost during trimming. High quality reads were aligned against the Arabidopsis genome (Araport11) using STAR (version 2.7.9a), with an average mapping rate of 94.3% (range 90.1 to 98.1%).
The raw number of reads uniquely mapped to each transcript was quantified using featureCounts and values normalised using the Trimmed Mean of M values (TMM) normalisation strategy. Removal of genes that were not expressed or that exhibited high variance was performed using the HTSFilter package. The overall quality of the experiment was assessed on the basis of the similarity between replicates by a Principal Component Analysis using the normalized gene expression values as the input. Differentially expressed genes were identified using edgeR, and only genes with a False Discovery Rate (FDR) value < 0.05 (Benjamini-Hochberg correction) were considered statistically significant. To identify the most enriched gene ontology (GO) categories in the differentially expressed gene sets a GO enrichment analysis was performed using PANTHER with default parameters. Significance was determined by Fisher’s exact test followed by Bonferroni adjustment.
Quantitative PCR analysis
Tissue (either single leaves or plates of seedlings) was frozen in liquid nitrogen and stored at -80 °C. Total RNA was extracted using using Trizol (Thermo Fisher Scientific), treated with TURBO DNase (Thermo Fisher Scientific) and cDNA synthesised from 1 μg of RNA using Superscript III (Thermo Fisher Scientific). Quantitative PCR was carried out using the Rotor Gene 6000 (Corbett Life Science). The RealQ Plus 2x Master Mix Green (Ampliqon) was used for all qPCR reactions according to the manufacturer’s guidelines. Each reaction was set up using 5 μL of RealQ Plus 2x Master Mix, forward and reverse primers (Tables 1) and 1 μL of template cDNA. The cycling parameters for qPCR were as follows; an initial step at 95 °C for 15 min, 40 cycles of (95 °C for 30 s, annealing temperature for 30 s, 72 °C for 30 s) and a final step of 72 °C for 1 min. The two-standard curve method was used to calculate relative gene expression76. Expression values were normalized to the geometric mean of the Actin- 2 (AT3G18780) and MON1 (AT2G28390) reference genes. All qPCR reactions were performed with two technical repeats per biological repeat, and repeated if the ΔCt between the two technical repeats was >0.5.
Chromatin immunoprecipitation assays
Two-week-old seedlings grown on MS plates were spray inoculated with B. cinerea spores (5 × 105 spores mL-1) or half-strength grape juice at ZT0 and harvested at ZT18. Two g of tissue were cross-linked in fixation buffer (0.4 M sucrose, 10 mM Tris-HCl pH 8.0, 1 mM EDTA, 1 mM PMSF and 1% (v/v) formaldehyde) at RT under vacuum for 30 min. The cross-linking reaction was quenched with 2 M glycine, added to a final concentration of 0.125 M and vacuum infiltrated for 10 min. The seedlings were ground to a fine powder in liquid nitrogen and cross-linked chromatin complexes were isolated in 25 mL of cold nuclei isolation buffer (0.25 M sucrose, 15 mM piperazine-N,N’-bis 2-ethanesulfonic acid (PIPES) pH 6.8, 5 mM MgCl2, 60 mM KCl, 15 mM NaCl, 1 mM CaCl2, 0.9% (v/v) Triton X-100, 1x Halt Protease Inhibitor Cocktail (ThermoScientific) and 1 mM PMSF). The solution was filtered through two layers of miracloth, incubated on ice for 30 min and then centrifuged at 11,000 x g for 15 min at 4 °C. The pellets were resuspended in 600 μL of cold nuclear lysis buffer (50 mM HEPES pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% (w/v) SDS, 0.1% (w/v) sodium deoxycholate, 1% (v/v) Triton X-100, 1 mM PMSF and 1x Halt Protease Inhibitor Cocktail) and sonicated for 18 min on a cycle of 20 s at maximum power followed by a 40 s rest using a QSonica Q700 sonicator. The sonicated chromatin was then clarified by centrifugation at 4 °C for 10 min at 13,800 x g.
Pre-blocking was performed by incubating 500 μL of protein A/G PLUS beads (Santa Cruz Biotechnology, USA), with 100 μg salmon sperm DNA (Sigma-Aldrich) and 1 mg mL-1 BSA in 2 mL ChIP dilution buffer (16.7 mM Tris-HCl pH 8, 167 mM NaCl, 1.2 mM EDTA, 1.1% (v/v) Triton X-100, 1 mM PMSF and 1x Halt Protease Inhibitor Cocktail) for 1 h at 4 °C with rotation. Thereafter the beads were collected by centrifugation and resuspended in 500 μL of ChIP dilution buffer. For input samples, 30 μg of chromatin was made up to 100 μL with nuclear lysis buffer and diluted with 400 μL of elution buffer (0.5% (w/v) SDS, 0.1 M NaHCO3). For immunoprecipitation, 80 μg of chromatin was made up to a final volume of 100 μL with nuclear lysis buffer and diluted with 900 μL of ChIP dilution buffer. The diluted chromatin was then pre-cleared by adding 30 μL pre-blocked protein A/G PLUS beads, followed by rotation for 1 h at 4 °C to reduce non-specific binding. The pre-cleared chromatin solutions were collected by centrifugation at 15,000 x g for 10 min and immunoprecipitated with 5 μg anti-GFP N-terminal antibody (Sigma-Aldrich #G1544) overnight at 4 °C with rotation. Negative control immunoprecipitations were performed using samples treated in the same manner but without the addition of antibody.
Following overnight immunoprecipitation, an additional 40 μL of pre-blocked protein A/G PLUS beads was added to each sample and incubated for a further 2 h at 4 °C. Immunoprecipitated samples were sequentially washed for 8 min with gentle rotation at 4 °C with 1 mL of the following buffers: once with low salt wash buffer (150 mM NaCl, 0.2% (w/v) SDS, 0.5% (w/v) Triton X-100, 2 mM EDTA, 20 mM Tris-HCl pH 8, 1 mM PMSF and 1x Halt Protease Inhibitor Cocktail) once with high salt wash buffer (as above but with 500 mM NaCl), once with LiCl wash buffer (0.25 M LiCl, 1% (v/v) NP-40, 1% (w/v) sodium deoxycholate, 1 mM EDTA, 10 mM Tris-HCl pH 8, 1 mM PMSF and 1x Halt Protease Inhibitor Cocktail) and then three times with TE wash buffer (10 mM Tris-HCl pH 8, 1 mM EDTA, 1 mM PMSF and 1x Halt Protease Inhibitor Cocktail). The DNA-protein complexes were released in 250 μL of elution buffer (0.5% (w/v) SDS, 0.1 M NaHCO3) by incubation at 65 °C for 15 min with gentle agitation followed by incubation at room temperature for an additional 15 min with rotation. Samples were collected by centrifugation at 4500 x g for 5 min at room temperature. The supernatant was removed and transferred into a new microfuge tube, and the elution step repeated. The two eluates were combined and 20 μL 5 M NaCl was added to the ChIP samples and the 30 μg input samples, followed by incubation at 65 °C overnight to reverse the cross-links. The following day, protein digestion was performed by adding 10 μL of 0.5 M EDTA, 20 μL of 1 M Tris-HCl pH 6.5 and 1 μL of 10 mg mL-1 Proteinase K (New England BioLabs) to each sample followed by incubation at 45 °C for 2 h. DNA from immunoprecipitated and input chromatin was purified by phenol/chloroform extraction.
The amount of target DNA present was quantified by qPCR (see Table 2 for primer sequences) using either 1 μL of input DNA or 2 μL of immunoprecipitated DNA in a final reaction volume of 10 μL. Two technical replicates were performed for each experiment and repeated if the ΔCt between the two technical repeats was >0.5. Enrichment at target promoters was calculated by the percentage input method77.
Western blot analysis
Total protein was extracted by grinding plant leaf tissue in 1 mL of extraction buffer (250 mM sucrose, 50 mM HEPES pH 7.5, 5% (v/v) glycerol, 50 mM NaPP, 1 mM NaMo, 25 mM NaF, 10 mM EDTA, 0.5% (v/v) PVP, 1 mM PMSF, 3 mM DTT, 1x Halt Protease Inhibitor Cocktail). Ten % (w/v) polyacrylamide gels prepared the TGX Stain-Free FastCast Acrylamide Kit (BioRad). Proteins were separated by electrophoresis on a 10% polyacrylamide gel at 110 V for 1 h and equal loading verified by UV-imaging. TOC1-YFP and MYC2-YPet proteins were detected using a 1:2000 dilution of anti-GFP N-terminal antibody (Sigma-Aldrich #G1544) at 4 °C overnight. The membrane was incubated with a 1:80,000 anti-rabbit horseradish peroxidase-conjugated secondary antibody (Sigma-Aldrich #12-348) for 1 h at room temperature. Working solutions of the luminol based Enhanced Chemiluminescent (ECL) solution were prepared and added to the membrane for 1 min78. Protein bands were detected using a ChemiDoc XRS+ system with the Image Lab software (version 5.2).
Statistics and reproducibility
All statistical analyses were performed in Prism (version 10). Prior to two-way ANOVA, data were transformed to ensure homogeneity of variance and normality of error by square root (B. cinerea susceptibility assays) or natural log (qPCR and ChIP-qPCR) transformation. The F-ratio, p-value and effect size (η2) for all factors significant at p < 0.05 are provided in the figure legends or in the supplementary tables. If one or more factors was significant at p < 0.05, a Fisher LSD post-hoc test was used to identify mean values significantly different at p < 0.05. The number of biological replicates in the experimental data presented, and the number of times the experiment was performed, are indicated in the figure legends. Biological replicates were single leaves from independent four-week-old plants for Botrytis cinerea infection assays (Fig. 1) and qPCR analysis of EIN3, EIL1, ORA47, ORA59 and WRKY33 in plants mis-expressing TOC1 (Fig. 3 and Supplementary Fig. 2), and individual MS plates with seedlings for ChIP and qPCR analysis of TMG, TMG myc2, YPet and YPet toc1 plants (Figs. 4 and 5, Supplementary Fig. 5). The RNA-seq experiment was performed with two biological replicates per genotype/infection/time of sampling combination, each consisting of RNA pooled from single detached leaves from six independent plants.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The RNA-seq datasets generated in this work have been deposited in NCBI under accession number PRJNA1270936. Trimmed Mean of M values (TMM)-normalised expression values for each sample in the RNA-seq experiment are provided in Supplementary Data 5. The source data used to generate Figs. 1 to 5 and Supplementary Fig. 2, 3 and 5 are provided in Supplemental Data 6. This study did not develop any custom code. All other data supporting the findings of this study are available from Robert Ingle (Robert.ingle@uct.ac.za) upon reasonable request.
Change history
22 May 2026
A Correction to this paper has been published: https://doi.org/10.1038/s42003-026-10254-4
References
Nozue, K. et al. Rhythmic growth explained by coincidence between internal and external cues. Nature 448, 358–361 (2007).
Bhardwaj, V., Meier, S., Petersen, L. N., Ingle, R. A. & Roden, L. C. Defence responses of Arabidopsis thaliana to infection by Pseudomonas syringae are regulated by the circadian clock. PLOS ONE 6, e26968 (2011).
Ingle, R. A. et al. Jasmonate signalling drives time-of-day differences in susceptibility of Arabidopsis to the fungal pathogen Botrytis cinerea. Plant J. 84, 937–948 (2015).
Wang, W. et al. Timing of plant immune responses by a central circadian regulator. Nature 470, 110–114 (2011).
Zhang, C. et al. LUX ARRHYTHMO mediates crosstalk between the circadian clock and defense in Arabidopsis. Nat. Commun. 10, 2543 (2019).
Zhang, C. et al. Crosstalk between the circadian clock and innate Immunity in Arabidopsis. PLOS Pathog. 9, e1003370 (2013).
Eichmann, R. & Schäfer, P. Growth versus immunity—a redirection of the cell cycle? Curr. Opin. Plant Biol. 26, 106–112 (2015).
Hevia, M. A., Canessa, P., Müller-Esparza, H. & Larrondo, L. F. A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana. Proc. Natl. Acad. Sci. 112, 8744–8749 (2015).
Schaffer, R. et al. The late elongated hypocotyl mutation of Arabidopsis disrupts circadian rhythms and the photoperiodic control of flowering. Cell 93, 1219–1229 (1998).
Strayer, C. et al. Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science 289, 768–771 (2000).
Wang, Z.-Y. & Tobin, E. M. Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 93, 1207–1217 (1998).
Alabadı́, D. et al. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293, 880–883 (2001).
Gendron, J. M. et al. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. 109, 3167–3172 (2012).
Huang, W. et al. Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 336, 75–79 (2012).
Pruneda-Paz, J. L., Breton, G., Para, A. & Kay, S. A. A functional genomics approach reveals CHE as a component of the Arabidopsis circadian clock. Science 323, 1481–1485 (2009).
Jones, J. D. G. & Dangl, J. L. The plant immune system. Nature 444, 323–329 (2006).
Ngou, B. P. M., Ding, P. & Jones, J. D. G. Thirty years of resistance: Zig-zag through the plant immune system. Plant Cell 34, 1447–1478 (2022).
Deslandes, L. & Rivas, S. Catch me if you can: bacterial effectors and plant targets. Trends Plant Sci. 17, 644–655 (2012).
Xiang, T. et al. Pseudomonas syringae effector AvrPto blocks innate immunity by targeting receptor kinases. Curr. Biol. 18, 74–80 (2008).
Chen, J. et al. NLR surveillance of pathogen interference with hormone receptors induces immunity. Nature 613, 145–152 (2023).
Fu, M. et al. A pathogen effector HaRxL10 hijacks the circadian clock component CHE to perturb both plant development and immunity. Nat. Commun. 16, 1538 (2025).
Yuan, M., Ngou, B. P. M., Ding, P. & Xin, X.-F. PTI-ETI crosstalk: an integrative view of plant immunity. Curr. Opin. Plant Biol. 62, 102030 (2021).
Mine, A. et al. The defense phytohormone signaling network enables rapid, high-amplitude transcriptional reprogramming during effector-triggered immunity. Plant Cell 30, 1199–1219 (2018).
Shigenaga, A. M., Berens, M. L., Tsuda, K. & Argueso, C. T. Towards engineering of hormonal crosstalk in plant immunity. Curr. Opini. Plant Biol. 38, 164–172 (2017).
Aerts, N., Pereira Mendes, M. & Van Wees, S. C. M. Multiple levels of crosstalk in hormone networks regulating plant defense. Plant J. 105, 489–504 (2021).
Melotto, M., Underwood, W., Koczan, J., Nomura, K. & He, S. Y. Plant stomata function in innate immunity against bacterial invasion. Cell 126, 969–980 (2006).
Melotto, M., Zhang, L., Oblessuc, P. R. & He, S. Y. Stomatal defense a decade later. Plant Physiol. 174, 561–571 (2017).
Covington, M. F., Maloof, J. N., Straume, M., Kay, S. A. & Harmer, S. L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol. 9, R130 (2008).
Mizuno, T. & Yamashino, T. Comparative transcriptome of diurnally oscillating genes and hormone-responsive genes in Arabidopsis thaliana: Insight into circadian clock-controlled daily responses to common ambient stresses in plants. Plant Cell Physiol. 49, 481–487 (2008).
Goodspeed, D., Chehab, E. W., Min-Venditti, A., Braam, J. & Covington, M. F. Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. 109, 4674–4677 (2012).
Nagel, D. H. et al. Genome-wide identification of CCA1 targets uncovers an expanded clock network in Arabidopsis. Proc. Natl. Acad. Sci. 112, E4802–E4810 (2015).
Shin, J., Heidrich, K., Sanchez-Villarreal, A., Parker, J. E. & Davis, S. J. TIME FOR COFFEE represses accumulation of the MYC2 transcription factor to provide time-of-day regulation of jasmonate signaling in Arabidopsis. Plant Cell 24, 2470–2482 (2012).
Chini, A. et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448, 666–671 (2007).
Dombrecht, B. et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 19, 2225–2245 (2007).
Joseph, R., Odendaal, J. L., Ingle, R. A. & Roden, L. C. The role of the jasmonate signalling transcription factors MYC2/3/4 in circadian clock-mediated regulation of immunity in Arabidopsis. Philos. Trans. R. Soc. B 380, 20230338 (2025).
Más, P., Alabadí, D., Yanovsky, M. J., Oyama, T. & Kay, S. A. Dual role of TOC1 in the control of circadian and photomorphogenic responses in Arabidopsis. Plant Cell 15, 223–236 (2003).
Makino, S., Matsushika, A., Kojima, M., Yamashino, T. & Mizuno, T. The APRR1/TOC1 quintet implicated in circadian rhythms of Arabidopsis thaliana: I. Characterization with APRR1-overexpressing Plants. Plant Cell Physiol. 43, 58–69 (2002).
Yamashino, T. et al. Involvement of Arabidopsis clock-associated pseudo-response regulators in diurnal oscillations of gene expression in the presence of environmental time cues. Plant Cell Physiol. 49, 1839–1850 (2008).
Más, P., Kim, W.-Y., Somers, D. E. & Kay, S. A. Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana. Nature 426, 567–570 (2003).
Windram, O. et al. Arabidopsis defense against Botrytis cinerea: Chronology and regulation deciphered by high-resolution temporal transcriptomic analysis. Plant Cell 24, 3530–3557 (2012).
Graf, A. et al. Parallel analysis of Arabidopsis circadian clock mutants reveals different scales of transcriptome and proteome regulation. Open Biol. 7, 160333 (2017).
Liu, T. L., Newton, L., Liu, M. J., Shiu, S. H. & Farré, E. M. A G-Box-like motif Is necessary for transcriptional regulation by circadian Pseudo-Response Regulators in Arabidopsis. Plant Physiol. 170, 528–539 (2016).
Yoo, S.-D., Cho, Y.-H., Tena, G., Xiong, Y. & Sheen, J. Dual control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signalling. Nature 451, 789–795 (2008).
Wang, H. et al. MED25 connects enhancer–promoter looping and MYC2-dependent activation of jasmonate signalling. Nat. Plants 5, 616–625 (2019).
Bonnot, T. & Nagel, D. H. Time of the day prioritizes the pool of translating mRNAs in response to heat stress. Plant Cell 33, 2164–2182 (2021).
Gao, M. et al. Circadian regulation of the GLYCINE-RICH RNA-BINDING PROTEIN gene by the master clock protein CIRCADIAN CLOCK-ASSOCIATED 1 is important for plant innate immunity. J. Exp. Bot. 74, 991–1003 (2022).
Chao, Q. et al. Activation of the ethylene gas response pathway in Arabidopsis by the nuclear protein ETHYLENE-INSENSITIVE3 and related proteins. Cell 89, 1133–1144 (1997).
Pré, M. et al. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 147, 1347–1357 (2008).
Solano, R., Stepanova, A., Chao, Q. & Ecker, J. R. Nuclear events in ethylene signaling: a transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev. 12, 3703–3714 (1998).
Frerigmann, H., Glawischnig, E. & Gigolashvili, T. The role of MYB34, MYB51 and MYB122 in the regulation of camalexin biosynthesis in Arabidopsis thaliana. Front Plant Sci. 6, 654 (2015).
Moffat, C. S. et al. ERF5 and ERF6 play redundant roles as positive regulators of JA/Et-mediated defense against Botrytis cinerea in Arabidopsis. PLOS One 7, e35995 (2012).
Yang, Z., Tian, L., Latoszek-Green, M., Brown, D. & Wu, K. Arabidopsis ERF4 is a transcriptional repressor capable of modulating ethylene and abscisic acid responses. Plant Mol. Biol. 58, 585–596 (2005).
McGrath, K. C. et al. Repressor- and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 139, 949–959 (2005).
Birkenbihl, R. P., Diezel, C. & Somssich, I. E. Arabidopsis WRKY33 Is a key transcriptional regulator of hormonal and metabolic responses toward Botrytis cinerea Infection. Plant Physiol. 159, 266–285 (2012).
Chen, H.-Y. et al. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. N. Phytol. 211, 599–613 (2016).
Soy, J. et al. Molecular convergence of clock and photosensory pathways through PIF3-TOC1 interaction and co-occupancy of target promoters. Proc. Natl. Acad. Sci. 113, 4870–4875 (2016).
Wasternack, C. & Hause, B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 111, 1021–1058 (2013).
Yan, J. et al. TOC1 clock protein phosphorylation controls complex formation with NF-YB/C to repress hypocotyl growth. EMBO J. 40, e108684 (2021).
Yan, J. et al. TOC1 phosphorylation disproportionally enhances chromatin binding at rhythmic gene promoters. Sci. Adv. 11, eadx7804 (2025).
Chico, J. M. et al. Repression of jasmonate-dependent defenses by shade involves differential regulation of protein stability of MYC transcription factors and their JAZ repressors in Arabidopsis. Plant Cell 26, 1967–1980 (2014).
Song, S. et al. MYC5 is Involved in jasmonate-regulated plant growth, leaf senescence and defense responses. Plant Cell Physiol. 58, 1752–1763 (2017).
Kazan, K. & Manners, J. M. MYC2: the master in action. Mol. Plant 6, 686–703 (2013).
Lorenzo, O., Chico, J. M., Sánchez-Serrano, J. J. & Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 16, 1938–1950 (2004).
Nickstadt, A. et al. The jasmonate-insensitive mutant jin1 shows increased resistance to biotrophic as well as necrotrophic pathogens. Mol. Plant Pathol. 5, 425–434 (2004).
Gautam, J. K., Giri, M. K., Singh, D., Chattopadhyay, S. & Nandi, A. K. MYC2 influences salicylic acid biosynthesis and defense against bacterial pathogens in Arabidopsis thaliana. Physiol. Plant 173, 2248–2261 (2021).
Liu, T., Carlsson, J., Takeuchi, T., Newton, L. & Farré, E. M. Direct regulation of abiotic responses by the Arabidopsis circadian clock component PRR7. Plant J. 76, 101–114 (2013).
Nakamichi, N. et al. Transcriptional repressor PRR5 directly regulates clock-output pathways. Proc. Natl. Acad. Sci. 109, 17123–17128 (2012).
Shen, C. et al. Structural Insight into DNA Recognition by CCT/NF-YB/YC Complexes in Plant Photoperiodic Flowering. Plant Cell 32, 3469–3484 (2020).
Para, A. et al. PRR3 Is a vascular regulator of TOC1 stability in the Arabidopsis circadian clock. Plant Cell 19, 3462–3473 (2007).
Wang, L., Fujiwara, S. & Somers, D. E. PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock. EMBO J. 29, 1903–1915 (2010).
Nakamichi, N. et al. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 22, 594–605 (2010).
Fraser, O. J. P., Spoel, S. H. & van Ooijen, G. TOC1 supresses PAMP-triggered immunity in Arabidopsis. bioRxiv, 2025.2007.2016.665052 (2025).
Gimenez-Ibanez, S. et al. JAZ2 controls stomata dynamics during bacterial invasion. N. Phytol. 213, 1378–1392 (2017).
Carstens, M. et al. Increased resistance to biotrophic pathogens in the Arabidopsis constitutive induced resistance 1 mutant is EDS1 and PAD4-dependent and modulated by environmental temperature. PLOS ONE 9, e109853 (2014).
Ingle, R. A. & Roden, L. C. in Plant Circadian Networks: Methods and Protocols (ed Dorothee Staiger) 273-283 (Springer New York, 2014).
Rasmussen, R. in Rapid Cycle Real-Time PCR: Methods and Applications (eds Stefan Meuer, Carl Wittwer, & Kan-Ichi Nakagawara) 21-34 (Springer Berlin Heidelberg, 2001).
Haring, M. et al. Chromatin immunoprecipitation: optimization, quantitative analysis and data normalization. Plant Methods 3, 11 (2007).
Mruk, D. D. & Cheng, C. Y. Enhanced chemiluminescence (ECL) for routine immunoblotting: An inexpensive alternative to commercially available kits. Spermatogenesis 1, 121–122 (2011).
Acknowledgements
We acknowledge Riccardo Aiese Cigliano (Sequentia Biotech) for his analysis of the RNA-Seq dataset. We also thank Lara Donaldson (ICGEB Cape Town) for C24 seed, Paloma Más (Universitat Autònoma de Barcelona) for toc1-2, TOC1-ox and TMG seeds and Matthew Lewsey (La Trobe University) for MYC2-YPet seed. This work was funded by the National Research Foundation of South Africa (Competitive support for rated researcher grant numbers 105819 and 118504).
Author information
Authors and Affiliations
Contributions
L.C.R. and R.A.I. conceptualised the study and obtained funding. SS performed the experimental work. S.S. and R.A.I. analysed the data. All authors contributed to the design of experiments and to writing of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: David Favero. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sparks, SL., Roden, L.C. & Ingle, R.A. The core clock transcription factor TOC1 binds directly to defence gene promoters regulating immunity in Arabidopsis. Commun Biol 9, 402 (2026). https://doi.org/10.1038/s42003-026-09667-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-026-09667-y
