
Abstract
Circadian rhythms play a preeminent role in our life, organizing our physiology and behavior on a daily basis to resonate with our fluctuating environment. However, recent studies reveal that hundreds of mouse and human genes are expressed with a 12-h pattern. We take a close look at mammalian 12-h rhythms, their potential mechanisms and functions, and evidence linking them to circatidal rhythms, which enable marine animals to adapt to tides.
Similar content being viewed by others


Introduction
Life on Earth is profoundly impacted by various environmental cycles of defined periodicities. Organisms cope with them with the help of biological clocks that closely match the period of the cycle they track1. Circadian (~24 hour [h]) clocks allow organisms to optimize their physiology and behavior with the time-of-day. For example, most of us humans sleep during the night and are active during the day. Our sleep/wake and feeding/fasting cycles, as well as many other rhythmic physiological processes, are coordinated by circadian pacemaker neurons located in the suprachiasmatic nucleus (SCN) of the hypothalamus. These neurons synchronize cell-autonomous circadian clocks present throughout our body, which regulate locally rhythmic gene expression2. At the molecular level, the core circadian molecular clock in animals is a negative transcriptional feedback loop that comprises the heterodimeric transcriptional activator Circadian Locomotor Output Cycles Kaput/Brain and Muscle ARNT-like 1 (CLOCK/BMAL1) and its own repressor complex, which, depending on the species, may contain Period (PER), Timeless (TIM) and/or Cryptochrome (CRY) proteins2,3. This core loop is interlocked with a second transcriptional loop that generates a wave of transcription with an opposite phase2. This second loop contains the transcription factors Vrille (VRI) and PAR-Domain-Protein 1 (PDP1) in fruit flies, and RAR-Related Orphan Receptor (ROR)/REV-ERB proteins in mammals.
Other biological timers can play key roles in adaptation to ever-changing environmental conditions, but they are not as well understood1. Circannual (~1 year) clocks are critical for seasonal adaptation, such as the timing of migration, hibernation, and reproduction. Circalunar clocks (~29.5 days) keep track of the phase of the moon and play a particularly important role for the timing of reproduction in the sea. Finally, circatidal clocks allow marine organisms to anticipate the changes linked with the 12.4-h tidal cycle (water level, food availability, currents, temperature, etc.)4,5. Interestingly, there is growing evidence that ca. 12-h rhythms are not limited to marine animals. They have also been observed in cyanobacteria6, diatoms7 and even in terrestrial animals such as Drosophila8,9, C. elegans10, mice10,11,12,13,14,15,16,17,18, and most recently in humans13,19. It has been proposed that 12-h rhythms (or circasemidian rhythms) in terrestrial animals are related to circatidal rhythms10,11,12,13. After introducing these ca. 12.4-h marine rhythms, this review focus on 12-h rhythms in terrestrial mammals, their potential mechanisms, and their role in physiology and human health.
Circatidal clocks
In 1903, the French biologist Georges Bohn brought back beach sand containing a green acoelomate: the Roscoff’s worm. Bohn had noticed that this simple animal, which contains a symbiotic micro-algea, sinks into the sand before the arrival of tides to avoid dispersion. After placing the collected sand in an elongated glass tube, Bohn observed a green ring moving down the tube in anticipation of tides, and moving back up when the low tide would have occurred at the beach of origin20. Since this seminal report of a circatidal rhythm, the nature of the circatidal clock has been hotly debated. Naylor proposed that circatidal rhythms are driven by a dedicated 12.4 -h oscillator21,22, but because the period of circatidal rhythms is so close to half that of the circadian clock, Enright instead proposed that a single clock would drive either circadian or circatidal rhythms, depending on the environmental cycles an animal is exposed to23. The period would be adjusted from 24 h to 24.8 h in the presence of tides, with two peaks of activity generated every 12.4-h. Finally, Palmer and Williams proposed the existence of a circatidal clock comprised of two coupled antiphase 24.8-h oscillators that would generate 12.4-h rhythms of activity24,25.
In coastal insects and crustaceans, behavioral studies strongly support the existence of distinct circadian and circatidal clocks (Fig. 1): circatidal rhythms of behavior can be modulated as a function of the time-of-day even under constant conditions, while circadian behavior can show circatidal influence (reviewed in ref. 4). Hybrid behaviors are observed in the amphipod Parhyale hawaiensis under naturally occurring diurnal or mixed tidal regimen, which deviate significantly from the most common regular 12.4-h tidal cycle26. Moreover, pioneer studies in the crustacean Eurydice pulchra and the mangrove cricket Apteronemobius asahinai further support the idea that circadian and circatidal clocks are distinct: circatidal rhythms of behavior were unaffected by knocking down two essential circadian genes, per27,28 and Clock29, through the abdominal injection of specific dsRNAs to trigger RNA interference (RNAi). Recently, however, two studies found that another essential circadian clock gene, Bmal1, is required for circatidal behavior. One study relied again on RNAi in E pulchra30, while the other used CRISPR/Cas9 mutagenesis to generate a null allele of Bmal1 in the amphipod P. hawaiensis31. In the latter organism, both circadian and circatidal rhythms were disrupted by loss of Bmal1. This indicates some mechanistic overlap between circadian and circatidal clocks. A recent manuscript actually raises the possibility that the mechanistic overlap is broader than expected from RNAi studies32, which have a significant caveat: the incomplete suppression of gene expression. Indeed, ca. 12-h rhythms of per and cry2 mRNAs were observed in a subset of clock neurons in P. hawaiensis animals entrained to rhythmic vibrations mimicking tides, while most clock neurons showed a circadian pattern of gene expression. The existence of distinct circadian and circatidal neurons would explain how rhythmic behavior can be both under circadian and circatidal control in this organism26.

In P. hawaiensis, interacting circadian and circatidal clocks are entrained by the light/dark and tidal cycles, respectively, to control rhythmic behavior. See main text for details. Created in BioRender. Emery, P. (2025) https://BioRender.com/caeejhf.
That per and cry2 might flexibly adopt 12.4 h or 24 h period of expression in P. hawaiensis is reminiscent of observations made in oysters. In these marine animals, circadian clock genes can show either 24-h or 12.4-h rhythms of expression, depending on whether animals are exposed to tides or only to a LD cycle33. The circatidal clock might thus have considerable mechanistic overlap with the circadian clock. Alternatively, circadian gene expression might be driven by a circatidal clock functioning upstream of the circadian clock. The development of novel models and genetic approaches to study circatidal rhythms will hopefully soon help elucidate the molecular mechanisms underlying circatidal clocks4.
Circadian or circatidal control of ~ 12-h rhythms?
Distinguishing circadian from circatidal rhythms can be challenging, both at the behavioral and molecular level, because their periodicities are so close to be harmonics. Genes under circadian clock control could be expressed with a 12-h period if their promoters or enhancers harbor binding sites for both the CLOCK/BMAL1 heterodimer and the transcription factors involved in the second transcriptional feedback loop, or other pairs of regulators active in antiphase17,34. Accordingly, the overlapping BMAL1/REV-ERBα/REV-ERBβ cistrome (genome-wide binding sites) contains a few rhythmic genes with a 12-h period expression pattern35. Nevertheless, the majority of the genes in this common cistrome presented a circadian pattern of expression, suggesting that such dual 12-h regulation is limited, at least for genes co-regulated by ROR/REV-ERB and CLOCK/BMAL1 transcription factors.
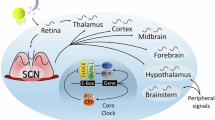
In addition, the circadian clock can generate 12-h rhythms non-cell-autonomously. For example, 12-h rhythms in the mouse liver appear to be dependent on both the local circadian clock and external signaling, presumably from the central circadian pacemaker in the SCN (Fig. 2, see also below)18. At the behavioral level, the circadian clock can generate two bouts of activity per day. For example, Drosophila melanogaster presents two daily activity peaks in the morning and evening, ca. 12 h apart. These peaks are controlled by two different set of circadian pacemaker neurons, referred to as Morning and Evening oscillators36,37. Both harbor the same circadian clock mechanism, but these neurons are either more active in the morning or in the evening38.

Both circadian and circasemidian rhythms of gene expression can be observed in mice (in whole animals or cell lines). They might be generated through three, non-mutually exclusive, mechanisms. Left: the circadian clock generates both 24 h and 12 h rhythms. Circasemidian rhythms could be the result of two distinct sets of transcriptional regulators taking turn to promote gene expression every 12 h (double arrow). Middle: the circadian clock collaborates with systemic factors controlled by feeding to generate circasemidian rhythms. Right: a dedicated 12-h oscillator, independent of the circadian clock, generates circasemidian rhythms, reminiscent of the independent circadian and circatidal clocks in crustaceans (see Fig. 1). Created in BioRender. Emery, P. (2025) https://BioRender.com/n22k1pa.
A key criterion to determine whether a ca. 12-h rhythm in a marine organism is driven by the circadian or the circatidal clock is to test if that rhythm is entrained (synchronized) by the light/dark (LD) cycle or by the tides. In the lab, the phase of the tidal cycle can be shifted, and a circatidal molecular or behavioral rhythm will shift accordingly (see for example31,32). In the field, animals can be collected at different periods of the lunar month (e.g.39). Ideally constant conditions should be used to ensure that the observed rhythms are not direct responses to environmental changes.
Beginning with the mussel Mytilus californianus40, multiple transcriptomics studies have been aimed at identifying genes under circatidal control (e.g.40,41,42,43,44,45,46). To our knowledge however, no such work has so far been designed to meet both key criteria for circatidal rhythms: that the rhythms in the 12-h range free-run and that tides entrain them. This is understandable given the cost and challenges of circatidal transcriptomics studies, particularly in the field, but it is important to keep in mind that at least some of the 12-h oscillations might not actually be circatidal. This issue is particularly acute when 12-h rhythms are designated as, or proposed to be, “circatidal” in animals exposed to LD cycles, in the complete absence of tidal input47,48. To briefly summarize these transcriptomics studies, it appears that hundreds to thousands of genes might be under circatidal control. Genes implicated in transcription, ER function, proteostasis, and metabolism are frequently expressed with a 12-h rhythm4.
The ultimate way to distinguish a circadian rhythm from a circatidal rhythm would be to determine whether a behavioral or molecular rhythm is eliminated when either the circadian or the circatidal clock is genetically disrupted. Combining RNAi or CRISPR/Cas9 gene editing with transcriptomics would be a powerful way to elucidate the mechanism underlying 12-h rhythms in marine organisms. Unfortunately, no gene dedicated to circatidal rhythms has been isolated in any species so far. However, as mentioned above, RNAi studies in crustaceans and insects targeting core circadian genes (per, cry2, and Clock) suggest that circatidal behavior is independent of the circadian clock27,28,29,30. If these observations are confirmed with stringent genome-editing methods, this should open a path to determine whether 12-h transcriptional rhythms persist after disruption of the circadian clock.
12-h rhythms of gene expression in mice
Soon after the initial identification of ca. 12-h rhythms of gene expression in mussels40, similar pattern of expression were unexpectedly discovered in mice. Taking advantage of then-recently developed transcriptomics technologies, Hughes et al. characterized hundreds of 12-h period genes in the liver and other organs17. Under constant darkness condition, the two daily peaks of expression were centered around two critical circadian markers: the beginning of the resting (subjective light) phase and the beginning of the active (subjective dark) phase. Further studies showed that the rhythmic activation of the Unfolded Protein Response (UPR), a signal transduction pathway adjusting critical cellular function to the accumulation of proteins inside the endoplasmic reticulum (ER)49, plays a critical role in the generation of murine circasemidian rhythms of gene expression, suggesting a transcription-led mechanism10,11,14,17. Nevertheless, recent evidence also demonstrates the additional role of mRNA degradation in the generation of 12-h rhythms of gene expression50.
A key question is whether murine 12-h rhythms are driven by the circadian clock or a dedicated circasemidian oscillator. The harmonic nature of the two oscillations again complicates matters. Early investigations indicated that systemic factors, combined with local circadian clocks, determine hepatic circasemidian rhythms (Fig. 2). Like circadian rhythms 12-h rhythms are maintained in constant darkness. However, the phase and amplitude of these rhythms are impacted by obesity and feeding rhythms, themselves controlled by the central circadian pacemaker14,17,18,51. As expected, several studies demonstrated the disappearance of the expression of 12-h rhythmic genes in clock deficient animals or cell cultures14,17,18,50,51,52. However, these 12-h rhythmic genes were expressed with a 24-h rhythm in circadian mutant animals when rhythmic feeding was maintained using time-restricted feeding or by restoring a functional circadian clock in the brain of clock-deficient animals14,17,18,52. Interestingly, only the peak associated with the dark/feeding phase was present.
Thus, based on these studies, it appears that murine 12-h rhythms are the results of a combination of food-related cues and local circadian regulation (Fig. 2). However, recent work by Bokai Zhu and collaborators challenges this conclusion10,11,53. This team has presented evidence for a dedicated cell-autonomous circasemidian oscillator, independent of BMAL1 and thus the circadian clock (Fig. 2). This pacemaker would be organized around XBP1, a key transcription factor in one of the branches of the UPR10,11,53. Accordingly, a large number of 12-h rhythmic genes were associated with the UPR and included genes involved in protein processing in the ER ang Golgi apparatus. In addition, circasemidian rhythms were found in other fundamental cellular processes such as mitochondrial activity, mRNA translation, cell cycle, and interferon/NF-kB pathways10,11,17.
The nature of the murine circasemidian oscillator is currently unclear. Liver-specific elimination of XBP1 significantly decreased the number of 12-h rhythmic transcripts in this organ, but circasemidian rhythms were far from abolished. Some RNAs even saw the amplitude of their 12-h rhythms increasing in the absence of XBP111. Thus, this transcription factor does not appear to be an essential part of the putative Bmal1-independent circasemidian oscillator, but rather an important downstream effector. It will be very important to figure out the root causes for the different conclusions reached on the role of circadian genes such as Bmal1 in the control of murine 12-h rhythms. Is it the use of different cell lines (MEFs10 vs primary hepatocytes, U2OS17, or NIH3T350), animal care, or statistical methods used to identify rhythmic genes? Could both the circadian clock and a distinct circasemidian oscillator generate 12-h rhythms? Clearly, much additional research is required to decipher how circasemidian rhythms are generated in mice.
12-h rhythms in humans
Recent studies indicate that 12-h rhythms in transcript level are also present in humans. In a very original study published in NJP Biological Timing and Sleep, Zhu and collaborators measured gene expression patterns in three volunteers13. Blood samples were collected over 48-h, and the transcriptome of peripheral blood cells was analyzed at high temporal resolution. As expected, thousands of genes showed an expression pattern in the circadian range (5453 genes to be precise), but a significant number showed rhythms in the circasemidian range (653). Importantly, Zhu et al. provide evidence that these genes are not simply controlled by the circadian clock. First, the average period of circadian and circasemidian genes is not a perfect harmonic. Second, the identity and functions of the two pools of genes are clearly different. These results thus support the existence of two different oscillatory mechanisms for ca. 12-h and 24-h rhythms. Interestingly, based on the meta-analysis of the three patients, the circasemidian rhythms appear to involve the UPR transcription factor XBP1, suggesting a conserved mechanism between mouse and human.
However, working with human subject comes with important limitations. The Zhu et al. study was not performed under constant conditions13. The volunteers received meals at specific times of the day and were in control of lightning in their environment. The time at which they switched off or on the lights, when precisely they ate, or when they fell asleep, was not reported. This is significant, particularly considering the impact of feeding rhythm on the activation of the UPR pathway18,51. Indeed, a very striking observation in this study is the remarkably tight phase distribution of rhythmic transcripts in each individual, but also the interindividual variability of phase. It would have been important to know what the volunteers were doing and when to determine whether their 12-h rhythms in gene expression were linked to behavioral, internal or environmental cues. At the very least, such cues might have contributed to the tight phase distribution. They could even be entirely responsible for the observed rhythms. The latter does not seem likely, however. Indeed, the genes that were 12-h rhythmic showed significant overlap with circasemidian transcripts in mouse liver11,17. Importantly, this mouse study was performed under constant conditions, and the distribution of transcript phase was much broader than in the human study.
Are circatidal and terrestrial 12-h rhythms evolutionarily related?
The similarity of 12-h rhythmic genes in humans and mice suggests a conserved underlying mechanism. Zhu et al.13 present evidence for an even deeper evolutionary connection as they found a statistically significant overlap with 12-h rhythms in a cnidarian: A. diaphana47. It would be really fascinating if mammalian 12-h rhythms are evolutionary remnants of marine circatidal rhythms, as the authors proposed. Such connection would be conceivable given the marine ancestry of tetrapod. However, we do not think that such conclusion is warranted yet. First, that genes implicated in splicing, protein synthesis, protein homeostasis, and fatty acid metabolism are regulated by a 12-h oscillator in mammals and Cnidarian could be the result of convergent evolution, given their critical importance for cell metabolism and physiology. Second, the expression studies in A. diaphana were performed under LD conditions, not tidal conditions47. As discussed above, this is a really important caveat, as it is unclear whether the 12-h rhythms observed are driven by a circadian or a circatidal clock, or even simply light-driven. The phase of the rhythmic transcripts in A. diaphana was, as in the human study, very tight, suggesting that acute response to the light cycle at least partially contributed to the observed rhythms. A previous study by Zhu et al. also compared murine circasemidian transcription with a second marine organism, which was exposed to both tides and a LD cycle: the limpet C. rota39. Again, there was significant overlap between the limpet and mouse 12-h rhythms, and the limpet study was based on time series generated at two different time of the lunar month. Thus, the 12-h rhythms observed were synchronized to the tides, not the LD cycle. It would have been interesting to determine whether the conservation of 12-h transcripts between the limpet and mouse extends to humans. There is further indication of an ancient origin for 12-h rhythms. 12-h oscillations in UPR-related genes have been broadly found among living species, including invertebrates9,10,40,54, cnidarian47,48, diatom7, and cyanobacteria6. However, most of these studies were conducted under rhythmic environmental conditions, and such apparent conservation could again be the result of convergent evolution.
In summary, whether there is an evolutionary connection between 12-h rhythms in mammals and circatidal rhythms in marine organisms remains uncertain. Determining the mechanisms of these ultradian oscillations is therefore critical. We know that circatidal behavioral rhythms are dependent on Bmal1 in P. hawaiensis31 and E. pulchra30, but RNAi studies in the latter suggest a separate mechanism for circatidal rhythms independent of per27 or cry230. In mammals, as mentioned above, different studies have come to different conclusions on the necessity of Bmal1 and the circadian clock for 12-h rhythms. While XBP1 seems to be an important contributor, other factors appear also involved, as observed for the UPR. Thus, reconciling contradicting conclusions in mammals and uncovering the mechanism of the marine circatidal clock will help answering the fundamentally important questions of the evolutionary origin of 12-h rhythms.
One can wonder why terrestrial animals would need ca. 12 h rhythms in gene expression since they are not subjected to tides. This might be simply the result of the day or night length averaging 12-h. In mice, the two peaks of gene expression correspond to the beginning of the fasting and feeding periods linked to their rhythmic behavior and physiology. Many animals, including Drosophila as mentioned above, are crepuscular and thus are active with a 12-h period. It is therefore possible that physiological phenomena happening 12 h apart independently activate the UPR and require increased protein synthesis and maturation as Zhu et al. proposed in their “rush hour” hypothesis11.
A recent study identified 12-h rhythms in transcript levels in brain samples from deceased controls and schizophrenic patients19. Curiously, there was a preferential disruption of these 12-h rhythms in the patient cohort, particularly for mRNAs encoding genes of the UPR response and involved in neuronal maintenance. It is unclear whether these rhythmic disruptions are relevant to the etiology of the disease or are a mere consequence of it. Even so, such disruption could contribute to the disease symptoms and thus be clinically relevant. Moreover, as discussed, metabolic conditions also impact 12-h rhythms in mice. It is therefore urgent to shed light on the mechanism underlying these rhythms, which until recently had been overlooked.
Data availability
No datasets were generated or analyzed during the current study.
References
Dunlap, J. C., Loros, J. L. & DeCoursey, P. J. Chronobiology - Biological TImekeeping. (Smaller Associates, Inc., 2004).
Weaver, D. R. & Emery, P. In Fundamental Neuroscience (ed L. R. Squire) 819–845 (Academic Press, 2012).
Kotwica-Rolinska, J. et al. Loss of timeless underlies an evolutionary transition within the circadian clock. Mol. Biol. Evol. 39, msab346. (2022).
Kwiatkowski, E. R., Rosenthal, J. J. C. & Emery, P. Clocks at sea: the genome-editing tide is rising. Trends Genet 40, 387–397 (2024).
Hafker, N. S. et al. Rhythms and Clocks in Marine Organisms. Ann. Rev. Mar. Sci. 15, 509–538 (2023).
Elvitigala, T., Stockel, J., Ghosh, B. K. & Pakrasi, H. B. Effect of continuous light on diurnal rhythms in Cyanothece sp. ATCC 51142. BMC Genomics 10, 226 (2009).
Bilcke, G. et al. Diurnal transcript profiling of the diatom Seminavis robusta reveals adaptations to a benthic lifestyle. Plant J. 107, 315–336 (2021).
Rhoades, S. D., Nayak, K., Zhang, S. L., Sehgal, A. & Weljie, A. M. Circadian- and light-driven metabolic rhythms in Drosophila melanogaster. J. Biol. Rhythms 33, 126–136 (2018).
Zhu, B. & Liu, S. Preservation of approximately 12-h ultradian rhythms of gene expression of mRNA and protein metabolism in the absence of canonical circadian clock. Front Physiol. 14, 1195001 (2023).
Zhu, B. et al. A cell-autonomous mammalian 12 h clock coordinates metabolic and stress rhythms. Cell Metab. 25, 1305–1319 e1309 (2017).
Pan, Y. et al. 12-h clock regulation of genetic information flow by XBP1s. PLoS Biol. 18, e3000580 (2020).
Zhu, B., Dacso, C. C. & O’Malley, B. W. Unveiling “Musica Universalis” of the cell: a brief history of biological 12-hour rhythms. J. Endocr. Soc. 2, 727–752 (2018).
Zhu, B. et al. Evidence for ~12-h ultradian gene programs in humans. NPJ Biol. Timing Sleep. 1, 4 (2024).
Cretenet, G., Le Clech, M. & Gachon, F. Circadian clock-coordinated 12 h period rhythmic activation of the IRE1alpha pathway controls lipid metabolism in mouse liver. Cell Metab. 11, 47–57 (2010).
Mauvoisin, D. et al. Circadian and feeding rhythms orchestrate the diurnal liver acetylome. Cell Rep. 20, 1729–1743 (2017).
Meng, H. et al. XBP1 links the 12-hour clock to NAFLD and regulation of membrane fluidity and lipid homeostasis. Nat. Commun. 11, 6215 (2020).
Hughes, M. E. et al. Harmonics of circadian gene transcription in mammals. PLoS Genet. 5, e1000442 (2009).
Hughes, M. E. et al. Brain-specific rescue of Clock reveals system-driven transcriptional rhythms in peripheral tissue. PLoS Genet. 8, e1002835 (2012).
Scott, M. R. et al. Twelve-hour rhythms in transcript expression within the human dorsolateral prefrontal cortex are altered in schizophrenia. PLoS Biol. 21, e3001688, https://doi.org/10.1371/journal.pbio.3001688 (2023).
Bohn, G. Sur les mouvements oscillatoires des Convoluta roscoffensis. Comptes rendus Hebd. de. l’Acedemie des. Sci. 137, 576–578 (1903).
Reid, D. G. & Naylor, E. Are there separate circatidal and circadian clocks in the shore Crab Carcinus maenas?. Mar. Ecol. Orig. Ser. 52, 1–6 (1989).
Naylor, E. Crab clockwork: the case for interactive circatidal and circadian oscillators controlling rhythmic locomotor activity of Carcinus maenas. Chronobiol. Int. 13, 153–161 (1996).
Enright, J. T. Plasticity in an isopod’s clockworks: shaking shapes form and affects phase and frequency. J. Comp. Physiol. 107, 13–37 (1976).
Palmer, J. D. & Williams, B. G. Comparativre studies of tidal rhythms. II. The dual clock control of the lovomotor rhythms of two decapod crustaceans. Mar. Behav. Physiol. 12, 269–278 (1986).
Palmer, J. D. Dueling hypotheses: circatidal versus circalunidian battle basics–second engagement. Chronobiol. Int 14, 431–433 (1997).
Kwiatkowski, E. R., Rosenthal, J. J. C. & Emery, P. Crosstalk between the circatidal and circadian clocks mediates behavioral adaptation to tidal patterns. Curr. Biol. 34, 5239–5251 e5232 (2024).
Zhang, L. et al. Dissociation of circadian and circatidal timekeeping in the marine crustacean Eurydice pulchra. Curr. Biol. 23, 1863–1873 (2013).
Takekata, H., Matsuura, Y., Goto, S. G., Satoh, A. & Numata, H. RNAi of the circadian clock gene period disrupts the circadian rhythm but not the circatidal rhythm in the mangrove cricket. Biol. Lett. 8, 488–491 (2012).
Takekata, H., Numata, H., Shiga, S. & Goto, S. G. Silencing the circadian clock gene Clock using RNAi reveals dissociation of the circatidal clock from the circadian clock in the mangrove cricket. J. Insect Physiol. 68, 16–22 (2014).
Zhang, L. et al. The circadian clock gene bmal1 is necessary for co-ordinated circatidal rhythms in the marine isopod Eurydice pulchra (Leach). PLoS Genet. 19, e1011011 (2023).
Kwiatkowski, E. R., Schnytzer, Y., Rosenthal, J. J. C. & Emery, P. Behavioral circatidal rhythms require Bmal1 in Parhyale hawaiensis. Curr. Biol. 33, 1867–1882 e1865 (2023).
Oliphant, A., Sia, C. Y., Kyriacou, C. P., Wilcockson, D. C. & Hastings, M. H. Circadian and circatidal oscillations of clock gene expression in brains of Eurydice pulchra and Parhyale hawaiensis. bioRxiv, https://doi.org/10.1101/2025.01.13.632323 (2025).
Tran, D., Perrigault, M., Ciret, P. & Payton, L. Bivalve mollusc circadian clock genes can run at tidal frequency. Proc. Biol. Sci. 287, 20192440 (2020).
Westermark, P. O. & Herzel, H. Mechanism for 12 hr rhythm generation by the circadian clock. Cell Rep. 3, 1228–1238 (2013).
Cho, H. et al. Regulation of circadian behaviour and metabolism by REV-ERB-alpha and REV-ERB-beta. Nature 485, 123–127 (2012).
Grima, B., Chelot, E., Xia, R. & Rouyer, F. Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 431, 869–873 (2004).
Stoleru, D., Peng, Y., Agosto, J. & Rosbash, M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 431, 862–868, https://doi.org/10.1038/nature02926 (2004).
Liang, X., Holy, T. E. & Taghert, P. H. Synchronous Drosophila circadian pacemakers display nonsynchronous Ca(2)(+) rhythms in vivo. Science 351, 976–981 (2016).
Schnytzer, Y. et al. Tidal and diel orchestration of behaviour and gene expression in an intertidal mollusc. Sci. Rep. 8, 4917 (2018).
Gracey, A. Y. et al. Rhythms of gene expression in a fluctuating intertidal environment. Curr. Biol. 18, 1501–1507, https://doi.org/10.1016/j.cub.2008.08.049 (2008).
Mat, A. M. et al. Biological rhythms in the deep-sea hydrothermal mussel Bathymodiolus azoricus. Nat. Commun. 11, 3454, https://doi.org/10.1038/s41467-020-17284-4 (2020).
Connor, K. M. & Gracey, A. Y. Circadian cycles are the dominant transcriptional rhythm in the intertidal mussel Mytilus californianus. Proc. Natl. Acad. Sci. USA 108, 16110–16115 (2011).
Satoh, A. & Terai, Y. Circatidal gene expression in the mangrove cricket Apteronemobius asahinai. Sci. Rep. 9, 3719 (2019).
Simpson, S. D., Ramsdell, J. S., Watson Iii, W. H. & Chabot, C. C. The Draft Genome and Transcriptome of the Atlantic Horseshoe Crab, Limulus polyphemus. Int. J. Genomics 2017, 7636513 (2017).
Payton, L. et al. Widely rhythmic transcriptome in Calanus finmarchicus during the high Arctic summer solstice period. iScience 24, 101927 (2021).
Yokomizo, T. & Takahashi, Y. Plasticity of circadian and circatidal rhythms in activity and transcriptomic dynamics in a freshwater snail. Heredity ((Edinb.)) 132, 267–274 (2024).
Sorek, M. et al. Setting the pace: host rhythmic behaviour and gene expression patterns in the facultatively symbiotic cnidarian Aiptasia are determined largely by Symbiodinium. Microbiome 6, 83, https://doi.org/10.1186/s40168-018-0465-9 (2018).
Rinsky, M. et al. Temporal gene expression patterns in the coral Euphyllia paradivisa reveal the complexity of biological clocks in the cnidarian-algal symbiosis. Sci. Adv. 8, eabo6467 (2022).
Hetz, C., Zhang, K. & Kaufman, R. J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 21, 421–438 (2020).
Unruh, B. A., Weidemann, D. E., Miao, L. & Kojima, S. Coordination of rhythmic RNA synthesis and degradation orchestrates 24- and 12-h RNA expression patterns in mouse fibroblasts. Proc. Natl. Acad. Sci. USA 121, e2314690121 (2024).
Vollmers, C. et al. Time of feeding and the intrinsic circadian clock drive rhythms in hepatic gene expression. Proc. Natl. Acad. Sci. USA 106, 21453–21458 (2009).
Atger, F. et al. Circadian and feeding rhythms differentially affect rhythmic mRNA transcription and translation in mouse liver. Proc. Natl. Acad. Sci. USA 112, E6579–E6588 (2015).
Dion, W. et al. Four-dimensional nuclear speckle phase separation dynamics regulate proteostasis. Sci. Adv. 8, eabl4150 (2022).
O’Neill, J. S. et al. Metabolic molecular markers of the tidal clock in the marine crustacean Eurydice pulchra. Curr. Biol. 25, R326–R327 (2015).
Acknowledgements
Patrick Emery’s work is supported by grants from the National Science Foundation (#2139765) and the National Institute of General Medical Sciences (1R35GM145253). Frédéric Gachon work is supported by the National Institute of Health (R01AG078241) and the Novo Nordisk Foundation (Hallas-Møller Ascending Investigator grant #0087882). Figures were created in Biorender.
Author information
Authors and Affiliations
Contributions
P.E. and F.G. wrote, edited, and revised the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Emery, P., Gachon, F. Biological rhythms: Living your life, one half-day at a time. npj Biol Timing Sleep 2, 21 (2025). https://doi.org/10.1038/s44323-025-00037-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44323-025-00037-1
