
Abstract
Biodiversity Hotspots (Hotspots), harboring exceptionally rich small-ranged species, are critical for mitigating biodiversity loss. As priorities for terrestrial conservation, Hotspots increasingly face threats from agriculture, the largest anthropogenic disturbance impacting biodiversity. Yet, the spatial dynamics of agricultural expansion and its impacts on biodiversity, especially small-ranged vertebrates, remain poorly understood. Using site-level observations and satellite imagery, we found that agricultural pressures reduce species richness by 25.8%, total abundance by 12.4%, and rarefied species richness by 8.7% relative to primary vegetation within Hotspots. However, cropland area within Hotspots expanded 12% from 2000–2019, exceeding the global average of 9%. Fine-scale analysis identified 3,483 risk spots (cropland expansion and high small-ranged vertebrate richness, ~1741 Mha); ~1031 Mha of these areas fall outside Protected Areas, particularly in the Atlantic Forest, Indo-Burma, Western Ghats, Sri Lanka, and Sundaland. These results underscore the urgent need for targeted conservation actions to prevent biodiversity loss from agricultural expansion.
Similar content being viewed by others

Introduction
Global biodiversity loss has been occurring at unprecedented rates in the Anthropocene1,2,3,4, already surpassing the planetary boundary5,6, and is becoming a major threat to human well-being7,8,9,10. Halting biodiversity loss is a global concern and the focus of numerous international commitments, particularly the recent Kunming-Montreal Global Biodiversity Framework (GBF)11. Given the need to optimally allocate scarce conservation resources and address trade-offs with other demands on land12,13,14,15, it has been proposed to focus on protecting species with small ranges16,17, which are critical for avoiding species extinction13,16,17,18,19.
Biodiversity Hotspots (hereinafter referred to as Hotspots) are regions that contain more than 1500 species of endemic vascular plants and have lost more than 70% of their primary native vegetation13,20,21. Not only that, but Hotspots also house the most undiscovered species22,23. Therefore, effective protection of Hotspots can result in preventing the triggering of a new episode of global extinction. However, the trend and pattern of human disturbances, as well as the effectiveness of conservation measures within Hotspots, remain unclear.
Agriculture, as a long-standing and major human activity, has long been identified as the largest driver of biodiversity collapse24,25,26,27,28, through its disruption, fragmentation, and elimination of natural habitats. To meet the demands of increasing food security29, global cropland area increased by 9% during 2000–201930, and this trend is expected to continue in the future31,32,33. However, the expansion of cropland within Hotspots is not yet clear. Closing this knowledge gap is essential for evaluating agricultural threats to small-range and yet undiscovered species. Furthermore, the impacts of agricultural expansion may vary in different regions34, with different intensities of cropland use. But the impacts of agricultural expansion on biodiversity within Hotspots have yet to be documented. This knowledge gap hinders our ability to implement effective measures to address biodiversity loss caused by agricultural expansion.
Establishing Protected Areas (PAs) has been considered as one effective solution for the long-term conservation of biodiversity35,36,37,38. The Aichi target aimed to generate 17% PA coverage of terrestrial habitats by 202039, but has not yet been fully achieved40. This poses a major challenge to the realization of GBF Target 3, which commits to protecting at least 30% of the world’s land and ocean areas by 2030 (30 × 30 Target)11,41,42. More concerningly, Hotspots should theoretically be prioritized for PA establishment, while the designated PAs previously have been largely biased toward non-Hotspots40, limiting their effectiveness in safeguarding small-ranged species. So, guidance is urgently needed to identify conservation priorities based on small-ranged vertebrates to mitigate agricultural threats to biodiversity within Hotspots, achieving the ambitious conservation commitments planned for the immediate future21,43.
To fill the aforementioned knowledge gaps, here we examined the increasing agricultural threat within Hotspots, identified the risk spots by combining cropland expansion and small-ranged species richness, and provided support for future PA designation. First, we employed the PREDICTS database44, a terrestrial assemblage database of unprecedented geographic and taxonomic coverage, to evaluate biodiversity responses to agricultural pressure at the ecological community level within Hotspots. Second, building upon these in-situ findings, we characterized the agricultural expansion pressure within Hotspots by using remotely sensed high-resolution cropland data. Third, the risk spots were identified by overlaying cropland expansion with high-resolution layers of small-ranged vertebrates (i.e., species with smaller than the median geographical range size), including a total of 9124 species (mammals: 2164; birds: 4796; amphibians: 2164). Finally, the effectiveness and priorities of conservation (i.e., PA) within Hotspots were documented. The knowledge generated by our study aims to understand the status of Hotspots in resisting agricultural activities, guide conservation efforts within Hotspots, and eventually prevent species extinction, even before species are discovered.
Results
Effects of agricultural pressure on site-level diversity within Hotspots
The analysis of the PREDICTS database revealed that agricultural expansion and intensification threaten the authenticity and distinctiveness of ecological community species composition within Hotspots. Specifically, the species diversity in croplands was lower than that in primary vegetation, and this pattern became more pronounced as the intensity of cropland use increased (Fig. 1).

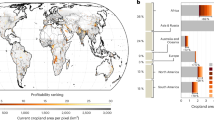
a Location of PREDICTS sites (blue points) within Hotspots (red area). b–d Each panel illustrates a linear or generalized linear mixed-effects model applied to species richness (Richness), total abundance (Abundance), and rarefied species richness (RSR). Symbols denote land use intensity: circles for minimal use, triangles for light use, and squares for intense use. Effect sizes were adjusted to a percentage by drawing fixed effects 1000 times based on the variance-covariance matrix, expressing each fixed effect within each random draw as a percentage of the baseline (primary vegetation minimal use), then calculating points to show the median value and error bars to show 95% confidence intervals.
Species richness in croplands declined from −7.26% at minimally used to −43.52% at intensely used. Total species abundance in croplands at minimally used was higher than the baseline (primary vegetation minimal usage), but significantly decreased with increasing cropland use intensity, dropping to −26.90% at intensely used croplands. Rarefied species richness (defined as the fewest individuals at any site in each study) also significantly decreased with increasing cropland use intensity, dropping from −3.19% at minimally used to −24.67% at intensely used croplands.
Characterizing cropland changes within Hotspots
An analysis based on the remote sensing-based cropland data at 30 m resolution shows a remarkable cropland expansion within Hotspots from 2000 to 2019, particularly acute in the tropics. From 2000 to 2019, the cropland area increased by 12% (36 million hectares, Mha) within Hotspots.
Both Kernel Density Estimation (KDE) and Optimized Hot Spot Analysis (HSA) (see “Methods”) show the cropland expansion within the Hotspots, accompanied by the loss of natural habitats. Cropland expansion was more significant in developing countries than in developed countries, particularly in countries within Eastern South America, south-central and northwestern Indo-Burma, the Middle East, Central Asia, and eastern Africa, which experienced the most substantial cropland expansions (Fig. 2a, b). Those regions generally have larger amounts of originally undeveloped natural habitats. The cropland contraction within the Hotspots from 2000 to 2019 mainly occurred in the developed regions (e.g., the northern region of the Mediterranean, coastal regions of the United States, and the Caribbean). Notably, both the expansion and contraction of cropland existed simultaneously in the Indo-Burma (Fig. 2a, b).

a Kernel Density Estimation of cropland changes; b Hot Spot Analysis of cropland changes. The Gi_Bin identifies statistically significant hot spots (red) and cold spots (blue). Statistical significance was based on the p-value and z-score (two-sided). c Cropland change area (bars) and percentage (connected points) within Hotspots.
Out of 36 Hotspots, 30 experienced cropland expansion, of which 18 were in tropical regions (Fig. 2c). The Cerrado, one of the most important and vulnerable worldwide arenas for the conflict between food production and biodiversity conservation, experienced the largest expansion of cropland area. Compared to 2000, the Cerrado’s cropland area increased by 10 Mha by 2019, which was nearly twice that of the second-ranking region (Atlantic Forest, also home to many distinctive and threatened species). In contrast, six Hotspots experienced a noticeable cropland contraction, of which five are situated in the temperate regions.
Types of cropland expansion within Hotspots
Overlaying the cropland changes with the original land cover data around the starting point of our study period (~2000, detailed information in Table S1), we found that the regions with the largest areas of cropland expansion were mainly at the expense of natural habitats, particularly in the Cerrado, Atlantic Forest, and eastern Africa. At the same time, the minimal-intensity cropland expansion, which can drive the biotic homogenization within Hotspots, was also severe, mainly located in the Indo-Burma and African regions (Fig. 3). Specifically, the Mosaic forest & cropland (Light) has seen the largest area increase, amounting to ~7.5 Mha. This was followed by the Mosaic forest & cropland (Minimal) with an increase of ~3.8 Mha. These lands were mainly located in the Cerrado and Atlantic Forest. The Primary_CS land (land is primary vegetation now, but suitable for cropland) had the highest cropland growth rate, reaching 116%, with an increase of 3.5 Mha, primarily in the Sundaland and eastern Africa.

Map of cropland changes between different LUI, and statistics on cropland changes area and percent between different LUI within Hotspots; Primary_CS indicates that land is primary vegetation now but suitable for cropland.
Since Hotspots are mainly located in tropical developing countries, where agricultural intensification levels are relatively low, the cropland with minimal use intensity has seen the largest increase, reaching 3.1 Mha (Fig. 3). This was followed by cropland with light use intensity, increasing by ~2.2 Mha, mainly in the Indo-Burma. However, cropland with intense use had shown a slight decrease, with a reduction of ~100 thousand ha. Considering this together with Fig. 1, cropland expansion at the expense of natural habitats within Hotspots may contribute to reductions in species richness, abundance, and rarefied species richness at the local scale, which could pose substantial risks to biodiversity. Consequently, identifying risk spots where cropland changes threaten small-ranged vertebrates was crucial.
Threats to biodiversity from cropland expansion within Hotspots
Spatial correlation analysis between the distribution of cropland changes and the distribution of small-ranged vertebrates showed that there are 3483 risk spots (i.e., H-H and L-H clusters, meaning cropland changes and high concentrations of small-ranged vertebrates) in Hotspots, accounting for 64.18% Hotspots, particularly small-ranged mammals (3326 risk spots, 61.29%) and small-ranged birds (3529 risk spots, 65.03%) (Fig. 4). At the same time, amphibians show relatively few risk spots.

Cropland changes with small-ranged species richness of vertebrates (a), mammals (b), birds (c), and amphibians (d). They are calculated using local indicators of spatial association (LISA) at 5000 km²/ Hexagon. H-H clusters indicate hotspot locations (red), in which areas with high cropland expansion were significantly associated with high species richness of small-ranged vertebrates (at 0.05 significance level). L-L clusters (blue) show cold spot locations, in which areas with low cropland expansion were significantly associated with low species richness of small-ranged vertebrates. L-H and H-L clusters show an inverse spatial association. Small-ranged vertebrates represent the sum of small-ranged mammals, small-ranged birds, and small-ranged amphibians.
Specifically, the Atlantic Forest, Eastern African, Southern African, and Madagascar Islands, Eastern Himalayan, and the Indo-Burma were characterized by large cropland expansion and abundant small-ranged vertebrate species richness (i.e., H-H), especially small-ranged mammals and birds (Fig. 4b, c), suggesting that agricultural activities should be constrained in those regions.
Conservation gaps and conservation priorities within Hotspots
As agricultural pressures mount, it is vital to determine whether existing PAs within Hotspots can function as refuges for the vertebrates’ survival and where Hotspots might efficiently expand their protection. We calculated the percentage of PA area within Hotspots (Fig. 5a) and the conservation gap (Fig. 5b) (see “Methods”).

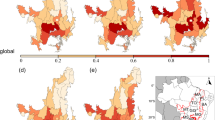
Conservation status (a), gaps (b), and conservation priorities of vertebrates (c), mammals (d), birds (e), and amphibians (f).
The conservation gaps were defined as the areas of net risk spots for small-ranged vertebrates (i.e., risk spots not covered by PAs). There is a total of 13 regions within Hotspots, such as the Cerrado, Atlantic Forest, Indo-Burma, Eastern Himalayan, Sundaland, Western Ghats and Sri Lanka, and Eastern Africa (existing cropland expansion hotspots), where PA coverage was less than 17% currently and 13 Hotspots had PA coverage of less than 30% (Fig. 5a). Meanwhile, a total of ~1031 Mha conservation gap remains within Hotspots, with net risk spots areas exceeding 30% of Hotspots’ area in the Atlantic Forest, Eastern Himalaya, Indo-Burma, Western Ghats, Sri Lanka, and Sundaland, over 17% in Eastern Africa, and below 17% in the Cerrado (Fig. 5b).
To prioritize lands for future PA expansion, we calculated species-level scores based on the proportion of unprotected range for small-ranged vertebrates within Hotspots, excluding cropland and urban areas, and aggregated the hexagonal grid (Fig. 5c–f) (see “Methods”). Summing up the priority scores across all taxa (Fig. 5c), the highest-priority regions were mostly in the Tropical Andes, Mesoamerica, Atlantic Forest, Eastern Afromontane, Madagascar, Western Ghats and Sri Lanka, Philippines, Sundaland, and Indo-Burma. Although the PA coverage was greater than 17% in the Tropical Andes, Central America, Eastern Afromontane, and Madagascar, there were still very high priority scores due to the exceptional richness of small-ranged vertebrates.
Discussion
Increasing agricultural pressures within Hotspots
We first analyzed the response of biodiversity metrics to varying cropland use intensities within Hotspots using the site-level PREDICTS database, and the results align with previous analyzes2,6,26,45. Although the total species abundance in minimal-intensity croplands within Hotspots was slightly higher than the baseline, it was accompanied by a decrease in species richness. This indicates a potential threat to the stability of ecological communities (Fig. 1). The effect of primary vegetation loss on biodiversity depends on the species’ habitat requirements46. Some species are susceptible to natural habitat loss because they depend on primary vegetation resources for shelter, feeding, and breeding, or they are adapted to the prevailing abiotic conditions47. However, other species tolerate and even proliferate in agricultural ecosystems47, because they can exploit additive resource subsidies across non-primary vegetation habitats46. Thus, land conversions can trigger a disturbance-sensitive/disturbance-tolerant species replacement47. In the long run, such winners-losers replacement can create a “negative feedback loop” that further reduces species richness, especially for undiscovered or small-range species that are not adapted to croplands.
Furthermore, our spatial analyzes showed an alarming cropland expansion within Hotspots, especially in the tropics, e.g., the Cerrado, Atlantic Forest, and Indo-Burma, concordant with previous research48,49,50,51. Cropland expansion is leading to the continued disruption or even loss of large areas of natural habitats, and many rarefied species are facing a dramatic reduction in living space within Hotspots52,53.
We further discovered a strong spatial correlation between cropland expansion and small-ranged vertebrates within Hotspots in many regions where widespread expansion of cropland poses a significant threat to overall biodiversity, including Central and South America, including the Tropical Andes and the Atlantic Forest, in the forests and savannahs of Central Africa and Madagascar, as well as in parts of South Africa, Eastern Australia and large portions of South-East Asia. Our analysis agrees with the previous efforts based on theoretical and empirical analyzes52,53,54,55,56.
Enhance PAs as vehicles for biodiversity conservation within Hotspots
Given the high biodiversity values of Hotspots16,23,57,58, and the agricultural threat to biodiversity documented in this study, more conservation measures should be implemented to prevent further habitat losses59. While PAs can mitigate biodiversity loss, their effectiveness for biodiversity conservation has been found to correlate most strongly with the density of guards60 and the level of anthropogenic pressure surrounding the PAs61,62. This study showed the insufficient PA coverage of vertebrate ranges within Hotspots, and a previous study also showed that most high-suitability habitats occurred outside of existing PAs63. Therefore, more PAs should be established urgently within Hotspots in a targeted manner to ensure the conservation of small-ranged and undiscovered species18, especially in the Atlantic Forest64, Tropical Andes65, Madagascar66, Sundaland, and Eastern Himalayan67.
In addition, cropland expansion is probably occurring not just in unprotected areas but also inside PAs68, suggesting that the management level of PAs located within Hotspots should be raised, and the internal patrolling activities of PAs should be strengthened69,70. Involving local communities in monitoring PAs could be an optional solution47,71. Conservation strategies must address the underlying drivers of cropland area expansion72. Administrative measures should also be taken to reduce domestic demand for more croplands, such as improving the productivity of existing areas. This may effectively reduce cropland expansion in PAs as a consequence.
Reconciling biodiversity conservation and food security
This study primarily focused on biodiversity conservation targets, aiming to curb cropland expansion, protect natural habitats, and mitigate the detrimental impacts of escalating food demand on biodiversity. However, food security is another global concern, especially for developing countries with limited food production capacity28. Therefore, the trade-off between food security and biodiversity conservation has become a topic of great concern29,38,73,74, also known as the trade-off between SDG 2 (“Zero Hunger”) and SDG 15 (“Life on Land”). We have observed that the PA gaps within Hotspots mainly happened in low-income countries, such as Brazil, Vietnam, and Ethiopia, characterized by rapid population and economic growth, as well as the demand for food production or export. For instance, in Brazil, cattle grazing in the Amazon and soy plantations in the Cerrado contribute to these challenges75.
Although international food trade could benefit biodiversity conservation and food security in low-income countries with high biodiversity76, it may also pose a threat to the biodiversity conservation efforts of food-exporting countries. Therefore, achieving balanced food and biodiversity security will require close cooperation between countries and organizations across the globe. Low-income countries should improve the productivity and efficiency of land use through advanced agricultural technologies and management measures. On the other hand, high-income countries should aim to increase food production and food exports to meet the food needs of low-income countries.
Concerningly, these cooperative efforts may be disrupted by geopolitical tensions and market fluctuations. For example, reduced access to fertilizer imports in key grain-exporting countries such as Brazil can substantially lower productivity in the short term, until alternative fertilizer supplies become available. Such disruptions may reshape food-trade policies and could, to some extent, accelerate cropland expansion56. Consequently, to avoid exacerbating deeply rooted inequalities, international organizations such as the UN, FAO, and WTO should actively coordinate to establish a fair and orderly international food trade order, to prevent food prices from rising, and to eliminate trade barriers to safeguard the rights and interests of low-income countries with limited resources for biodiversity protection.
Limitations and uncertainties
The PREDICT database used in this study exhibits taxonomic and geographic biases. As shown in Fig. S1, the data is heavily skewed toward birds and concentrated in the Mediterranean Basin, reflecting the uneven accumulation of research: birds are among the most studied taxa, and European countries in this region have strong research infrastructure and a culture of data sharing. These biases may, to some extent, influence the study’s conclusions. Future integration of emerging datasets, such as BioTIME and Living Planet Index, will be crucial for robust, site-level analyzes of how land-use change impacts biodiversity.
Historically, invertebrates, particularly the highly diverse insects, have been largely overlooked in conservation planning due to the lack of systematic research and available data77,78. At the same time, many countries have not timely updated their PA information, which may result in systematic biases in identifying conservation priorities40. Therefore, the estimates of priority Hotspots presented in this study should be considered as a current baseline. As more data becomes available for underrepresented taxa, this baseline is likely to increase. Future studies should build upon the existing framework by integrating new species data and dynamic distribution information, thereby enhancing the scientific rigor, comprehensiveness, and robustness of conservation planning across taxa and regions, and providing more reliable guidance for Hotspots.
Methods
Effects of land conversion on biodiversity within Hotspots
We used the PREDICTS database (https://data.nhm.ac.uk/dataset/the-2016-release-of-the-predicts-database-v1-1 and https://data.nhm.ac.uk/dataset/release-of-data-added-to-the-predicts-database-november-2022) to model responses of biodiversity to different land-use intensities on cropland2. The database contains variables for land-use intensity (minimal, low, and high) and land-use type (primary vegetation, secondary vegetation, plantation, pasture, cropland, and urban). Land-use intensity for each land-use type is defined based on a series of variables, including fertilizer and pesticide application, mechanization, and hunting79 (detailed information is provided in Table S1).
We performed a series of data preprocessing steps and retained a total of 12,153 data entries. First, we selected data entries where sites were located in Hotspots using the shapefile; removed all sites with unknown land use types and land use intensities; and excluded sites in secondary vegetation in an unknown stage of recovery. Next, we combined the factors of land use type and intensity (hereafter referred to as LUI) to create a single variable as described by Joseph Millard et al.45. We then calculated site-level biodiversity metrics, including species richness, total abundance, and rare species richness (defined as the fewest individuals at any site in each study), for different land use and intensities, as described by Tim Newbold et al.2. Finally, we performed a log(x + 1) transformation on the total abundance data to normalize model residuals.
We developed generalized linear mixed-effects models based on Poisson error distributions for the effects of LUI on species richness and rare species richness, and linear mixed-effects models for the effects of LUI on total abundance. Due to the nested nature of the database, random effects considered included study to account for variation in sampling methods, sampling effort, and broad geographic variation between studies, and block to account for the spatial arrangement of sites within the study. An additional (observation-level) random interception of site identity was included in the species richness and rare species richness model to control the over-dispersion present in species richness estimates. We checked for overdispersion in the species richness models using the function GLMEROverdispersion in the R package StatisticalModels. We compared each model against an intercept-only model and discarded any main model for which the AIC was greater than the null model.
Cropland change analyses within Hotspots
We used the 30-m global cropland time-series dataset developed by Potapov et al.30, which provides continuous maps for 2000–2019. Their cropland definition closely follows that of the Food and Agriculture Organization of the United Nations68. To reduce misclassification caused by fallow cycles, the dataset aggregates observations into five 4-year epochs (2000–2003, 2004–2007, 2008–2011, 2012–2015, and 2016–2019), yielding one representative cropland layer for each period (2003, 2007, 2011, 2015, and 2019)68. We chose this product as our main source because it underwent extensive validation and demonstrated the highest accuracy among currently available 30-m global cropland datasets (overall accuracy > 97%)68. It also showed strong agreement with FAO statistics (R² > 0.94 from sample-based assessments) and reliably captured cropland changes, with 71% of change events correctly identified68.
We spatially aggregated the 30-m resolution cropland data into 1-km cropland area fractional grid cells by using the mean calculator in ArcGIS Pro v3.0. Then, we combined the cropland data with the shapefile data of Hotspots to extract the cropland data located within Hotspots. The absolute change and relative change methods were used to explore cropland dynamics within each Hotspots. Because hexagonal grids optimize the representation of spatial heterogeneity80, minimize statistical bias80, and enhance the robustness of ecological inferences80, we adopted hexagonal grids of 5000 km² to balance the capture of regional-scale land-use dynamics with the reduction of noise from local fluctuations, and to calculate the average rate of cropland change within each grid from 2000 to 2019.
Optimized HSA can be used to find where spatial clustering of high- or low-value elements occurs81. The calculation formula is shown in Eq. (1). KDE can be used to assess the density of spatial data distributions. The calculation formula is shown in Eq. (2).
Here, xi is the attribute value of the element i, and wij is the spatial weight between the element i and the element j.
Here: i represents the event point; popi represents the value of the population field of the time point; disti is the distance from the valuation point to the event point; radius represents the radius of the search.
We used HSA and KDE in ArcGIS Pro v3.0 to identify spatial distribution density and statistically significant spatial hot spots and cold spots of cropland changes from 2000 to 2019.
Assessing threats of cropland expansion on biodiversity analyses
We used BiodiversityMapping17 (https://biodiversitymapping.org/index.php/download) to analyze threats of cropland expansion on biodiversity. BiodiversityMapping17 was calculated through the spatial overlap of range maps for birds, mammals, and amphibians (the three common vertebrate taxa) with an equal-area 10 km grid cell. This dataset also identified the spatial ranges of threatened amphibians, mammals, and birds, and small-ranged amphibians, mammals, and birds. Here, the vulnerable, endangered, or critically endangered species in the IUCN Red List are considered threatened species; small-ranged species refer to those living in a geographic range that is smaller than the median range size for that taxon.
First, we calculated species richness within each hexagonal grid. Then, we used the Local Indicator of Spatial Association (LISA) in GeoDA 1.22 to analyze hot spots where biodiversity could be most affected by cropland changes.
The LISA results have four types: High–High (H–H), High–Low (H–L), Low–High (L–H), and Low–Low (L–L). H–H clusters indicate areas with high cropland expansion were significantly associated with high species richness (at 0.05 significance level). H–L clusters indicate areas with high cropland expansion that were significantly associated with low species richness (at 0.05 significance level). L–H clusters indicate areas with low cropland expansion (below-average but present) were significantly associated with high species richness (at 0.05 significance level). L–L clusters indicate areas with low cropland expansion were significantly associated with low species richness (at 0.05 significance level).
Because H–H and L–H clusters represent regions experiencing cropland expansion while harboring exceptionally high species richness, they face potential threats from agricultural expansion, so we considered them as risk spots.
Assessing the conservation gaps and conservation priorities within Hotspots
The data for PAs within Hotspots were obtained from the January 2024 edition of the World Database on Protected Area (WDPA, www.protectedplanet.net). We did not consider PAs that only have point data, as we are interested in precisely determining where land is protected within species ranges. So, we only used the polygon boundary data layer, and the point data layer was excluded from our analysis. The IUCN classifies PAs such as Ia (strict nature reserve), Ib (wilderness area), II (national park), III (natural monument or feature), IV (habitat or species management area), V (protected landscape or seascape), or VI (PA with sustainable use of natural resources). PAs without an IUCN category are referred to as No Category.
We performed filtering to eliminate potential duplicates and resolve overlapping issues with PAs. We also addressed international designations that are typically excluded due to duplication with national designations. Since China has not updated its PA boundary data in the WDPA, we used PA data from the April 2018 WDPA82 for China.
We calculated the percentage of PA area within Hotspots and determined the conservation gap. The conservation gaps were defined as the areas of net risk spots of small-ranged vertebrates (i.e., risk spots are not converged by PAs).
To prioritize lands for future conservation, our focus was on the small-ranged vertebrates, including mammals, birds, and amphibians (detailed information in Table S2). For each species, we calculated a priority score by dividing the proportion of the species’ range that is unprotected (i.e., not in PAs) by the area of the species’ range (i.e., not in PAs)83. Conversely, if a large proportion of the species’ range is within PAs, the score decreases accordingly83. Priority maps sum scores across all small-ranged species within each taxonomic group and across all taxonomic groups83.
Data availability
All underlying raw model data are publicly available online. Potapov et al’s cropland data are available at https://glad.umd.edu/dataset/croplands; the species richness data can be obtained from https://biodiversitymapping.org/index.php/download; the PREDICTS database can be obtained from https://data.nhm.ac.uk/dataset/the-2016-release-of-the-predicts-database-v1-1 and https://data.nhm.ac.uk/dataset/release-of-data-added-to-the-predicts-database-november-2022; Biodiversity Hotspots’ shapefile is available at https://zenodo.org/records/3261807. Other ancillary datasets are available at https://zenodo.org/records/17790423.
Code availability
All scripts for the data analyses and visualization are available upon request by contacting the corresponding authors.
References
Cardinale, B. J. et al. Biodiversity loss and its impact on humanity. Nature 486, 59–67 (2012).
Newbold, T. et al. Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50 (2015).
Johnson, C. N. et al. Biodiversity losses and conservation responses in the Anthropocene. Science 356, 270–275 (2017).
Pereira, H. M. et al. Global trends and scenarios for terrestrial biodiversity and ecosystem services from 1900 to 2050. Science 384, 458–465 (2024).
Steffen, W. et al. Planetary boundaries: Guiding human development on a changing planet. Science 347, 1259855 (2015).
Newbold, T. et al. Has land use pushed terrestrial biodiversity beyond the planetary boundary? A global assessment. Science 353, 288–291 (2016).
Tilman, D. et al. Future threats to biodiversity and pathways to their prevention. Nature 546, 73–81 (2017).
Nielsen, K. S. et al. Biodiversity conservation as a promising frontier for behavioural science. Nat. Hum. Behav. 5, 550–556 (2021).
Blicharska, M. et al. Biodiversity’s contributions to sustainable development. Nat. Sustain. 2, 1083–1093 (2019).
Dobson, A. P. et al. Ecology and economics for pandemic prevention. Science https://doi.org/10.1126/science.abc3189 (2020).
Nicholson, E. et al. Scientific foundations for an ecosystem goal, milestones and indicators for the post-2020 global biodiversity framework. Nat. Ecol. Evol. 5, 1338–1349 (2021).
Waldron, A. et al. Reductions in global biodiversity loss predicted from conservation spending. Nature 551, 364–367 (2017).
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000).
Watson, J. E. M. et al. Protect the last of the wild. Nature 563, 27–30 (2018).
Luby, I. H., Miller, S. J. & Polasky, S. When and where to protect forests. Nature 609, 89–93 (2022).
Jenkins, C. N., Pimm, S. L. & Joppa, L. N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl Acad. Sci. USA 110, E2602–E2610 (2013).
Pimm, S. L. et al. The biodiversity of species and their rates of extinction, distribution, and protection. Science 344, 1246752 (2014).
Pimm, S. L. & Raven, P. Extinction by numbers. Nature 403, 843–845 (2000).
Pimm, S. L. & Raven, P. H. Norman Myers (1934–2019. Nat. Ecol. Evol. 4, 177–178 (2020).
Brooks, T. M. et al. Global biodiversity conservation priorities. Science 313, 58–61 (2006).
Brancalion, P. H. S. et al. Global restoration opportunities in tropical rainforest landscapes. Sci. Adv. 5, eaav3223 (2019).
Joppa, L. N., Roberts, D. L., Myers, N. & Pimm, S. L. Biodiversity hotspots house most undiscovered plant species. Proc. Natl Acad. Sci. USA 108, 13171–13176 (2011).
Scheffers, B. R., Joppa, L. N., Pimm, S. L. & Laurance, W. F. What we know and don’t know about Earth’s missing biodiversity. Trends Ecol. Evol. 27, 501–510 (2012).
Lark, T. J., Spawn, S. A., Bougie, M. & Gibbs, H. K. Cropland expansion in the United States produces marginal yields at high costs to wildlife. Nat. Commun. 11, 4295 (2020).
Bateman, I. & Balmford, A. Current conservation policies risk accelerating biodiversity loss. Nature 618, 671–674 (2023).
Outhwaite, C. L., McCann, P. & Newbold, T. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 605, 97–102 (2022).
Usubiaga-Liaño, A., Mace, G. M. & Ekins, P. Limits to agricultural land for retaining acceptable levels of local biodiversity. Nat. Sustain. 2, 491–498 (2019).
Tilman, D. et al. Forecasting agriculturally driven global environmental change. Science 292, 281–284 (2001).
Crist, E., Mora, C. & Engelman, R. The interaction of human population, food production, and biodiversity protection. Science 356, 260–264 (2017).
Potapov, P. et al. Global maps of cropland extent and change show accelerated cropland expansion in the twenty-first century. Nat. Food 3, 19–28 (2022).
Kehoe, L. et al. Biodiversity at risk under future cropland expansion and intensification. Nat. Ecol. Evol. 1, 1129–1135 (2017).
Zabel, F. et al. Global impacts of future cropland expansion and intensification on agricultural markets and biodiversity. Nat. Commun. 10, 2844 (2019).
Molotoks, A. et al. Global projections of future cropland expansion to 2050 and direct impacts on biodiversity and carbon storage. Glob. Change Biol. 24, 5895–5908 (2018).
Newbold, T., Oppenheimer, P., Etard, A. & Williams, J. J. Tropical and Mediterranean biodiversity is disproportionately sensitive to land-use and climate change. Nat. Ecol. Evol. 4, 1630–1638 (2020).
Watson, J. E. M., Dudley, N., Segan, D. B. & Hockings, M. The performance and potential of protected areas. Nature 515, 67–73 (2014).
Brennan, A. et al. Functional connectivity of the world’s protected areas. Science 376, 1101–1104 (2022).
Brodie, J. F. et al. Landscape-scale benefits of protected areas for tropical biodiversity. Nature 620, 807–812 (2023).
De Marco, P. et al. The value of private properties for the conservation of biodiversity in the Brazilian Cerrado. Science 380, 298–301 (2023).
Allan, J. R. et al. The minimum land area requiring conservation attention to safeguard biodiversity. Science 376, 1094–1101 (2022).
Farhadinia, M. S. et al. Current trends suggest most Asian countries are unlikely to meet future biodiversity targets on protected areas. Commun. Biol. 5, 1221 (2022).
Yang, R. et al. Cost-effective priorities for the expansion of global terrestrial protected areas: Setting post-2020 global and national targets. Sci. Adv. 6, eabc3436 (2020).
Xu, H. et al. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 5, 411–418 (2021).
Murali, G., Gumbs, R., Meiri, S. & Roll, U. Global determinants and conservation of evolutionary and geographic rarity in land vertebrates. Sci. Adv. 7, eabe5582 (2021).
Hudson, L. N. et al. The PREDICTS database: a global database of how local terrestrial biodiversity responds to human impacts. Ecol. Evol. 4, 4701–4735 (2014).
Millard, J. et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 12, 2902 (2021).
Filgueiras, B. K. C., Peres, C. A., Melo, F. P. L., Leal, I. R. & Tabarelli, M. Winner–loser species replacements in human-modified landscapes. Trends Ecol. Evol. 36, 545–555 (2021).
Auliz-Ortiz, D. M. et al. Underlying and proximate drivers of biodiversity changes in Mesoamerican biosphere reserves. Proc. Natl Acad. Sci. USA 121, e2305944121 (2024).
Laurance, W. F., Sayer, J. & Cassman, K. G. Agricultural expansion and its impacts on tropical nature. Trends Ecol. Evol. 29, 107–116 (2014).
Sala, O. E. et al. Global Biodiversity Scenarios for the Year 2100. Science 287, 1770–1774 (2000).
Alroy, J. Effects of habitat disturbance on tropical forest biodiversity. Proc. Natl Acad. Sci. USA 114, 6056–6061 (2017).
Silva Junior, C. H. L. et al. Northeast Brazil’s imperiled Cerrado. Science 372, 139–140 (2021).
He, X. et al. Accelerating global mountain forest loss threatens biodiversity hotspots. One Earth 6, 303–315 (2023).
Searchinger, T. D. et al. High carbon and biodiversity costs from converting Africa’s wet savannahs to cropland. Nat. Clim. Change 5, 481–486 (2015).
Hu, X., Huang, B., Verones, F., Cavalett, O. & Cherubini, F. Overview of recent land-cover changes in biodiversity hotspots. Front. Ecol. Environ. 19, 91–97 (2021).
Kong, X., Zhou, Z. & Jiao, L. Hotspots of land-use change in global biodiversity hotspots. Resour. Conserv. Recycl. 174, 105770 (2021).
Chai, L. et al. Telecoupled impacts of the Russia–Ukraine war on global cropland expansion and biodiversity. Nat. Sustain. https://doi.org/10.1038/s41893-024-01292-z (2024).
Orme, C. D. L. et al. Global hotspots of species richness are not congruent with endemism or threat. Nature 436, 1016–1019 (2005).
Raven, P. H. et al. The distribution of biodiversity richness in the tropics. Sci. Adv. 6, eabc6228 (2020).
Williams, D. R. et al. Proactive conservation to prevent habitat losses to agricultural expansion. Nat. Sustain. 4, 314–322 (2020).
Bruner, A. G., Gullison, R. E., Rice, R. E. & Da Fonseca, G. A. B. Effectiveness of parks in protecting tropical biodiversity. Science 291, 125–128 (2001).
de Lima, R. A. F. et al. The erosion of biodiversity and biomass in the Atlantic Forest biodiversity hotspot. Nat. Commun. 11, 6347 (2020).
Wolf, C., Levi, T., Ripple, W. J., Zárrate-Charry, D. A. & Betts, M. G. A forest loss report card for the world’s protected areas. Nat. Ecol. Evol. 5, 520–529 (2021).
Crooks, K. R. et al. Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc. Natl Acad. Sci. USA 114, 7635–7640 (2017).
Haddad, N. M. et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, e1500052 (2015).
Perez-Quezada, J. F. & Scherson, R. Protect central Chile’s biodiversity. Science 382, 165–165 (2023).
Ralimanana, H. et al. Madagascar’s extraordinary biodiversity: threats and opportunities. Science https://doi.org/10.1126/science.adf1466 (2022).
Chen, X. & Hull, V. Border conservation in Hindu Kush-Himalaya. Science 380, 1330–1332 (2023).
Meng, Z. et al. Post-2020 biodiversity framework challenged by cropland expansion in protected areas. Nat. Sustain. 1–11 https://doi.org/10.1038/s41893-023-01093-w (2023).
Jones, K. R. et al. One-third of global protected land is under intense human pressure. Science 360, 788–791 (2018).
Arneth, A. et al. Making protected areas effective for biodiversity, climate and food. Glob. Change Biol. 29, 3883–3894 (2023).
Appleton, M. R. et al. Protected area personnel and ranger numbers are insufficient to deliver global expectations. Nat. Sustain. 5, 1100–1110 (2022).
Barlow, J. et al. The future of hyperdiverse tropical ecosystems. Nature 559, 517–526 (2018).
Pimm, S. We can have biodiversity and eat too. Nat. Food 3, 310–311 (2022).
Henry, R. C. et al. Global and regional health and food security under strict conservation scenarios. Nat. Sustain. 5, 303–310 (2022).
Albert, J. S. et al. Human impacts outpace natural processes in the Amazon. Science 379, eabo5003 (2023).
Chung, M. G. & Liu, J. International food trade benefits biodiversity and food security in low-income countries. Nat. Food 3, 349–355 (2022).
Chowdhury, S. et al. Protected areas and the future of insect conservation. Trends Ecol. Evol. 38, 85–95 (2023).
Noori, S. et al. Extensive mismatch between protected areas and biodiversity hotspots of Iranian Lepidoptera. Insect Conserv. Divers. 17, 938–952 (2024).
Hudson, L. N. et al. The database of the PREDICTS (Projecting Responses of Ecological Diversity In Changing Terrestrial Systems) project. Ecol. Evol. 7, 145–188 (2017).
Birch, C. P. D., Oom, S. P. & Beecham, J. A. Rectangular and hexagonal grids used for observation, experiment and simulation in ecology. Ecol. Model. 206, 347–359 (2007).
Li, G. et al. Global impacts of future urban expansion on terrestrial vertebrate diversity. Nat. Commun. 13, 1628 (2022).
Mi, C. et al. Global Protected Areas as refuges for amphibians and reptiles under climate change. Nat. Commun. 14, 1389 (2023).
Jenkins, C. N., Van Houtan, K. S., Pimm, S. L. & Sexton, J. O. US protected lands mismatch biodiversity priorities. Proc. Natl Acad. Sci. USA 112, 5081–5086 (2015).
Acknowledgements
This research is supported by the National Key Research and Development Program of China (2022YFF0802400). X. Xiao was supported by the research grants from the U.S. National Science Foundation (1911955, 2200310).
Author information
Authors and Affiliations
Contributions
J.D. and C.Y. conceptualized the study; C.Y. performed research, analyzed data, made the visualizations, and wrote the initial draft; C.Y., J.D., C.J., G.Z., X.Z., Y.L., Z.M., K.M., L.Z., R.G., E.E., and X.X. edited the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Sajad Noori, Jing Gan and the other anonymous reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Edmond Totin, Martina Grecequet, and Mengjie Wang. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang, C., Dong, J., Jenkins, C.N. et al. Identifying conservation priorities in global Biodiversity Hotspots to protect small-ranged vertebrates from agricultural pressure. Commun Earth Environ 7, 78 (2026). https://doi.org/10.1038/s43247-025-03099-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-03099-y
