
Abstract
Mono-ADP-ribosylation, a modification of both proteins and nucleic acids, is implicated in innate immunity. Intracellularly, this modification is catalyzed by PARP enzymes, some induced in response to interferons. Mono-ADP-ribosylation is reversed by hydrolases including proteins with macrodomains, which are conserved across all kingdoms of life. Macrodomains encoded by certain positive-sense single-stranded RNA viruses, such as Chikungunya virus and SARS-CoV-2, antagonize host MARylation to enhance viral replication and suppress the immune response. While macrodomain hydrolase activity is essential for CHIKV replication, in SARS-CoV-2 it predominantly contributes to immune evasion, underscoring viral macrodomains as potential antiviral drug targets. Efforts to develop macrodomain inhibitors include computational modeling, crystallography-based methods, and in vitro assays. However, tools to study macrodomain activity directly in cells remain rare. Here, we established a cell-based assay using PARP15 isoform 1, which we found forms nuclear foci dependent on its ADP-ribosyltransferase activity. Enzymatically active macrodomains dissolve these foci, enabling hydrolase activity monitoring in living cells. Using stable cell lines, this system allows the screening of macrodomain inhibitors while simultaneously addressing cell permeability, toxicity, and physiological relevance. Adaptable to various macrodomains, our platform offers a versatile tool to study macrodomain function in living cells, analyzing mutants, and advancing drug discovery efforts.

Similar content being viewed by others
Introduction
Macrodomains are highly conserved protein domains closely associated with mono-ADP-ribosylation (MARylation), a posttranslational modification of proteins and, recently identified as posttranscriptional modification of RNA1,2,3. Intracellular MARylation is primarily catalyzed by some enzymes of the ADP-ribosyltransferase diphtheria toxin-like (ARTD) family, commonly known as the PARP family1,4. These enzymes consume NAD+ to transfer a single ADP-ribose to a substrate molecule, releasing nicotinamide.
A decade ago, hydrolase activity was attributed to several cellular macrodomain-containing proteins, confirming MARylation as a reversible post-translational modification5,6,7,8. These protein domains are present across all kingdoms of life9,10. In addition, some positive-sense single-stranded RNA ((+)ssRNA) viruses encode macrodomains, including Chikungunya virus (CHIKV)11,12, a member of the Alphavirus genus, and SARS-CoV-213,14, a representative of the Betacoronavirus genus10.
Recent studies, including our own, have identified these viral macrodomains as hydrolases that counteract intracellular MARylation15,16,17,18. For CHIKV, macrodomain hydrolase activity is essential for viral replication. Mutations that disrupt ADP-ribose binding and consequently abolish hydrolase activity result in a replication-deficient virus17,19. One function of the CHIKV macrodomain is the reactivation of the essential viral protease, which is inhibited by intracellular PARP enzymes19. In contrast, while the hydrolase activity of the SARS-CoV-2 macrodomain 1 has little effect on viral replication, it significantly affects the immunomodulatory functions of the macrodomain. Specifically, a hydrolase-lacking SARS-CoV-2 permits a robust antiviral immune response, promoting viral clearance. Conversely, an active SARS-CoV-2 macrodomain contributes to viral pathogenesis by suppressing the innate immune response20. These findings establish viral macrodomains as promising antiviral drug targets21,22.
The findings summarized above, together with the recognition that various PARP family members are linked to innate immunity23, and the SARS-CoV-2 pandemic, suggest that viral macrodomains represent potential therapeutic drug targets. Therefore, substantial interest has been dedicated to screening for compounds that inhibit viral macrodomain activity. Recent efforts included in silico modeling, fragments screening, crystallography-based approaches, and in vitro screening techniques, focusing on identifying small molecules that bind in the catalytic pocket of the macrodomain24,25,26,27,28,29,30. Furthermore, novel assays have been developed to monitor macrodomain hydrolase activity in vitro31,32,33,34.
In this study, we developed an assay to investigate the hydrolase activity of macrodomains in cellulo. While studying PARP15 in the context of stress granule biology, a suggestion based on a previous report35, we noticed that GFP-tagged PARP15 isoform 1 (PARP15.1) formed nuclear foci. These foci were unrelated to stress granules. They appeared to be dependent on the ADP-ribosyltransferase activity of PARP15.1. This led to the question whether PARP15.1 foci formation can be used to measure MAR hydrolase activity in cell nuclei. Indeed, we found that PARP15.1 foci dissolve in response to catalytically active hydrolases. Building on these findings, we established stable cell lines enabling us to monitor macrodomain hydrolase activity and, consequently, the screening for small molecule inhibitors directly in living cells. Furthermore, the cell lines provide a platform to study macrodomain mutants and assess compounds that target PARP15 directly. Together, the established system is easily adaptable to different macrodomains and offers the advantage of simultaneously addressing compound toxicity, cell permeability, target engagement, and activity in a physiologically relevant setting.
Results
PARP15 isoform 1 forms nuclear foci dependent on its catalytic activity
To study the subcellular localization of PARP15.1, doxycycline-inducible lentiviral vectors were generated that express GFP-tagged PARP15.1 or a catalytically inactive mutant, PARP15.1-H559Y. These proteins were introduced into U2OS cells, which express endogenously tagged tdTomato-G3BP136. The loss of enzymatic activity of the H559Y mutant was confirmed by in vitro ADP-ribosylation assays comparing the automodification activities of the bacterially expressed catalytic domains of PARP15 (PARP15cat) and PARP15cat-H559Y (Fig. 1a).

a In vitro ADP-ribosylation assay. Recombinant His6-PARP15cat or His6-PARP15cat-H559Y proteins were incubated with radioactively labeled 32P-NAD+ for 30 min at 30 °C. Proteins were separated by SDS-PAGE and visualized by Coomassie blue (CB) staining. The incorporated radioactive label was visualized by exposure to X-ray films (32P). b The expression of PARP15.1 fusion proteins in U2OS-tdTomato-G3BP1/GFP-PARP15 or U2OS-tdTomato-G3BP1/GFP-PARP15-H559Y was induced by increasing doxycycline concentrations as indicated. Protein expression was evaluated by immunoblotting 24 h post-induction. PARP15 fusion proteins were visualized using a GFP antibody. γ-Tubulin was detected as loading control. c U2OS-tdTomato-G3BP1/GFP-PARP15 or U2OS-tdTomato-G3BP1-GFP-PARP15-H559Y were treated with 0.5 µg/ml doxycycline for 16 h to induce expression of GFP-PARP15.1 variants. Cells were fixed, and PARP15.1 localization investigated by confocal microscopy. PARP15.1 and its catalytically inactive mutant (H559Y) are stained in green, G3BP1 in magenta. Nuclei were visualized by Hoechst staining (blue). Scale bar, 10 µM.
The expression of GFP-PARP15.1 fusion proteins was evaluated following overnight treatment with varying doxycycline concentrations, as assessed by immunoblotting (Fig. 1b). Both GFP-PARP15.1 and GFP-PARP15.1-H559Y showed dose-dependent expression in response to doxycycline, which was used to determine the optimal concentration of 0.5 µg/ml for subsequent subcellular localization experiments. The parental U2OS-tdTomato-G3BP1 cell line allowed for a straightforward visualization of stress granules because G3BP1 is a key factor for stress granule formation36,37. Therefore, we compared the localization of tdTomato-G3BP1 and GFP-PARP15.1 using confocal microscopy (Fig. 1c). GFP-PARP15.1-expressing cells were fixed and stained with Hoechst to visualize nuclei. PARP15.1 displayed exclusive nuclear localization, whereas G3BP1 was cytoplasmic as expected. Moreover, wildtype GFP-PARP15.1 appeared in small dot-like structures, which we refer to as foci in the following. In contrast, the catalytically inactive H559Y mutant exhibited a diffuse nuclear distribution (Fig. 1c). Recent studies have linked ADP-ribosylation to the regulation of biomolecular condensates38. Based on these initial findings, we hypothesized that PARP15 catalytic activity similarly regulates its own nuclear foci formation.
Establishment of an assay that allows for studying PARP15 activity and the hydrolase activity of viral macrodomains in cells
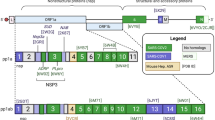
Based on these initial findings, we tested whether co-expression of hydrolytically active macrodomains was able to dissolve GFP-PARP15.1 foci in the nucleus (as schematically depicted in Fig. 2a). We established a system allowing for the specific recruitment of a GFP-targeting nanobody-macrodomain fusion protein to mEGFP-PARP15.119. To achieve equimolar and controllable expression of GFP-PARP15.1 and the 3xFLAG-NLS-nanobodyGFP (NBGFP) fusion proteins, the two parts were linked with a T2A site. Furthermore, the sequence coding for the NBGFP was followed by a Gateway cassette integrated into a pcDNA5/FRT/TO plasmid (Fig. 2b). This construct allows for an easy recombination of any macrodomain or respective mutant of interest into the plasmid using Gateway cloning. Further, it can be used to generate stable doxycycline-inducible Flp-In T-REx cells, e.g., in HeLa, HEK293 or U2OS cells39.

a Schematic representation of the assay illustrating foci formation of MARylated PARP15.1, which is reversed upon hydrolase activity of a macrodomain (created in BioRender. Verheugd, P. (2026) https://BioRender.com/zm1a7k2). b Schematic representation of constructs designed to generate cell lines stably expressing the indicated fusion proteins. The integration of a T2A site between GFP-PARP15 and the 3xFLAG-NLS-NBGFP (a GFP-specific nanobody) enables the simultaneous expression of both fusion proteins in equimolar ratios. Additionally, the inclusion of a Gateway cassette downstream of the sequence encoding the NBGFP facilitates recombination of various macrodomains or their respective mutants for analysis. GFP green fluorescent protein, MD macrodomain, NLS nuclear localization signal, NB nanobody. c The indicated constructs were transiently expressed in HEK293 cells. Protein expression was analyzed by immunoblotting. GFP-PARP15.1 was detected using a specific PARP15 antibody, 3x-FLAG-NLS-NBGFP fusion proteins were visualized by an anti-FLAG antibody. γ-Tubulin served as loading control. d U2OS cells were transiently transfected with constructs as shown in (b) expressing GFP-PARP15.1 and the indicated NBGFP, either without the MD, with the SARS-CoV-2 MD, the CHIKV MD, or its catalytically inactive mutant (V33E). Cells were fixed, nuclei visualized by Hoechst staining (blue) and localization of GFP-PARP15.1 (green) and 3xFLAG-NLS-NBGFP variants (red) determined by confocal microscopy.
We initially tested these constructs for expression and functionality (Fig. 2c, d). Therefore, we transiently transfected constructs that express GFP-PARP15.1-T2A-NBGFP-macrodomain fusions of SARS-CoV-2 and CHIKV and mutants thereof in HEK293 cells. Protein expression was analyzed by immunoblotting (Fig. 2c). From all constructs analyzed in this study, GFP-PARP15.1 or GFP-PARP15.1-KR (a mutant in which two ubiquitin acceptor lysines were changed to arginine, see below) as well as the respective macrodomain fusions (Chikungunya virus macrodomain (CHIKV MD) and the catalytically inactive mutant CHIKV MD V33E (VE)) or SARS-CoV-2 macrodomain 1 (CoV2 MD) along with mutants N40R, G130D, F132V, D22V, G48V, or G130V) were expressed as individual proteins, confirming the functionality of the T2A site (Fig. 2c).
To visualize the recruitment of the 3xFLAG-NLS-NBGFP-macrodomains to GFP-PARP15.1, U2OS cells were transiently transfected with the respective constructs, and the localization of the two fusion proteins was analyzed by confocal microscopy (Fig. 2d). In the control experiment without expression of a macrodomain, GFP-PARP15.1 accumulated in nuclear foci, corroborating our initial observations. Furthermore, signals from the NBGFP stained with a FLAG-selective antibody and from the fluorescence of the GFP-PARP15.1 fusion protein overlapped, demonstrating that the 3x-FLAG-NLS-NBGFP was efficiently recruited to the GFP-tagged fusion protein (Fig. 2d).
As hypothesized, fusion of the viral macrodomain from either SARS-CoV-2 or CHIKV to 3xFLAG-NLS-NBGFP resulted in a uniform nuclear distribution of GFP-PARP15.1, similar to the pattern observed with the catalytically inactive PARP15.1-H559Y mutant (Fig. 1c). In contrast, when the catalytically inactive CHIKV macrodomain variant (V33E), which shows very limited deMARylation activity16, was fused to the NBGFP, PARP15.1 retained its accumulation in nuclear foci (Fig. 2d).
In summary, our findings demonstrate that the accumulation of GFP-PARP15.1 in nuclear foci is MARylation-dependent. Recruitment of hydrolytically active viral macrodomains effectively disrupts foci formation, leading to a uniform nuclear distribution of GFP-PARP15.1, consistent with reversing MARylation.
PARP15 inhibitors dissolve GFP-PARP15.1 foci
Based on our initial findings, we utilized pcDNA5/FRT/TO plasmids encoding GFP-PARP15.1-T2A-3xFLAG-NLS-NBGFP, either fused or not to a viral macrodomain, to generate stable HeLa Flp-In TRex cells39. These cells enable doxycycline-dependent induction of the fusion proteins. To evaluate their potential for future screenings, we tested PARP15.1 foci formation and its dependence on MARylation activity by applying the recently developed PARP15 inhibitors OUL232 and OUL243 (Figs. 3a, b, and S1)40. HeLa Flp-In TRex cells expressing GFP-PARP15.1-T2A-3xFLAG-NLS-NBGFP were seeded for live-cell imaging. Protein expression was induced, and cells were either left untreated or treated with DMSO (vehicle control), OUL232 or OUL243. Nuclei were stained with SpyDNA, and GFP-PARP15.1 localization was subsequently analyzed via live-cell confocal microscope (Figs. 3b and S1). In untreated or DMSO-treated control cells, GFP-PARP15.1 exhibited nuclear foci. However, treatment with OUL232, which inhibits PARP15 MARylation activity, disrupted foci formation, resulting in a uniform nuclear distribution of GFP-PARP15.1.

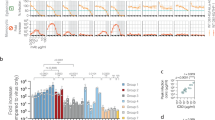
a Schematic representation of the assay when used to evaluate small molecule inhibitors targeting PARP15 (created in BioRender. Verheugd, P. (2026) https://BioRender.com/a35t588). MARylation-dependent PARP15.1 foci are resolved upon addition of OUL232 or OUL243, resulting in a shift from foci formation to a uniform, spread signal distribution. b–e HeLa-Flp-In TRex-GFP-PARP15.1-T2A-3xFLAG-NBGFP cells were treated with 1 µg/ml doxycycline to induce protein expression. Additionally, the cells were treated or not with the indicated amounts of compounds OUL232 or OUL243 overnight. DMSO served as the vehicle control. One hour prior to analysis by confocal microscopy, SpyDNA was added to stain nuclei. Live-cell imaging was performed using confocal microscopy. b Representative confocal microscopy images showing GFP-PARP15.1 (magenta), SpyDNA (blue) or the merged images. c Schematic representation of the established CellProfiler pipeline. d, e Images obtained from confocal microscopy were analyzed with CellProfiler. d Data were plotted with ggplot in R to indicate percentages of cells showing either nuclear foci (blue), uniformly distributed, spread signal (black), or a combination of both (yellow) for the indicated treatments. e Boxplot overlaid with individual values illustrating the number of foci per nucleus for the indicated treatments, differentiating between foci only (blue), spread signal (black) or a combination of both (yellow). Outliers were removed by the IQR method and p-values were calculated using pairwise Wilcoxon test, adjusted by false discovery rate applying the rstatix package in R. (**p ≤ 0.01; ****p ≤ 0.0001; ns, not significant; of 2–5 biological replicates, a total of 182 (control (n = 4)); 103 and 56 (DMSO 0.05% (n = 3) and 0.1% (n = 2), respectively); 77, 95 and 147 (OUL232 1.25 µM (n = 3), 2.5 µM (n = 4) and 5 µM (n = 5), respectively); 103 and 59 (OUL243 5 µM (n = 3) and 10 µM (n = 2), respectively) nuclei with GFP signal were evaluated).
To quantitatively assess foci formation versus uniform nuclear distribution as well as the numbers of foci per nucleus, we established an analysis pipeline using CellProfiler41. This pipeline enabled the automated analysis of the obtained data from microscopic analysis (Fig. 3c). Applying this pipeline to images from this inhibitor study revealed the proportions of cells displaying uniformly distributed GFP-PARP15.1 (“spread”), distinct nuclear foci, or a combination of both patterns (Fig. 3d, e, Supplementary Data 1 and 2). In control samples (untreated or DMSO-treated), the majority of cells displayed either PARP15.1 nuclear foci or a combination of foci and spread signals, with only a small fraction of cells exhibiting uniformly distributed PARP15.1. Treatment with OUL232 resulted in a dose-dependent increase of the proportion of cells with spread GFP-PARP15.1 localization, while the proportion of cells exhibiting foci or both localization patterns decreased. Similarly, treatment with OUL243 also reduced the number of cells displaying PARP15.1 foci and increased the proportion of cells with uniformly distributed PARP15.1, albeit OUL243 having a less potent response than OUL232, consistent with the reduced potency of the former inhibiting PARP15 catalytic activity in vitro40 (Fig. 3d, e). This trend was further reflected in the number of GFP-PARP15.1 foci per nucleus. Even at concentrations of 1.25 µM, OUL232 significantly reduced the number of nuclear foci, with complete ablation observed at 5 µM. In contrast, OUL243 required a concentration of 10 µM to significantly reduce foci formation (Fig. 3e).
In summary, these findings demonstrate that the stable HeLa Flp-In TRex cells expressing GFP-PARP15.1-T2A-3xFLAG-NLS-NBGFP form nuclear foci in a MARylation-dependent manner. Thus, this assay represents a reliable tool for screening and studying potential inhibitory compounds targeting PARP15 catalytic activity in cells.
In silico analysis of the SARS-CoV-2 macrodomain identifies catalytically important amino acids
To identify amino acids critical for ADP-ribose binding and thus potentially hydrolase activity, an in silico alanine scan was conducted on the SARS-CoV-2 macrodomain guided by the structure of an with ADP-ribose X-ray structure (PDB ID: 7KQP)24. This computational technique systematically replaces amino acid residues in a protein, for example, at the ligand binding site interface, with alanine to evaluate their contributions to binding energy42. Alanine was selected due to its small size, nonpolar nature, and lack of reactive side chain groups, making it a good choice to assess the energetic contributions of side chain interactions. The alanine scan identified four key residues predicted to be critical for ADP-ribose binding, namely N40, V49, F132, and F156 (Fig. 4a). In addition, residues D22, G48, and G130 were included in the analysis due to their high conservation across viral macrodomains. Interestingly, the disruption of the corresponding positions N40 (CHIKV N24), V49 (CHIKV V33) and F132 (CHIKV Y114) in the CHIKV macrodomain has been shown to impair hydrolase activity16.

a In silico alanine scan docking scores for all residues framing the binding site. The docking scores refer to AutoDock 4.1. b Computational introduction of point mutations at the seven selected sites and re-docking of ADP-ribose. The docking scores refer to the Schrodinger 2022.1 standard protocol glide score. c RMSD of the SARS-CoV-2 macrodomain in comparison to the three indicated mutants. d RMSF of the in (c) indicated macrodomain variants.
Subsequently, each of the identified residues was mutated to amino acids other than the wild-type residue, and the impact of each substitution was assessed by re-docking ADP-ribose to the macrodomain mutants. The docking scores of the ADP-ribose in complex with macrodomain mutants were then compared to those in complex with the wildtype macrodomain (Fig. 4b). This procedure enabled the identification of the most disruptive mutants for each critical position identified in the alanine scan (Fig. 4a). Based on the results of the in silico mutational analyses, multiple sequence analyses and previous mutation studies of the CHIKV macrodomain, three specific mutants, N40R, G130D, and F132V were selected for experimental validation and further computational studies.
The structural stability of these mutants was subsequently evaluated using molecular dynamics simulations to predict potential local unfolding events. All mutants demonstrated stable conformations with backbone RMSD (root mean square deviation) values averaging 1.7 Å (Fig. 4c). Additionally, the secondary structures of all macrodomain mutants were predicted to be preserved. However, RMSF (root mean square fluctuation) analysis for the F132V mutant revealed slight conformational changes in the side chains of residues within the ADP-ribose binding site (Fig. 4d).
A detailed analysis of the impact of each substitution on ADP-ribose binding, compared to the wild-type macrodomain, revealed minor alterations in the ADP-ribose binding pocket (Fig. 5a). While the adenosine subpocket appeared unaltered, mutant-dependent variations were observed in the diphosphate and distal ADP-ribose subpockets (Fig. 5b–d). Across all three mutants, ADP-ribose consistently formed two hydrogen bonds in the adenosine binding subpocket: one between D22 and the primary amine of the adenine moiety, and the other between I23 and the adenine ring, comparable to the wildtype macrodomain (Fig. 5b–d). However, in the diphosphate and the distal ribose subpockets, the interaction patterns are different among the mutants.

a Macrodomain wildtype in complex with ADP-ribose (sites for substitution are indicated in magenta). Lower panel shows the protein–ligand interaction diagram. b–d 3D representation of the indicated mutants in complex with ADPr and 2D representation of the protein-ligand interactions to the right site. b N40R mutant c F132V mutant d G130D mutant.
In detail, in the diphosphate subpocket, the number of contacts varied among mutants. The N40R and F132V mutants displayed interaction patterns similar to the wild-type macrodomain, with ADP-ribose forming hydrogen bonds with the backbone of V49, S128, G130, I131, and F132 (or V132 in the case of the F132V mutant) (Fig. 5b, c). In contrast, the G130D mutant maintained a comparable number of hydrogen bonds to the wildtype, but these involved interactions with V49, S128, F132, D130, D50, L126, and G49 (Fig. 5d). Considering the distal ADP-ribose subpocket for the F132V and G130D mutants, ADP-ribose formed hydrogen bonds with N40 and G46 as in the wildtype, while the N40R mutant exhibited hydrogen bonding exclusively through G46 (Fig. 5a–d). Concluding from the in silico interaction analyses, the N40R, G130D, and F132V mutants are likely to reduce ADP-ribose binding without affecting the structural integrity of the mutant macrodomains.
Evaluation of viral macrodomain mutants in cells
Next, we sought to use the developed in cellulo assay to evaluate the potential hydrolase deficiencies of the SARS-CoV-2 macrodomain mutants, which were expressed to comparable levels (Fig. 2c), as predicted from the previous in silico analysis (Figs. 4 and 5). Therefore, we employed HeLa Flp-In T-REx cells expressing GFP-PARP15.1-T2A-3xFLAG-NLS-NBGFP-SARS-CoV-2 macrodomain or its mutant variants (N40R, G130D, and F132V) to analyze GFP-PARP15.1 foci formation (Fig. 6a). As shown before in our transient assays, GFP-PARP15.1 formed nuclear foci in the absence of a macrodomain, which were dissolved upon co-expression of the wild-type SARS-CoV-2 macrodomain (Fig. 6b, c). Although the G130D substitution in the catalytic cleft of the macrodomain was predicted to cause steric hindrance of ADP-ribose binding and was confirmed as inactive in in vitro deMARylation assays10, co-expression of SARS-CoV-2 macrodomain G130D resulted in spread GFP-PARP15.1 signals. Thus, this mutant was active in our cellular assay, unlike what was predicted from the in silico analysis (Figs. 4 and 5) and in vitro assays10.

a Schematic representation of the assay when used to evaluate macrodomain mutants (created in BioRender. Verheugd, P. (2026) https://BioRender.com/j53d353). MARylation-dependent PARP15.1 foci are resolved upon recruitment of a hydrolytically active macrodomains, resulting in a shift from foci formation to a uniform, spread signal distribution. In contrast, mutations that disrupt hydrolase activity (viral macrodomain mutants, such as CHIKV MD V33E or SARS-CoV-2 MD F132V, G130D, N40R, see b, c) preserve PARP15.1 foci formation. b Stable HeLa Flp-In T-REx cells were induced for expression of GFP-PARP15.1 and the different 3xFLAG-NBGFP-macrodomain fusion proteins by 1 µg/ml doxycycline overnight. One hour prior to live-cell imaging using confocal microscopy, nuclei were stained using SpyDNA. Representative images of each cell line are shown with GFP-PARP15.1 in magenta, SpyDNA staining in blue and merged pictures. c Images obtained from confocal microscopy were analyzed using the CellProfiler pipeline. The data were plotted with ggplot in R to indicate percentages of cells showing either nuclear foci (blue), uniformly distributed, spread signal (black), or both (yellow) in the presence of the respective macrodomain variant.
Additionally, we tested the N40R and F132V SARS-CoV-2 macrodomain mutants that were predicted to lose hydrolytic activity in our in silico analyses (Figs. 4 and 5). Co-expression of either of the two SARS-CoV-2 macrodomain mutants resulted in the retention of PARP15.1 nuclear foci, consistent with a loss of hydrolytic activity (Fig. 6b, c). As controls, we included the CHIKV macrodomain and its catalytically inactive mutant, V33E. As expected and analogous to the SARS-CoV-2 macrodomain, the CHIKV macrodomain disrupted GFP-PARP15.1 foci, leading to a uniform nuclear distribution of the GFP signal. Co-expression of the CHIKV V33E macrodomain mutant, however, allowed GFP-PARP15.1 to persist in nuclear foci (Fig. 6b, c). These findings are consistent with previous biochemical and in cellulo analyses showing that the V33E substitution in the catalytic cleft of the CHIKV macrodomain results in a loss of hydrolase activity16 and does not support efficient viral replication19. The observed localization and distribution patterns of GFP-PARP15.1 were further validated through quantitative analysis using our CellProfiler pipeline (Fig. 6c, Supplementary Data 1 and 3).
In conclusion, our assay not only provides a robust platform for testing potential inhibitors targeting PARP15 activity but also facilitates the in cellulo evaluation of hydrolase-deficient mutants by assessing the disappearance or persistence of PARP15.1 nuclear foci.
Evaluation of recently published potential SARS-CoV-2 macrodomain inhibitors in cells
Our assay provides the opportunity to screen for compounds targeting the individual viral macrodomain in cellulo (Fig. 7a). Based on our findings described above, we expected that inhibition of the macrodomain prevents loss of MARylation and thus results in reappearance of PARP15.1 nuclear foci. To evaluate this, we tested compounds that have been reported to target the SARS-CoV-2 macrodomain in vitro25,43,44. However, none of the tested compounds showed effects on the PARP15.1 localization, suggesting that they were not active in our in cellulo assay (Fig. 7b, c, Supplementary Data 1 and 4). Thus, although these compounds were predicted to inhibit the macrodomain or were found to interfere with SARS-CoV-2 replication, possibly through inhibiting the macrodomain, they were nonfunctional in our cell-based assay despite using high concentration of the compounds.

a Schematic representation of the assay when used to investigate the inhibitory potential of small molecular compounds targeting the SARS-CoV-2 macrodomain (created in BioRender. Verheugd, P. (2026) https://BioRender.com/q91u685). Recruitment of a hydrolytically active macrodomain results in resolution of PARP15.1 foci. Inhibitory compounds targeting the macrodomain preserve PARP15.1 foci formation. b, c HeLa Flp-In T-REx cells were treated with 1 µg/ml doxycycline to induce the expression of GFP-PARP15.1-T2A-3xFLAG-NBGFP-SARS-CoV-2 macrodomain. The cells were treated with the indicated inhibitors overnight. One hour prior to evaluation by confocal microscopy, nuclei were stained using SpyDNA. b Representative images from confocal microscopy are shown with GFP-PARP15.1 (magenta), SpyDNA (blue), or the merged picture. c Images from confocal microscopy were evaluated with the CellProfiler Pipeline and data plotted with ggplot in R. The percentage of cells showing foci (blue), spread signal (black), or both (yellow) is illustrated upon the indicated treatments. For control the 3xFLAG-nanobody construct alone, untreated cells, or DMSO vehicle was included.
Evolving the in cellulo assay by employing PARP15-KR, a more stable PARP15 mutant
When inducing protein expression in our stable cell lines in parallel, we observed that protein levels of PARP15.1 wildtype and its catalytically inactive PARP15.1-HY mutant varied considerably when analyzed in parallel (Fig. 8a). While PARP15.1 wildtype protein was barely detectable on Immunoblots, the PARP15.1-HY mutant exhibited robust protein expression. Inhibiting PARP15 catalytic activity with OUL232 increased PARP15.1 protein abundance, suggesting that PARP15 is destabilized in response to its MARylation function (Fig. 8a). In support, OUL232 did not further stabilize PARP15.1-HY (Fig. 8a). Because protein turnover is often mediated by ubiquitination, we substituted lysine residues at positions 128 and 132, which are unique to isoform 1 and are known as ubiquitin acceptor sites (according to PhophoSitePlus), with arginine. This PARP15.1-K128/135R (PARP15-KR) mutant was indeed more stable than the wild-type protein, comparable to the catalytically inactive mutant (Figs. 8a and S2a). Moreover, PARP15.1-KR formed foci efficiently, which appeared somewhat larger and which is consistent with preserved MARylation activity (Fig. 8b).

a Stable U2OS Flp-In TREx cells were induced by 0.5 µg/ml doxycycline for the expression of GFP-PARP15.1 (wt, -HY, -KR). The cells were treated subsequently with DMSO or OUL232 as indicated. Cell lysates were separated and analyzed for protein abundance via immunoblotting. b Representative images from confocal microscopy indicating the localization pattern of GFP-PARP15.1 and mutant variants (magenta) in the stable U2OS Flp-In TREx upon doxycycline-induced expression. Nuclei were stained using SpyDNA, applied one hour prior to live-cell imaging (blue). c Stable HeLa Flp-In T-REx cells were induced for expression of GFP-PARP15.1-KR and the indicated 3xFLAG-NBGFP-macrodomain fusion proteins by 1 µg/ml doxycycline overnight. One hour prior to live-cell imaging by confocal microscopy, nuclei were stained using SpyDNA. Representative images of each cell line are shown with GFP-PARP15.1-KR (magenta), SpyDNA (blue) and merged. d Images obtained from confocal microscopy were analyzed using the CellProfiler pipeline. The data were plotted with ggplot in R to indicate percentages of cells showing either nuclear foci (blue), uniformly distributed, spread signal (black), or both (yellow) in the presence of the respective macrodomain variant. e Stable HeLa Flp-In T-REx cells were induced for expression of GFP-PARP15.1-KR and the SARS-CoV2 MD wt and subsequently left untreated, treated with DMSO or the indicated inhibitors overnight. Representative images of each condition are shown with GFP-PARP15.1-KR (magenta), SpyDNA (blue) and merged. f As in (d), but here upon treatment with the indicated inhibitors.
Next, we expanded the repertoire of SARS-CoV2 MD mutants by introducing D22V, G48V, and G130V variants10, which we tested in the PARP15.1-KR system due to its higher frequency of foci formation compared with the wild-type protein (Figs. 8b–d and S2b). Expression of the SARS-CoV2-MD wildtype, consistent with our previous findings, disrupted PARP15.1-KR foci and resulted in a uniform nuclear signal (Fig. 8c). In contrast, co-expression with the SARS-CoV2 MD D22V or G130V variants led to persistence of PARP15.1-KR foci, indicating that these mutants are hydrolytically inactive, in agreement with published data10. Though the G48V mutant affected foci formation to a lesser extend, the proportion of cells with both foci and spread signal still increased (Fig. 8c, d, Supplementary Data 1 and 5).
Finally, we tested a set of recently published MD inhibitors in the improved assay employing PARP15.1-KR (Fig. 8e, f)30. Treatment with compounds 5a, 5c, or 6b resulted in minor effects on the nuclear distribution of the GFP-PARP15.1-KR signal, comparable to the control treated with DMSO, suggesting that these compounds are inactive against the SARS-CoV2 MD in our in cellulo assay. In contrast, treatment with compounds 6d and 6e restored partially PARP15.1-KR foci formation, indicating that these inhibitors engage their target in cells (Fig. 8e, f, Supplementary Data 1 and 6), consistent with recent reports30.
In summary, the establishment of the PARP15.1-KR mutant in our in cellulo assay provides a more robust and reliable readout of foci formation versus diffuse nuclear signal compared to the assay including the PARP15 wildtype protein. This enhanced system enabled us to characterize additional SARS-CoV2 MD variants and, importantly, to assess the inhibitory potential of newly reported MD inhibitors, supporting the suitability of this assay as a screening platform for compounds targeting viral macrodomains.
Discussion
With the onset of the SARS-CoV-2 pandemic, viral macrodomains have garnered attention as potential therapeutic targets due to their roles in immunomodulation and viral replication21,22. Numerous studies aimed at the development of assays that facilitate the identification of small molecule inhibitors targeting the SARS-CoV-2 macrodomain24,25,26,27,29,32,33,34,40,43,45,46,47. Most of these approaches focused on identifying molecules that bind to the ADPr binding pocket of the SARS-CoV-2 macrodomain24,25,26,27,29,43,45,46. For example, Schuller et al. used fragment-based crystallography to visualize high-resolution binding modes of diverse fragments and explore the chemical binding space of the SARS-CoV-2 macrodomain24,46. Promising candidates were further investigated by biophysical techniques, such as homogenous time-resolved fluorescence (HTRF) assay, differential scanning fluorimetry (DSF), or isothermal titration calorimetry (ITC), all of which measure binding of a ligand to a macrodomain24,46. Brosey et al. analyzed previously developed poly-ADP-ribose glycohydrolase inhibitor (PARGi) fragments to assess their interaction with the SARS-CoV-2 ADPr binding pocket, providing additional structural insights27. Similarly, Ni et al. determined by co-crystallization that the remdesivir analog GS441524 binds to the SARS-CoV-2 macrodomain’s ADPr pocket, which was confirmed by ITC studies43.
Other studies have employed displacement assays using ADP-ribosylated peptide tracers to measure competitive binding of potential inhibitors. While these assays effectively evaluate binding, they are unable to assess the hydrolytic activity of the macrodomain due to the nature of the tracer peptide. Moreover, the binding affinities of these tracers can vary significantly across different macrodomains, limiting their applicability to a broad spectrum of macrodomains25,26,29,45,47,48. For instance, Gahbauer et al. performed in silico design of merged fragments and, as a second approach, large library docking to identify potential SARS-CoV-2 macrodomain inhibitors. These candidates were subsequently evaluated using ITC, DSF, and HTRF assays, followed by structural studies via crystallographic soaking experiments. However, these compounds with promising binding profiles displayed poor cell permeability. Also, effects on viral pathogenicity or immunomodulation remain unexplored under physiological conditions. This highlights not only the potential limitations of the identified molecules but also the need for suitable cell-based assays to evaluate these leads in more biologically relevant settings25.
Recent advances have focused on the development of assays, which directly assess the enzymatic activity of macrodomains.32,34,47. The so-called ADPr-Glo assay, a luminescence-based assay, measures the de-MARylation activity of macrodomains by quantifying free ADP-ribose. This is subsequently converted to AMP via the ADP-ribose pyrophosphatase NudF. AMP levels are then quantified using a commercially available AMP-Glo assay. While effective, this method involves multiple enzymatic steps requiring extensive controls and countermeasures32. A more streamlined approach developed by Ildefeld et al. leverages HTRF to monitor macrodomain hydrolase activity34. This assay uses an auto-MARylated PARP10 catalytic domain coupled to a terbium-cryptate as a substrate and exploits subsequent specific interaction with GFP-labeled PARP14 macrodomains 2 and 334,49,50. In all these in vitro assays, parameters are highly controlled with only limited reaction partners, yet they fail to address critical properties, such as cell permeability, cytotoxicity, and antiviral or immunomodulatory effects, all of which are essential for evaluating therapeutic potential in physiologically relevant conditions. The lack of robust cell-based assays remains a significant barrier to fully characterize the therapeutic efficacy of potential macrodomain inhibitors.
One notable advance toward cellular studies was made by Russo et al., who developed a method to assess macrodomain hydrolase activity in cells. They observed that treatment with type I or II interferons increased ADP-ribose fluorescence intensity in a PARP9/DTX3L-dependent manner, which can be modulated by expression of the SARS-CoV-2 macrodomain. While promising, this approach relies on antibody staining of fixed cells prior to confocal microscopy-based analysis, which is time-consuming and likely limits its throughput and scalability33.
In the current study, we describe a novel in cellulo assay for evaluating hydrolase activity of both cellular and viral macrodomains, including the SARS-CoV-2 and the CHIKV macrodomains. This assay exploits the observation that MARylated PARP15.1 forms nuclear foci, a pattern disrupted by hydrolytically active macrodomains (Figs. 1–3). This phenomenon can be visualized in live cells, providing a straightforward, physiologically relevant readout for macrodomain hydrolase activity. Importantly, the catalytic domain of PARP15 has been validated as a suitable substrate for multiple viral macrodomains16, making this assay readily adaptable to study diverse macrodomains and mutants thereof. However, the relatively low expression of PARP15.1 in cells has made the analysis somewhat challenging. Therefore, to further improve our assay, we developed the PARP15-K128R/K135R mutant, which is more stable than wild-type PARP15 due to the substitution of the two ubiquitin acceptor lysines with arginine. This results in a more robust and reliable readout of hydrolase activity in cells.
Using this PARP15.1 in cellulo assay, we analyzed SARS-CoV-2 macrodomain mutants to investigate their hydrolase activity. Based on in silico analyses of the macrodomain structure in complex with ADP-ribose, potentially critical amino acids in the ADPr binding pocket were substituted (N40R, G130D, and F132V) (Figs. 4 and 5). In addition, we included previously identified catalytically inactive mutant variants (D22V, G48V, and G130V)10, which we analyzed in our advanced assay system based on PARP15-KR (Fig. 8c, d). In agreement with previous in vitro findings10, mutations at positions N40 or F132 of the SARS-CoV2 MD significantly diminished hydrolase activity (Fig. 6). The G130V mutant completely abolished catalytic activity in vitro10, and was inactive in cells (Fig. 8c, d). In contrast, the G130D substitution retained hydrolase activity in cells (Fig. 6b, c), which is likely attributed to the specific amino acid substitution. Furthermore, the D22V and G48V mutant variants revealed a loss of hydrolase activity (Fig. 8c, d), consistent with published in vitro data10.
In addition, we also tested recently identified compounds, including the remdesivir analog GS-44152443, MDOLL-022947 and a series of small molecules (Z853_0023, Z8539_0029, Z3122, LL1_0023, LRH-0003, Z8539) identified by Gahbauer et al.25. Despite their promising binding profiles and application at high concentrations, none of these compounds showed hydrolase inhibitory activity in cells (Fig. 7). This underscores the critical need to evaluate candidate inhibitors in cell-based systems that account for physiological factors. A plausible explanation for the lack of activity may be poor cell-permeability, as previously noted for compounds identified by Gahbauer and colleagues25, or poor target engagement in cells.
GS441524, a nucleoside analog of AMP, was initially shown to inhibit the SARS-CoV-2 RNA-dependent RNA polymerase, reducing viral RNA synthesis and thus replication51,52. GS441524 binds to and inhibits the SARS-CoV-2 macrodomain with a KD of roughly 10 µM in vitro, as demonstrated by crystallography and ITC43. Considering that the concentration of ADP-ribose and of MARylated proteins is low under unstressed conditions53 and that GS441524 is taken up by cells52,54, we expected some inhibition of catalytic activity. However, our results indicate that the SARS-CoV-2 MD is not targeted by GS441524 in a cellular context. Similarly, the MDOLL-0229, which was identified via a peptide displacement assay and subsequently validated in a SARS-CoV-2 replication assay, failed to inhibit hydrolase activity in our in cellulo system. In the original study, pretreatment of Calu-3 cells with MDOLL-0229 and IFNγ prior to infection with SARS-CoV-2 reduced plaque-forming units, suggesting antiviral activity and cell permeability47. However, target engagement in cells was not addressed, and our results raise the possibility that the observed reduction in viral replication may be due to indirect effects rather than specific inhibition of macrodomain activity.
As none of the previously tested inhibitors exhibited functionality in our system, we expanded the inhibitor analysis by a set of pyrollo-pyrimidine-based compounds recently identified as SARS-CoV2 MD inhibitors (Fig. 8d, e)30. In contrast to the earlier compounds, but consistent with the published data, compounds 6d and 6e, previously shown to inhibit the MD and consequently the replication of murine hepatitis virus (MHV)30, also displayed inhibitory effects in our system.
In summary, our assay provides a valuable tool to study the hydrolytic activity of various macrodomains under physiologically relevant conditions. In addition, it allows to evaluate the catalytic activity of PARP15 in cells. It enables the evaluation of the effects of candidate inhibitory compounds of the catalytic activities of both macrodomains and PARP15, while simultaneously assessing cell toxicity and cellular availability of such candidate compounds. However, the detection of effects on PARP15 MARylation alone might not fully reflect inhibition of macrodomain-dependent signaling or viral replication. Complementary analysis, such as assessing viral propagation, will be necessary to fully validate the functional impact of candidate compounds. Nonetheless, this live-cell imaging approach represents a significant advancement to directly evaluate macrodomain inhibitors, bridging the gap between in vitro biochemical assays and more complex physiological or viral models.
Material and methods
Cell lines and cell cultures
U2OS-tdTomato-G3BP1 (provided by Paul Taylor, St. Jude Children’s Hospital, Memphis, USA)36, U2OS-tdTomato-G3BP1/EGFPm-PARP15 (this study), U2OS (originally obtained from DSMZ, ACC 785) and stable U2OS Flp-In T-Rex (provided by Dorothee Dormann, Johannes Gutenberg University, Mainz, Germany), stable HeLa Flp-In T-REx cells (originally provided by Steven Taylor, the University of Manchester, Manchester, UK), as well as HEK293 cells (originally obtained from DSMZ, ACC 305) were cultivated in DMEM supplemented with 10% heat-inactivated fetal calf serum (FCS) at 37 °C in 5% CO2. Stable HeLa Flp-In T-REx cell lines were additionally supplemented with 15 µg/mL Blasticidin S (Invivogen, ant-bl-5b) and 200 µg/mL Hygromycin B (Invivogen, ant-hg-1) for selection during every second passage. Thawed cells were regularly tested for mycoplasma. Therefore, genomic DNA was isolated using the peqGOLD tissue DNA Mini Kit (Peglab, 123396) and subsequently a PCR was performed to detect potential mycoplasma DNA using specific primers (GPO-1: 5′-ACTCCTACGGGAGGCAGCAGTA-3′, MGSO: 5′-TGCACCATCTGTCACTCTGTTAACCTC-3′)19.
U2OS-tdTomato-G3BP1-EGFPm-PARP15.1 and -PARP15.1-H559Y were constructed via lentiviral transduction. To generate lentiviral particles, HEK293T cells were transiently transfected with pLIX403-EGFPm-PARP15.1 or pLIX403-EGFPm-PARP15.1-H559Y, psPAX2, and pCMV-VSV-G using the calcium phosphate precipitation method. Two days after transfection, the supernatant containing mature lentiviral particles was collected and filtered using a 0.45 µm sterile filter. For lentiviral transduction, U2OS-tdTomato-G3BP1 cells were grown to 40% confluence, and normal DMEM replaced by 1 mL 10x Polybrene-DMEM, before adding 4.5 mL of the virus-containing supernatant and 4.5 mL DMEM. Six hours after transduction, the polybrene-containing medium was replaced by normal DMEM. This procedure was repeated the next day. Cells were selected using 1 µg/mL Puromycin.
U2OS Flp-In TREx cells were transfected with pcDNA5/FRT/TO-EGFPm-PARP15.1, -PARP15.1-HY, or PARP15.1-KR whereas HeLa Flp-In T-REx cells were transfected with pcDNA5/FRT/TO-EGFPm-PARP15.1-T2A-3xFLAG-NLS-NBGFP, -SARS-CoV-2 MD, -SARS-CoV-2 MD N40R, SARS-CoV-2 MD G130D, SARS-CoV-2 MD F132V, -CHIKV MD, or CHIKV MD V33E, or pcDNA5/FRT/TO-EGFPm-PARP15.1KR-T2A-3xFLAG-NLS-NBGFP-SARS-CoV-2 MD, -SARS-CoV2 MD D22V, -SARS-CoV2 MD G48V, or -SARS-CoV2 MD G130V and pOG44 (Invitrogen) using Lipofectamine 3000 according to the manufacturer’s protocol. Cells were selected for efficient integration using 15 µg/mL blasticidin S (Invivogen, ant-bl-5b) and 200 µg/mL hygromycin B (Invivogen, ant-hg-1).
Reagents and antibodies
The following reagents were used in this study: ß-NAD+ (Sigma-Aldrich, N6522), 32P-NAD+ (Hartmann, FP-321), Protease inhibitor cocktail (Sigma-Aldrich, P8340; MedChem Express, HY-K0011), polybrene (Hexadimethrinbromid, Sigma Aldrich, 107689), doxycycline (Sigma-Aldrich, D9891), Z8539_0023 (Enamine-ID: Z5010894404; 4-hydroxy-1-{4-[(phenylcarbamoyl)amino]benzamido}-2.3-dihydro-1H-indene-2-carboxylic acid), Z8539_0029 (Enamine-ID: Z5010894407; 1-{4-[(cyclopropylcarbamoyl)amino]benzamido}-2,3-dihydro-1H-indene-2-carboxylic acid), Z3122 (Enamine-ID: Z5389393122; 3-(4-bromophenyl)-3-({7H-pyrrolo[2,3-d]pyrimidin-4-yl}formamido)propanoic acid), LL1_0023 (Enamine-ID: Z5030903496, 8-fluoro-6-{7-fluoro-9H-pyrimido[4,5-b]indol-4-yl}-6-azaspirol[3.4]octane-8-carboxylic acid), LRH-0003 (Enamine-ID: Z5265428218, 1-({7-fluoro-9H-pyrimido[4,5-b]indol-4-yl}amino)pyrrolodin-2-one), Z8539 (Enamine-ID: Z4718398539, 1-{4-[(cyclopropylcarbamyol)amino]benzamido}-4-hydroxy-2,3-dihydro-1H-indene-2-carboxylic acid)25; OUL232 (MedChem express HY-148566), OUL 24340; GS441524 (Tocris, #7227, (2R,3R,4S,5R)-2-(4-Aminopyrrolo[2,1-f][1,2,4]triazin-7-yl)-3,4-dihydroxy-5-(hydroxymethyl)tetrahydrofuran-2-carbonitrile); MDOLL-022947; the pyrollo-pyrimidine-based compounds (7H-pyrrolo[2,3-d]pyrimidin-4-yl)-L-tryptophanate (5a), (7H-pyrrolo[2,3-d]pyrimidin-4-yl)-L-tryptophanate (5c), (S)-2-((7H-pyrrolo[2,3-d]pyrimidin-4-yl)amino)-N-((2-chloropyridin-4-yl)methyl)-3-(1H-indol-3-yl)propenamide (6b), (S)-2-((7H-pyrrolo[2,3-d]pyrimidin-4-yl)amino)-3-(1H-indol-3-yl)-N-(pyridin-2-ylmethyl)propenamide (6d), (R)-2-((7H-pyrrolo[2,3-d]pyrimidin-4-yl)amino)-3-(1H-indol-3-yl)-N-(pyridin-2-ylmethyl)propenamide (6e)30. TALON metal affinity resin (Takara, 635501), anti-GFP (Rockland, 600-101-2015, WB: 1:2000), anti-gamma-Tubulin (Sigma-Aldrich, GTU88, WB: 1:2000), anti-GAPDH (Santa Cruz, sc-32233, WB 1:2000), anti-PARP15 (Proteintech, 18126-1-AP, WB: 1:2000), anti-FLAG (Sigma Aldrich, F3165, clone M2, WB: 1:5000, IF: 1:500), goat-anti-mouse-HRP (Jackson Immunoresearch, 115-036-068, WB: 1:10000), rabbit-anti-goat-HRP (Santa Cruz, sc-2768, WB: 1:5000), goat-anti-rabbit IgG (Cell Sinalling, 7074P2, WB: 1:2000), goat-anti-mouse-Alexa-Flour-633 (Invitrogen, A21052, IF: 1:500), Spy650-DNA (tebu-bio, sc501, 1:1000), Immu-Mount (Epredia, 9990402), Hoechst33258 (Sigma-Aldrich, 94403), Lipofectamine 3000 (Thermo Scientific, L3000001), NEBuilder HiFi°DNA°Assembly°Master Mix (NEB, E2621L), CloneAampTM HiFi PCR Premix (Takara Bio, 639298), Gateway™ BP Clonase™ II Enzym-Mix (Thermo Scientific, 11789020), Gateway™ LR Clonase™ II Enzym-Mix (Thermo Scientific, 11791020), AflII (New England Biolabs, R0520S), NheI-HF (New England Biolabs, R3131), AgeI-HF (New England Biolabs, R3552).
Cloning and mutagenesis
Cloning of GW-pcDNA5/FRT/TO-EGFPm-PARP15.1-T2A-3xFLAG-NLS-NBGFP was achieved via Gibson assembly as follows: first, we generated a plasmid encoding the 3xFLAG-NLS-NBGFP. A nanobodyGFP construct19 was opened using NheI and AgeI restriction enzymes for the insertion of 3xFLAG-NLS, which was amplified in two steps via nested PCR. The 3xFLAG sequence was amplified using primer 1_for and primer 1_rev with GW-pCMV-3xFLAG as template (Table 1). The product from this reaction was used as template for the next amplification with primers primer 1_for and primer 1a_rev. Primers were designed in that way that they allowed for Gibson assembly of the final PCR product with the opened nanobodyGFP construct, resulting in a 3xFLAG-NLS-NBGFP construct.
Thereafter, three fragments were generated to be cloned via Gibson assembly into GW-pcDNA/FRT/TO to gain the final construct GW-pcDNA5/FRT/TO-EGFPm-PARP15.1-T2A-3xFLAG-NLS-NBGFP. First, EGFP was amplified using GWpEGFPm as a template and the following primers primer 2_for and primer 2_rev, resulting in a EGFPm-fragment with overhangs for assembly with the GWpcDNA5/FRT/TO backbone at the 5’ end and PARP15.1 at the 3’ end.
Second, the PARP15.1 fragment was generated via nested PCR. In a first step, PARP15.1 was amplified using GWpEGFPm-PARP15.1 as template and primer 3_for and primer 3_rev as oligos. The product resulting from this PCR was amplified using primer 3_for and primer 3a_rev. This resulted in a PARP15.1 fragment with overhangs to GFP at the 5′ end and containing part of the T2A site at the 3′ end for Gibson assembly. Third, the part harboring the 3xFLAG-NLS-NBGFP fragment was generated. Therefore, the previously generated 3xFLAG-NLS-NBGFP construct was used as a template and amplified via nested PCR. For the first amplification primer, 4_for and primer 4_rev were used. The resulting product was subsequently amplified using primer 4a_for and primer 4_rev. This resulted in a 3xFLAG-NLS-NBGFP fragment containing part of the T2A site at the 5′ end and overhangs for assembly with the GW-pcDNA/FRT/TO backbone. Finally, the GW-pcDNA5/FRT/TO was opened using the AFlII restriction enzyme, and the three fragments were inserted via Gibson assembly, resulting in the final GW-pcDNA5/FRT/TO-mEGFP-PARP15.1-T2A-3xFLAG-NLS-NBGFP construct.
Viral macrodomains were ordered as gBlocks from IDT and cloned by BP-reaction using the Gateway™ BP Clonase™ II Enzyme-Mix into GW-pDONRZeo. In a second step, the macrodomains were recombined into the GW-pcDNA5/FRT/TO-mEGFP-PARP15-T2A-3xFLAG-NLS-NBGFP construct by LR-reaction using the Gateway™ LR Clonase™ II Enzyme-Mix. All constructs were validated by sequencing.
To generate the PARP15.1-K128/135R mutant, a gBlock (IDT) containing StuI and BstBI restriction sites was synthesized, enabling replacement of the original sequence with the mutant insert upon restriction digestion and ligation.
Expression test of GFP-PARP15 and 3xFlag-NBGFP
HEK293 cells were transiently transfected with GW-pcDNA5/FRT/TO-EGFPm-PARP15.1-T2A-3xFLAG-NLS-NBGFP or GW-pcDNA5/FRT/TO-EGFPm-PARP15.1-T2A-3xFLAG-NLS-NBGFP-viral macrodomain using the calcium phosphate precipitation method. Two days after transfection, cells were lysed in RIPA buffer (10 mM Tris, pH 7.4; 150 mM CaCl; 1% NP-40; 1% DOC; 0.1% SDS; PIC), sonicated, and lysates centrifuged. Samples were separated by SDS-PAGE and analyzed via immunoblotting.
Evaluation of PARP15.1 protein expression and stability
Stable U2OS Flp-In TREx- pcDNA5/FRT/TO-EGFPm-PARP15.1, -HY, or -KR were seeded in 6-well plates. Protein expression was induced with 0.5 µg/ml Doxycycline for 16 h. Cells were either left untreated or were treated with DMSO (vehicle control) or OUL232 (10 µM) overnight. Following treatment, cells were harvested and lysed using RIPA lysis buffer (10 mM Tris, pH 7.4; 150 mM CaCl; 1% NP-40, 1% DOC; 0,1% SDS; PIC). Lysates were sonicated and subjected to SDS-PAGE followed by Western blotting to analyze protein expression. Similarly, stable HeLa Flp-In T-Rex- pcDNA5/FRT/TO-EGFPm-PARP15.1KR-T2A-3xFLAG-NLS-NBGFP-SARS-CoV-2 MD, -SARS-CoV2 MD D22V, -SARS-CoV2 MD G48V, or -SARS-CoV2 MD G130V cells were tested for protein expression.
Purification of His6-tagged fusion proteins
His6-tagged fusion proteins were expressed in E. coli Lemo21 (DE3). Fusions were enriched and purified via affinity chromatography using TALON metal affinity resin according to standard protocols.
In vitro ADP-ribosylation assay
The ADP-ribosylation reaction was carried out in ADP-ribosylation buffer (50 mM Tris, pH 8.0, 2 mM TCEP, 4 mM MgCl2, 0,1% NP-40) in the presence of 50 µM ß-NAD+ and 1 µCi γ-32P-NAD+ in a final volume of 30 µl. Reactions were incubated at 30 °C for 30 min and stopped by the addition of SDS sample buffer and subsequent heating at 95 °C for 5 min. Samples were separated by SDS-PAGE, and protein visualized by Coomassie brilliant blue staining of the gels. Thereafter, the gels were dried and exposed to X-ray films for the detection of the incorporated label.
Compound screening in cells
For inhibitor screening, 40,000 cells/well were seeded into a µ-Slide 8-well ibiTreat 1.55 mm Coverslip (IBIDI, 80826-90) in 250 µl DMEM + 10% FCS. After 6 h, cells were induced with Doxycyclin (1 µg/ml) and treated with compounds for 16 h. One hour prior to analysis by confocal microscopy, Spy650-DNA (tebu-bio, sc501) (1:1000) was added to the cells to visualize the nuclei.
Colocalization of 3xFlag-NLS-NB with GFP-PARP15 foci
U2OS cells were transiently transfected with pcDNA5/FRT/TO-mEGFP-PARP15.1_T2A_3xFLAG-NLS-NBGFP or pcDNA5/FRT/TO-mEGFP-PARP15_T2A_3xFLAG-NLS-NBGFP-macrodomain (SARS-CoV-2 MD1, CHIKV MD or CHIKV MD V33E) using Lipofectamine3000 according to the manual. Cells were fixed with 3.7% paraformaldehyde for 20 min at room temperature, quenched with 50 mM NH4Cl in PBS + 0.1% Triton-X100 for 5 min in the dark at RT, then permeabilized and blocked in PBS + 0.1% Triton-X100 + 1% BSA at room temperature for 30 min. Fixed cells were stained with α-Flag (Sigma Aldrich, F3165, Clone M2) 1:500 in PBS + 0.2% BSA for 45 min at 37 °C in a wet chamber, washed and incubated with goat-anti-mouse-Alexa-Flour-633 (Invitrogen, A21052) 1:500 as described before. Nuclei were stained with Hoechst33258 (SigmaAldrich) diluted to 1 µg/ml in ddH2O. Samples were mounted with Immu-Mount (Epredia, 9990402).
For imaging of fixed cells (initial PARP15.1. localization or FLAG co-staining): Images were captured using a Zeiss LSM710 Confocal Laser Scanning Microscope equipped with an AxioCam (Zeiss) and C-Apochromat 40× water immersion objective. For all pictures, the pinhole was set 1 airy unit for the longest wavelength and dimensions of 1024 × 1024 pixels (initial PARP15.1 localization studies) or 1180 × 1180 pixels (FLAG co-staining) were chosen. Pictures were taken in plane mode with beam splitters MBS 488/561/633 and MBS 405.
For initial PARP15.1 localization studies, the EGFP fluorochrome (emission wavelength of 528 nm) was excited at a wavelength of 488 nm by an argon laser and detected at 494–562 nM bandpass filter. The Hoechst33258 staining (emission wavelength of 453 nm) was excited at 352 nm and detected using a 410–496 nm band pass filter.
For FLAG co-staining, the EGFP fluorochrome (emission maximum: 509 nm) was excited at a wavelength of 488 nm by an argon laser and detected using a 488 nm single-channel photomultiplier (PMT) and a 494–582 nm bandpass filter. The Hoechst33258 staining was excited at a wavelength of 405 nm by a Diode 405-30 laser and detected using a 410–484 nm bandpass filter. The AF633 fluorochrome was excited at a wavelength of 633 nm by helium-neon laser and detected using a 638–747 nm bandpass filter.
For live-cell imaging: images were captured using a Zeiss LSM710 confocal laser scanning microscope equipped with an AxioCam (Zeiss) and C-Apochromat 40× water immersion objective. For all pictures, the pinhole was set 1 airy unit for the longest wavelength and dimensions of 980 × 980 pixels was chosen. The EGFP fluorochrome (emission maximum: 509 nm) was excited at a wavelength of 488 nm by an argon laser and detected using a 488 nm single-channel photomultiplier (PMT) and a 493–557 nm bandpass filter. The Spy650DNA was excited at a wavelength of 633 nm by helium-neon laser and detected using a 644–735 nm bandpass filter.
Images were analyzed for nuclear foci or spread GFP signal using a CellProfiler pipeline adapted from https://github.com/CellProfiler/examples/tree/master/ExampleSpeckles (Supporting Table 1).
Statistics and reproducibility
Postprocessing of the Nuclei-file resulting from the CellProfiler analysis and visualization of results was done using R55 in RStudio with the following packages: tidyverse, ggplot2, ggpubr, ggthemes, scales and rstatix. For the jitter plot, p-values were calculated with pairwise Wilcoxon test, adjusted by false discovery rate, and outliers were removed using the IQR method. To ensure reproducibility, experiments in which cultured cells were used for microscopy were performed at least three times, documenting at least six different frames with cells. In vitro experiments using recombinant purified proteins were performed twice. Expression profiles of proteins induced by doxycycline in the stable cells were performed once, as this was only for testing and confirming stable expression of the protein of interest.
Computational analyses
The crystal structure of the SARS-CoV-2 macro domain Mac1 complexed with its indigenous ligand ADP-ribose (PBD ID: 7KQP) is reported in the Protein Data Bank with a resolution of 0.88 Å. After the removal of all water molecules, one monomer of the dimeric crystal structure was used for all the following steps.
An in silico alanine scan was performed with the ABS-scan webserver42. Using this application, the in silico mutagenesis was carried out with the Modeler library on all residues within 4.5 Å of the complexed ligand and was followed by energy minimization of the complex using conjugate gradient and steepest descent algorithms, successively. The binding poses of the single-point mutated complexes were rescored employing the AutoDock 4.1 force field. Utilizing the Schrodinger Workspace application, 21 single point mutants were generated for seven different positions (ASP 22, ASN 40, GLY 48, VAL 49, GLY 130, PHE 132, and PHE 156), respectively. The binding pose of ADP-ribose in each of the mutated structures was refined using Schrodinger’s MM-GBSA protocol. Applying the Schrodinger Glide standard protocol56, ADP-ribose was redocked into all in silico mutated protein structures. The centers of the Glide grids were specified by the crystal structure complex. Default settings were used for the grid generation as well as for the molecular docking procedure. Following the docking, the binding poses were subjected to post-docking minimization to optimize the overall geometry of ADP-ribose.
Subsequently, changes in stability and flexibility of the point mutants N40R, G130D and F132V were evaluated by unbiased molecular dynamics simulations in GROMACS 2021.757. All simulations were performed using the AMBER ff99SB-ILDN force field for protein and ions, while the TIP3P model was used for explicit water molecules58. Hydrogen bonds were restrained by the LINCS algorithm59. Particle-mesh Ewald scheme with a grid spacing of 1.2 Å was applied for the calculation of long-range interactions, while the short-range electrostatic as well as van der Waals interactions were calculated with a distance cutoff of 12 Å. In all simulations, protein and solvent were coupled to separate heat baths. The velocity-rescale method60 was employed to control the temperature, while the Parrinello-Rahman method61 was used for pressure coupling. Prior to the molecular dynamics simulation, each system was minimized with the steepest descent algorithm. In a heating approach, each system was gradually heated to 310 K for 2 ns and subsequently equilibrated in a NVT ensemble for 10 ns while applying position restraints on the protein. Further equilibration was achieved in an NPT ensemble for 10 ns without any position restraints, followed by a 500 ns production run.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Image data are available in the BioImage Archive (http://www.ebi.ac.uk/bioimage-archive) under the accession number S-BIAD1673. The uncropped gels and membranes are included in the Supplementary Information.
Code availability
The R-code used to visualize the data obtained from microscopy is available at https://doi.org/10.5281/zenodo.1845103462.
References
Luscher, B. et al. ADP-ribosylation, a multifaceted posttranslational modification involved in the control of cell physiology in health and disease. Chem. Rev. 118, 1092–1136 (2018).
Weixler, L. et al. ADP-ribosylation of RNA and DNA: from in vitro characterization to in vivo function. Nucleic Acids Res 49, 3634–3650 (2021).
Rack, J. G. M., Palazzo, L. & Ahel, I. (ADP-ribosyl)hydrolases: structure, function, and biology. Genes Dev. 34, 263–284 (2020).
Luscher, B. et al. ADP-ribosyltransferases, an update on function and nomenclature. Febs J. https://doi.org/10.1111/febs.16142 (2021).
Jankevicius, G. et al. A family of macrodomain proteins reverses cellular mono-ADP-ribosylation. Nat. Struct. Mol. Biol. 20, 508–514 (2013).
Sharifi, R. et al. Deficiency of terminal ADP-ribose protein glycohydrolase TARG1/C6orf130 in neurodegenerative disease. Embo J. 32, 1225–1237 (2013).
Rosenthal, F. et al. Macrodomain-containing proteins are new mono-ADP-ribosylhydrolases. Nat. Struct. Mol. Biol. 20, 502–507 (2013).
Feijs, K. L., Forst, A. H., Verheugd, P. & Luscher, B. Macrodomain-containing proteins: regulating new intracellular functions of mono(ADP-ribosyl)ation. Nat. Rev. Mol. Cell Biol. 14, 443–451 (2013).
Rack, J. G., Perina, D. & Ahel, I. Macrodomains: structure, function, evolution, and catalytic activities. Annu. Rev. Biochem. https://doi.org/10.1146/annurev-biochem-060815-014935 (2016).
Rack, J. G. M. et al. Viral macrodomains: a structural and evolutionary assessment of the pharmacological potential. Open Biol. 10, 200237 (2020).
Constant, L. E. C. et al. Overview on chikungunya virus infection: from epidemiology to state-of-the-art experimental models. Front. Microbiol. 12, 744164 (2021).
Silva, L. A. & Dermody, T. S. Chikungunya virus: epidemiology, replication, disease mechanisms, and prospective intervention strategies. J. Clin. Investig. 127, 737–749 (2017).
Lamers, M. M. & Haagmans, B. L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 20, 270–284 (2022).
V’Kovski, P., Kratzel, A., Steiner, S., Stalder, H. & Thiel, V. Coronavirus biology and replication: implications for SARS-CoV-2. Nat. Rev. Microbiol. 19, 155–170 (2021).
Li, C. et al. Viral macro domains reverse protein ADP-ribosylation. J. Virol. 90, 8478–8486 (2016).
Eckei, L. et al. The conserved macrodomains of the non-structural proteins of chikungunya virus and other pathogenic positive strand RNA viruses function as mono-ADP-ribosylhydrolases. Sci. Rep. 7, 41746 (2017).
McPherson, R. L. et al. ADP-ribosylhydrolase activity of chikungunya virus macrodomain is critical for virus replication and virulence. Proc. Natl. Acad. Sci. USA 114, 1666–1671 (2017).
Alhammad, Y. M. O. et al. The SARS-CoV-2 conserved macrodomain is a mono-ADP-ribosylhydrolase. J. Virol. 95, https://doi.org/10.1128/JVI.01969-20 (2021).
Krieg, S. et al. Mono-ADP-ribosylation by PARP10 inhibits chikungunya virus nsP2 proteolytic activity and viral replication. Cell Mol. Life Sci. 80, 72 (2023).
Alhammad, Y. M. et al. SARS-CoV-2 Mac1 is required for IFN antagonism and efficient virus replication in cell culture and in mice. Proc. Natl. Acad. Sci. USA 120, e2302083120 (2023).
Leung, A. K. L., Griffin, D. E., Bosch, J. & Fehr, A. R. The conserved macrodomain is a potential therapeutic target for coronaviruses and alphaviruses. Pathogens 11, https://doi.org/10.3390/pathogens11010094 (2022).
Fu, W. et al. The search for inhibitors of macrodomains for targeting the readers and erasers of mono-ADP-ribosylation. Drug Discov. Today 26, 2547–2558 (2021).
Luscher, B., Verheirstraeten, M., Krieg, S. & Korn, P. Intracellular mono-ADP-ribosyltransferases at the host-virus interphase. Cell Mol. Life Sci. 79, 288 (2022).
Schuller, M. et al. Fragment binding to the Nsp3 macrodomain of SARS-CoV-2 identified through crystallographic screening and computational docking. Sci. Adv. 7, https://doi.org/10.1126/sciadv.abf8711 (2021).
Gahbauer, S. et al. Iterative computational design and crystallographic screening identifies potent inhibitors targeting the Nsp3 macrodomain of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 120, e2212931120 (2023).
Roy, A. et al. Discovery of compounds that inhibit SARS-CoV-2 Mac1-ADP-ribose binding by high-throughput screening. Antivir. Res. 203, 105344 (2022).
Brosey, C. A. et al. Targeting SARS-CoV-2 Nsp3 macrodomain structure with insights from human poly(ADP-ribose) glycohydrolase (PARG) structures with inhibitors. Prog. Biophys. Mol. Biol. 163, 171–186 (2021).
Bajusz, D. et al. Exploring protein hotspots by optimized fragment pharmacophores. Nat. Commun. 12, 3201 (2021).
Sherrill, L. M. et al. Design, synthesis and evaluation of inhibitors of the SARS-CoV-2 nsp3 macrodomain. Bioorg. Med. Chem. 67, 116788 (2022).
Pfannenstiel, J. J. et al. Identification of a series of pyrrolo-pyrimidine-based SARS-CoV-2 Mac1 inhibitors that repress coronavirus replication. mBio 16, e0386524 (2025).
Wazir, S., Maksimainen, M. M., Alanen, H. I., Galera-Prat, A. & Lehtio, L. Activity-based screening assay for mono-ADP-ribosylhydrolases. SLAS Discov. 26, 67–76 (2021).
Dasovich, M. et al. High-throughput activity assay for screening inhibitors of the SARS-CoV-2 Mac1 macrodomain. ACS Chem. Biol. 17, 17–23 (2022).
Russo, L. C. et al. The SARS-CoV-2 Nsp3 macrodomain reverses PARP9/DTX3L-dependent ADP-ribosylation induced by interferon signaling. J. Biol. Chem. 297, 101041 (2021).
Ildefeld, N., Steinhilber, D., Proschak, E. & Heering, J. HTRF-based assay for detection of mono-ADP-ribosyl hydrolyzing macrodomains and inhibitor screening. iScience 27, 110333 (2024).
Leung, A. K. et al. Poly(ADP-ribose) regulates stress responses and microRNA activity in the cytoplasm. Mol. Cell 42, 489–499 (2011).
Yang, P. et al. G3BP1 is a tunable switch that triggers phase separation to assemble stress granules. Cell 181, 325–345 (2020).
Guillen-Boixet, J. et al. RNA-induced conformational switching and clustering of G3BP drive stress granule assembly by condensation. Cell 181, 346–361 (2020).
Rhine, K., Odeh, H. M., Shorter, J. & Myong, S. Regulation of biomolecular condensates by poly(ADP-ribose). Chem. Rev. 123, 9065–9093 (2023).
Herzog, N. et al. Caspase-dependent cleavage of the mono-ADP-ribosyltransferase ARTD10 interferes with its pro-apoptotic function. Febs J. 280, 1330–1343 (2013).
Nizi, M. G. et al. Potent 2,3-dihydrophthalazine-1,4-dione derivatives as dual inhibitors for mono-ADP-ribosyltransferases PARP10 and PARP15. Eur. J. Med. Chem. 237, 114362 (2022).
Stirling, D. R. et al. CellProfiler 4: improvements in speed, utility and usability. BMC Bioinform. 22, 433 (2021).
Anand, P., Nagarajan, D., Mukherjee, S. & Chandra, N. ABS-scan: in silico alanine scanning mutagenesis for binding site residues in protein-ligand complex. F1000Res 3, 214 (2014).
Ni, X. et al. Structural insights into plasticity and discovery of remdesivir metabolite GS-441524 binding in SARS-CoV-2 macrodomain. ACS Med. Chem. Lett. 12, 603–609 (2021).
Wazir, S. et al. Discovery of 2-Amide-3-methylester Thiophenes that Target SARS-CoV-2 Mac1 and Repress Coronavirus Replication, Validating Mac1 as an Antiviral Target. J. Med. Chem. 67, 6519–6536 (2024).
Anmangandla, A. et al. A fluorescence polarization assay for macrodomains facilitates the identification of potent inhibitors of the SARS-CoV-2 macrodomain. ACS Chem. Biol. 18, 1200–1207 (2023).
Schuller, M. et al. Discovery and development strategies for SARS-CoV-2 NSP3 macrodomain inhibitors. Pathogens 12, https://doi.org/10.3390/pathogens12020324 (2023).
Wazir, S. et al. Discovery of 2-amide-3-methylester thiophenes that target SARS-CoV-2 Mac1 and repress coronavirus replication, validating Mac1 as an antiviral target. J. Med. Chem. 67, 6519–6536 (2024).
Sowa, S. T. et al. Preparation of screening assays for ADP-ribosyl readers and erasers using the GAP-tag as a binding probe. STAR Protoc. 3, 101147 (2022).
Forst, A. H. et al. Recognition of mono-ADP-ribosylated ARTD10 substrates by ARTD8 macrodomains. Structure 21, 462–475 (2013).
Butepage, M. et al. Assessment of intracellular auto-modification levels of ARTD10 using mono-ADP-ribose-specific macrodomains 2 and 3 of murine Artd8. Methods Mol. Biol. 1813, 41–63 (2018).
Gordon, C. J., Tchesnokov, E. P., Feng, J. Y., Porter, D. P. & Gotte, M. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus. J. Biol. Chem. 295, 4773–4779 (2020).
Li, Y. et al. Remdesivir metabolite GS-441524 effectively inhibits SARS-CoV-2 infection in mouse models. J. Med. Chem. 65, 2785–2793 (2022).
Ribeiro, V. C., Russo, L. C. & Hoch, N. C. PARP14 is regulated by the PARP9/DTX3L complex and promotes interferon gamma-induced ADP-ribosylation. Embo J. 43, 2908–2928 (2024).
Yan, V. C. & Muller, F. L. Advantages of the parent nucleoside GS-441524 over remdesivir for Covid-19 treatment. ACS Med. Chem. Lett. 11, 1361–1366 (2020).
R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/ (2023).
Friesner, R. A. et al. Glide: a new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 47, 1739–1749 (2004).
Abraham, M. J. et al. GROMACS: high performance molecular simulations though multi-level parallelism from laptops to supercomputers. Softw. X 1-2, 19–25 (2015).
Lindorff-Larsen, K. et al. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 78, 1950–1958 (2010).
Hess, B. P-LINCS: a parallel linear constraint solver for molecular simulation. J. Chem. Theory Comput. 4, 116–122 (2008).
Bussi, G., Donadio, D. & Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 126, 014101 (2007).
Parrinello, M. & Rahman, A. Polymorphic transitions in single crystals: a new molecular dynamics method. J. Appl. Phys. 52, 7182–7190 (1981).
Knapp, S. knappsarah/Foci-Assay: Screening assay to monitor mono-ADP-ribosylhydrolase activity of viral macrodomains in cells [Data Set]. Zenodo. https://doi.org/10.5281/zenodo.18451034 (2026).
Acknowledgements
We thank Lari Lethiö for providing us with OUL243 and the MDOLL-0229 compounds, Fabian Schmidt and Roko Zaja for providing us with the cDNA encoding PARP15.1, Dorothee Dormann for providing us with U2OS-Flp-In Trex cells, Paul Taylor for providing us with the U2OS-tdTomato-G3BP1 cells, Ksenia Krooß (NFDI4BIOIMAGE, DFG project ID 501864659) for help with metadata annotation and upload to the BioImage Archive repository. This work was supported by the Confocal Microscopy Facility of the Interdisciplinary Center for Clinical Research (IZKF) within the Faculty of Medicine at RWTH Aachen University, by the START program of the Medical School of the RWTH Aachen University to Patricia Korn (123/21) and Sarah Knapp (119/23), by the Deutsche Forschungsgemeinschaft DFG (VE1093/1-1) to Patricia Korn, and by the Volkswagen Foundation (9A870) to Giulia Rossetti, Stefan Knapp, Andreas Ladurner, Bernhard Lüscher and Patricia Korn.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualization and supervision, P.K., B.L., and G.R.; formal analysis and investigation, Sa.Kn., V.W., M.V., A.S., T.W., Le.He, Lu.H., Le.Hi., A.G., Ba.L., Sa.Kr., P.K.; resources, D.F., St.K., A.L., G.R., B.L., and P.K.; writing-original draft, P.K.; writing-review and editing, all authors; visualization, Sa.Kn., V.W., G.R., B.L., and P.K.; funding acquisition, St.K., A.L., G.R., B.L., and P.K.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Rachy Abraham and the other anonymous reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Joanna Timmins and Laura Rodríguez Pérez. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Knapp, S., Weber, V., Verheirstraeten, M. et al. Screening assay to monitor mono-ADP-ribosylhydrolase activity of viral macrodomains in cells. Commun Biol 9, 386 (2026). https://doi.org/10.1038/s42003-026-09832-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-026-09832-3
