
Abstract
The Holocene Thermal Maximum has been considered an analog for near-future climate. Terrestrial records show that this period culminated in Southwest Greenland with the Greenland Ice Sheet retreating behind its present-day position. However, there is a paucity of Holocene coastal marine records proximal to the ice sheet from which to infer marine conditions. Here we present a multi-proxy record from Nuup Kangerlua covering the past ~10,500 years, supported by a one-year sediment trap time-series. We infer modern sea-surface conditions comparable to those following the fjord’s deglaciation from 10,000 to 8000 calibrated years before present. Warmer temperatures led to a period of pronounced meltwater discharge and peak marine productivity by 7500 calibrated years before present. We detect an exceptional oceanographic regime with no recent analog from ~7000 to 3000 calibrated years before present, when reduced ice-sheet extent was coeval with entrainment of subpolar mode water (of Atlantic origin) into the fjord.
Similar content being viewed by others
Introduction
Warming of the Arctic region is leading to mass loss of the Greenland Ice Sheet (GIS), enhancing freshwater discharge to the surrounding seas1. Coastal areas around Greenland receive ~1 Tt of meltwater every year2, and the freshwater runoff rate has increased by 21% during the period 2011–2020 CE compared to 1980–2010 CE3. According to recent estimates, the GIS will lose 3.3 ± 0.9% of its total volume within the twenty-first century, regardless of the emission pathway humanity takes4. These changes are expected to significantly impact fjord environments, including important marine ecosystems that support the local fisheries and subsistence livelihoods.
Global temperature reconstructions show elevated temperatures over Greenland between 10 and 5 cal ka (BP) (hereafter, ka BP)5,6 The Holocene Thermal Maximum (HTM) is defined by a positive temperature anomaly of 0.3–1.8 °C relative to the 19th century5. In South Greenland, marine records show a delayed HTM from ~6 to 3 ka BP7,8,9,10,11,12. During the HTM, the GIS retreated behind its present-day extent13,14, perennial sea-ice cover in the Arctic shrank15, and several of the present-day ice caps were reduced or disappeared altogether14,16. The HTM was primarily driven by astronomical forcing, whereas today’s warming is driven by anthropogenic greenhouse gas forcing17. Still, this period has been considered a potentially useful analog for near-future conditions6, with global surface temperatures between 2011–2020 already reaching ~1.1 °C relative to 1850–1900 CE18.
Marine climate variability in the Arctic is closely linked to sea-level pressure differences between the Arctic Ocean and the middle latitudes related to the Arctic Oscillation (AO)19,20,21. In a positive AO phase (AO+), a weakening of the Beaufort Gyre20 causes the strengthening of the East Greenland Current (EGC), expanding the Subpolar Gyre and leading to a decreased entrainment of Atlantic waters to the West Greenland Current (WGC)22,23,24. The counterpart is a negative AO (AO-) where a weakening of the EGC gives rise to a relatively contracted subpolar gyre facilitating entrainment of warm saline Atlantic waters to the WGC22,23. In the mid-Holocene, a long period with predominantly AO-phase15,25,26 has been identified in several coastal sediment records from South Greenland during the HTM (from ~6 to 3 ka BP)7,8,9,10,11.
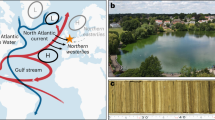
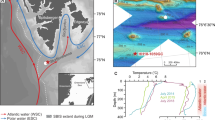
Nuup Kangerlua (Godthåbsfjord) is the largest fjord system in Southwest Greenland (Fig. 1), covering an area of ~2013 km2. Geological records indicate that, during the Last Glacial Maximum, the ice margin was located 20–40 km offshore of the present-day coastline15. The ice margin retreated from the outer coast to the present extent between ~11.4–10.4 ka BP27,28. The GIS margin in Nuup Kangerlua subsequently retreated further inland, reaching a minimum extent at ~5 ka BP28,29. The fjord has been a key area for the Greenland Ecosystem Monitoring program, where oceanographic observations have been conducted over the last 15 years30,31,32. The fjord hydrography is controlled by seasonal variation in circulation modes related to dense coastal inflows from the outer shelf, tidal mixing, glacial freshwater runoff, and sub-glacial circulation33. The system receives meltwater from the GIS through three marine-terminating glaciers (Narsap Sermia, Akullersuup Sermia, and Kangiata Nunaata Sermia) and three land-terminating glaciers (Saqqap Sermia, Qamanaarsuup Sermia, and Kangilinnguata Sermia) (Fig. 1).

a Greenland and the dominant ocean circulation pattern. The blue lines indicate the fresh and cold East Greenland Current (EGC), Baffin Island Current (BIC), and Labrador Current (LC). The red lines indicate the warm and saline North Atlantic Current (NAC), Irminger Current (IC), and the West Greenland Current (WGC) (which results from IC and EGC mixing). The dark gray area outlines a contracted Subpolar Gyre scenario, and the light gray area outlines an expanded Subpolar Gyre. The black box delineates the extent of figure b. b The red line shows the upper Subpolar Mode Water (uSPMW) and deep Subpolar Mode Water (dSMW), the blue line shows the Baffin Bay Polar Water (BBPW), and the purple line shows the Southwest Greenland Coastal Water (CW)30. The location of the sediment cores and CTD (SA13-ST6-36R, SA13-ST6-40G, and SA13-ST6-41CTD—all named SA13-ST6) and the sediment trap (PPS 4/3 from 2017-18) analyzed in this study are marked by yellow stars, situated within Kapisillit Kangerluat (KK). The main glaciers are indicated as: KS (Kangaasarsuup Sermia), NS (Narsap Sermia), QS (Qamanaarsuup Sermia), AS (Akullersuup Sermia), and KNS (Kangiata Nunaata Sermia). The black dashed line follows the Last Glacial Maximum (LGM) end moraine, and the white dashed line follows the Kapisigdlit stade moraine deposited between ~10.4 and 10 ka BP27,28,29. c CTD (conductivity/salinity, depth, temperature) and fluorescence profile at the core site (SA13-ST6-41CTD) obtained at the time of core collection (14-08-2013)49.
Today, the coastal inflow to the fjord results from a mixture of three water masses; the Subpolar Mode Water, Southwest Greenland Coastal Water, and Baffin Bay Polar Water30,33,34,35 (Fig. 1). The Subpolar Mode Water of Atlantic origin is transported to the region by the WGC and it is an important heat source for marine-terminating glaciers30,36. The cold and fresh Southwest Greenland Coastal Water originates from the sea-ice-loaded Polar Waters of the upper EGC and freshwater runoff from land30,37. The Baffin Bay Polar Water is carried by the Baffin current and blocks the northward transport of the Southwest Greenland Coastal Water30. Both the warm and saline Subpolar Mode Water (T ≈ 5 °C and S ≈ 35) and the colder and fresher Southwest Greenland Coastal Water (T ≈ 0 °C and S ≈ 33) can be observed outside the fjord34. Southwest Greenland Coastal Water can restrict the Subpolar Mode Water from entering the fjord system35. In addition, the fjord is constrained by several sills, including a main sill (170 m water depth) at the mouth. Inside the fjord, the summer surface layer is formed from freshwater runoff, and the properties of the layer vary with the season and distance to the source, with surface salinity varying from ~5 to 28 and reaching a minimum in mid-August, and sea-surface temperatures ranging ~0–10 °C34.
Sub-glacial freshwater discharge from the marine-terminating glaciers in Nuup Kangerlua brings up nutrient-rich deep water by upwelling32,38, including nitrate, the main growth-limiting resource in the area31,39. This is an important mechanism for stimulating primary production32,39,40. Today, this is a relatively productive fjord system, with a total annual primary production of ~120 g C m-2 yr-1 39,41. Sediment core records covering the last ~3300 years have (i) shown that long-term changes in primary productivity are mainly controlled by climate variability and associated enhanced meltwater discharge, and (ii) linked increased primary production to enhanced meltwater discharge into the fjord42,43.
To obtain new insights into the Holocene environmental history of Nuup Kangerlua, we generated a multi-proxy record covering the last ~10.5 ± 0.3 cal ka BP. Our record includes grain-size distribution data (including ice-rafted debris (IRD)), biogeochemical data (total organic carbon (TOC), total nitrogen (TN), calcium carbonate (CaCO3), the stable isotopic composition of bulk organic matter (δ13C and δ15N), and biogenic silica (BSi) as well as dinoflagellate cyst (dinocyst) assemblages.
Dinoflagellates are single-celled protists, and around 15–20% of the species are known to produce resting cysts as part of their life cycle44. The cysts are composed of highly resistant organic material and are thus generally well-preserved in sediments. Dinocysts are particularly useful proxies at high latitudes, where the preservation of siliceous (e.g., diatoms) and calcareous (e.g., foraminifera) microfossils can be problematic. Dinocysts are useful indicators of sea-surface conditions45,46, and the group includes both primary producers and heterotrophic species acting as micrograzers47. To further validate interpretations based on dinocyst assemblages in our sediment record, we analyzed seasonal changes in vertical particulate matter and dinocyst species fluxes from material collected by a sediment trap deployed over one year in the fjord (May 2017–April 2018). Sediment trap studies from high latitudes are relatively rare48, but important for determining local species responses and the seasonal nature of biological proxy data.
Our multi-proxy record from Nuup Kangerlua revealed that, in the context of the Holocene, sea-surface conditions at present are similar to those following the rapid deglaciation of the fjord. In contrast, inferred conditions during peak ice-sheet retreat in the mid-to-late Holocene indicate an exceptional oceanographic regime, with entrainment of Atlantic-sourced water. While the reconstructed Holocene Thermal Maximum conditions have no modern analog, they indicate a plausible future scenario in which sustained warming would enhance the inflow of relatively warm and saline water masses into the fjord.
Results
Core retrieval, geochronology, and grain-size distribution
The fjord sediment archives analyzed in this study consist of a gravity core (SA13-ST6-40G) and Rumohr-lot core (SA13-ST6-36R) (Table 1). The cores and CTD profile (Table 1) were retrieved from the intra-fjord channel Kapisillit Kangerluat onboard the Greenlandic research vessel Sanna in 2013 (Fig. 1b)49. The automated Technicap PPS 4/3 sediment trap GF10 was deployed in the main fjord branch at a depth of 300 m, ~290 m above the seafloor, i.e., at a site with 590-m water depth (Table 1) (Fig. 1b). The sediment trap samples were collected over a full year at a 20- to 45-day interval.
An age-depth model was constructed using 14 radiocarbon ages (14C) on organic material (polychaete tubes) and benthic foraminifera in OxCal 4.4 (see “Methods”). A minor reversal in the radiocarbon dates was observed at 377.5 cm, based on the dating of benthic foraminifera. The very small sample size of this date, however, was associated with a large error of ~330 years, making it compatible with surrounding dates, and it was therefore not treated as an outliner. There was also strong agreement between the two dates conducted on different materials at 400.5 cm and 403.5 cm. The two cores show marine sedimentation spanning the past ~10.5 ka BP with sedimentation rates ranging from 0.04 to 0.08 cm yr-1 in the gravity core, giving an average sample resolution of 18 years per centimeter (Table 2 and Fig. 2).

High-resolution optical image, sedimentation rate (cm yr-1) in black, magnetic susceptibility (MagSus) in red, and grain-size distribution (fine sand fraction in orange and coarse sand fraction in brown) for gravity core SA13-ST6-40G. The grain-size fraction >63 μm is interpreted as IRD. The relationship between depth and age is shown for both cores, SA13-ST6-40G and SA13-ST6-36R (inserted). Green dots are 210Pb-based dates (Supplementary Table 1), blue dots denote 14C dates of polychaete tubes, and orange dots denote 14C dates from mixed benthic foraminifera. The uncertainties associated with each individual radiocarbon date are shown in Supplementary Fig. 1. The gray area displays the 2σ uncertainty age range, and the dark gray box indicates when the fjord system was deglaciated based on terrestrial studies101.
The gravity core consists of 48–99% clay and silt (<63 μm), 1-14% fine sand (63–150 μm), and 0–46% coarse sand (>150 μm) (Fig. 2). Sand grains (>63 μm) are interpreted as IRD. Several peaks in magnetic susceptibility and IRD concentrations are present in the core data. The largest from ~10.5 to 9.5 ka BP (553–512 cm) contains 6.3-46.4% of grains >63 μm. Additional peaks with >6% of grains >63 μm are present from ~9.3 to 9.2 ka BP (498–496 cm), ~8.1 to 7.8 ka BP (417–400 cm), at ~6.7 ka BP (345–344 cm) and from ~150 to −31 yrs BP (8–0 cm) (Fig. 2).
Biogeochemical analysis used to infer productivity changes and organic matter sources
Biogenic silica (BSi) can be a useful proxy for siliceous primary production (see methods). The BSi concentration varies from 9.1 to 29.9 mg g-1 dry mass (mean 17.32 mg g-1 dry mass) throughout the record (Fig. 3). The BSi concentration increases from a minimum in the lowermost part of the core to a maximum at ~3.2 ka BP (208 cm), after which it sharply decreases and remains low to the top of the core. The highest BSi concentrations (>20 mg g-1 dry mass) were measured between ~5.6 and 3.2 ka BP (304-208 cm). The BSi flux was based on sediment accumulation rates and BSi concentrations. Throughout the record, the BSi flux ranges from 0.5 to 1.4 mg cm2 yr-1 (mean 0.931 cm2 yr-1) (Fig. 3). The BSi flux remains >0.6 mg cm2 yr-1 until ~4 ka BP with a small decrease between ~7 and 4 ka BP (Fig. 3). After 2 ka BP an increase is observed and the BSi flux remains >0.7 mg cm2 yr-1 for the most recent part of the core.

Sedimentation rate in black, the BSi content in dark green and the BSi flux in light green, the total organic carbon (TOC) in dark green and the TOC flux in light green, nitrogen isotopic composition (δ15N), total N (TN), organic carbon isotopic composition (δ13C) and the C/N ratio in dark green. The arrows indicate how the data are interpreted, and the dashed line separates the origin of organic matter (terrestrial versus marine).
Total organic carbon (TOC) content in sediments can be used as a proxy for primary production over time, although at coastal sites, organic carbon can also be of terrestrial origin and impacted by mineralization50. TOC content varies from 0.21 to 1.37% (mean 0.92%) throughout the record (Fig. 3). Minimum TOC values are observed in the bottom part of the record and increase upwards, reaching maximum values at ~2.9 ka BP (192 cm core depth). Thereafter, the TOC values decrease towards the top of the record. TOC flux was based on sediment accumulation rates. TOC flux varies from 1.9 to 8.9 g C m2 yr-1 (mean 4.772 g C m2 yr-1) throughout the record (Fig. 3). The TOC flux is lowest in the bottom part of the record until ~9 ka BP, where it increases, reaching maximum values between ~2 to 1.1 ka BP after which it declines towards the present.
Increasing δ15N values may indicate increasing productivity since phytoplankton preferentially consume the lighter nitrogen isotopes51. δ15N values gradually decline towards the top of the core and vary from 2.8 to 8.3‰ (mean 5.5‰), with the greatest variation occurring in the Early Holocene (Fig. 3). Total N varies from 0.02 to 0.19 wt% (mean 0.12 wt%). The C/N ratio and δ13C can be used to determine the origin of organic matter (terrestrial, marine, or sea-ice environments). The C/N ratio varies from 6.8 to 13.1 (mean 8), indicating a predominantly aquatic organic matter source52, and δ13C values vary from −25.7 to −19.8‰ (mean −22.5‰), which is closer to values typically for marine production53,54 (Fig. 3). The C/N ratio and δ13C display a change from a relatively high terrestrial organic component in the bottom part of the record towards increased marine organic influence in the upper part of the record.
Dinocyst assemblages
Fossil assemblages in sediment cores
A total of 17 dinocyst taxa were identified in the gravity core and the Rumohr-lot core, combined (Supplementary Figs. 2 and 3). In the gravity core, dinocyst concentrations are ca. 1800-95400 cysts g-1 (average 27241 cysts g-1) while fluxes span 197-5741 cysts cm-2 yr-1 (average 1420 cysts cm-2 yr-1) (Fig. 4). Dinocyst assemblages show compositional shifts throughout the record, which we summarized into six different zones via cluster analysis (Fig. 4). The bottom of the core is referred to as zone 1 (~10.5 to 9.2 ka BP) and is characterized by the lowest cyst concentrations and the highest proportion of heterotrophic species, dominated by Islandinium minutum subsp. minutum (I. minutum) and Brigantedinium spp. Zone 2 ( ~ 9.2 to 8.5 ka BP) is characterized by an increase in cyst concentration and a shift from a mainly heterotrophic assemblage to a diverse, primarily autotrophic assemblage, with Pentapharsodinium dalei (P. dalei) cysts making up >50% of the total assemblage. Zone 3 (~8.5 to 7 ka BP) is characterized by a further increase in autotrophic species, and a peak in dinocyst concentration mainly driven by P. dalei. In Zone 4 (~7 to 3.2 ka BP) the proportion of Nematosphaeropsis labyrinthus (N. labyrinthus) increases substantially (up to 22%) and its presence is nearly exclusive to this interval. In Zone 5 (~3.2 to 0.5 ka BP) N. labyrinthus was not observed, while the relative abundance of Spiniferites ramosus increased. In Zone 6 (~0.5 ka BP to present), the assemblage is still mainly composed of autotrophs but with a gradual increase in the proportion of heterotrophic taxa. The dinocyst cf. Biecheleria spp. and freshwater ciliate Halodinium spp. are also more abundant in this zone. The dinocyst assemblage in the Rumohr-lot core (Fig. 4) has a similar composition to zone 6 of the gravity core.

The autotrophic dinoflagellate species are shown in green, and the heterotrophic dinoflagellate species in brown. Cyst concentration, autotrophic/mixotrophic versus heterotrophic species, and cyst flux are plotted. Fluxes for pollen grains and spores, copepod eggs, foraminiferal linings, and Halodinium spp. are shown in gray. The six zones are indicated by horizontal lines and are based on cluster analysis.
Modern assemblages in the sediment trap
Fourteen morphotypes were identified in the sediment trap samples, all of which were also present in the sediment record. The most abundant species in the traps were I. minutum and P. dalei (Fig. 5), while Ataxiodinium choane, Bitectatodinium tepikiense and N. labyrinthus, which were abundant in parts of the sediment cores, were not detected in the trap samples. The dinocyst species in the traps exhibit clearly differing seasonal patterns. In spring from May to June 2017, simultaneously with drifting ice mélange and icebergs at the site, downward sediment flux consisted primarily of I. minutum subsp. minutum and Echinidinium karaense (E. karaense). For this same period, diatom assemblages analyzed previously consisted of cold-water species48. From July to August, fluxes of the autotrophic taxa P. dalei and Biecheleria spp., and the heterotroph Brigantedinium peaked. The dinocyst data are compared with previously published mass and TOC fluxes27. The mass flux was relatively steady throughout the year (ranging from 2 to 7 g m-2 d-1)48 while TOC flux varied from 40 to 140 mg C m-2 d-1. The highest TOC fluxes in summer (August-September 2017) co-occurred with the highest dinocyst diversity.

Plotted are dinocyst fluxes as recorded by a one-year automated sediment trap deployment. The green bars show autotrophic dinoflagellate species, and the brown bars show heterotrophic dinoflagellate species. The dark color represents cysts with cell content, and the light color represents empty cysts. The total mass flux and TOC flux48 are plotted with gray bars.
Discussion
Deglaciation followed by increased fjord productivity
Rising air temperatures55,56 and ocean forcing9 in the Early Holocene led to a retreat of the ice margin to the inner part of Nuup Kangerlua by ~11.4–10.4 ka BP27 (Fig. 6), where the Kapisigdlit stade moraine was deposited ~10.4–10 ka BP28 (Fig. 1). This is evidenced in our record by the high sedimentation rate and relatively large number of grains >63 μm, interpreted as IRD. Low dinocyst concentrations and fluxes, low BSi and TOC levels, and relatively high C/N ratios, all indicate low productivity in the fjord when terrestrial inputs remained high. The dinocyst assemblages at this time were dominated by heterotrophic taxa (I. minutum, Islandinium? cezare/P. tricingulatum, E. karaense, Brigantedinium spp., and Selenopemphix quanta). A comparison with data from the sediment trap shows that Early Holocene dinocyst assemblages most closely resemble those of modern cold surface-water conditions. The sediment trap data shows peak abundances of I. minutum and E. karaense in June during the spring bloom. The breakup of an ice mélange (mixture of sea ice and icebergs) located in the inner part of the fjord system feeds the site with icebergs from June to September, supporting the idea that I. minutum and E. karaense thrive in cold surface-water conditions influenced by icebergs and ice melt45,57,58,59,60 (Fig. 4). Combined, the sedimentology, dinocyst and biogeochemical records indicate low sea-surface temperatures and melting ice consistent with the Early Holocene deglaciation of the fjord and in line with other reconstructions from West Greenland61.

The block shaded in gray indicates a period of enhanced influence of Atlantic Water. The white boxes indicate glacier dynamics based on terrestrial evidence28,29. The dark gray boxes in the bottom panel show the timing of the increased influence of Atlantic Water inferred from other marine records from West Greenland, plotted from south to north7,8,9,10,11,12. a GAMs on the principal component analysis (PC) axis 1 and 2 scores (PCA on dinocyst species data). b Total dinocyst flux and concentration. c Organic matter δ13C values, where higher values indicate increased marine carbon. d Atlantic Water influence as indicated by the concentration of dinocyst N. labyrinthus. e Atlantic Water influence as indicated by foraminiferal assemblages from Disko Bay9,102. f Grains >63 μm interpreted as IRD (light blue) and sedimentation rate (dark blue). g Camp Century ice core δ18O-based56 air temperature reconstruction.
The generalized additive models (GAMs)62,63 of the first two principal components of the dinocyst assemblages indicate changing environmental conditions from deglaciation to ∼8 ka BP (Methods, Supplementary Figs. 5 and 6). The change described by the GAMs primarily reflects the shift from heterotrophic species dominance to autotrophic species dominance (Ataxiodinium choane, cyst of P. dalei, Operculodinium centrocarpum, Spiniferites ramosus and Spiniferites elongatus). Furthermore, the site became more productive during this period, as evidenced by an increase in BSi concentrations, TOC content, dinocyst concentration, and dinocyst flux. The observed decrease in IRD, suggests reduced calving/iceberg activity. At ~7.5 ka BP, the dinocyst concentration and dinocyst flux peaks, and a substantial increase in the δ13C values is observed. Higher δ13C values suggest eithera reduced influence of terrestrial carbon50 or an increase of organic marine carbon. δ13C values of −20 to −22‰ are typical for marine phytoplankton, while in high-latitude regions, the δ13C signal of marine organic carbon has been reported to vary between −16.7 and −30.4‰ (∼ −24‰) reflecting potential contributions from enriched sea-ice carbon (∼ −15 to −20‰) and depleted carbon (up to −30‰) produced in cold surface waters64,65. Decreased IRD suggests a change from an ice-sheet proximal setting to a more marine environment during the Early Holocene, consisting of reduced terrestrial carbon flux to the core site. The sediment trap (Fig. 5) supports the notion that most dinoflagellate primary producers bloom in the summer months39, when the highest dinocyst diversity and TOC are observed. Thus, the increased primary productivity, dinocyst diversity, and shift towards more phototrophic taxa all suggest overall higher sea-surface temperatures (Fig. 6h) and increased light availability. This is consistent with an increase in P. dalei cysts (>50% of the total assemblage), which can be associated with a stratified water-column during summer66.
Minimum ice-sheet extent and increased salinity
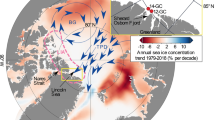
From ~7 to 3 ka BP, there is a compositional shift in the dinocyst assemblage, reflected by the cluster analysis (zone 4) (Fig. 4). This shift coincides with lower sedimentation rates and reduced IRD. While the sedimentation rate is lower, a larger proportion of the deposited material is BSi, supported by relatively high productivity based on the BSi and dinocyst concentrations. More striking is the relative abundance of N. labyrinthus, which is unaffected by sedimentation processes. At ~7 ka BP, the relative abundance of N. labyrinthus increases (Fig. 6), and at ~5 ka BP, its relative abundance represents >20% of the total assemblage. The modern-day distribution of N. labyrinthus in Northern Hemisphere surface sediments46 with respect to sea-surface temperature and salinity67,68,69 shows that the species is found in areas influenced by relatively warm and saline Atlantic Water (Fig. 7). To assess this further, the relative abundance of N. labyrinthus was plotted against sea-surface temperature and sea-surface salinity (Supplementary Fig. 6). The species was detected in the sediment at sites with summer sea-surface temperatures ranging from −2 to 30 °C and summer salinities ranging from 28 to 38. Some sites have likely been affected by transport, so these ranges likely do not accurately reflect the species optima but rather a broader range. However, they indicate that the species likely tolerates a larger temperature range compared to salinity (Supplementary Fig. 6).

a Distribution map for N. labyrinthus based on the Northern Hemisphere dataset of “modern” dinocyst dataset46. The points are samples, and the green color indicates the relative abundance of the species in the samples ranging from 0 to 80%, b Sea-surface salinity ranging from 15 to >35 from the World Ocean Atlas 201867,68, c Sea-surface temperature ranging from -5 to 20 degrees from the World Ocean Atlas 201867,69. All maps were created using Ocean Data View100.
The Northern Hemisphere dataset of “modern” dinocyst assemblage samples from 1968 sites across the Northern Hemisphere46 were plotted against summer sea-surface salinity and temperature (Fig. 8). In our record, N. labyrinthus reached a relative abundance of >20%. Today, summer sea-surface salinity ranges from ~5–28, and summer sea-surface temperature ranges from ~0–10 °C in Nuup Kangerlua34. None of the modern-day surface samples with an abundance of >10% and only one sample with >5% of N. labyrinthus is observed under the present-day sea-surface salinity and temperature conditions in the fjord. Thus, the period with high N. labyrinthus relative abundance is indicative of either much increased salinity in the fjord, or stronger advection of Atlantic-derived waters.

The relative abundance of dinocyst N. labyrinthus from the Northern Hemisphere dataset of “modern” dinocyst dataset46 plotted against sea-surface temperature and sea-surface salinity from the World Ocean Atlas 201867,68,69. The dashed box shows the range of sea-surface salinities and sea-surface temperatures measured in Nuup Kangerlua at present-day.
At ~5 ka BP, the ice sheet grounding line retreated inland from its present position, and meltwater runoff to the study site was reduced10. Ice sheet retreat would have enhanced freshwater runoff to the fjord, lowering its surface salinity, and resulting in unfavorable conditions for N. labyrinthus even though phytoplankton blooms occur at the sub-surface. A possible scenario that explains the high abundance of N. labyrinthus during minimum ice-sheet extent is that a stronger advection of Atlantic-derived waters led to N. labyrinthus being brought into the fjord.
The modern sediments from the fjord had low abundances of this cyst taxon (maximum 2% of the total assemblage). The sediment trap samples deployed in the main channel over a full year at a vertical depth position of 300 m contained no cysts of N. labyrinthus. Atlantic-derived water is often observed at depths between 200 and 800 m, so the absence of N. labyrinthus exclusively in the trap points towards the possibility of the species being advected into the fjord at present. It should, however, be considered that the trap data covers only one year (2017–2018). Thus, it is possible that conditions favorable for N. labyrinthus blooms can take place in other years.
At ~5 ka BP, enhanced inflow of Atlantic Water (Subpolar Mode Water) to the WGC has been recorded in several marine sediment records7,8,9,10,11,12,70,71,72,73, and attributed to a dominant AO-phase15,25,26 (Fig. 6). Here, the formation of multiyear sea ice reduced the export of freshwater and ice through Fram Strait. This led to a weakening of the EGC, giving rise to a relatively contracted subpolar gyre that facilitated entrainment of warm saline Atlantic Waters to the WGC22,23, leading to mild winters in West Greenland. Thus, minimum ice-sheet extent and limited Southwest Greenland Coastal Water in the fjord system would have made it possible for the relatively warm and saline Subpolar Mode Water to enter the fjord from ~7 to 3 ka BP. A study of reworked marine bivalve shells provides supporting evidence for optimum oceanographic conditions close to 5.0 ka BP in a period of maximum retreat of the ice sheet margin in West Greenland74.
On a much larger spatial scale, the period from 6.1 to 4.0 ka BP is notable for the anomalous warming of Atlantic surface-water masses south of Iceland75. Marked thermohaline changes also occurred farther north in the Arctic sector of the Atlantic. In West Spitsbergen fjords, an ice-rafting event and changes in hydrography and bottom water ventilation were found and dated between 4.4 and 3.8 ka BP76. As the Subpolar Mode Water component of the WGC is an important heat source for glacier mass balance30,36, the enhanced inflow of this water mass would likely lead to further melting of the marine-terminating glaciers in Nuup Kangerlua. Evidence for mid-Holocene tidewater glacier retreat is available from a site in Disko Bay, where an episode of major IRD deposition and a meltwater discharge event was found to have occurred close to 4.0 ka BP77. We suggest that the exceptionally high N. labyrinthus abundance during this period reflects anomalous atmospheric and sea-surface warming. Such warming is manifest in the NGRIP ice core as a 5-degree K positive temperature anomaly from ca. 4.4 to 3.9 ka BP. Furthermore, positive temperature spikes are observed in the Agassiz ice core (Ellesmere Island), Greenland Renland ice core, and Camp Century ice core between 4.29 and 3.79 ka BP56,78 (Fig. 5). Regional high-latitude climate warming during this period may be related to an atmospheric circulation regime forcing widespread climate anomaly79,80.
Cooling and ice sheet re-advance linked to freshening
The cluster analysis indicates a compositional shift in the dinocyst assemblage at ~3 ka BP, coinciding with an increased sedimentation rate. The general increasing trend in the percentage of heterotrophic dinocysts suggests colder surface-water conditions during the Neoglacial period29. The dinocyst N. labyrinthus was not detected in the assemblages from this period. In contrast, N. labyrinthus is present in a sediment record from outside the fjord system during this time interval73. This suggests that Subpolar Mode Water is present on the shelf, but either Southwest Greenland Coastal Water limited its inflow into the fjord system or a reduction in surface salinity precluded the species from thriving in the fjord during that period. Previous studies from Nuup Kangerlua suggest a mild climate during the early part of the Neoglacial, promoting glacial melt and leading to enhanced productivity until ~2.8 ka BP43. Our findings support decreased productivity from ~3 ka BP. Within the overall trend of Neoglacial cooling, the period from ~1.8–1.5 ka BP coincides with a temporary increase in productivity and Atlantic Water indicators (Fig. 6), and is consistent with relatively warmer surface-water conditions during the Roman Warm Period, as reported in other West Greenland studies7,8,9,10,11,12.
The last millennium
The GAMs indicate that from ~1 ka BP to the present, a transition to a new state is ongoing. The last millennium is marked by a relatively high sedimentation rate and increased IRD content. Furthermore, relatively high productivity levels are observed in our record from ~1950 CE to the present, based on dinocyst concentrations and a relatively high BSi. High productivity levels have been shown to be sustained by the upwelling of nutrient-rich waters driven by sub-glacial discharge40. Thus, this suggests enhanced meltwater discharge to Nuup Kangerlua, leading to increased productivity. This finding was also evident in studies of high-resolution sediment records from the fjord system covering the last 3300 years42,43. Here, Oksman and co-authors (2022 and 2024) found that primary productivity levels increased substantially in ~1900 CE and onwards which they associated with enhanced meltwater input42.
The most recent dinocyst assemblages show an increase in heterotrophic species, including cold-water and ice-melt indicators such as I. minutum45. This observation implies that the surface waters in Nuup Kangerlua are cooling relative to the period prior to ~1950 CE. Modern monitoring data covering the last 15 years, show that freshwater input to Nuup Kangerlua has varied substantially on both a seasonal and interannual time scale30,31,32,33,38. However, the inner part of the fjord is categorized by large runoff81. Our long-term record indicates that meltwater runoff has increased from ~1950 CE, as shown by the increased concentrations of the freshwater ciliate Halodinium and the first observations of the brackish-water indicator dinocyst Biecheleria spp., both found in areas with low surface salinity45. Our results support the idea that the cooling of the surface waters resulted from increased freshwater runoff, including meltwater and ice discharge into Nuup Kangerlua during the last decades.
Implications for a future warmer climate
Today, fjord conditions are akin to the period following deglaciation when a warmer climate facilitated ice-sheet retreat and enhanced freshwater discharge. Heterotrophic dinocyst species, which thrive during periods of ice melt, are dominant at present, and diatoms are the main primary producers in the fjord31. The period between ~7 and 3 ka BP with minimum ice-sheet extent has no modern analog. Today, the relatively fresh Southwest Greenland Coastal Water limits the inflow of warm Subpolar Mode Water into Nuup Kangerlua. Southwest Greenland Coastal Water originates from the EGC and runoff from the Southwest Greenland coast30,37. A warmer climate would facilitate positive AO-like conditions with enhanced export of freshwater and ice through Fram Strait. This could lead to a stronger component of the EGC to the WGC, increasing the amount of Coastal Water. Thereby, a warmer (near-future) climate will likely lead to enhanced Southwest Greenland Coastal Water restricting the Subpolar Mode Water from entering the fjord system. Enhanced Southwest Greenland Coastal Water would likely change the water-column properties, leading to more stratified waters in summer, higher turbidity rates impeding nutrient supply, and limiting light penetration, all affecting the coastal ecosystems38. Under continued warming and ice-sheet retreat, it is plausible that the hydrography of Nuup Kangerlua will change due to stronger advection of Atlantic-derived waters and/or reduced freshwater discharge to the area, facilitating an increase of Atlantic-derived inflows into the fjord, such as those for the mid-to-late Holocene described in this study.
There are, however, significant differences between the HTM and ongoing warming. While the HTM was driven by increased boreal summer insolation, the unprecedented modern-day warming is primarily caused by elevated greenhouse gas concentrations in the atmosphere. Furthermore, present-day warming is occurring at an unprecedented rate at high latitudes, making it the fastest observed warming during the entire Holocene period6,82. During the HTM, the ice sheet retreated inland, reaching a minimum extent farther inland than its current position. The warming predicted for this century is expected to drive even faster ice mass loss than what has been modeled for the last 12,000 years, including the HTM6,83. When considering this, the increased advection of Atlantic-derived waters to West Greenland will likely proceed at a faster pace during this century than during the mid-Holocene. Enhanced advection of warmer, saltier Atlantic-derived waters will drive significant changes in the distribution of ocean heat and salinity. These changes are expected to accelerate ice sheet retreat, amplify sea ice loss, and intensify water-column stratification. Such shifts will have profound and potentially detrimental impacts on marine ecosystems, altering biodiversity and food-web dynamics.
Methods
Sediment trap
The automated Technicap PPS 4/3 sediment trap GF10 was deployed at a site that was ice-free throughout the sampling period, apart from drifting ice mélange from the inner part of the fjord system during early May to June and from late October to November. The sediment trap samples were collected over a full year at a 20- to 45-day interval. Prior to the deployment, the trap collector cups were filled with 100% formalin and sodium borate with salinity adjusted to 37. The samples were wet split into eight subsamples using a HAVER RPT Rotating Sample Reducer at Aarhus University. We used two of the eight subsamples to conduct dinocyst and geochemical analysis.
Geochronology and grain-size distribution
The chronology for Rumohr-lot core SA13-ST6-36R was based on 10Pb and 137Cs and presented in Oksman et al.42. For gravity core SA13-ST6-40G the chronology is based on 210Pb, 137Cs, and 11 AMS 14C dates done on mixed benthic foraminifera (minimum 0.5 mg) and polychaete tubes (Table 2). Polychaetes burrow into the sediments, and careful assessment was made to determine the uppermost layer penetrated by the tube, as this represents the layer inhabited by polychaetes. The 210Pb activity was measured at the Department of Geosciences and Natural Resource Management, University of Copenhagen, using a Canberra ultralow-background Ge-detector using the Constant Rate of Supply (CRS) model and validated by 137Cs data84.
The AMS 14C dates were obtained from the accelerator mass spectrometry (AMS) facilities at the Aarhus AMS Center in Denmark (AAR in Table 2) and the Laboratory of Ion Beam Physics in Zurich in Switzerland (ETH in Table 2). The radiocarbon dates were calibrated using the Marine20 radiocarbon calibration curve85 and a local reservoir correction (ΔR) of −70 ± 54 years86 before age-depth modeling in OxCal 4.487,88.
The grain-size analyses were performed on 37 samples at the Geological Survey of Denmark and Greenland (GEUS) through wet sieving freeze-dried sediment (ca. 2 gram) to obtain the percentages of clay and silt (<63 μm), fine sand (63–150 μm), and coarse sand (> 150 μm).
Biogeochemical analysis
The biogenic silica (BSi) content was measured in 72 samples at GEUS. Freeze-dried samples (30 ± 1 mg) were leached with 40 ml of 1% sodium carbonate (Na2CO3) in a warm water bath (85 °C) for 5 hours. The samples were subsampled (1 ml) and mixed with 0.021 M HCL (9 ml) after 3, 4, and 5 h of extraction89. The subsamples were measured on a Perkin Elmer lambda 25UV/VIS spectrophotometer following the blue ammonium molybdate method90. The average was used to calculate the BSi concentration, assuming that all BSi is dissolved after two hours of extraction91. The BSi flux was calculated by multiplying the sediment mass accumulation rate with the BSi concentration. The sediment mass accumulation rate was found by multiplying the sediment accumulation rate with the dry bulk density of the sediment.
The total organic carbon (TOC), total nitrogen (TN), and stable isotopic composition (δ13C and δ15N) of organic matter on the sediment cores were analyzed in 83 samples at the RMS lab, Department of Geosciences and Natural Resource Management, University of Copenhagen. Homogenized freeze-dried samples (between 20 and 30 mg) were treated with 10% hydrochloric acid (HCl) to remove inorganic carbon before being measured on an elemental analyzer coupled to a Finnigan MAT Delta PLUS or Thermo Delta V Advantage isotope ratio mass spectrometer. The TOC flux was calculated by multiplying the sediment mass accumulation rate with the TOC concentration. The TOC and mass flux analyses for the sediment trap samples were done at GEUS and are described in Luostarinen et al.48.
Dinocysts and other palynomorphs
Palynological microscopy slides from core SA13-ST6-40G were prepared at Ghent University at a 4–8 cm resolution (number: 75 slides), whereas the slides from SA13-ST6-36R (number: 7 slides), and the sediment trap (number: 12 slides), were prepared at GEUS. One tablet (batch no. for core SA13-ST6-40G: 161018201 and 110118351 and batch no. for core SA13-ST6-36R: 110118351) with Lycopodium spores (containing 10000 spores) were added to each freeze-dried sample (minimum 1.5 grams of sediment). Palynological treatment consisted of treating the samples with 2 M HCl and 40% hydrofluoric acid (HF) (both at room temperature) to remove carbonates and silicates. After adding deionized water, the sediment samples were sonicated and sieved with a nylon mesh (10–11 µm for the core samples, and 6 µm for the trap samples) and gently centrifuged92. The samples were mounted on microscope slides with glycerin jelly and counted under an Olympus BX60 light microscope at magnifications of 400x and 1000x. The slides are stored at GEUS. To obtain representative relative and absolute abundances, a minimum of 300 dinocysts (Supplementary Figs. 2 and 3) or 500 Lycopodium marker spores were counted per slide, and the slides were scanned for rare species93. Other palynomorphs (e.g., acritarch’s, pollen, copepod eggs, foraminiferal linings, and Halodinium spp.) were also counted, and their concentrations were calculated following the marker-grain method. The permanent microscope slides from the Holocene sediment record and the sediment trap samples are archived at GEUS, Department of Glaciology and Climate.
Statistical analyses
To define stratigraphic zones in the gravity core, an age-constrained cluster analysis was performed on the dinocyst data using the open-source software R94 and the Rioja package95. This method is used to find temporal clusters of similar dinocyst composition. A constrained hierarchical clustering was applied, and the broken stick method was used to determine the number of clusters96 resulting in six clusters being retained. Then a principal component analysis (PCA) was carried out to describe the main patterns in the dataset and reduce the data dimensions. This was done by transforming the relative dinocyst abundance data via a Hellinger transformation97 and running the PCA using the vegan package98 for R94. The PCA1 explained ca. 33% of the variations in the dataset (eigenvalue: 4.54 and proportion of variance explained: 0.33), and PCA2 explained ca. 19% of the variations in the dataset (eigenvalue: 2.66 and proportion of variance explained: 0.19) (Supplementary Fig. 4).
The scores from the PCA1 and PCA2 were plotted against time (Supplementary Fig. 5). To better detect trends in these ordination scores, a GAMs62 was used (Supplementary Fig. 5). GAMs allow us to estimate the local mean of time-series with uneven spacing and where variable accumulation rate is an issue99. Smooths were parameterized using a thin plate regression spline basis with 9 basis functions after identifiability constraints were absorbed into the basis. To estimate rates of change in dinocyst assemblages, the first derivatives of the estimated trend and their 95% simultaneous intervals were used. A statistically significant change is observed when the simultaneous interval of the first derivative does not include 099.
Distribution of N. labyrinthus in relation to sea-surface temperature and salinity
A distribution map was created to investigate the affinity of dinocyst N. labyrinthus to summer sea-surface salinity and temperature. This was done by using the “modern” dinocyst database that includes dinocyst assemblage data from 1968 sites across the Northern Hemisphere46. The dinocyst counts for N. labyrinthus were extracted from the dataset and plotted in Ocean Data View (ODV)100. Two distribution maps with the environmental parameters of summer sea-surface temperature and summer sea-surface salinity were plotted. The datasets are from the World Ocean Atlas 2018 (WOA18: https://www.ncei.noaa.gov/access/metadata/landing-page/bin/iso?id=gov.noaa.nodc:NCEI-WOA18)67,68,69. The World Ocean Atlas is a collection of oceanographic conditions based on in-situ measurements. The statistical mean from 1955 to 2017 with a spatial resolution of 1/4-degree was plotted in ODV100. The WOA18 dataset includes more data compared to WOA13 (from 1955 to 2012) that was used in the dinocyst database46.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All the proxy data presented in this manuscript are available in the supplementary material and a public data repository (GEUS Dataverse https://doi.org/10.22008/FK2/21WRYA).
References
Meredith, M. et al. Polar regions. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate (eds H.-O. Pörtner, H.-O. & D.C. Roberts, D. C. et al.) (Cambridge University Press, Cambridge, UK and New York, NY, USA, 2019).
Mankoff, K. D. et al. Greenland liquid water discharge from 1958 through 2019. Earth Syst. Sci. Data 12, 2811–2841 (2020).
Slater, T. et al. Increased variability in Greenland Ice Sheet runoff from satellite observations. Nat. Commun. 12, 1–9 (2021).
Box, J. E. et al. Greenland ice sheet climate disequilibrium and committed sea-level rise. Nat. Clim. Chang. 12, 808–813 (2022).
Kaufman, D. et al. Holocene global mean surface temperature, a multi-method reconstruction approach. Sci. Data 7, 1–13 (2020).
Axford, Y., De Vernal, A. & Osterberg, E. C. Past warmth and its impacts during the holocene thermal maximum in greenland. Annu. Rev. Earth Planet. Sci. 49, 279–307 (2021).
Limoges, A. et al. Learning from the past: impact of the Arctic Oscillation on sea ice and marine productivity off northwest Greenland over the last 9,000 years. Glob. Chang. Biol. 26, 6767–6786 (2020).
Ouellet-Bernier, M.-M., Vernal, A., De, Hillaire-Marcel, C. & Moros, M. Paleoceanographic changes in the Disko Bugt area, West Greenland, during the Holocene. Holocene 24, 1573–1583 (2014).
Perner, K., Moros, M., Jennings, A., Lloyd, J. M. & Knudsen, K. L. Holocene palaeoceanographic evolution off West Greenland. Holocene 23, 374–387 (2012).
Moros, M. et al. Surface and sub-surface multi-proxy reconstruction of middle to late Holocene palaeoceanographic changes in Disko Bugt, West Greenland. Quat. Sci. Rev. 132, 146–160 (2016).
Gibb, O. T., Steinhauer, S., Fréchette, B., de Vernal, A. & Hillaire-Marcel, C. Diachronous evolution of sea surface conditions in the Labrador Sea and Baffin Bay since the last deglaciation. Holocene 25, 1882–1897 (2015).
Seidenkrantz, M. S. et al. Hydrography and climate of the last 4400 years in a SW Greenland fjord: Implications for Labrador Sea palaeoceanography. Holocene 17, 387–401 (2007).
Larsen, N. K. et al. The response of the southern Greenland ice sheet to the Holocene thermal maximum. Geology 43, 291–294 (2015).
Young, N. E. & Briner, J. Holocene evolution of the western Greenland Ice Sheet: assessing geophysical Ice-Sheet models with geological reconstructions of ice-margin change. Quat. Sci. Rev. 114, 1–17 (2015).
Funder, S. et al. A 10.000-year record of arctic ocean sea-ice variability-view from the beach. Science 333, 747–750 (2011).
Kelly, M. A. & Thomas, V. L. Fluctuations of local glaciers in Greenland during latest Pleistocene and Holocene time. Quat. Sci. Rev. 28, 2088–2106 (2009).
Kaufman, D. et al. Recent warming reverses long-term arctic cooling. Science 325, 1236–1239 (2009).
IPCC. Sections. In: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [eds. Core Writing Team, Lee, H. & J. Romero]. 35–115 (IPCC, Geneva, Switzerland, 2023).
Cohen, J. & Barlow, M. The NAO, the AO, and global warming: how closely related? J. Clim. 18, 4498–4513 (2005).
Rigor, I. G., Wallace, J. M. & Colony, R. L. Response of sea ice to the Arctic oscillation. J. Clim. 15, 2648–2663 (2002).
Thompson, D. W. J. & Wallace, J. M. The Arctic oscillation signature in the wintertime geopotential height and temperature fields. Geophys. Res. Lett. 25, 1297–1300 (1998).
Morley, A., Rosenthal, Y. & DeMenocal, P. Ocean-atmosphere climate shift during the mid-to-late Holocene transition. Earth Planet. Sci. Lett. 388, 18–26 (2014).
Sarafanov, A. On the effect of the North Atlantic Oscillation on temperature and salinity of the subpolar North Atlantic intermediate and deep waters. ICES J. Mar. Sci. 66, 1448–1454 (2009).
Box, J. E. Survey of greenland instrumental temperature records: 1873-2001. Int. J. Climatol. 22, 1829–1847 (2002).
Darby, D. A., Ortiz, J. D., Grosch, C. E. & Lund, S. P. 1,500-year cycle in the Arctic Oscillation identified in Holocene Arctic sea-ice drift. Nat. Geosci. 5, 897–900 (2012).
Staines-Urías, F., Kuijpers, A. & Korte, C. Evolution of subpolar North Atlantic surface circulation since the early Holocene inferred from planktic foraminifera faunal and stable isotope records. Quat. Sci. Rev. 76, 66–81 (2013).
Larsen, N. K. et al. Rapid early Holocene ice retreat in West Greenland. Quat. Sci. Rev. 92, 310–323 (2014).
Young, N. E. et al. In situ cosmogenic 10Be-14C-26Al measurements from recently deglaciated bedrock as a new tool to decipher changes in Greenland Ice Sheet size. Clim 17, 419–450 (2021).
Weidick, A., Bennike, O., Citterio, M. & Nørgaard-Pedersen, N. Neoglacial and historical glacier changes around Kangersuneq fjord in southern West Greenland. Geological Survey of Denmark and Greenland Bulletin. https://doi.org/10.34194/geusb.v27.4694 (2012).
Rysgaard, S. et al. An updated view on water masses on the pan-West Greenland continental shelf and their link to proglacial fjords. J. Geophys. Res. Ocean. 125, 1–10 (2020).
Meire, L. et al. High export of dissolved silica from the Greenland Ice Sheet. Geophys. Res. Lett. 43, 9173–9182 (2016).
Hopwood, M. J. et al. Non-linear response of summertime marine productivity to increased meltwater discharge around Greenland. Nat. Commun. 9, 1–9 (2018).
Mortensen, J., Bendtsen, J., Lennert, K. & Rysgaard, S. Seasonal variability of the circulation system in a west Greenland tidewater outlet glacier fjord, Godthåbsfjord (64°N). J. Geophys. Res. Earth Surf. 300, 316 (2014).
Mortensen, J., Lennert, K., Bendtsen, J. & Rysgaard, S. Heat sources for glacial melt in a sub-Arctic fjord (Godthåbsfjord) in contact with the Greenland Ice Sheet. J. Geophys. Res. Ocean. 116, 1–13 (2011).
Mortensen, J. et al. Local coastal water masses control heat levels in a West Greenland tidewater outlet glacier Fjord. J. Geophys. Res. Ocean. 123, 8068–8083 (2018).
Holland, D. M., Thomas, R. H., De Young, B., Ribergaard, M. H. & Lyberth, B. Acceleration of Jakobshavn Isbr triggered by warm subsurface ocean waters. Nat. Geosci. 1, 659–664 (2008).
Lin, P., Pickart, R. S., Torres, D. J. & Pacini, A. Evolution of the freshwater coastal current at the Southern Tip of Greenland. J. Phys. Oceanogr. 48, 2127–2140 (2018).
Hopwood, M. J. et al. Review article: How does glacier discharge affect marine biogeochemistry and primary production in the Arctic? Cryosphere 14, 1347–1383 (2020).
Juul-Pedersen, T. et al. Seasonal and interannual phytoplankton production in a sub-Arctic tidewater outlet glacier fjord, SW Greenland. Mar. Ecol. Prog. Ser. 524, 27–38 (2015).
Meire, L. et al. Marine-terminating glaciers sustain high productivity in Greenland fjords. WILEY, Glob. Chang. Biol. 23, 5344–5357 (2017).
Meire, L. et al. Glacier retreat alters downstream fjord ecosystem structure and function in Greenland. Nat. Geosci. 16, 671–674 (2023).
Oksman, M. et al. Impact of freshwater runoff from the southwest Greenland Ice Sheet on fjord productivity since the late 19th century. Cryosphere 16, 2471–2491 (2022).
Oksman, M. et al. Climate variability and glacier dynamics linked to fjord productivity changes over the last ca. 3300 years in Nuup Kangerlua, Southwest Greenland. Paleoceanogr. Paleoclimatology 39, 1–20 (2024).
Head, M. J. Chapter 30. Modern dinoflagellate cysts and their biological affnities. Jansonius, J. McGregor, D.C. (ed.), Palynol. Princ. Appl. Am. Assoc. Stratigr. Palynol. Found. 3 1197–1248 (1996).
Zonneveld, K. A. F. et al. Atlas of modern dinoflagellate cyst distribution based on 2405 data points. Rev. Palaeobot. Palynol. 191, 1–197 (2013).
de Vernal, A. et al. Distribution of common modern dinoflagellate cyst taxa in surface sediments of the Northern Hemisphere in relation to environmental parameters: The new n=1968 database. Mar. Micropaleontol. 159, 101796 (2020).
Dale, B. The sedimentary record of dinoflagellate cysts: Looking back into the future of phytoplankton blooms. Sci. Mar. 65, 257–272 (2001).
Luostarinen, T. et al. An annual cycle of diatom succession in two contrasting Greenlandic fjords: from simple sea-ice indicators to varied seasonal strategists. Mar. Micropaleontol. 158, 101873 (2020).
Seidenkrantz, M., Røy, H., Lomstein, B. A. & Meire, L. Cruise Report: Godthåbsfjord system and the West Greenland shelf with ‘R / V Sanna’. 11–16. August 2013 (Fieldreport, 2013).
Andreasen, N. et al. From land to sea: provenance, composition, and preservation of organic matter in a marine sediment record from the North-East Greenland shelf spanning the Younger Dryas–Holocene. Boreas 52, 459–475 (2023).
Kienast, S. S., Calvert, S. E. & Pedersen, T. F. Nitrogen isotope and productivity variations along the northeast Pacific margin over the last 120 kyr: surface and subsurface paleoceanography. Paleoceanography 17, 7-1–7–17 (2002).
Lamb, A. L., Wilson, G. P. & Leng, M. J. A review of coastal palaeoclimate and relative sea-level reconstructions using δ13C and C/N ratios in organic material. Earth Sci. Rev. 75, 29–57 (2006).
Belicka, L. L. & Harvey, R. H. The sequestration of terrestrial organic carbon in Arctic Ocean sediments: a comparison of methods and implications for regional carbon budgets. Geochim. Cosmochim. Acta 73, 6231–6248 (2009).
Naidu, A. S. et al. Organic carbon isotope ratios (d13C) of Arctic Amerasian continental shelf sediments. Int. J. Earth Sci. 89, 522–532 (2000).
Dahl-Jensen, D. et al. Past temperatures directly from the Greenland Ice Sheet. Sci.ence 282, 268–271 (1998).
Vinther, B. M. et al. Holocene thinning of the Greenland ice sheet. Nature 461, 385–388 (2009).
Head, M. J., Harland, R. & Matthiessen, J. Cold marine indicators of the late Quaternary: The new dinoflagellate cyst genus Islandinium and related morphotypes. J. Quat. Sci. 16, 621–636 (2001).
Kunz-Pirrung, M. Dinoflagellate cyst assemblages in surface sediments of the Laptev Sea region (Arctic Ocean) and their relationship to hydrographic conditions. J. Quat. Sci. 16, 637–649 (2001).
De Vernal, A. et al. Reconstruction of sea-surface conditions at middle to high latitudes of the Northern Hemisphere during the Last Glacial Maximum (LGM) based on dinoflagellate cyst assemblages. Quat. Sci. Rev. 24, 897–924 (2005).
Ribeiro, S., Moros, M., Ellegaard, M. & Kuijpers, A. Climate variability in West Greenland during the past 1500 years: evidence from a high-resolution marine palynological record from Disko Bay. Boreas 41, 68–83 (2012).
Seidenkrantz, M. S. et al. Early Holocene large-scale meltwater discharge from Greenland documented by foraminifera and sediment parameters. Palaeogeogr. Palaeoclimatol. Palaeoecol. 391, 71–81 (2013).
Hastie, T. J. & Tibshirani, R. J. Generalized Additive Models. (CHAPMAN & HALL/CRC, 1990).
Hastie, T. & Tibshirani, R. Generalized additive models. Stat. Sci. 1, 297–318 (1986).
Kumar, V., Tiwari, M., Nagoji, S. & Tripathi, S. Evidence of anomalously low δ13C of marine organic matter in an Arctic Fjord. Sci. Rep. 6, 1–9 (2016).
Holding, J. M. et al. Autochthonous and allochthonous contributions of organic carbon to microbial food webs in Svalbard fjords. Limnol. Oceanogr. 62, 1307–1323 (2017).
Heikkilä, M. et al. Surface sediment dinoflagellate cysts from the Hudson Bay system and their relation to freshwater and nutrient cycling. Mar. Micropaleontol. 106, 79–109 (2014).
Boyer, T. P. et al. World Ocean Atlas 2018. [temperature and salinity]. NOAA National Centers for Environmental Information. Dataset. Available at: https://www.ncei.noaa.gov/archive/accession/NCEI-WOA18. Accessed [10.03.22] (2018).
Locarnini, R. A. et al. World Ocean Atlas 2018, Volume 1: Temperature. A. Mishonov Tech. Ed. 52 (NOAA National Centers for Environmental Information, 2019).
Zweng, M. M. et al. World Ocean Atlas 2018, Volume 2: Salinity. A. Mishonov Tech. Ed. 50 (NOAA National Centers for Environmental Information, 2019).
Caron, M., Rochon, A., Montero-Serrano, J. C. & St-Onge, G. Evolution of sea-surface conditions on the northwestern Greenland margin during the Holocene. J. Quat. Sci. 34, 569–580 (2019).
Hansen, K. E., Giraudeau, J., Wacker, L., Pearce, C. & Seidenkrantz, M. S. Reconstruction of Holocene oceanographic conditions in eastern Baffin Bay. Clim 16, 1075–1095 (2020).
Saini, J. et al. Holocene variability in sea ice and primary productivity in the northeastern Baffin Bay. Arktos 6, 55–73 (2020).
Allan, E. et al. Insolation vs. meltwater control of productivity and sea surface conditions off SW Greenland during the Holocene. Boreas 50, 631–651 (2021).
Briner, J. P., Kaufman, D. S., Bennike, O. & Kosnik, M. A. Amino acid ratios in reworked marine bivalve shells constrain Greenland Ice Sheet history during the Holocene. Geology 42, 75–78 (2014).
Orme, L. C. et al. Subpolar North Atlantic sea surface temperature since 6 ka BP: Indications of anomalous ocean-atmosphere interactions at 4-2 ka BP. Quat. Sci. Rev. 194, 128–142 (2018).
Brice, C., de Vernal, A., Francus, P., Forwick, M. & Nam, S. Il. Millennial-scale oscillations and an environmental regime shift around the Middle to Late Holocene transition in the North Atlantic region based on a multiproxy record from Isfjorden, West Spitsbergen. Boreas 52, 42–59 (2023).
Lloyd, J. M., Kuijpers, A., Long, A., Moros, M. & Park, L. A. Foraminiferal reconstruction of mid- to late-Holocene ocean circulation and climate variability in Disko Bugt, West Greenland. Holocene 17, 1079–1091 (2007).
Gkinis, V., Simonsen, S. B., Buchardt, S. L., White, J. W. C. & Vinther, B. M. Water isotope diffusion rates from the NorthGRIP ice core for the last 16,000 years—Glaciological and paleoclimatic implications. Earth Planet. Sci. Lett. 405, 132–141 (2014).
Weiss, H., Kuijpers, A. & Fisher, D. The 4.2 ka BP event: Global rearrangement of ocean-atmosphere interaction patterns forced Eastern Hemisphere societal collapses. Abstr. EGU Gen. Assem. Vienna, EGU 23-3591. https://doi.org/10.5194/egusphere-egu23-3591 (2023).
Jeong, H., Park, H. S., Stuecker, M. F. & Yeh, S. W. Distinct impacts of major El Niño events on Arctic temperatures due to differences in eastern tropical Pacific sea surface temperatures. Sci. Adv. 8, 1–11 (2022).
Mortensen, J. et al. On the seasonal freshwater stratification in the proximity of fast-flowing tidewater outlet glaciers in a sub-Arctic sill fjord. J. Geophys. Res. Ocean. 118, 1382–1395 (2013).
Lecavalier, B. S. et al. High Arctic Holocene temperature record from the Agassiz ice cap and Greenland ice sheet evolution. Proc. Natl Acad. Sci. USA 114, 5952–5957 (2017).
Briner, J. P. et al. Rate of mass loss from the Greenland Ice Sheet will exceed Holocene values this century. Nature 586, 70–74 (2020).
Andersen, T. J. In Applications of Paleoenvironmental Techniques in Estuarine Studies. 121–140 (Springer Nature, 2017).
Heaton, T. J. et al. Marine20—The marine radiocarbon age calibration curve (0-55,000 cal BP). Radiocarbon 62, 779–820 (2020).
Pieńkowski, A. J., Coulthard, R. D. & Furze, M. F. A. Revised marine reservoir offset (ΔR) values for molluscs and marine mammals from Arctic North America. Boreas. https://doi.org/10.1111/bor.12606 (2022).
Ramsey, B. C. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Ramsey, C. & Lee, S. Recent and planned developments of the program OxCal. Radiocarbon 55, 720–730 (2013).
DeMaster, D. J. Measuring biogenic silica in marine sediments and suspended matter. In Marine Particles: Analysis and Characterization. Geophysical Monograph Series Vol. 63. Am. Geophys. Union (eds. Hurd, D. C. & Spencer, D. W.) 363–367 (AGU publications, Washington, D. C, 1991).
Mullin, J. & Riley, J. The colorimetric determination of silicate with special reference to sea and natural waters. Anal. Chim. Acta 12, 162–176 (1955).
Barão, L. et al. Alkaline-extractable silicon from land to ocean: a challenge for biogenic silicon determination. Limnol. Oceanogr. Methods 13, 329–344 (2015).
Quaijtaal, W., Donders, T. H., Persico, D. & Louwye, S. Characterising the middle Miocene Mi-events in the Eastern North Atlantic realm: a first high-resolution marine palynological record from the Porcupine Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 399, 140–159 (2014).
Mertens, K. N. et al. Determining the absolute abundance of dinoflagellate cysts in recent marine sediments: the Lycopodium marker-grain method put to the test. Rev. Palaeobot. Palynol. 157, 238–252 (2009).
R Core Team. R: A Language and Environment for Statistical Computing. https://www.R-project.org/. (R Foundation for Statistical Computing, Vienna, Austria2021).
Juggins, S. rioja: Analysis of Quaternary Science Data, R Package Version (0.9-26). (https://cran.r-project.org/package=rioja) (2020).
Bennett, K. D. Determination of the number of zones in a biostratigraphical sequence. N. Phytol. 132, 155–170 (1996).
Legendre, P. & Gallagher, E. Ecologically meaningful transformations for ordination of species data. Oecologia 2, 271–280 (2001).
Oksanen, J. et al. W. J. _vegan: Community Ecology Package_. R package version 2.6-2, https://CRAN.R-project.org/package=vegan (2022).
Simpson, G. L. Modelling palaeoecological time series using generalised additive models. Front. Ecol. Evol. 6, 1–21 (2018).
Schlitzer, R. Ocean Data View. Available at: odv.awi.de. (2023).
Larsen, N. K. et al. Strong altitudinal control on the response of local glaciers to Holocene climate change in southwest Greenland. Quat. Sci. Rev. 168, 69–78 (2017).
Perner, K. et al. An oceanic perspective on Greenland’s recent freshwater discharge since 1850. Sci. Rep. 9, 1–11 (2019).
Acknowledgements
The required permits for geological investigations, collection, and export of sediments were obtained from the Mineral License and Safety Authority, Government of Greenland. This study was funded by Geocenter Denmark (project Greenshift) and the Danish Independent Research Fund (grant 9064-00039B to S.R.). M.H. received funding from the Research Council of Finland (grant 334509), while M.-S.S. and C.P. have received funding through the European Union’s Horizon 2020 research and innovation program under Grant Agreement No. 869383 (ECOTIP) and the European Union’s Horizon Europe research and innovation program under Grant Agreement No. 101136480 (SEA-Quester). We thank Charlotte Olsen for laboratory assistance.
Author information
Authors and Affiliations
Contributions
A.B.K. wrote the manuscript and generated all figures with input from all co-authors. S.R. conceived the study and provided extensive feedback and comments on the manuscript. M.-S.S. and C.P. provided the marine sediment core material and geochronology. L.M. was responsible for the sediment trap deployment and recovery. M.H. provided the dinocyst trap dataset. A.B.K., S.R., and M.H. contributed to the interpretation of the datasets. G.L.S. contributed with expert knowledge of the statistical methods. N.K.L. contributed with expert knowledge on the area and provided terrestrial context to the interpretations. A.K. contributed with expert knowledge of the area and the broader palaeoceanographic context. All co-authors commented on the manuscript and approved its content.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Thomas R. Lakeman and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Carolina Ortiz Guerrero. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kvorning, A.B., Heikkilä, M., Pearce, C. et al. A Holocene fjord record from Greenland reveals exceptional Atlantic water influence during minimum ice-sheet extent. Commun Earth Environ 6, 326 (2025). https://doi.org/10.1038/s43247-025-02282-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02282-5
This article is cited by
-
Mid-Holocene retreat of the Greenland Ice Sheet indicated by subglacial methane release
Nature Geoscience (2026)
