
Abstract
Recent genetic studies have found common genomic risk variants among psychiatric disorders, strongly suggesting the overlaps in their molecular and cellular mechanism. Our research group identified the variant in ASTN2 as one of the candidate risk factors across these psychiatric disorders by whole-genome copy number variation analysis. However, the alterations in the human neuronal cells resulting from ASTN2 variants identified in patients remain unknown. To address this, we used patient-derived and genome-edited iPS cells with ASTN2 deletion; cells were further differentiated into neuronal cells. A comprehensive gene expression analysis using genome-edited iPS cells with variants on both alleles revealed that the expression level of ZNF558, a gene specifically expressed in human forebrain neural progenitor cells, was greatly reduced in ASTN2-deleted neuronal cells. Furthermore, the expression of the mitophagy-related gene SPATA18, which is repressed by ZNF558, and mitophagy activity were increased in ASTN2-deleted neuronal cells. These phenotypes were also detected in neuronal cells differentiated from patient-derived iPS cells with heterozygous ASTN2 deletion. Our results suggest that ASTN2 deletion is related to the common pathogenic mechanism of psychiatric disorders by regulating mitophagy via ZNF558.
Similar content being viewed by others


Introduction
Schizophrenia (SCZ), autism spectrum disorder (ASD), and bipolar disorder (BP) are very common psychiatric disorders, with high lifetime prevalence rates of ~0.5% [1, 2], 2.5% [3,4,5], and 1% [6, 7], respectively. They are characterized by their duration as chronic diseases, from childhood to the time of death. This results in a longer treatment period and extensive social and personal damage [8, 9]. However, the biological pathogenesis of these psychiatric disorders is not well understood. Their understanding is urgently needed to develop novel therapies that significantly improve the functional outcomes of patients.
The Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition, has established the diagnostic criteria for SCZ, ASD, and BP as independent and separate diseases. In contrast, in clinical settings, the comorbid diagnoses are also allowed, and in some patients, drawing the diagnose line is difficult, suggesting that these psychiatric disorders partially share clinical symptoms and phenotypes. Indeed, studies reported that psychiatric disorders show common alterations, such as dysfunction in the white matter microstructure in the body of the corpus callosum [10] and brain responses in the ventral striatum during reward anticipation [11]. Additionally, recent genetic studies have uncovered common risk genomic variants in these psychiatric disorders [12,13,14,15]. These findings imply that there are common underlying mechanisms in these psychiatric disorders, prompting us to examine the molecular and cellular alterations caused by the common risk variants across these psychiatric disorders.
Recently, we identified the variants in ASTN2 as a candidate risk factor for psychiatric disorders by whole-genome copy number variation (CNV) analysis [13, 16]. Moreover, a study reported that deletions in ASTN2 that would disrupt almost all transcript isoforms (Fig. 1A, B, shaded gray region) are significantly enriched in patients with neurodevelopmental disorders, such as ASD, attention-deficit/hyperactivity disorder, and intellectual disability [17]. In support of these results, ASTN2 is also listed in multiple databases, such as SFARI (https://gene.sfari.org/) and DBDGD (https://dbd.geisingeradmi.org/), which are centered on genes implicated in these psychiatric disorders.

A Diagram of the deleted region of ASTN2 in patients with psychiatric disorders identified by our previous CNV analysis. Red bars show the deletion region in each patient. B Diagram of ASTN2 splicing variants. Exons in yellow are the most upstream exons of those common to all splicing variants. Regions in gray are those that have been previously identified as significantly associated with neurodevelopmental disorders. C The indel pattern of nonhomologous end joining. The red letters represent PAM sequences. D Immunostaining of pluripotent stem cell markers (NANOG and TRA-1-60) in iPS cell lines. The scale bar represents 500 µm. E RT-qPCR analysis of ASTN2 in each iPS cell line. N = 3. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. *p < 0.05; **p < 0.01.
ASTN2 encodes a large vertebrate-specific transmembrane protein, astrotactin-2, which has been implicated in mouse models to regulate neuronal migration [18] and synaptic function [19]. In addition, ASTN2 knockout mice show monoaminergic dysfunction and abnormal neuronal morphogenesis with shrinkage of the neuronal soma, implying some behavioral abnormalities and some emotional or cognitive impairments [20]. However, the molecular and cellular mechanisms caused by ASTN2 variants identified in patients remain unknown. Moreover, considering the differences in structures and gene expressions between rodent and human brains, the examination of human neuronal cells is also required to understand the molecular and cellular alterations [21,22,23].
To address this, we applied a strategy using patient-derived and genome-edited iPS cells with ASTN2 deletion. iPS cells have the genetic and molecular characteristics of individual subjects, allowing the researchers to overcome the limitations of animal models and to examine disease- and genetic-related phenotypes at the human cellular level [24, 25]. Here, we differentiated each iPS cell into neuronal cells and further examined them at the cellular and molecular levels. Our results suggest that as a pathogenic mechanism for psychiatric disorders, ASTN2 deletion promotes mitophagy by downregulating ZNF558 expression in human neuronal cells.
Material and methods
Subject
Previously, we had established iPS cells from one male Japanese patient with SCZ from eight patients with ASTN2 deletion (SCZ3 in Fig. 1A, chr9:118,490,453–118,538,170, NCBI36/hg18) [26]. The two cloned iPS cell lines from this patient with SCZ were used in this study (PT_1 and PT_2). Two healthy Japanese subjects, a 34-year-old man (CON1) and a 30-year-old woman (CON2), were selected as the sources for control iPS cells, which had no pathogenic CNVs [27]. The given ages of the subjects were those at the time of the blood sampling for iPS cell generation. The use of human samples and genomic analyses were approved by the Ethics Committee of Nagoya University (approval number: 2012-0184). All subjects provided written informed consent.
Establishment of iPS cells and differentiation into neurospheres (NSs)
The iPS cells were established from the peripheral blood of each subject and cultured as previously reported [26]. The cells were differentiated into NSs, as previously reported, with minor modifications [28]. After the passage of iPS cells onto feeder cells (Day 0), the iPS cells were cultured in an iPS cell medium comprising Dulbecco’s Modified Eagle Medium (DMEM/F12) supplemented with 20% knockout serum replacement (KSR), 2 -mM l-glutamine, 0.1-mM nonessential amino acids, 0.1-mM 2-mercaptoethanol, 100-units/mL penicillin, 100-μg/mL streptomycin, and 4-ng/mL basic fibroblast growth factor (bFGF). On day 1, SB431542 (3 μM), CHIR99021 (3 μM), and dorsomorphin (3 μM) were added to the iPS cell medium, and the iPS cells were incubated for 6 days (days 1–7). On day 7, they were dissociated into single cells with 0.5× TrypLE select and cultured in NS medium consisting of DMEM/F12 supplemented with 1× N-2 supplement, 0.6% glucose, 100-units/mL penicillin, 100-μg/mL streptomycin, 5-mM HEPES, 1× B-27 supplement, 20-ng/mL bFGF, 10-ng/mL hLIF, 10-μM Y-27632, 3-μM CHIR99021, and 2-μM SB431542 for 2 weeks (days 7–21). The cells were dissociated into single cells once on day 14 for passage. Then, the cells were cultured in a 5% CO2/18–22% O2 atmosphere during all experiments.
Assessment of the size of NSs
The differentiated NSs (day 21) were imaged in a bright field using a BZ-X810 fluorescence microscope (KEYENCE, Japan). Induction into the NSs was performed twice independently, and 50 NSs were randomly selected for measurement. Each NS was identified as a circle or oval on the image using Adobe Illustrator 26.5 (Adobe Systems Inc, CA, USA), and subsequently, the area of each NS was measured using ImageJ.
Evaluation of neurite outgrowth
On day 21, the differentiated NSs were dissociated into single cells and plated onto poly-l-ornithine/laminin/fibronectin-coated dishes in a pan-neuron medium (DMEM/F12 supplemented with 1× N-2 supplement, 0.6% glucose, 100-units/mL penicillin, 100-μg/mL streptomycin, 5-mM HEPES, 1× B-27 supplement, 10-μM DAPT, 20-ng/mL BDNF, 20-ng/mL GDNF, 0.2-mM ascorbic acid, 1-ng/mL TGF-β3, and 0.5-mM dbcAMP) for 3 days. For the mitophagy induction study, H2O2 (5 μM/20 μM) or rapamycin (0.5 μM/2 μM) was added as a mitophagy inducer [29, 30] and for the mitophagy inhibition study, chloroquine (0.1 μM/0.5 μM) was added as a mitophagy inhibitor to the pan-neuron medium. Timelapse images were acquired by IncuCyte ZOOM™ software (sartorius, MI, USA) for 3 days.
Immunocytochemistry
The cultured cells were fixed with 4% paraformaldehyde for 15 min at room temperature, permeabilized and blocked in phosphate-buffered saline (PBS) containing 0.3% Triton X-100 and 1% bovine serum albumin for 60 min at room temperature, and then incubated with each of the primary antibodies overnight at 4 °C. After washing with PBS, the immunolabeled cells were incubated with appropriate fluorophore-conjugated secondary antibodies for 1 h at room temperature. DAPI (Dojindo, Japan) was used for nuclei. The antibodies used for immunocytochemistry are listed in Supplementary Table 1. Images were captured using a BZ-X810 fluorescence microscope.
Five images in each cell line were taken, and the number of βIII-tubulin positive, vGLUT1-positive, and DAPI-positive were counted. In measuring the neurite length, the 15 longest neurites were selected for each image. Neurite lengths were measured and calculated using ImageJ and its plug-in NeuronJ.
Comprehensive expression analysis
Expression analysis was performed using the SurePrint G3 Human GE 8x60K V2 Microarray Kit (Agilent Technology, CA, USA), according to the manufacturer’s instructions. The sample number for the microarray was n = 3. All data (GEO: GSE260713) analyses were performed using the GeneSpring GX Software Program (version 13; Agilent Technology). Absolute expression values were normalized using quantile normalization. The statistical criteria for differential expression were as follows: moderated t-test, Bonferroni correction for multiple testing correction, corrected p value <0.05, and fold changes >2. Gene Ontology (GO) analysis (using GOTERM_BP_FAT, GOTERM_MF_FAT, and GOTERM_CC_FAT) was performed using DAVID’s functional annotation tool (https://david.ncifcrf.gov/).
Analysis of ZNF558 binding sites
To analyze the ZNF558 binding sites to DNA, we used three sets of chromatin immunoprecipitation sequencing (ChIP-seq) data available on the Sequence Read Archive (SRA) (https://www.ncbi.nlm.nih.gov/sra) (SRR5197178, SRR5111604, SRR5111605). ZNF224 (SRR5197084) was used as a control gene. The data were analyzed based on a previous report [31]. Briefly, adapter sequences were trimmed from raw datasets using Trimmomatic, followed by quality control using Fastq. Then, the data were mapped to the GRCh38 reference sequence using Bowtie2 with the sensitive-local setting. Multi-mapped reads were filtered out. Polymerase chain reaction (PCR) duplicates were then removed using samtools and repeat regions on the genome were removed using bedtools. Peak calling was performed using MACS2. Then, the processed called peaks were analyzed using GREAT, version 4.0.4 [32]. Binding sequence motif analysis was performed using HOMER.
Evaluation of mitophagy
On day 21, NSs were dissociated into single cells and stained with Mtphagy Dye (Dojindo 1000:1) for 30 min in a 5% CO2/18–22% O2 atmosphere. The cells were washed with PBS and incubated in 96-well U-bottom plates at 5.0 × 104 cells per well. Twenty-four hours later, they were stained with DAPI (Dojindo 5000:1) for 15 min, then dissociated again into single cells, and applied into BD FACS Canto II (BD Bioscience). The data were analyzed using FlowJo.
Quantitative reverse transcription PCR (RT-qPCR) and measurement of mitochondrial DNA and VNTR (variable number tandem repeat) copy numbers
Total RNA was extracted using the RNeasy Plus Mini Kit (QIAGEN, Netherlands). Then, cDNA was generated using the High-Capacity RNA-to-cDNA Kit (Applied Biosystems, CA, USA). RT-qPCR was performed using KAPA SYBR Fast qPCR Kit (KAPA BIOSYSTEMS,) and detected using QuantStudio5 (Applied Biosystems). The gene expression values were normalized to the housekeeping gene RPS18. The primers for mitochondrial DNA (mtDNA) and nuclear DNA (nucDNA) copy number measurement were used as previously reported [33]. As previously reported, the VNTR copy numbers were measured [31]. Albumin was used as the positive control, and the values were calculated using the copy number of albumin as 2. The primers used are listed in Supplementary Table 2.
Statistics
Statistical comparisons were conducted using repeated-measure multiple analysis of variance for the timelapse imaging study. For the other experiments, an unpaired t-test (two tails) or two-way analysis of variance followed by Dunnett’s post hoc test was used. The sample size was determined based on pilot data from our laboratory and previous similar studies. Statistical Package for the Social Sciences, version 28.0.1.0 (IBM, USA), was used to analyze the data. The significance level was set at 0.05 or 0.01.
Results
Deleted region of ASTN2 in each patient with psychiatric disorders and establishment of ASTN2-deleted iPS cells by CRISPR/Cas9 genome editing
Our research group recently identified heterozygous ASTN2 deletions in eight patients with psychiatric disorders (three patients with SCZ, one patient with ASD, and four patients with BP) by a whole-genome CNV analysis [13, 16, 34] (Fig. 1A). Among these patients, we could establish iPS cells from one patient (SCZ3), which were used in the subsequent analysis as PT cell lines in this study.
To understand the biological effect of ASTN2 deletion on human neuronal cells, we also worked on the generation of an ASTN2-deleted iPS cell line from healthy control iPS cells (CON1) using the CRISPR/Cas9 system (Supplementary Methods). Five individual single-guide RNAs (sgRNAs) were constructed near the start point of the most upstream exon that all splicing variants have in common (ENSE00003567870, Fig. 1B and Supplementary Fig. 1A). Among the constructed sgRNAs, sgRNA#5 showed the strongest cleavage activity as examined by T7EI assay (Supplementary Fig. 1B). We also performed an off-target search using CCTop. CCTop predicted 13 potential off-target sites for sgRNA#5 in the human genome under the following conditions: core length = 12; max core mismatches = 2; max total mismatches = 3. However, the only target at an exonic position was in the ASTN2 gene region (Supplementary Table 4). Taken together, sgRNA#5 was selected for ASTN2-targeted genome editing.
As a result, one ASTN2-deleted iPS cell line was established (CON1_ASTN2−/− cell line). The deletion of each homologous chromosome in the CON1_ASTN2−/− cell line was confirmed by Sanger sequencing, with a 1-bp deletion on one allele and a 14-bp deletion on the other (Fig. 1C). The CON1_ASTN2−/− cell line expressed the pluripotent marker (TRA-1-60 and NANOG) as well as its parent iPS cell line CON1 (Fig. 1D), whereas the expression of ASTN2 in the CON1_ASTN2−/− iPS cell line was reduced compared with that in CON1 (Fig. 1E). We also verified the downregulation of ASTN2 expression in the PT_1 and PT_2 cell lines (Fig. 1E).
Differentiation of iPS cells into NSs and measurement of expression levels of neuronal markers
To investigate the mechanisms of ASTN2 deletion in neuronal cells, we differentiated each iPS cell line into NSs, which predominantly contain neuronal progenitors or stem cells. A differentiation scheme to NSs is shown in Fig. 2A. Each iPS cell line formed cell aggregates, including NSs, and no significant differences in their size were observed (Fig. 2B, C). Then, we examined ASTN2 mRNA expression levels in the NSs on day 21. Compared with the parental cell lines, the CON1_ ASTN2−/− and PT cell lines showed decreased mRNA expression of ASTN2, whereas the mRNA expression levels of ASTN1 were significantly increased (Fig. 2D). No significant differences in the expression levels of PAX6, TUBB3, and MAP2, which are markers of the pan-neuron cells, were found between each cell line (Fig. 2E), suggesting that they are equally differentiated into the neuronal cells.

A Schematic timeline for NS differentiation from iPS cells. B Phage images of the NS (Day 21) of each cell line. The scale bar indicates 500 µm. C Comparison of NS area in each cell line. N = 50. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. D, E Relative mRNA expression levels for ASTN1, ASTN2, PAX6, TUBB3, and MAP2 in NSs (day 21). N = 6. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. **p < 0.01.
We also evaluated the expression of ASTN2 protein in neurospheres using immunoblotting using anti-ASTN2 antibody (Sigma-Aldrich #ZRB1283). We obtained a band at ~105 kDa that are expected size by the manufacturer’s document. However, the immunogen used to generate the anti-ASTN2 antibody (18 amino acids from the C-terminal half, lumenal domain of ASTN2) contains the short isoform region of ASTN2 (~50 kDa). Therefore, we evaluated the ASTN2 expression levels considering both the bands (~105 and ~50 kDa). The expression level of ~50 kDa ASTN2 rather than ~105 kDa was decreased in ASTN2-deleted neurospheres (patient-derived and genome-edited neurospheres) compared to control neurospheres (Supplementary Fig. 2).
NSs derived from a patient with ASTN2-deleted SCZ and CON1_ASTN2−/− cell line showed suppressed neurite outgrowth
Studies have shown that the decreased expression of psychiatric disorder-associated genes suppresses neurite outgrowth [27, 35,36,37]. Therefore, we observed how ASTN2 deletion affects neurite outgrowth.
NSs from each cell line were dissociated into single cells once and seeded onto poly-L-ornithine/laminin/fibronectin-coated plates and observed by timelapse imaging for 3 days. As a result, the CON1_ASTN2−/− and PT cell lines showed suppressed neurite outgrowth compared with the control cell lines at almost all time points (Fig. 3A, B). In agreement with this, the immunocytochemistry analysis showed that the βIII-tubulin (a marker for neurons)-expressing neurites in ASTN2-deleted cell lines were shorter than those in the control cell lines 72 h after seeding onto the plates (Fig. 3C, D). Considering that no differences in the ratio of βIII-tubulin- and vGLUT1-expressing cells were observed between cell lines (Supplementary Fig. 3), the suppressed neurite outgrowth in ASTN2-deleted cell lines was not due to the differences in the degree of differentiation of each cell line.

A Timelapse images of pan-neuronal cell outgrowth of each cell line using IncuCyte ZOOM™ software. Scale bars indicate 200 µm. Green: cell body, Pink: neurite. B Neurite length measured in the indicated time after plating. N = 5. Bars represent the mean ± standard error. Repeated-measure multiple ANOVA method was used for analysis. Moreover, Bonferroni adjustment for multiple comparisons was performed. C Representative ICC images of pan-neuronal cells 72 h after plating. Immunostained for βIII-tubulin and DAPI. The white bars in the image indicate 200 µm. D Measurement of the longest neurite using ICC images. N = 75. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. **p < 0.01.
Comprehensive expression analysis of the CON1_ ASTN2−/− cell line compared with its parental cell line showed decreased expression of ZNF558
Next, to explore the molecular and biological effects of ASTN2 deletion, CON1_ASTN2−/− and its parental NSs were applied to the comprehensive gene expression analysis. The analysis of differentially expressed genes in the CON1_ASTN2−/− cell line identified 1241 upregulated entities and 741 downregulated entities (T-test cutoff p < 0.05, fold change >2, Supplementary Fig. 4A, and Supplementary Table 5). The upregulated genes were mainly associated with cell localization or motility (Supplementary Fig. 4B and Supplementary Table 6) In contrast, the downregulated genes were enriched in GO terms in transcription factor activity (Supplementary Fig. 4B and Supplementary Tables 6,7). The Bonferroni correction for multiple comparisons corrected 1,982 entities to 42 entities (including four upregulated and 16 downregulated protein-coding genes), of which ZNF558 had been identified as the most downregulated gene in the CON1_ASTN2−/− cell line compared with the parental NSs (Fig. 4A). The expression of ZNF558 was extremely downregulated in the subsequent RT-qPCR in both the CON1_ASTN2−/− and PT cell lines (Fig. 4B). This result suggests that ASTN2 deletion is involved in the pathogenesis of psychiatric disorders by downregulating the expression of ZNF558. Therefore, we focused on ZNF558 and performed further analysis.

A Differentially expressed genes (protein coding) with fold change >2, Bonferroni correction of p value <0.05. Red/blue indicates genes that have increased/decreased in CON1_ASTN2−/− vs. CON1. B Relative mRNA expression level of ZNF558. N = 6. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. **p < 0.01. C Number of peaks overlapping between datasets. D Image of called peaks most plausible in each dataset. E Five most significant genes around the peaks called for each ChIP-seq dataset and its distance from TSS. F Relative mRNA expression level of SPATA18. N = 6. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. *p < 0.05; **p < 0.01.
Analysis of the DNA binding sites of ZNF558
The zinc finger protein 558 encoded by ZNF558 is a protein that belongs to the Krüppel-associated box domain zinc finger protein (KZFP) family and is known to bind to DNA [38, 39]. A report suggested that ZNF558 binds with the DNA region of SPATA18, a gene involved in mitophagy [31]. We attempted to confirm the reproducibility by analyzing multiple ChIP-seq datasets targeting ZNF558 from public databases.
We used three sets of ChIP-seq data available on the SRA (https://www.ncbi.nlm.nih.gov/sra) (SRR5197178, SRR5111604, and SRR5111605) for ZNF558 using the HEK293 cell line. SRR5111604 and SRR5111605 were duplicate data from the same sample (Fig. 4C). The analysis pipeline and the result of region-gene association is shown in Supplementary Fig. 5A–D. The highest scoring peak was called near the transcriptional start sight of SPATA18 in each dataset and even merged data from the three datasets (Fig. 4D, E and Supplementary Table 8). In agreement with this result, RT-qPCR analysis showed that the expression level of SPATA18 was significantly increased in the PT and CON1_ASTN2−/− cell lines compared with that in the control cell lines (Fig. 4F).
Increased mitophagy and decreased ratio of the copy number of mtDNA/nucDNA in the CON1_ ASTN2−/− and PT cell lines
According to previous reports, the mitophagy-related gene SPATA18 (also known as mitochondria-eating protein) enhances mitophagy in the extracellular matrix [31, 33, 40]. Therefore, we compared the intensity of mitophagy among the cell lines. The NSs were stained with mitophagy dye on day 21, and their intensity was measured using flow cytometry. The experimental scheme is shown in Fig. 5A. As a result, the PT and CON1_ASTN2-/- cell lines increased the fraction of high fluorescence intensity by 2–3 folds compared with the control cell lines (Fig. 5B and Supplementary Fig. 6A,B). Additionally, previous reports suggest that increased mitophagy consequently alters the mtDNA/nucDNA copy number ratio [31, 40]. In agreement with the result of flow cytometry analysis, we found that the mtDNA/nucDNA copy number ratio was reduced in the PT and CON1_ASTN2−/− cell lines (Fig. 5C). These results confirm the enhanced mitophagy in ASTN2-deleted cell lines.

A Schematic timeline for flow cytometry analysis. B Results of flow cytometry analysis. N = 4. The red bar indicates the average value. Multiple comparison was performed using Dunnett’s test. **p < 0.01. C MtDNA/nucDNA copy number ratio. N = 6. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. *p < 0.05; **p < 0.01. D Representative images of the CON2 cell line with H2O2 or rapamycin. Images were obtained 72 h after plating. Green: cell body, Pink: neurite. The scale bar indicates 200 µm. E Neurite length of the CON2 cell lines 72 h after H2O2 or rapamycin was added. N = 4. Bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. **p < 0.01. F The results of the flow cytometry analysis with the addition of H2O2 or rapamycin. N = 3. Multiple comparison was performed using Dunnett’s test. *p < 0.05; **p < 0.01. G The representative images of the PT cell line with 0.1 µM or 0.5 µM chloroquine. The images were obtained 72 h after plating. Green, cell body; pink, neurite. The scale bar indicates 200 µm. H The neurite length of the PT cell line was 72 h after 0.1 µM or 0.5 µM chloroquine was added (N = 4). The bars represent the mean ± standard error. Multiple comparison was performed using Dunnett’s test. *p < 0.05.
Finally, we investigated whether the increased mitophagy affects neurite outgrowth. As shown in Fig. 5D–F, H2O2 [30] and rapamycin [29], which increase mitophagy, attenuated the neurite outgrowth of CON2 cell lines compared with the vehicle. Moreover, we found that chloroquine, an inhibitor of mitophagy, improved neurite outgrowth in the PT cell line (Fig. 5G, H). Overall, these findings suggest that the impaired neurite outgrowth of ASTN-deleted cells is attributed to mitophagy promotion.
Discussion
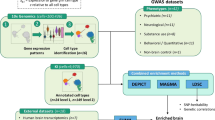
In this study, we showed how ASTN2 deletion, a candidate cross-disorder risk variant for psychiatric disorders, affects human neuronal cells. To the best of our knowledge, this study is the first report of molecular biological analysis using patient-derived iPS cells with ASTN2 deletion. We found that both the patient-derived and genome-edited ASTN2-deleted neuronal cells showed shortened neurite outgrowth, reduced expression of ZNF558, increased expression of SPATA18, increased mitophagy, and reduced mtDNA/nucDNA copy number ratio compared with healthy control neuronal cells (Fig. 6). These findings suggest that ASTN2 is involved in regulating mitochondrial quality, which may be implicated in the pathogenesis of various psychiatric disorders.

Graphical abstract of this research.
To determine the effects of ASTN2 deletion in human neuronal cells, we performed gene editing, in addition to using iPS cells derived from a patient with ASTN2 deletion. The exons we targeted using the CRISPR/Cas9 system to create ASTN2-deleted cell lines were not the same as the deleted exons in the SCZ3 patient. The CON1_ASTN2−/− and PT cell lines differed in the following aspects: frameshift due to single and 14 bases of deletion in ENST00000313400.9 exon 17, or a frameshift due to 410 bases of deletion over exons 14, 15, and 16. However, these two cell lines showed almost similar phenotypes, in terms of the degree of neurite outgrowth, the mRNA expression of ZNF558 and SPATA18, and the intensity of mitophagy. To address this, we aimed to quantify the ASTN2 protein of neurospheres. ASTN2-deleted neurospheres (patient-derived and genome-edited neurospheres), showed a decrease in ~50 kDa ASTN2 protein levels (Supplementary Fig. 2). ASTN2 has many splicing variants (Fig. 1B). Thus, it is probable that the ~50 kDa ASTN2 protein is the mainly expressed variant in neurospheres and the ~105 kDa band protein is a nonspecific one. However, the unexpected result in which the genome-edited cells (CON1_ASTN2−/−)—with biallelic out-of-frame variants leading to an almost complete knockout of ASTN2—continued to express ASTN2 protein, remains unexplained. The potential limitations of the CRISPR technology may explain this result. Genome editing using the CRISPR-Cas9 system may sometimes cause the expression of unanticipated proteins, similar in size to the wild-type protein, from the out-of-frame variant alleles [41, 42]. Moreover, the non-sense mediated mRNA decay efficiency of CRISPR-Cas9 system-induced frameshift knock-out variants is inconsistent; and complete loss of protein expression is not achieved [43]. This may explain the continued expression of ASTN2 in our genome-edited cells (CON1_ASTN2−/− cells).
Based on our results, it seems that patient-derived and genome-edited cells have the similar reduced expression level of ASTN2. Hence, it can be concluded that these phenotypes are caused by the reduced expression of ASTN2. Moreover, given that this region of ASTN2, including the target site of CRISPR/Cas9 and the deleted region in SCZ3, is a disease-susceptible region (gray-shaded area in Fig. 1A) [17], these phenotypes will also reflect the vulnerabilities for the onset of psychiatric disorders at the cellular and molecular levels.
The brain is composed of various types of neuronal and glial cells, all of which are generated by the differentiation of neural stem cells (NSCs). Such NSC activities from embryonic to early childhood must be precisely controlled. The function of transcription factors, a protein family that regulates gene transcription, is crucial for differentiating NSCs into various nervous system cells and their subsequent formation into various brain regions [44]. In a study of postmortem brains, Voineagu et al. revealed that gene expression dysregulation related to brain development in the embryonic period may be associated with the occurrence of ASD [45]. Furthermore, a recent CNV analysis study of patients with SCZ and ASD has shown that CNV was significantly enriched in enhancers and promoters in brain tissue, suggesting that gene expression dysregulation is involved in the pathogenesis of SCZ and ASD [13]. C2H2 zinc finger proteins are the largest family of human transcription factors [46] and are highly expressed in the developing brain. They have been shown to regulate transcription patterning of the early central nervous system and play an important role in regulating brain development [47]. A 335.4-kb duplication in the Xp11.2p11.3 region, which contains the C2H2 zinc finger proteins, ZNF81 and ZNF182, was identified in a patient showing developmental delays, autistic features, and growth and speech delays [48]. Another study on a large cohort of probands with ASD in China identified ZNF292 as a candidate risk gene [49]. Additionally, another analysis of the Autism Genetic Research Exchange cohort, consisting of approximately 1,000 multiplex ASD families, identified homozygous and compound heterozygous variants in the ZNF18 gene [50]. The KRAB zinc finger protein family, a subtype of the C2H2-ZNF protein family characterized by its ZNF and KRAB domains, is expressed characteristically in the human brain compared with other primates, such as chimpanzees. The KRAB zinc finger protein is necessary for the transcriptional network that drives the unique features of the human brain, such as increased cognitive activity and larger brain size [47]. In this study, we found that the ZNF558 gene was most differentially expressed in the CON1_ASTN2−/− cell line compared with that in its parental cell line. This result was also confirmed in the PT cell line, a heterozygous cell line with ASTN2 deletion. Another report has shown that not only by the knockout of ASTN2 but also by the knockout of other ASD-related genes, such as ATRX, AFF2, KCNQ2, SCN2A, and DLGAP2, the expression of ZNF558 is decreased [51]. Similarly, the expression of several other C2H2-ZNF family proteins was also decreased by the knockout of ASD-related genes [51]. These findings suggest that phenotypes detected in the ASTN2-deleted neurons, which are deficient in ZNF558 expression, may reflect the dysregulation of transcriptional systems or dysfunction of some higher brain functions related to ASD and SCZ.
Another aspect of the important function of NSCs in normal brain development is neural cell migration [52]. The important thing is to migrate to the right place at the right time. Reports have shown that functional impairment of proteins involved in neural cell migration, such as RELN [53,54,55] and NRG1/ERBB4 [56], is also associated with the pathogenesis of some neurological syndromes, including SCZ and BP [57]. ASTN1 is a ligand for neuron–glia junctions during the migration of neural progenitors along the radial glia in early embryos. In contrast, rather than directly controlling neuron–glia adhesions, ASTN2 regulates ASTN1 surface expression by forming a complex with ASTN1 in endocytosis—indirectly controlling the neural cell migration [58]. Thus, ASTN1 and ASTN2 complementarily and conjugately regulate neuronal migration during brain development. As an important case study, Mitani et al. reported brain malformation and abnormally shaped skulls as a result of the presence of double-heterozygous variants of both ASTN1 and ASTN2 [59]. These findings suggest that ASTN2 and ASTN1 play important roles in neuron localization and brain morphogenesis in early embryos or immature nervous systems. In this study, the mRNA expression level of ASTN1 was increased in both the PT and CON1_ASTN2−/− NSs compared with that in healthy control NSs. NSs consist of immature neural progenitors. Considering the indispensability and cooperativeness of ASTN1 and ASTN2, the upregulation of ASTN1 expression in ASTN2-deleted NSs may be due to compensation for the deletion of ASTN2. In fact, in the comprehensive expression analysis of NSs conducted in this study, the upregulated genes in the ASTN2-deleted cell line were most enriched in the category related to cell migration, cell localization, and cell motility.
In this study, we analyzed the DNA binding sites of ZNF558 using ChIP-seq data on the SRA. These are the results performed using HEK293T cells; therefore, we cannot rule out the possibility of different results when performed using neuronal cells, such as NSs. Nevertheless, the results were considered reasonable, as all three datasets showed the most specific binding to the transcription start sites (TSS) flanking region of SPATA18, and mRNA expression levels in the NS also showed differences between cell lines. Furthermore, a report confirmed that knocking out ZNF558 predominantly increased SPATA18 expression [31], which is consistent with the present results. This report also indicated that the expression of ZNF558 is regulated by the copy number of its downstream VNTR [31]. Therefore, we measured the respective VNTR copy numbers in the healthy control, PT, and CON1_ASTN2−/− cell lines and found that the VNTR copy number increased in the PT cell lines but did not differ from that in the healthy control in the CON1_ASTN2−/− cell lines (Supplementary Fig. 7). This suggests that a causal relationship was observed between ASTN2 and ZNF558 other than by VNTR copy number. As mentioned previously, the knockout of other ASD-related genes, aside from ASTN2, also causes the decreased expression of ZNF558. The identification of the regulatory system of ZNF558 downregulation will cut through the revealing molecular mechanism of psychiatric disorders, which should be addressed in future research.
We confirmed increased levels of mitophagy and reduced mtDNA/nucDNA copy number ratio in the PT and CON1_ASTN2−/− cell lines, both of which showed upregulated expression of SPATA18. According to a previous report, the overexpression of SPATA18 may efficiently eliminate abnormal mitochondria and reduce the mtDNA/nucDNA ratio [60]. Abnormal mitochondrial morphology has been reported in various neurodevelopmental disorders, including SCZ and tuberous sclerosis, which is a comorbidity associated with ASD in approximately half of patients [61,62,63]. Additionally, reduced numbers and size of mitochondria have been observed in SCZ patients [64, 65]. It is possible that abnormal mitochondria accumulate in the PT and CON1_ASTN2−/− cell lines where mitophagy eliminates these abnormal mitochondria.
We found that neurite outgrowth is inhibited in ASTN2-deleted neuronal cells and this impairment is associated with their increased mitophagy. Neurites develop immediately after a neuron differentiates from NSCs, and one of several neurites begins to elongate rapidly and takes on the characteristics of an axon during brain development. On that occasion, mitochondria aggregate at the base of the future axon before neurite outgrowth, indicating that mitochondria play an important role in neurite outgrowth [66]. Moreover, if treated with EtBr, mtDNA is selectively and irreversibly damaged, and cells are no longer able to gain polarity and develop axons [67]. Based on the findings of these previous reports and those of the present study, further research on mitochondria is needed to better understand the underlying mechanisms of psychiatric disorders based on ASTN2 deletion.
Here, we used rapamycin and H2O2 to induce mitophagy. Rapamycin induces mitophagy by inhibiting the PINK1 and PRKN-activated mTOR pathway [68]. Indeed, rapamycin-treated cells showed increased mitophagy with shortened neurites (Fig. 5D–F). Chloroquine, an inhibitor of mitophagy, improves neurite outgrowth. Thus, the impaired neurite outgrowth in rapamycin-treated cells is probably due to increased mitophagy. In contrast, H2O2-treated cells show impaired neurite outgrowth, but mitophagy remains unaltered (Fig. 5D–F). H2O2, a reactive oxygen species (ROS), induces mitophagy by generating highly oxidative hydroxyl radicals [69]. Counterintuitively, H2O2 may inhibit autophagy by impairing LC3 lipidation which is a key process for autophagosome induction and autophagosome-lysosome integration [70]. This may explain why H2O2-treatment failed to increase mitophagy in this study. Moreover, cell damage induced by ROS may explain the shortened neurites observed in the H2O2-treated cells, here.
Our final goal was to elucidate the molecular and cellular pathophysiology in the brains of patients with ASTN2 deletion. To address this, we used induced pluripotent stem cell-derived pan-neuronal cells as a lead to understand them in this study. Pan-neuronal cells mainly consist of cortical neurons but partially include the nonspecific neuronal subtype, and, therefore, are useful for getting an overview of the findings in neuronal cells. Indeed, ASTN2-deleted pan-neuronal cells provided us with novel findings, as mentioned above. For the next strategy, we can focus on each neuronal subtype, such as Purkinje cells in the cerebellum, where ASTN2 is highly expressed. Large-scale de novo variant analyses in patients with ASD have shown that ASD-related candidate genes in early embryos are enriched in the cerebellum [71], and cerebellar dysfunction seems deeply implicated in the pathophysiology of ASD [72]. Moreover, Purkinje cells are representative cells with increasing rates of mitophagy [30]. Because the technology to generate cerebellar organoids from iPS cells has been recently reported, it will be possible to apply that technology to our iPS cells [73]. Starting with our study, more detailed analyses are expected to achieve the goal.
This study has several limitations. First, variants of ASTN2 have been identified in several psychiatric disorders such as SCZ, ASD, and BP. However, we could only access the SCZ patient with ASTN2 deletion to generate iPS cells, which is considered the main limitation of this study. To determine the mechanism of cross-disorder psychiatric disorders based on ASTN2, further studies using iPS cells derived from ASD and BP patients are needed. Second, validation of our findings by direct analysis of neurons in a fetal patient’s brain is not feasible because psychiatric disorders are not diagnosed during the fetal period. However, for over three decades, SCZ has been regarded as a neurodevelopmental disorder [74]. Consistent with this, patient iPS cell-derived neurons often exhibit shortened neurites [75, 76], implying that the impairment of neurite length during the neurodevelopmental stage is a hallmark of psychiatric disorders, including SCZ. Therefore, our findings will contribute to reveal the mechanism of psychiatric disorders in addition to previous studies.
In conclusion, by studying disease- and genetic-related phenotypes at the human cellular level, this study provided one possible hypothesis on how ASTN2, a candidate cross-disorder risk variant for psychiatric disorders, is involved in the pathogenesis of these disorders and throws new insights into the understanding of their molecular and cellular mechanisms. This research will play a role as a stepping stone to reveal the pathomechanisms of psychiatric disorders and contribute to the development of even more effective therapies in the future.
Data availability
All data supporting our findings can be found in the main paper or in supplementary files. Gene expression data are available via the GEO accession number: GSE260713.
References
Baba K, Guo WJ, Chen YR, Nosaka T, Kato T. Burden of schizophrenia among Japanese patients: a cross-sectional National Health and Wellness Survey. BMC Psychiatry. 2022;22:410.
Simeone JC, Ward AJ, Rotella P, Collins J, Windisch R. An evaluation of variation in published estimates of schizophrenia prevalence from 1990-2013: a systematic literature review. BMC Psychiatry. 2015;15:193.
Saito M, Hirota T, Sakamoto Y, Adachi M, Takahashi M, Osato-Kaneda A, et al. Prevalence and cumulative incidence of autism spectrum disorders and the patterns of co-occurring neurodevelopmental disorders in a total population sample of 5-year-old children. Mol Autism. 2020;11:35.
Sasayama D, Kuge R, Toibana Y, Honda H. Trends in autism spectrum disorder diagnoses in Japan, 2009 to 2019. JAMA Netw Open. 2021;4:e219234.
Schendel DE, Thorsteinsson E. Cumulative incidence of autism into adulthood for birth cohorts in Denmark, 1980-2012. JAMA. 2018;320:1811–3.
Kato T, Baba K, Guo WJ, Chen YR, Nosaka T. Impact of bipolar disorder on health-related quality of life and work productivity: estimates from the national health and wellness survey in Japan. J Affect Disord. 2021;295:203–14.
Clemente AS, Diniz BS, Nicolato R, Kapczinski FP, Soares JC, Firmo JO, et al. Bipolar disorder prevalence: a systematic review and meta-analysis of the literature. Braz. J. Psychiatry. 2015;37:155–61.
Nishi D, Ishikawa H, Kawakami N. Prevalence of mental disorders and mental health service use in Japan (vol 73, pg 458, 2019). Psychiatry Clin Neurosci. 2019;73:767.
Demyttenaere K, Bruffaerts R, Posada-Villa J, Gasquet I, Kovess V, Lepine JP, et al. Prevalence, severity, and unmet need for treatment of mental disorders in the World Health Organization World Mental Health Surveys. JAMA. 2004;291:2581–90.
Koshiyama D, Fukunaga M, Okada N, Morita K, Nemoto K, Usui K, et al. White matter microstructural alterations across four major psychiatric disorders: mega-analysis study in 2937 individuals. Mol Psychiatry. 2020;25:883–95.
Schwarz K, Moessnang C, Schweiger JI, Baumeister S, Plichta MM, Brandeis D, et al. Transdiagnostic prediction of affective, cognitive, and social function through brainreward anticipation in schizophrenia, bipolar disorder, major depression, and autism spectrum diagnoses. Schizophr Bull. 2020;46:592–602.
Nakatochi M, Kushima I, Ozaki N. Implications of germline copy-number variations in psychiatric disorders: review of large-scale genetic studies. J Hum Genet. 2021;66:25–37.
Kushima I, Aleksic B, Nakatochi M, Shimamura T, Okada T, Uno Y, et al. Comparative analyses of copy-number variation in autism spectrum disorder and schizophrenia reveal etiological overlap and biological insights. Cell Rep. 2018;24:2838–56.
Gandal MJ, Haney JR, Parikshak NN, Leppa V, Ramaswami G, Hartl C, et al. Shared molecular neuropathology across major psychiatric disorders parallels polygenic overlap. Science. 2018;359:693–7.
Guan JT, Cai JJ, Ji GL, Sham PC. Commonality in dysregulated expression of gene sets in cortical brains of individuals with autism, schizophrenia, and bipolar disorder. Transl. Psychiatry. 2019;9:152.
Kushima I, Nakatochi M, Aleksic B, Okada T, Kimura H, Kato H, et al. Cross-disorder analysis of genic and regulatory copy number variations in bipolar disorder, schizophrenia, and autism spectrum disorder. Biol Psychiatry. 2022;92:362–74.
Lionel AC, Tammimies K, Vaags AK, Rosenfeld JA, Ahn JW, Merico D, et al. Disruption of the ASTN2/TRIM32 locus at 9q33. 1 is a risk factor in males for autism spectrum disorders, ADHD and other neurodevelopmental phenotypes. Hum Mol Genet. 2014;23:2752–68.
Wilson PM, Fryer RH, Fang Y, Hatten ME. Astn2, a novel member of the astrotactin gene family, regulates the trafficking of ASTN1 during glial-guided neuronal migration. J Neurosci. 2010;30:8529–40.
Behesti H, Fore TR, Wu P, Horn Z, Leppert M, Hatten ME. ASTN2 modulates synaptic strength by trafficking and degradation of surface proteins. Proc Natl Acad Sci USA. 2018;115:E9717–E9726.
Ito, T, Yoshida M, Aida T, Kushima I, Hiramatsu Y, Ono M et al. Astrotactin2 (ASTN2) regulates emotional and cognitive functions by affecting neuronal morphogenesis and monoaminergic systems. J. Neurochem. 2023;65(2):211–29.
Fang RX, Xia CL, Close JL, Zhan M, He J, Huang ZK, et al. Conservation and divergence of cortical cell organization in human and mouse revealed by MERFISH. Science. 2022;377:56.
Kaiser T, Feng GP. Modeling psychiatric disorders for developing effective treatments. Nat. Med. 2015;21:979–88.
Hodge RD, Bakken TE, Miller JA, Smith KA, Barkan ER, Graybuck LT, et al. Conserved cell types with divergent features in human versus mouse cortex. Nature. 2019;573:61.
Lee KM, Hawi ZH, Parkington HC, Parish CL, Kumar PV, Polo JM, et al. The application of human pluripotent stem cells to model the neuronal and glial components of neurodevelopmental disorders. Molecular Psychiatry. 2020;25:368–78.
Ardhanareeswaran K, Mariani J, Coppola G, Abyzov A, Vaccarino FM. Human induced pluripotent stem cells for modelling neurodevelopmental disorders. Nat Rev Neurol. 2017;13:265–78.
Arioka Y, Kushima I, Kubo H, Mori D, Ozaki N. Induced pluripotent stem cells derived from a schizophrenia patient with ASTN2 deletion. Stem Cell Res. 2018;30:81–4.
Arioka Y, Shishido E, Kushima I, Suzuki T, Saito R, Aiba A, et al. Chromosome 22q11.2 deletion causes PERK-dependent vulnerability in dopaminergic neurons. EBioMedicine. 2021;63:103138.
Fujimori K, Matsumoto T, Kisa F, Hattori N, Okano H, Akamatsu W. Escape from pluripotency via inhibition of TGF-β/BMP and activation of Wnt signaling accelerates differentiation and aging in hPSC progeny cells. Stem Cell Rep. 2017;9:1675–91.
Palikaras K, Lionaki E, Tavernarakis N. Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat Cell Biol. 2018;20:1013–22.
Dan XL, Babbar M, Moore A, Wechter N, Tian JY, Mohanty JG, et al. DNA damage invokes mitophagy through a pathway involving Spata18. Nucleic Acids Res. 2020;48:6611–23.
Johansson PA, Brattas PL, Douse CH, Hsieh PH, Adami A, Pontis J, et al. A cis-acting structural variation at the ZNF558 locus controls a gene regulatory network in human brain development. Cell Stem Cell. 2022;29:52.
McLean CY, Bristor D, Hiller M, Clarke SL, Schaar BT, Lowe CB, et al. GREAT improves functional interpretation of cis-regulatory regions. Nat Biotechnol. 2010;28:495–U155.
Rooney JP, Ryde IT, Sanders LH, Howlett EH, Colton MD, Germ KE, et al. PCR based determination of mitochondrial DNA copy number in multiple species. Methods Mol Biol. 2015;1241:23–38.
Kushima I, Aleksic B, Nakatochi M, Shimamura T, Shiino T, Yoshimi A, et al. High-resolution copy number variation analysis of schizophrenia in Japan. Mol Psychiatry. 2017;22:430–40.
Hashimoto R, Nakazawa T, Tsurusaki Y, Yasuda Y, Nagayasu K, Matsumura K, et al. Whole-exome sequencing and neurite outgrowth analysis in autism spectrum disorder. J Hum Genet. 2016;61:199–206.
Sekiguchi M, Sobue A, Kushima I, Wang C, Arioka Y, Kato H, et al. ARHGAP10, which encodes Rho GTPase-activating protein 10, is a novel gene for schizophrenia risk. Transl Psychiatry. 2020;10:247.
Toyoshima M, Akamatsu W, Okada Y, Ohnishi T, Balan S, Hisano Y, et al. Analysis of induced pluripotent stem cells carrying 22q11.2 deletion. Transl Psychiatry. 2016;6:e934.
Ecco G, Imbeault M, Trono D. KRAB zinc finger proteins. Development. 2017;144:2719–29.
Imbeault M, Helleboid PY, Trono D. KRAB zinc-finger proteins contribute to the evolution of gene regulatory networks. Nature. 2017;543:550–4.
Okuyama K, Kitajima Y, Egawa N, Kitagawa H, Ito K, Aishima S, et al. Mieap-induced accumulation of lysosomes within mitochondria (MALM) regulates gastric cancer cell invasion under hypoxia by suppressing reactive oxygen species accumulation. Sci. Rep. 2019;9:2822.
Makino S, Fukumura R, Gondo Y. Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9. Sci Rep. 2016;6:39608.
Tuladhar R, Yeu Y, Tyler Piazza J, Tan Z, Rene Clemenceau J, Wu X, et al. CRISPR-Cas9-based mutagenesis frequently provokes on-target mRNA misregulation. Nat Commun. 2019;10:4056.
Smits AH, Ziebell F, Joberty G, Zinn N, Mueller WF, Clauder-Münster S, et al. Biological plasticity rescues target activity in CRISPR knock outs. Nat Methods. 2019;16:1087–93.
Silbereis JC, Pochareddy S, Zhu Y, Li MF, Sestan N. The cellular and molecular landscapes of the developing human ventral nervous system. Neuron. 2016;89:248–68.
Voineagu I, Wang XC, Johnston P, Lowe JK, Tian Y, Horvath S, et al. Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature. 2011;474:380.
Emerson RO, Thomas JH. Adaptive evolution in zinc finger transcription factors. PLos Genet. 2009;5:e1000325.
Nowick K, Gernat T, Almaas E, Stubbs L. Differences in human and chimpanzee gene expression patterns define an evolving network of transcription factors in brain. Proc Natl Acad Sci USA. 2009;106:22358–63.
Alesi V, Bertoli M, Barrano G, Torres B, Pusceddu S, Pastorino M, et al. 335.4 kb microduplication in chromosome band Xp11.2p11.3 associated with developmental delay, growth retardation, autistic disorder and dysmorphic features. Gene. 2012;505:384–7.
Wang TY, Guo H, Xiong B, Stessman HAF, Wu HD, Coe BP, et al. De novo genic mutations among a Chinese autism spectrum disorder cohort. Nat. Communications. 2016;7:13316.
Chahrour MH, Yu TW, Lim ET, Ataman B, Coulter ME, Hill RS, et al. Whole-exome sequencing and homozygosity analysis implicate depolarization-regulated neuronal genes in autism. PLos Genet. 2012;8:236–44.
Deneault E, White SH, Rodrigues DC, Ross PJ, Faheem M, Zaslavsky K, et al. Complete disruption of autism-susceptibility genes by gene editing predominantly reduces functional connectivity of isogenic human neurons. Stem Cell Rep. 2019;12:427–9.
Bond AM, Ming GL, Song HJ. Adult mammalian neural stem cells and neurogenesis: five decades later. Cell Stem Cell. 2015;17:385–95.
Arioka Y, Hirata A, Kushima I, Aleksic B, Mori D, Ozaki N. Characterization of a schizophrenia patient with a rare RELN deletion by combining genomic and patient-derived cell analyses. Schizophr Res. 2020;216:511–5.
Folsom TD, Fatemi SH. The involvement of Reelin in neurodevelopmental disorders. Neuropharmacology. 2013;68:122–35.
Arioka Y, Shishido E, Kubo H, Kushima I, Yoshimi A, Kimura H, et al. Single-cell trajectory analysis of human homogenous neurons carrying a rare RELN variant. Transl Psychiatry. 2018;8:12.
Banerjee A, MacDonald ML, Borgmann-Winter KE, Hahn CG. Neuregulin 1-erbB4 pathway in schizophrenia: From genes to an interactome. Brain Res Bull. 2010;83:132–9.
Valiente M, Marin O. Neuronal migration mechanisms in development and disease. Curr Opin Neurobiol. 2010;20:68–78.
Horn Z, Behesti H, Hatten ME. N-cadherin provides a cis and trans ligand for astrotactin that functions in glial-guided neuronal migration. Proc Natl Acad Sci USA. 2018;115:10556–63.
Mitani T, Isikay S, Gezdirici A, Gulec EY, Punetha J, Fatih JM, et al. High prevalence of multilocus pathogenic variation in neurodevelopmental disorders in the Turkish population. Am J Hum Genet. 2021;108:1981–2005.
Mussazhanova Z, Shimamura M, Kurashige T, Ito M, Nakashima M, Nagayama Y. Causative role for defective expression of mitochondria-eating protein in accumulation of mitochondria in thyroid oncocytic cell tumors. Cancer Sci. 2020;111:2814–23.
Ebrahimi-Fakhari D, Saffari A, Wahlster L, DiNardo A, Turner D, Lewis TL Jr, et al. Impaired mitochondrial dynamics and mitophagy in neuronal models of tuberous sclerosis complex. Cell Rep. 2016;17:2162.
Hjelm BE, Rollins B, Mamdani F, Lauterborn JC, Kirov G, Lynch G, et al. Evidence of mitochondrial dysfunction within the complex genetic etiology of schizophrenia. Mol Neuropsychiatry. 2015;1:201–19.
Rajasekaran A, Venkatasubramanian G, Berk M, Debnath M. Mitochondrial dysfunction in schizophrenia: pathways, mechanisms and implications. Neurosci Biobehav Rev. 2015;48:10–21.
Uranova N, Orlovskaya D, Vikhreva O, Zimina I, Kolomeets N, Vostrikov V, et al. Electron microscopy of oligodendroglia in severe mental illness. Brain Res Bull. 2001;55:597–610.
Shivakumar V, Rajasekaran A, Subbanna M, Kalmady SV, Venugopal D, Agrawal R, et al. Leukocyte mitochondrial DNA copy number in schizophrenia. Asian J Psychiatry. 2020;53:102193.
Mattson MP, Gleichmann M, Cheng A. Mitochondria in neuroplasticity and neurological disorders. Neuron. 2008;60:748–66.
Mattson MP, Partin J. Evidence for mitochondrial control of neuronal polarity. J Neurosci Res. 1999;56:8–20.
Wu L, Feng Z, Cui S, Hou K, Tang L, Zhou J, et al. Rapamycin upregulates autophagy by inhibiting the mTOR-ULK1 pathway, resulting in reduced podocyte injury. PLoS ONE. 2013;8:e63799.
Zhou J, Li XY, Liu YJ, Feng J, Wu Y, Shen HM, et al. Full-coverage regulations of autophagy by ROS: from induction to maturation. Autophagy. 2022;18:1240–55.
Frudd K, Burgoyne T, Burgoyne JR. Oxidation of Atg3 and Atg7 mediates inhibition of autophagy. Nat Commun. 2018;9:95.
Takata A, Miyake N, Tsurusaki Y, Fukai R, Miyatake S, Koshimizu E, et al. Integrative analyses of de novo mutations provide deeper biological insights into autism spectrum disorder. Cell Rep. 2018;22:734–47.
Kelly E, Escamilla CO, Tsai PT. Cerebellar dysfunction in autism spectrum disorders: deriving mechanistic insights from an internal model framework. Neuroscience. 2021;462:274–87.
Muguruma K, Nishiyama A, Kawakami H, Hashimoto K, Sasai Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Rep. 2015;10:537–50.
Owen MJ, O’Donovan MC. Schizophrenia and the neurodevelopmental continuum:evidence from genomics. World Psychiatry. 2017;16:227–35.
Grunwald LM, Stock R, Haag K, Buckenmaier S, Eberle MC, Wildgruber D, et al. Comparative characterization of human induced pluripotent stem cells (hiPSC) derived from patients with schizophrenia and autism. Transl Psychiatry. 2019;9:179.
Ishii T, Ishikawa M, Fujimori K, Maeda T, Kushima I, Arioka Y, et al. In vitro modeling of the bipolar disorder and schizophrenia using patient-derived induced pluripotent stem cells with copy number variations of PCDH15 and RELN. ENeuro. 2019;6:ENEURO.0403–18.2019.
Acknowledgements
We thank all of the patients and their families for participating in this study. In addition, we thank all staff at the Department of Psychiatry, Nagoya University, for their assistance. This research was supported by the AMED under grant No. JP21wm0425007, JP19dm0207075, JP19ak0101113, JP21dk0307103, JP22ek0109601, JP22tm0424222, JP23ak0101215, JP22gm1410011, JP23gm1910005, JP23ek0109678, and the MEXT KAKENHI Grant Nos. 20K20602 and 21H04815.
Author information
Authors and Affiliations
Contributions
These authors contributed equally: Yu Hayashi and Hiroki Okumura. YA and NO conceived and designed the study. YH and HO performed the experiments and analyzed the data. YH, HO, and YA wrote the paper. YA, IK, DM, TL, GO, HK, YN, HK, BA, and NO reviewed and edited the manuscript. All authors have approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
Dr. N. Ozaki has received research support or speakers’ honoraria from or has served as a joint researcher with, or a consultant to, Sumitomo Pharma, Otsuka, Viatris, Eisai, Mochida, Kyowa Pharmaceutical Industry, Nihon Medi-Physics, Nippon Chemiphar, Medical Review, Nippon Boehringer Ingelheim, SUSMED, outside the submitted work.
Ethics approval and consent to participate
This study was performed in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Nagoya University (approval number: 2012-0184). All subjects provided written informed consent.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hayashi, Y., Okumura, H., Arioka, Y. et al. Analysis of human neuronal cells carrying ASTN2 deletion associated with psychiatric disorders. Transl Psychiatry 14, 236 (2024). https://doi.org/10.1038/s41398-024-02962-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-024-02962-4
