
Abstract
Objectives:
Excessive weight gain and obesity are currently among the world’s major threats to health. Women show significantly higher rates of obesity and eating disorders relative to men, but the factors contributing to these gender differences remain uncertain. We examined the correlations between regional brain responses to images of high-calorie versus low-calorie foods and self-reported motivational status, including ratings of general appetite, overeating propensity, state hunger and desire for specific foods.
Subjects:
Thirty-eight healthy right-handed adults (22 male; 16 female) ages 18–45 participated. There were no differences between males and females with regard to age or body mass index (BMI).
Results:
Overall, motivational status correlated significantly with activation within the amygdala, insula and orbitofrontal cortex. Regional activation was then used to predict BMI, an indicator of long-term food consumption and energy expenditure. The combined model was significant, accounting for 76% of the variance in BMI for women, whereas the same regions were not predictive of weight status among men.
Conclusions:
Findings suggest that long-term weight status is related to visual responsiveness to calorie-dense food imagery among women.
Similar content being viewed by others

Introduction
Problems with overweight and obesity are currently among the major public health concerns facing westernized societies. Within the United States, 2 of every 3 adults are classified as overweight,1 and over 1 in 3 now meet criteria for obesity.2 Excess weight gain is associated with numerous long-term health problems, including increased risk of type 2 diabetes, hypertension, stroke, cardiovascular disease, and a range of other negative health outcomes.3 In addition, those meeting criteria for obesity are twice as likely than their normal weight peers to succumb to premature death from a variety of causes.3 Women, in particular, appear to show the greatest problems with excess weight gain,2 extreme obesity,4 and higher rates of eating disorders relative to men.5, 6, 7 Food consumption and weight gain among humans is an extraordinarily complex phenomenon, regulated by genetic,8 central,9 and peripheral neurobiological factors,10 as well as complex social, cognitive and psychological variables.11, 12, 13, 14 Consequently, the basis for these sex differences in weight gain and food consumption remain unclear, but some evidence suggests that there are distinctions between men and women in their behavioral responses to food stimuli,7 and even in the responsiveness of critical brain regions involved in regulating appetite and food intake.15, 16
Individuals vary in their motivational status and behavioral control when confronted with food stimuli.17 Incentive to consume food in the immediate environment is dictated by an individual’s current hunger state18 and general hedonic preference for specific foods.19, 20 While the long-term ability to regulate food consumption and to maintain a stable weight is associated with individual hedonic preferences for food stimuli, it may also be related to a general capacity to inhibit behavior and affective responses.21 In particular, the ability to modulate behavior in response to tempting food is strongly linked to several aspects of impulse control, such as executive attention, inhibitory control and affect regulation,22 capacities that are often associated with the behavioral23 and emotion24 regulation functions of the prefrontal cortex. The medial orbitofrontal cortex, in particular, appears to be one among several nodes within a complex neurocircuitry involved in responding to food stimuli25, 26, 27, 28, 29 and regulating food intake,30, 31 a system that also likely includes the amygdala and posterior insula, among others. In the present study, we used functional magnetic resonance imaging (fMRI) to examine the responsiveness of this system to visual images of unhealthy high-calorie versus relatively healthy low-calorie foods, and correlated that activation with several self-report variables important to food motivation, including general appetite level (Appetite), the propensity to overeat (Overeating), state hunger (Hunger), and hedonic attraction to the individual foods (Food Desire). Based on prior research described above, we restricted our primary analyses to the amygdala, insula and medial orbitofrontal cortex. Activation within the regions found to be related to each of these food motivational indices was then used to predict body mass index (BMI), a stable measure of long-term food consumption. Based on our prior findings that cerebral responses to images of food stimuli were stronger in women than men,16 and similar findings reported by others,15 we hypothesized that such responses would be related to BMI among women, but would be weaker or non-existent among men.
Materials and Methods
Participants
The participants included thirty-eight healthy right-handed adults (22 male; 16 female) recruited via internet advertisements and posted flyers from the Boston metropolitan region, ranging in age from 18 to 45 years (M=30.1, s.d.=8.3). There were no differences between males (M=31.5, s.d.=9.3) and females (M=28.3, s.d.=7.5) with regard to age. A trained research technician screened all potential volunteers during a semi-structured telephone interview. Based on this screening, enrolled participants were deemed to be free from any history of severe medical conditions, head injury, loss of consciousness >30 min, brain tumors, seizures, neurologic conditions, symptoms consistent with Axis I psychopathology, or drug or alcohol treatment. Additionally, potential participants were excluded for current or recent use of any psychoactive medications, illicit substances or excessive alcohol intake. Normal or corrected normal visual acuity with contact lenses was required. BMI was determined through self-reported height and weight recorded on the prescan questionnaire. Men and women ranged from low normal BMI to Stage I obesity (males=24.24, s.d.=3.60, range=19.80–33.47; females=25.08, s.d.=4.01; range=19.84–34.78), which did not differ significantly between the groups. Written informed consent was obtained before enrollment and all participants were compensated for their time. This research study was reviewed and approved by the McLean Hospital Institutional Review Board.
Materials and procedure
Informed consent and prescan procedures began for each participant between 0900 and 1100 hours, during which participants completed a number of self-report inventories querying about demographic information, dietary intake and appetite/food consumption behavior. For the present analysis, participants answered the following questions: (1) ‘what is your appetite like?’ on a 10-point scale (Appetite: 1=never hungry; 10=always hungry), and (2) ‘do you feel you eat more than you intend to’ on a 10-point scale (Overeating: 1=never; 10=always). In order to ensure some variability in hunger ratings, participants were permitted to consume food if desired throughout the prescan period, although all intake throughout the day was documented on a food diary. However, no food was permitted for an hour before the fMRI scans. Men and women did not differ on any of these scales (all P-values >0.05).
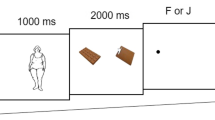
Functional neuroimaging occurred between 1230 and 1500 hours. During fMRI, participants completed a food perception task, similar to the task we have reported in previous papers.16, 25, 26, 27, 29, 32, 33 Briefly, the food perception task consisted of a series of visual images of various food and non-food items. Images were presented in 30-second blocks that alternated between images of high-calorie (H) foods (for example, ice cream, cheeseburgers, cake, French fries, candy), low-calorie (L) foods (for example, fruits, vegetables, fresh salads, whole-grain bread and fresh fish) or control (C) images (that is, non-edible flowers, rocks and shrubs). Each block consisted of 10 images, each displayed for 3 s. A fixation cross (+) was displayed for 15 s at the beginning and end of the task to allow stabilization of the signal. The food perception task followed a constant presentation order (+, C, L, H, C, H, L, C, +) and lasted for a total duration of 240 s. Participants were asked to try to attend to the images in order to identify them in a later recognition test. After the scan, participants indicated their current level of hunger (that is, Hunger: 1=not at all hungry; 7=extremely hungry). Finally, participants were again shown all of the previously seen images and asked to rate ‘how much you would like to eat each item right now’ (that is, Food Desire: 1=do not want to eat it; 7=strongly desire to eat it).
Magnetic resonance imaging parameters
Neuroimaging was completed on a 3.0 Tesla SIEMENS Tim Trio scanner (Erlangen, Germany) using a 12-channel head coil. A T1-weighted 3D MPRAGE sequence (repetition time/echo time/flip angle=2.1 s/2.25 ms/12°) was collected over 128 sagittal slices (256 × 256 matrix) with a slice thickness of 1.33 mm (voxel size=1 × 1 × 1.33 mm). For the 4-minute fMRI during the food perception task, a T2*-weighted echo-planar imaging sequence (repetition time/echo time/flip angle=3.0 s/30 ms/90°) was collected over 43 transverse interleaved slices with 80 images per slice (3.5 mm thickness, no skip; 22.4 cm field of view; 64 × 64 acquisition matrix), with a voxel size of 3.5 × 3.5 × 3.5 mm. The first 3 functional scans were discarded in order to achieve a steady-state equilibrium before data collection.
Image processing
Functional neuroimaging data were preprocessed and analyzed using SPM8 software (Wellcome Department of Cognitive Neurology, London, UK). Standard realignment and motion correction algorithms were employed to remove the effects of participant movement. The echo-planar images were coregistered to each individual’s own T1 anatomical image, and spatially normalized to the template of the Montreal Neurological Institute. An isotropic Gaussian kernel (full width at half maximum=6 mm) was used to spatially smooth the images, which were then resliced to 2 × 2 × 2 mm. During preliminary statistical modeling, the time series was convolved with the canonical hemodynamic response function and a first-level autoregressive model was used to remove the effects of serial autocorrelation. Low frequency drift in the signal was removed by applying the default 128-second high pass filter.
Statistical analysis
The fMRI data were analyzed using a two-stage process. First, the various conditions (that is, high-calorie foods, low-calorie foods and control images) were each modeled against an implicit baseline and contrasts comparing the various conditions were constructed (for example, high-calorie versus low-calorie conditions). The high greater than low calorie contrast image for each subject was then used as the dependent variable in a second-level random effects multiple regression analysis. In this analysis, individual responses to the questions about Appetite, Overeating, Hunger and Desire were entered as separate predictor variables. The linear relation between each predictor variable and brain activation was examined separately while holding the effects of the other variables constant. Based on our a priori hypotheses, we restricted the primary analyses to six bilateral search territories (that is, bilateral amygdala, insula and medial orbitofrontal cortex) as defined by the Automated Anatomical Labeling Atlas,34 implemented within the Wake Forest University SPM8 Toolbox PickAtlas Utility.35 Activation maps for the regression analyses were initially thresholded at P<0.001, k (extent) ⩾10 contiguous voxels, and then subjected to small volume correction for multiple comparisons within each search territory at P<0.05, corrected for family-wise error. Finally, to determine the role of these activation regions in long-term responses to food, brain activation data were extracted from the entire activated cluster in each SPM analysis and entered simultaneously into a multiple linear regression analysis to predict BMI in SPSS 20. Based on prior evidence of sex differences in brain responses to food, we also evaluated this prediction separately for men and women. The multiple correlation coefficients from the separate regression models for men and women were compared directly using Fisher’s r-to-z transform.
Results
Scale intercorrelations
Scale intercorrelations among the various items are presented in Table 1. General Appetite was only significantly correlated with Food Desire. Overeating was significantly correlated with greater BMI and higher Food Desire ratings. State Hunger at the time of the scan was only related to Food Desire ratings of the images following the scan. Other associations were not significant. Together, these correlations provide preliminary evidence of the convergent and discriminant validity of the scales.
Appetite correlations
The relation between self-reported general Appetite ratings and brain responses to the high- versus low-calorie food perception condition was evaluated using multiple linear regression analysis. After statistically controlling for the influence of the other three variables in the regression (that is, Overeating, Hunger and Food Desire), Appetite was not significantly correlated with greater activation within any of the regions of interest to the high-calorie versus low-calorie food images (see Table 2). However, Appetite was associated with significantly reduced task-related activation of a cluster of voxels within the left amygdala (see Figure 1). Table 3 shows the R2 for the overall model and individual β contributions of each of the predictor variables to the activation of this cluster. Figure 1 and Table 2 also show that Appetite was correlated with reduced activation of a cluster within the left posterior insula.

The figure depicts the results of the primary regression analyses for each of the four food motivation scales on brain responses to the high-calorie > low-calorie food contrast, while holding the other three scales constant. The first three columns show the locus of the primary cluster of activation revealed in the regression, and the right hand column depicts the partial correlation scatterplot between the food motivation variable and cluster signal intensity for the data in the highlighted cluster. The figure shows that general Appetite was negatively correlated with clusters in the (a) left amygdala and (b) left insula. Overeating was associated with greater responsiveness within (c) the right medial orbitofrontal cortex (OFC). Hunger ratings were positively correlated with a cluster in the (d) right amygdala, while Food Desirability ratings were positively correlated with a cluster in the (e) left amygdala.
Overeating correlations
The relationship between self-reported Overeating and task-related brain responses was also evaluated. Holding other variables constant (Appetite, Hunger and Food Desire), self-reported Overeating was associated with increased responsiveness to the high-calorie food condition for an activation cluster located within the right medial orbitofrontal gyrus (see Table 2 and Figure 1). In contrast, there were no negative correlations between Overeating and task-related brain responses within any of the search regions.
Hunger correlations
Hunger ratings taken immediately after the scan were also examined independently in the regression. After controlling for the other variables (that is, Appetite, Overeating and Food Desire), self-rated Hunger was positively correlated with activation within a small cluster of the right amygdala (see Table 2 and Figure 1). There were no activation clusters showing a negative correlation between Hunger and task-related brain activation within the regions of interest.
Food desire correlations
Actual ratings of the food images obtained immediately after the scan were also examined for their independent contribution to brain responses for the high- versus low-calorie foods. With the other variables (that is, Appetite, Overeating and Hunger) statistically controlled, Food Desire was positively correlated with activation within a small cluster within the left amygdala (see Table 2 and Figure 1), but no clusters showed any negative correlation with Food Desire during the task.
Exploratory whole-brain analyses
To aid in generation of future hypotheses, each of the preceding regression analyses were also examined at the whole-brain level (that is, not constrained to the hypothesized regions of interest). However, no regions of activation survived whole brain (family-wise error P<0.05) correction for multiple comparisons within any of the analyses.
Multiple regression to predict BMI
The final goal was to determine whether the combined brain activation clusters identified in the preceding analyses could be used to predict an independently obtained indicator of an individual’s long-term eating behavior; in this case we attempted to predict BMI from these cluster responses. For each of the five activation clusters identified in the previous analyses, the cluster eigenvariate was extracted and entered as a predictor variable in a multiple regression analysis with BMI as the dependent variable. Standard regression diagnostics were undertaken to identify particularly influential observations that may have affected the analyses. No participants scored more than 3 s.d. from the mean BMI score and no cases showed excessive influence on the regression analysis (that is, high leverage values or Cook’s Distance scores). For the sample as a whole, a model including all five activation clusters as predictors did not significantly predict BMI, R2=0.166, P=0.30. However, when the same model was tested separately by sex, we found striking differences in model prediction. Whereas there was no significant relation between combined activation from the extracted regions and BMI, R2=0.138, P=0.76 for males, activation within these same regions was highly predictive of BMI for females, R2=0.756, P=0.007. To directly compare the variance explained by these two models, we used a Fisher’s r-to-z transformation of the two multiple correlation coefficients and compared the resulting difference using the z-distribution. This comparison was significant (z=2.62, P=0.009), suggesting that the combined activation from the five brain regions was significantly more predictive of BMI for males than for females. The results were virtually unchanged when menstrual phase was statistically controlled as a nuisance covariate. These findings suggest that long-term weight status among females is closely related to the responsiveness of these brain regions to images of calorie-rich foods. Figure 2 presents the partial correlation plots showing the association between BMI and the standardized predicted scores from the combined activation clusters for males and females.

The figure depicts the partial correlation plots from the secondary regression analyses. In this analysis, the combined brain responses extracted from the primary analyses were used to predict BMI scores separately for men and women. Whereas the extracted functional clusters were not related to BMI among men, they were highly significantly related for women (R2=0.76, *P=0.007).
Discussion
We examined the covariation between regional brain responses to food images and several components of food motivation that might contribute to weight gain and obesity, including general appetite ratings, overeating propensity, current hunger status and ratings of food desirability. These motivational variables were each related to activation within several regions hypothesized a priori to be central in regulating food intake, including the amygdala, posterior insula and medial orbitofrontal cortex. These regions were selected based on prior evidence of their role in processing of visual images of food25, 26, 27, 28, 29 and regulating food intake,30, 31 but likely reflect only a subset of potential brain regions that may be involved. Overall, within this limited set of regions of interest, we found that those who reported greater general appetite tended to show reduced activation within the left amygdala and posterior left insula to images of high- relative to low-calorie foods, while those reporting a tendency to overeat showed greater responsiveness within the medial orbitofrontal cortex to such images. Greater self-rated hunger at the time of the scan was associated with increased responsiveness of voxels within the right amygdala to the high-calorie images, while greater desire to eat the foods depicted was associated with increased activation within the left amygdala. These regions together appear to be reliably responsive to food imagery and correlate significantly with several behaviorally relevant dimensions of eating behavior that may contribute to unhealthy weight gain. Moreover, when the activation within these regions was combined to predict a global measure of long-term food consumption (that is, BMI), there were clear distinctions between men and women in the relation between these brain responses and weight status. Whereas brain responses within these specific food-responsive regions were essentially unrelated to BMI for males, combined activation in these same regions accounted for 76% of the variance in body mass among females. These findings suggest that motivational processing of food images within the brain regions studied here may be reliably related to weight status among women, but may be less so among men.
Several important findings emerge from this study. First, we confirm that specific aspects of food motivation are related to the responsiveness of a core set of brain regions that have been implicated in prior studies of visual food imagery.36 General appetite, which reflects an individual’s self-reported persistent desire for food across settings, was inversely correlated with left amygdala and insular responses to images of high-calorie foods, such that greater responses within these regions were associated with lower appetite ratings. The amygdala has long been implicated in studies of appetitive behavior and food motivation,37 and lesions to the amygdala often result in severe changes in food seeking and consumption.38, 39 In a prior study, appetite ratings were suppressed following a 6-week regimen of daily citicoline administration, and the magnitude of appetite decline was inversely correlated with amygdala and insular responses to images of high-calorie foods,29 suggesting that these structures may have a role in appetite for food. The present findings are also consistent with the hypothesized role of the amygdala in detecting and responding to potential threat or harmful stimuli in the environment,40, 41 and the role of the insula in internally generated sensations of disgust.42, 43 The insula is believed to be part of the extended gustatory cortex and a key region involved in visceral sensation and interoceptive awareness.44 Activation of this region occurs in response to satiety,45 perception of painful and disgusting stimuli,46 and with greater sensitivity to the visceral somatic sensations associated with anxiety.47 This may be important for general appetite, as individuals with greater disgust sensitivity tend to be more restrained in their eating.48 When considered in light of existing research on these brain regions, we speculate that the present finding suggest that individuals with a lower general appetite might have a broad propensity to perceive calorie-rich foods as less appealing, more aversive or even potentially threatening, leading to increased amygdala and insular responses to such stimuli. Of course, the causal direction of this association cannot be inferred from these cross-sectional data, so it remains to be determined whether reduced appetite leads to increased amygdala and insula responses to food, or whether the activation of these regions contributes causally to a decreased desire for food.
Regardless of general appetite, some individuals are particularly prone to eat more than they intend when snacking or consuming a meal. In response to images of calorie-rich foods, self-reported overeating was uniquely associated with increased activation within the medial orbitofrontal cortex, a region that is consistently implicated in reward processes and food preferences.49, 50 For example, in one compelling study, participants underwent positron emission tomography scanning while eating pieces of chocolate to the point of satiety.51 Early in the scan, when the chocolate was still perceived as highly pleasurable, elevated brain activity was found within the caudal regions of the medial orbitofrontal cortex, proximal to the region activated here, but as participants continued to consume additional pieces of chocolate to the point of repulsion, this activation diminished and was replaced by activation within the lateral prefrontal cortex.51 A number of studies have now suggested that the medial orbitofrontal cortex directly tracks the subjective pleasantness of stimuli,52 and this region may contribute directly to decision-making processes that involve pleasure and reward.53 Higher scores on a food addiction scale also correlated with greater activation within the medial orbitofrontal cortex when anticipating the receipt of highly palatable food,54 and this region is also more responsive to food images following a fasting relative to a satiated state.55 In fact, some studies have pointed to an association between altered functioning32, 56 or structure57 of the medial orbitofrontal cortex and general weight status. Greater responsiveness of the medial orbitofrontal cortex to high-calorie food cues may reflect a hyper-sensitivity to the reward value of such foods,54, 58 and might even serve as a risk factor for obesity. Future research may explore whether the responsiveness of this region to rewarding food stimuli may be predictive of long-term weight gain.
We also examined acute hunger at the time of the scan and found that it correlated positively with activation within the right amygdala during perception of the high- versus low-calorie food images, after controlling for other motivational variables. The present findings corroborate prior work showing that acute hunger is a powerful modulator of amygdala responses to images of food.18, 20, 30, 55 For instance, significantly greater right amygdala activation was found in response to food images during a hungry state (that is, 14 h fasting) compared with satiation (that is, an hour after ingesting pizza ad libitum).59 A recent meta-analysis supported the modulating effect of hunger on right amygdala responses to food pictures.36 These findings are also consistent with other research suggesting that the amygdala has an important role in determining the motivational salience of a stimulus,60 and suggest that this salience detection system may be influenced by the motivational status of the individual. Similarly, after controlling for global appetite, overeating and hunger, we also found that the strength of desire to eat the foods depicted in the images was associated with greater response magnitude within the left amygdala to the high- versus low-calorie foods. It is particularly interesting to note that a cluster within the left amygdala was positively correlated with actual ratings of food desirability while a nearby cluster of activation was negatively correlated with general appetite, as described earlier. Although the resolution of our data precludes precise localization within the amygdala, we did find that the cluster associated with ratings of greater food desirability was located slightly more posteriorly than that associated with lower general appetite. Both clusters were collocated within an area corresponding to the superficial and centromedial nucleus groups,61 which project extensively to the orbitofrontal cortex62 and are broadly implicated in generating autonomic, behavioral and emotional signals based on prior learning.63
The primary goal of the present study was to determine the degree to which the identified food-responsive brain activation patterns might relate to long-term weight status among men and women. We found that motivation-related responses to the calorie-rich food images were highly predictive of BMI for women, accounting for up to 76% of variance in weight status, but these same regional brain responses were essentially unrelated to body mass among men. These findings raise the possibility that different factors may contribute to body weight composition for men and women. For women, body mass appears to be significantly related to specific cortico-limbic responses when confronted with visual food cues, particularly images of foods high in calorie density. On the other hand, such an association between brain responses to visual food cues and BMI was essentially absent for men, suggesting that body mass among men is likely to be more affected by any of a number of other factors that were not examined in the current study. The present findings build upon prior work showing that women tend to show greater cortico-limbic responses to visual images of high-calorie food compared with men,16, 64 and further suggest that long-term body weight status in women may be associated with greater responsiveness of the food motivation network to visual images depicting highly palatable food. Although further research will be necessary to determine the extent to which these findings may relate to actual food consumption and weight gain, these preliminary findings may have important implications regarding the higher rates of obesity2, 4 and eating disorders5, 6, 7 among women. In light of these findings, future interventions may benefit by focusing on developing methods to circumvent the neurobehavioral links between visual responses to food and eventual food consumption. Even simple awareness of the possibility that women may be particularly responsive to visual cues of food stimuli may serve as a potential method for curtailing food intake by overtly restricting exposure to such cues.
Several limitations should be borne in mind when interpreting these findings. First, we only explored self-reported food motivation, including self-ratings of general appetite, overeating, hunger and food preferences. As self-reported motivation may differ from actual behavior, it will be important to corroborate these findings using experimental techniques that involve measuring objective eating behavior. Second, participants were screened to exclude psychopathology, and no attempts were made to recruit based on weight status, so the findings may have limited generalizability to patients with eating disorders or those at the extremes of the weight continuum. Third, with the exception of preventing food intake for an hour prior to the scan, we did not directly manipulate hunger status or total calorie intake. This permitted us to measure brain responses across a normal spectrum of hunger and satiety, but may have also introduced error variance that potentially reduced our power to detect some statistical effects. Future research may benefit from direct manipulation of hunger status by holding calorie intake constant for a longer interval before the scan. Fourth, although we found significant differences between men and women in the relationship between regional brain responses and BMI, we cannot exclude the possibility that the findings were driven by other factors that were not examined or controlled in the current study. It is conceivable that body mass among men may be better accounted for by some other combination of elementary biological or physiological factors such as serum testosterone, age-related somatic changes, activity level or even to social or gender-role variables that influence the circumstances surrounding food and beverage consumption in western cultures. It is also possible that the present sex difference emerged because of the greater variation in lean muscle mass as a component of BMI among men versus women65 or even that men were simply less reliable at reporting height and weight than women. Future work should examine other more direct indices of fat to lean muscle mass to verify the currently observed sex differences. Finally, our primary hypotheses only focused on a small number of discrete brain regions. Although whole-brain analyses failed to show additional regions of correlation following stringent corrections for multiple comparisons, it is likely that at less stringent thresholds, other critically important regions may also emerge as significant. Thus, we make no claims that the regions observed here are the only ones that may be important in this process. Future work examining other regions important for food processing may also enhance our understanding of the neural underpinnings leading to weight gain and obesity. Despite these limitations, the present findings provide further support for a key network of regions involved in food motivation and further suggest that the responses within this network during visual perception of high-calorie foods are directly and strongly related to long-term weight status among women. These findings raise the speculative possibility that the vulnerabilities to weight gain, obesity and eating disorders, which predominate among women may be influenced to some extent by a greater neurocognitive responsiveness to the visual cues associated with food images.
References
Flegal KM, Carroll MD, Ogden CL, Johnson CL . Prevalence and trends in obesity among US adults, 1999-2000. JAMA 2002; 288: 1723–1727.
Ogden CL, Carroll MD, Kit BK, Flegal KM . Prevalence of obesity in the United States, 2009-2010. NCHS Data Brief 2012; 82: 1–8.
Overweight, obesity, and health risk. National task force on the prevention and treatment of obesity. Arch Intern Med 2000; 160: 898–904.
Ogden CL, Carroll MD, Curtin LR, McDowell MA, Tabak CJ, Flegal KM . Prevalence of overweight and obesity in the United States, 1999-2004. JAMA 2006; 295: 1549–1555.
Lewinsohn PM, Seeley JR, Moerk KC, Striegel-Moore RH . Gender differences in eating disorder symptoms in young adults. Int J Eat Disord 2002; 32: 426–440.
Striegel-Moore RH, Bulik CM . Risk factors for eating disorders. Am Psychol 2007; 62: 181–198.
Striegel-Moore RH, Rosselli F, Perrin N, DeBar L, Wilson GT, May A et al. Gender difference in the prevalence of eating disorder symptoms. Int J Eat Disord 2009; 42: 471–474.
Yeo GS, Heisler LK . Unraveling the brain regulation of appetite: lessons from genetics. Nat Neurosci 2012; 15: 1343–1349.
Zeltser LM, Seeley RJ, Tschop MH . Synaptic plasticity in neuronal circuits regulating energy balance. Nat Neurosci 2012; 15: 1336–1342.
Williams KW, Elmquist JK . From neuroanatomy to behavior: central integration of peripheral signals regulating feeding behavior. Nat Neurosci 2012; 15: 1350–1355.
Gregersen NT, Moller BK, Raben A, Kristensen ST, Holm L, Flint A et al. Determinants of appetite ratings: the role of age, gender, BMI, physical activity, smoking habits, and diet/weight concern. Food Nutr Res 2011; 55, doi: 10.3402/fnr.v55i0.7028.
Hooper N, Sandoz EK, Ashton J, Clarke A, McHugh L . Comparing thought suppression and acceptance as coping techniques for food cravings. Eat Behav 2012; 13: 62–64.
Moore CJ, Cunningham SA . Social position, psychological stress, and obesity: a systematic review. J Acad Nutr Diet 2012; 112: 518–526.
Verstuyf J, Patrick H, Vansteenkiste M, Teixeira PJ . Motivational dynamics of eating regulation: a self-determination theory perspective. Int J Behav Nutr Phys Act 2012; 9: 21.
Cornier MA, Salzberg AK, Endly DC, Bessesen DH, Tregellas JR . Sex-based differences in the behavioral and neuronal responses to food. Physiol Behav 2010; 99: 538–543.
Killgore WDS, Yurgelun-Todd DA . Sex differences in cerebral responses to images of high versus low-calorie food. Neuroreport 2010; 21: 354–358.
Lawrence NS, Hinton EC, Parkinson JA, Lawrence AD . Nucleus accumbens response to food cues predicts subsequent snack consumption in women and increased body mass index in those with reduced self-control. Neuroimage 2012; 63: 415–422.
LaBar KS, Gitelman DR, Parrish TB, Kim YH, Nobre AC, Mesulam MM . Hunger selectively modulates corticolimbic activation to food stimuli in humans. Behav Neurosci 2001; 115: 493–500.
Lutter M, Nestler EJ . Homeostatic and hedonic signals interact in the regulation of food intake. J Nutr 2009; 139: 629–632.
Piech RM, Lewis J, Parkinson CH, Owen AM, Roberts AC, Downing PE et al. Neural correlates of appetite and hunger-related evaluative judgments. PLoS One 2009; 4: e6581.
Nederkoorn C, Houben K, Hofmann W, Roefs A, Jansen A . Control yourself or just eat what you like? Weight gain over a year is predicted by an interactive effect of response inhibition and implicit preference for snack foods. Health Psychol 2010; 29: 389–393.
Hofmann W, Friese M, Roefs A . Three ways to resist temptation: the independent contributions of executive attention, inhibitory control, and affect regulation to the impulse control of eating behavior. J Exp Soc Psychol 2009; 45: 431–435.
Batterink L, Yokum S, Stice E . Body mass correlates inversely with inhibitory control in response to food among adolescent girls: an fMRI study. Neuroimage 2010; 52: 1696–1703.
Gruber AJ, McDonald RJ . Context, emotion, and the strategic pursuit of goals: interactions among multiple brain systems controlling motivated behavior. Front Behav Neurosci 2012; 6: 50.
Killgore WDS, Young AD, Femia LA, Bogorodzki P, Rogowska J, Yurgelun-Todd DA . Cortical and limbic activation during viewing of high- versus low-calorie foods. Neuroimage 2003; 19: 1381–1394.
Killgore WDS, Yurgelun-Todd DA . Developmental changes in the functional brain responses of adolescents to images of high and low-calorie foods. Dev Psychobiol 2005; 47: 377–397.
Killgore WDS, Yurgelun-Todd DA . Positive affect modulates activity in the visual cortex to images of high calorie foods. Int J Neurosci 2007; 117: 643–653.
Siep N, Roefs A, Roebroeck A, Havermans R, Bonte ML, Jansen A . Hunger is the best spice: an fMRI study of the effects of attention, hunger and calorie content on food reward processing in the amygdala and orbitofrontal cortex. Behav Brain Res 2009; 198: 149–158.
Killgore WDS, Ross AJ, Kamiya T, Kawada Y, Renshaw PF, Yurgelun-Todd DA . Citicoline affects appetite and cortico-limbic responses to images of high-calorie foods. Int J Eat Disord 2010; 43: 6–13.
Mehta S, Melhorn SJ, Smeraglio A, Tyagi V, Grabowski T, Schwartz MW et al. Regional brain response to visual food cues is a marker of satiety that predicts food choice. Am J Clin Nutr 2012; 96: 989–999.
Murdaugh DL, Cox JE, Cook EW, Weller RE . fMRI reactivity to high-calorie food pictures predicts short- and long-term outcome in a weight-loss program. Neuroimage 2012; 59: 2709–2721.
Killgore WDS, Yurgelun-Todd DA . Body mass predicts orbitofrontal activity during visual presentations of high-calorie foods. Neuroreport 2005; 16: 859–863.
Killgore WDS, Yurgelun-Todd DA . Affect modulates appetite-related brain activity to images of food. Int J Eat Disord 2006; 39: 357–363.
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002; 15: 273–289.
Maldjian JA, Laurienti PJ, Kraft RA, Burdette JH . An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 2003; 19: 1233–1239.
van der Laan LN, de Ridder DT, Viergever MA, Smeets PA . The first taste is always with the eyes: a meta-analysis on the neural correlates of processing visual food cues. Neuroimage 2011; 55: 296–303.
Goddard GV . Functions of the Amygdala. Psychol Bull 1964; 62: 89–109.
Loscher W, Brandt C, Ebert U . Excessive weight gain in rats over extended kindling of the basolateral amygdala. Neuroreport 2003; 14: 1829–1832.
Malkova L, Mishkin M, Suomi SJ, Bachevalier J . Long-term effects of neonatal medial temporal ablations on socioemotional behavior in monkeys (Macaca mulatta). Behav Neurosci 2010; 124: 742–760.
Eippert F, Veit R, Weiskopf N, Erb M, Birbaumer N, Anders S . Regulation of emotional responses elicited by threat-related stimuli. Hum Brain Mapp 2007; 28: 409–423.
Phelps EA, O'Connor KJ, Gatenby JC, Gore JC, Grillon C, Davis M . Activation of the left amygdala to a cognitive representation of fear. Nat Neurosci 2001; 4: 437–441.
Britton JC, Phan KL, Taylor SF, Welsh RC, Berridge KC, Liberzon I . Neural correlates of social and nonsocial emotions: an fMRI study. Neuroimage 2006; 31: 397–409.
Fitzgerald DA, Posse S, Moore GJ, Tancer ME, Nathan PJ, Phan KL . Neural correlates of internally-generated disgust via autobiographical recall: a functional magnetic resonance imaging investigation. Neurosci Lett 2004; 370: 91–96.
Augustine JR . Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res 1996; 22: 229–244.
Morris JS, Dolan RJ . Involvement of human amygdala and orbitofrontal cortex in hunger-enhanced memory for food stimuli. J Neurosci 2001; 21: 5304–5310.
Benuzzi F, Lui F, Duzzi D, Nichelli PF, Porro CA . Does it look painful or disgusting? Ask your parietal and cingulate cortex. J Neurosci 2008; 28: 923–931.
Killgore WDS, Britton JC, Price LM, Gold AL, Deckersbach T, Rauch SL . Neural correlates of anxiety sensitivity during masked presentation of affective faces. Depress Anxiety 2011; 28: 243–249.
Houben K, Havermans RC . A delicious fly in the soup. The relationship between disgust, obesity, and restraint. Appetite 2012; 58: 827–830.
Kringelbach ML, Rolls ET . The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog Neurobiol 2004; 72: 341–372.
Rolls ET . The orbitofrontal cortex and reward. Cereb Cortex 2000; 10: 284–294.
Small DM, Zatorre RJ, Dagher A, Evans AC, Jones-Gotman M . Changes in brain activity related to eating chocolate: from pleasure to aversion. Brain 2001; 124 (Pt 9): 1720–1733.
Grabenhorst F, Rolls ET . Different representations of relative and absolute subjective value in the human brain. Neuroimage 2009; 48: 258–268.
Grabenhorst F, Rolls ET . Value, pleasure and choice in the ventral prefrontal cortex. Trends Cogn Sci 2011; 15: 56–67.
Gearhardt AN, Yokum S, Orr PT, Stice E, Corbin WR, Brownell KD . Neural correlates of food addiction. Arch Gen Psychiat 2011; 68: 808–816.
Holsen LM, Zarcone JR, Thompson TI, Brooks WM, Anderson MF, Ahluwalia JS et al. Neural mechanisms underlying food motivation in children and adolescents. Neuroimage 2005; 27: 669–676.
Volkow ND, Wang GJ, Telang F, Fowler JS, Thanos PK, Logan J et al. Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. Neuroimage 2008; 42: 1537–1543.
Smucny J, Cornier MA, Eichman LC, Thomas EA, Bechtell JL, Tregellas JR . Brain structure predicts risk for obesity. Appetite 2012; 59: 859.
Nolan-Poupart S, Veldhuizen MG, Geha P, Small DM . Midbrain response to milkshake correlates with ad libitum milkshake intake in the absence of hunger. Appetite 2013; 60: 168–174.
Fuhrer D, Zysset S, Stumvoll M . Brain activity in hunger and satiety: an exploratory visually stimulated FMRI study. Obesity (Silver Spring) 2008; 16: 945–950.
Santos A, Mier D, Kirsch P, Meyer-Lindenberg A . Evidence for a general face salience signal in human amygdala. Neuroimage 2011; 54: 3111–3116.
Kim HJ, Kim N, Kim S, Hong S, Park K, Lim S et al. Sex differences in amygdala subregions: evidence from subregional shape analysis. Neuroimage 2012; 60: 2054–2061.
Bach DR, Behrens TE, Garrido L, Weiskopf N, Dolan RJ . Deep and superficial amygdala nuclei projections revealed in vivo by probabilistic tractography. J Neurosci 2011; 31: 618–623.
LeDoux JE . Emotion circuits in the brain. Annu Rev Neurosci 2000; 23: 155–184.
Frank S, Laharnar N, Kullmann S, Veit R, Canova C, Hegner YL et al. Processing of food pictures: influence of hunger, gender and calorie content. Brain Res 2010; 1350: 159–166.
Gallagher D, Visser M, Sepulveda D, Pierson RN, Harris T, Heymsfield SB . How useful is body mass index for comparison of body fatness across age, sex, and ethnic groups? Am J Epidemiol 1996; 143: 228–239.
Acknowledgements
This research was supported by a USAMRAA grant (W81XWH-09-1-0730).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Killgore, W., Weber, M., Schwab, Z. et al. Cortico-limbic responsiveness to high-calorie food images predicts weight status among women. Int J Obes 37, 1435–1442 (2013). https://doi.org/10.1038/ijo.2013.26
Received:
Revised:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/ijo.2013.26
Keywords
This article is cited by
-
Neuroimaging Investigations of Obesity: a Review of the Treatment of Sex from 2010
Current Obesity Reports (2023)
-
Obesity and brain structure in schizophrenia – ENIGMA study in 3021 individuals
Molecular Psychiatry (2022)
-
Sex/Gender Differences in Obesity Prevalence, Comorbidities, and Treatment
Current Obesity Reports (2021)
-
Association between body mass index and subcortical brain volumes in bipolar disorders–ENIGMA study in 2735 individuals
Molecular Psychiatry (2021)
-
Laparoscopic sleeve gastrectomy improves brain connectivity in obese patients
Journal of Neurology (2020)
