
Abstract
Macrolactam antibiotics incorporating β-amino acid polyketide starter units, isolated primarily from Actinomycetes species, show significant biological activities. This review provides a detailed analysis into the biosynthetic studies of vicenistatin, a macrolactam antibiotic with a 3-aminoisobutyrate starter unit, as well as biosynthetic research on related macrolactam compounds. Firstly, the elucidation of a common mechanism for the incorporation of β-amino acid starter units into the polyketide synthase (PKS) is described. Secondly, the unique biosynthetic mechanisms of the β-amino acids that are used to supply the main macrolactam biosynthetic pathways with starter units are discussed. Thirdly, some distinctive post-PKS modification mechanisms that complete macrolactam antibiotic biosynthesis are summarized. Finally, future directions for creating new macrolactam compounds through engineered biosynthesis pathways are described.
Similar content being viewed by others

Introduction
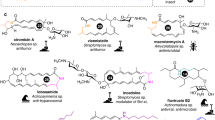
Macrolactam antibiotics that incorporate β-amino acids as polyketide starter units have been isolated mostly from the Actinomycetes species. These compounds exhibit significant biological activities, including antibacterial, antifungal, and antitumor effects [1]. Vicenistatin (1) [2], incednine (2) [3], cremimycin (3) [4], hitachimycin (stubomycin) (4) [5, 6], and fluvirucin B2 (Sch 38518, 5) [7, 8] are examples of such macrolactam antibiotics that have been selected as research targets in our laboratory (Fig. 1); however, many other related macrolactam compounds have also been discovered [9,10,11]. Vicenistatin (1) features 3-aminoisobutylate (3AIB, 6) as the polyketide starter unit. Similarly, incednine (2) has 3-aminobutyrate (3ABA, 7), cremimycin (3) has 3-aminononanoate (3ANA, 8), hitachimycin (4) has β-phenylalanine (β-Phe, 9), and fluvirucin B2 (5) has β-alanine (β-Ala, 10). The polyketide chain elongates from the β-amino acid starter unit and cyclizes between the β-amino group of the starter unit and the carboxylate moiety of the final extension product, yielding the corresponding macrolactam. As such, this class of macrolactams are biosynthesized via the typical polyketide pathway but with the incorporation of the unique β-amino acid starter units.

Macrolactam antibiotics studied in our laboratory
The biosynthesis of macrocyclic polyketides, and especially that of macrolactones, has been extensively studied [12,13,14] (Fig. 2). In general, acetate or propionate serves as the starter unit, and malonate and/or methylmalonate act as the extender units to construct the macrocyclic polyketide skeleton. The extender units typically exist as coenzyme A (CoA) thioesters, such as malonyl-CoA and methylmalonyl-CoA, which are transferred to the acyl carrier protein (ACP) domain of the polyketide synthase (PKS) by an acyltransferase (AT) domain, yielding malonyl-ACP/methylmalonyl-ACP [15]. Similarly, starter units are ligated to the ACP, forming acyl-ACP, and there are several methods to achieve this ligation [16]. The starter acyl-ACP is recognized by the β-ketosynthase (KS) domain. The acyl group is transferred to a cysteine residue of the active site of the KS domain and condensed with the extender malonyl-ACP/methylmalonyl-ACP to produce β-ketoacyl-ACP with the release of carbon dioxide. The β-carbonyl group of β-ketoacyl-ACP is subsequently reduced by the β-ketoreductase (KR) domain to yield β-hydroxyacyl-ACP, which is further processed by the dehydratase (DH) domain to generate α,β-unsaturated acyl-ACP. Finally, the enoyl reductase (ER) domain reduces α,β-unsaturated acyl-ACP to provide a fully reduced acyl-ACP.

General mechanism of type I polyketide synthase (PKS) for macrocyclic polyketides in bacteria
The second round of polyketide chain elongation is catalyzed by a different set of catalytic domains to extend the chain by one acetate unit. The number of rounds of extension determine the length of the corresponding polyketide and each round is catalyzed by a PKS module consisting of the essential catalytic domains: AT, ACP and KS. The degree of reduction of the polyketide chain is determined by the particular combination of additional tailoring domains (KR, DH, ER), within the module. For example, if the KR domain is absent, the β-carbonyl group remains in the polyketide chain; if the DH domain is absent, the β-hydroxy group remains; and if the ER domain is absent, an olefinic moiety remains. Finally, the thioesterase (TE) domain, located in the terminal PKS module, catalyzes an acyl transfer from the final thioester of the ACP-bound polyketide chain to form an acyl-TE complex, subsequently facilitating lactonization with a hydroxyl group on the elongated polyketide yielding a macrolactone. Post-PKS modifications, including polyketide skeletal modification, oxidation (hydroxylation, epoxidation), glycosylation, methylation, and acylation, are required to complete the biosynthesis of the dead-end polyketide compound [17].
In the biosynthesis of macrolactam antibiotics, the aforementioned PKS reaction is used to construct the polyketide skeleton; however, unique nitrogen-containing starter units are employed. There are several key points of interest regarding β-amino acid starter units. Firstly, their mechanism of incorporation into the PKS machinery. Secondly, as most β-amino acids are non-proteinogenic, their biosynthetic mechanisms are presumably unique. Lastly, the post-PKS modification of macrolactams appears crucial for their biological activities.
This review provides a detailed summary of the biosynthetic studies of vicenistatin, covering the entire biosynthetic pathway. A common mechanism for the incorporation of β-amino acids into the PKS machinery is outlined, emphasizing that adenylation enzymes, selective for β-amino acids, act as gatekeepers, thereby determining incorporation of the unique β-amino acid starter units. Next, the unique mechanisms of β-amino acid biosynthesis and post-PKS modification are described. Finally, future perspectives for creating new molecules based on these biosynthetic studies are discussed.
Vicenistatin: a macrolactam antibiotic with a β-amino acid starter unit
Vicenistatin (1) is a macrolactam antibiotic with 3-aminoisobutyric acid (3AIB, 6) as the starter unit of the polyketide backbone [2]. Vicenistatin (1) and its congener, vicenistatin M, are produced by Streptomyces halstedii HC34 [18]. Recently, Streptomyces parvus SCSIO Mla-L010 was also found to produce vicenistatin [19]. Vicenistatin exhibits antitumor activity against Co-3 human colon carcinoma cells [2].
3AIB (6) is a non-proteinogenic β-amino acid with a unique biosynthetic mechanism. Initially, it was unclear how 3AIB (6) was incorporated into the polyketide pathway, however, extensive feeding experiments with candidate amino acids have elucidated these mechanisms [20,21,22]. Critically, l-glutamic acid (11) and (2S,3S)-3-methylaspartic acid (3-MeAsp, 12) are incorporated into vicenistatin (Fig. 3) by the biosynthetic machinery, but 3AIB (6) and (2S,3R)-3-MeAsp (13) are not. Therefore, C–C bond rearrangement of l-Glu (11) results in the formation of (2S,3S)-3-MeAsp (12), presumably catalyzed by adenosylcobalamin (AdoCbl) and pyridoxal 5’-phosphate (PLP)-dependent glutamate mutase [23]. Decarboxylation of 12 likely occurs after it is incorporated into the polyketide biosynthetic pathway, probably as a thioester derivative with CoA or ACP. Furthermore, epimerization at the C-3 position of 12 most likely occurs during biosynthesis. The other carbons of the aglycon of vicenistatin, called vicenilactam (14), are derived from the typical polyketide pathway with malonyl-CoA and methylmalonyl-CoA, as evidenced by feeding experiments with 13C-labeled acetate and propionate [20].

Incorporation studies to investigate the origins of vicenistatin
The biosynthetic gene cluster (BGC) of vicenistatin was identified using probes targeting NDP-glucose 4,6-dehydratase and NDP-4-keto-6-deoxyglucose 2,3-dehydratase genes [24]. The vicenistatin BGC comprises typical type I PKS genes, deoxyaminosugar biosynthetic enzyme genes, a glutamate mutase gene and putative enzyme genes responsible for incorporating (2S,3S)-3-MeAsp (12) into the polyketide pathway.
Functional analysis of the candidate enzymes revealed that an adenylation enzyme, VinN, specifically recognizes 3-MeAsp (12), but not 3AIB (6), and binds it with a standalone ACP, VinL (15), in the presence of ATP to form 3-MeAsp-VinL (16) (Fig. 4) [25]. A PLP-dependent enzyme, VinO, then catalyzes the decarboxylation of 3-MeAsp-VinL (16) to produce 3AIB-VinL (17) [25]. Therefore, it is clear that thioester formation of 3-MeAsp is required before decarboxylation. VinO may not only recognize the 3-MeAsp moiety, but also the ACP moiety VinL. Additionally, epimerization at C-3 presumably occurs during decarboxylation by VinO, yielding (R)-3AIB-VinL (17), although this has not been clarified. Another adenylation enzyme, VinM, is encoded in the vicenistatin BGC and was found to activate l-Ala, l-Ser, and Gly, but not 3-MeAsp. Moreover, it was discovered that 3AIB-VinL (17) serves as an acceptor, generating l-Ala-3AIB-VinL (18) [25]. VinM selectively recognizes 3AIB-VinL (17), distinguishing it from VinL (15) and 3-MeAsp-VinL (16). The generated l-Ala-3AIB-VinL (18) is recognized by an acyltransferase, VinK, and the dipeptide group is transferred to the N-terminal ACP domain of the loading module (Ld-ACP) of PKS VinP1 (Fig. 4) [25].

Incorporation mechanism of 3-MeAsp into the polyketide synthase (PKS) in vicenistatin biosynthesis
Vicenistatin polyketide synthase (PKS) comprises four typical type I PKSs, VinP1, VinP2, VinP3, and VinP4, and is likely responsible for vicenilactam formation (Fig. 5) [24]. The PKS domain structure corresponds well to the chemical structure of vicenilactam (14), including specificity for the extender unit and the degree of reduction at the β-position. However, the mechanism of the C9–C10 double-bond formation remains unclear. The DH domain in module 5 of VinP5 is presumably involved in a unique dehydration process that generates the corresponding trisubstituted olefin. Ultimately, the C-terminal thioesterase domain of the VinP4 PKS is responsible for macrolactamization, resulting in the formation of vicenilactam (14) [26, 27].

PKS for vicenilactam in vicenistatin biosynthesis
Prior to macrolactamization, an amidohydrolase, VinJ, removes the terminal alanyl moiety from the elongated polyketide chain (Fig. 4) [25, 28]. VinJ appears to recognize the elongated polyketide chain with l-Ala at its terminus, although the exact substrate of recognition and timing of which VinJ recognizes, remain unclear. We propose that the attachment of l-Ala to form l-Ala-3AIB-VinL (18) is likely a protective step, blocking the nucleophilic amino group in 3AIB during polyketide elongation (the thermodynamically preferable six-membered lactam can form after the first polyketide extension step if the β-amino group is free; however, the terminal amino group of l-Ala in the dipeptide intermediate cannot access the thioester moiety on the ACP domain of PKS due to the rigid conformation of the amide bond). Thus, the removal of l-Ala from the elongated polyketide intermediate appears to be a deprotection step, generating the nucleophilic amino group derived from 3AIB. This protection–deprotection logic resembles the methodology employed in organic synthesis. Nature seems to favor this type of methodology for the efficient biosynthesis of natural products; for example, amidohydrolase-mediated cleavage reactions are employed in the selective biosynthesis of other dead-end natural products, such as desertomycin [29] and butirosin [30].
A glycosyltransferase, VinC, is responsible for attaching the unique deoxyaminosugar, vicenisamine, to complete the biosynthesis [24]. The substrate, dTDP-vicenisamine (19), is biosynthesized from d-glucose 1-phosphate (20) through the concerted action of the following enzymes: dTDP-glucose synthase VinA, dTDP-glucose 4,6-dehydratase VinB, dTDP-4-keto-6-deoxyglucose 2,3-dehydratase VinD, dTDP-3,4-diketo-2,6-dideoxyglucose 3-ketoreductase VinE, aminotransferase VinF, and N-methyltransferase VinG (Fig. 6) [19, 24]. Knocking out the N-methyltransferase gene resulted in the production of 4’-N-demethylvicenistatin, which exhibited potent antibacterial activities [19].

Post-PKS modification in vicenistatin biosynthesis
Incorporation of β-amino acids into the polyketide synthase: a common mechanism
A series of biosynthetic studies on vicenistatin revealed a common logic regarding the mechanism of β-amino acid starter unit incorporation into the polyketide pathway (Fig. 4) [31]. First, a β-amino acid-selective adenylation enzyme activates a pathway-specific β-amino acid and ligates it to a standalone ACP, yielding β-aminoacyl-ACP. Second, another adenylation enzyme activates l-Ala, l-Ser, or Gly, and catalyzes an amide bond-forming reaction with β-aminoacyl-ACP to give dipeptidyl-ACP, presumably for protection of the β-amino group. Third, dipeptidyl-ACP is recognized by a dipeptidyltransferase and transferred to the loading ACP domain at the start of the initial PKS module. Finally, the terminal aminoacyl group is removed by an amidohydrolase prior to macrolactam formation. We have identified the BGCs for incednine (2) [32], cremimycin (3) [33], hitachimycin (4) [34], and fluvircin B2 (5) [35]; these studies indicated that homologous enzymes are encoded in all BGCs. Thus, this biosynthetic logic is commonly utilized in the biosynthesis of macrolactam antibiotics with β-amino acid starter units. The existence of five homologous enzymes, including a standalone ACP, highlights the potential for BGC identification corresponding to other macrolactam antibiotics that incorporate β-amino acid starter units [9, 10]. An important detail in the biosynthesis of vicenistatin and fluvircin B2 is that a decarboxylation reaction (removal of the β-carboxy group from β-aminoacyl-ACP) must occur before the second aminoacylation to give dipeptidyl-ACP.
Among the five common homologous biosynthetic enzymes, the β-amino acid-selective adenylation enzymes are unique to each biosynthetic pathway [31, 36]. β-Amino acid-selective adenylation enzymes can be classified based on their substrate specificity. These enzymes can recognize β-amino acids with a carboxy group at the β-position, such as 3-MeAsp (12) by VinN [24] /VtlN [37] and Asp (21) by FlvN [35], a methyl group at the β-position (3-aminobutylic acid, 3ABA, 7) by IdnL1 [32]/LobL [38]/MmlL [39], a medium-chain alkyl group at the β-position, such as 3-aminononanoic acid (3ANA, 8) by CmiS6 [33]/BecJ [40]/HerJ [41, 42]/MlaJ [43], and a phenyl group at the β-position (β-phenylalanine, 9) by HitB [34] (Fig. 7). Crystal structure analysis of VinN [44], CmiS6 [45], IdnL1 [45], and HitB [46] revealed the mechanism of selective recognition of β-amino acids. It is now possible to predict the substrate specificity of β-amino acid-selective adenylation enzymes by comparing their amino acid sequences, particularly for the enzymes involved in this class of macrolactam biosynthesis.

Non-ribosomal (amino acid specificity-conferring) codes for representative β-amino acid selective adenylation enzymes. Unique amino acid residues conserved in each family of enzymes are highlighted in color
It is also intriguing how these β-amino acid-selective adenylation enzymes recognize their cognate standalone ACPs (Fig. 8). To elucidate the mechanism by which β-amino acid-selective adenylation enzymes recognize standalone ACPs, we constructed a cross-linked complex using HitB and HitD in hitachimycin biosynthesis [47]. In this system, the original cysteamine moiety in pantetheine is replaced with 1,2-ethylenediamine and further acylated with α-bromoacetic acid, to yield the α-bromoacetamide-containing pantetheine mimic C2Br (25). This analog provides an electrophilic functional group for the cross-linking reaction. C2Br (25) is enzymatically converted to the corresponding acylated CoA mimic by the CoA biosynthetic enzymes CoaA, CoaD, and CoaE [48]. A promiscuous phosphopantetheinyl transferase, Sfp [49], is then used to transfer the C2Br mimic to the apo form of a standalone ACP to give C2Br-ACP (26) as crypto-ACP. Next, we mutated an aspartate residue, which is conserved among adenylation enzymes (shown here as A domain), to cysteine. In the wild-type enzyme, the aspartate residue interacts with the amino group of the β-amino acid substrate. When the ACP is correctly recognized by the adenylation enzyme, the thiol group of the cysteine residue functions as a nucleophile, reacting with the α-bromoacetyl group of crypto-ACP to form a cross-linked complex. The crystal structure of the HitB–HitD cross-linked complex revealed the interface between these proteins, illustrating their favorable interaction. This unique protein–protein interaction (PPI) appears to be a key factor for the selective transfer of the β-amino acid to the cognate ACP to produce β-aminoacyl-ACP in selective biosynthesis.

Cross-linking reaction of adenylation enzyme (A domain) and crypto-ACP
It is also intriguing how VinM selectively recognizes 3AIB-VinL (17) by distinguishing it from the holo form of VinL (15) and 3-MeAsp-VinL (16). To address the importance of the length of the β-aminoacyl moiety in 3AIB-VinL (17), we prepared longer versions of the α-bromoacetamide pantetheine mimic by replacing the 1,2-ethylenediamine moiety with 1,3-propanediamine, 1,4-propanediamine, or 1,6-hexanediamine, yielding C3Br (27), C4Br (28), and C6Br (29) (Fig. 8) [50]. When we used C6Br-crypto-VinL for the cross-linking reaction with the VinM cysteine mutant, the cross-linked complex was efficiently produced. The crystal structure of the VinM–VinL complex with C6Br (29) illustrated the PPI between VinM and VinL. Interestingly, the pantoate moiety of C6Br is bent, and the dimethyl group interacts with a tyrosine residue that is conserved among the VinM family of adenylation enzymes. Therefore, VinM appears to recognize the pantoate moiety in 3AIB-VinL (17) and adjusts the location of the nucleophilic β-amino group for the amide bond-forming reaction to give l-Ala-3AIB-VinL (18). This result indicates that the phosphopantetheinyl group is not only a linker that connects the ACP and the acyl group, but is also selectively recognized by enzymes that transfer acyl groups.
The cross-linked complex between the dipeptidyltransferase VinK and VinL, using bismaleimide and C2Br (25) as cross-linkers, revealed an appropriate PPI between these proteins [51, 52]. Furthermore, VinK has a large cavity at its active site, which accommodates the dipeptide moiety of l-Ala-3AIB-VinL (18). Therefore, VinK seems to distinguish l-Ala-3AIB-VinL (18) from 3-MeAsp-VinL (16) and 3AIB-VinL (17) and selectively transfers the dipeptidyl moiety to the loading ACP domain. Overall, these studies allowed us to elucidate the detailed mechanism of the biosynthesis of vicenistatin.
Biosynthesis of β-amino acids in macrolactam antibiotics: unique mechanisms in each pathway
β-Amino acids, being non-proteinogenic amino acids, are biosynthesized in each macrolactam producer strain. In vicenistatin biosynthesis, (2S,3S)-3-MeAsp (12) appears to be biosynthesized from l-glutamic acid (11) through the action of AdoCbl and a PLP-dependent glutamate mutase [23], comprising the E subunit VinI and the S subunit VinH (Fig. 9) [24]. Disruption of the vinI gene abolishes vicenistatin production, which confirms the involvement of the glutamate mutase E subunit VinI in the formation of (2S,3S)-3-MeAsp (12). However, mutants with the disrupted vinI gene produced 18-desmethylvicesistain, supposedly biosynthesized from l-Asp (21) [53]. Since VinN recognizes l-Asp (21) with low efficiency [25], downstream enzymes may accommodate the l-Asp-derived intermediates to biosynthesize 18-desmethylvicenistatin.

Biosynthesis of β-amino acids
In incednine biosynthesis, (S)-3-aminobutyric acid (3ABA, 7) is biosynthesized from l-glutamic acid (11), as determined by feeding experiments [54, 55]. Interestingly, incorporation studies revealed that β-glutamate (30) is the biosynthetic intermediate. A PLP-dependent decarboxylase, IdnL3, was found to recognize β-glutamate (30) and catalyze decarboxylation to give (S)-3ABA (7) (Fig. 9) [32]. In the incednine BGC, a radical S-adenosyl-l-methionine-dependent enzyme, IdnL4, is encoded and is likely responsible for rearranging the α-amino group of l-Glu (11) to the β-position to give β-glutamate (30) (functioning as a PLP-dependent glutamate 2,3-aminomutase [56]). We disrupted the idnL4 gene to confirm that idnL4 is involved in the biosynthesis of 3ABA (7) [57], although the expected enzymatic activity of IdnL4 has not been observed in vitro. Furthermore, we demonstrated mutasynthesis of incednine analogs using the ΔidnL4 strain with 3-aminopentanoic acid (3APA) as an alternative substrate for IdnL1. Since IdnL1 shows strict substrate specificity against β-amino acid substrates, only 3ABA (7) and 3APA were incorporated into the pathway to produce natural incednine and 28-methylincednine [57].
In hitachimycin biosynthesis, (S)-β-Phe (9) appears to be biosynthesized from l-α-phenylalanine by the action of phenylalanine aminomutase HitA (Fig. 9). The enzymatic activity of HitA has been confirmed [34]. Additionally, the hitA gene was disrupted to verify its involvement in the biosynthesis of hitachimycin [34]. Supplementation of (S)-β-Phe (9) into the ΔhitA strain was required to recover hitachimycin production. Interestingly, HitB shows tolerant substrate specificity, recognizing several (S)-β-Phe derivatives [46]. Thus, mutasynthesis of hitachimycin analogs was conducted with the ΔhitA strain [46] (Fig. 10). In mutasynthesis, it was clarified that HitB functions as a gatekeeper in the biosynthesis, as most (S)-β-Phe derivatives recognized by HitB were incorporated into the pathway, yielding the corresponding hitachimycin analogs.

Mutasynthesis of hitachimycin analogs
In cremimycin biosynthesis, 3-aminononanoic acid (3ANA, 8) is biosynthesized via the polyketide pathway, as determined by feeding experiments [58]. In the cremimycin BGC, two sets of type I PKSs, one for 3ANA biosynthesis and one for macrolactam biosynthesis, are encoded [33]. The PKS for 3ANA biosynthesis consists of CmiP4, CmiP3, and CmiP2. Interestingly, a DH domain is split into CmiP3 and CmiP2 (Fig. 11). Furthermore, there is no TE domain in the PKS to release the polyketide chain for 3ANA (8) biosynthesis. A split DH domain was artificially connected to verify the dehydration activity. As a result, the split DH domain catalyzed the dehydration of (R)-2-hydroxynonanoyl-N-acetylcysteamine (NAC) thioester to yield (E)-2-nonenoate [59]. Additionally, a unique dual-functional thioesterase, CmiS1, was found to catalyze the Michael addition of glycine to the β-position of (E)-2-nonenoate and the subsequent hydrolysis of the thioester to give N-carboxymethyl-3-aminononanoic acid (31) [33]. Crystal structure analysis of a CmiS1 homolog SAV606 revealed a unique substrate recognition and reaction mechanism [60]. N-Carboxymethyl-3-aminononanoic acid (31) is then recognized by a flavin adenine dinucleotide (FAD)-dependent dehydrogenase CmiS2 and converted to (R)-3ANA (8), a unique β-amino acid macrolactam starter unit in cremimycin biosynthesis [33]. CmiS2 seems to catalyze dehydrogenation, to generate an imine intermediate, and subsequent hydrolysis, to afford glyoxylic acid and (R)-3ANA (8) [61]. Interestingly, CmiS1/SAV606-type thioesterase is involved in the biosynthesis of isonitrile-containing natural products to produce N-carboxymethyl-3-amino-fatty acid [62], which is converted to a fatty acid with an isonitrile group at the β-position by the action of α-ketoglutarate-dependent non-heme iron oxygenase [63,64,65,66].

Biosynthesis of (R)-3-aminononanoic acid in cremimycin biosynthesis
Overall, unique β-amino acids are biosynthesized by pathway-specific enzymes in each macrolactam producer strain to supply the appropriate starter units.
Unique mechanisms of post-PKS modification
In vicenistatin biosynthesis, the aglycone vicenilactam (14) undergoes glycosylation with dTDP-vicenisamine (19), catalyzed by the glycosyltransferase VinC, to complete the biosynthesis (Fig. 6) [24]. This post-PKS modification is crucial for the biological activity of vicenistatin. The unique glycosyl donor dTDP-vicenisamine (19) is biosynthesized by unique deoxyaminosugar biosynthetic enzymes, as described earlier [19]. A similar scenario is also seen in fluvirucin B2 biosynthesis (Figs. 12 and 13) [35]. The glycosylation of the aglycon of fluvirucin B2 with dTDP-l-mycosamine (32) by FlvS5 is essential to complete the biosynthesis. dTDP-l-mycosamine (32) is likely biosynthesized by a set of NDP-deoxyaminosugar biosynthetic enzymes: dTDP-glucose synthase FlvS1, dTDP-glucose 4,6-dehydratase FivS2, dTDP-4-keto-6-deoxyglucose 3,5-epimerase FlvS3, and dTDP-sugar aminotransferase FlvS4 (Fig. 12). The unique combinations of aglycones and deoxyaminosugars are significant in the biosynthesis of specific macrolactam compounds with distinct biological activities.

Putative biosynthetic pathways of NDP-deoxy(amino)sugars

Putative glycosylation reaction in the biosynthesis of fluvirucin B2 and incednine
In incednine biosynthesis, the aglycone incednam (33) appears to undergo glycosylation with two unique sugars, N-methylxylosamine and N-demethylforosamine, presumably by glycosyltransferases IdnS4 and IdnS14, and possibly with the assistance of an auxiliary cytochrome P450-type protein, IdnO2 (Fig. 13) [32]. UDP-N-methylxylosamine (34) is likely biosynthesized from UDP-N-acetyl-d-glucosamine (GlcNAc, 35) through the action of UDP-GlcNAc 6’-dehydrogenase IdnS2, UDP-N-acetylxylosamine (XylNAc) synthase IdnS3, UDP-XylNAc deacetylase IdnS1, and N-methyltransferase IdnS5 (Fig. 12). dTDP-N-demethylforosamine (36) is presumably biosynthesized from d-glucose 1-phosphate (20) through the sequential action of the following enzymes: dTDP-glucose synthase IdnS7, dTDP-glucose 4,6-dehydratase IdnS6, dTDP-4-keto-6-deoxyglucose 2,3-dehydratase IdnS10, dTDP-3,4-diketo-2,6-dideoxyglucose 3-ketoreductase IdnS11, dTDP-4-keto-2,6-dideoxyglucose 3,4-dehydratase IdnS12, aminotransferase IdnS13, and N-methyltransferase IdnS8 (Fig. 12). Incednam (33) is unlikely to be the macrolactam that is initially formed by PKSs IdnP1, IdnP2, IdnP3, IdnP4, and IdnP5, as it contains a hydroxy group at the C-10 position. This hydroxy group is presumably introduced by a cytochrome P450 monooxygenase, IdnO1, during post-PKS modification, although the exact point at which it does in the enzymatic pathway remains unclear.
Cremimycin (3) and hitachimycin (4) share a common bicyclic polyketide skeleton, with the C-8 and C-12 positions presumably linked by post-PKS modification enzymes (Fig. 14). Comparison of the BGCs of cremimycin and hitachimycin suggests the involvement of five homologous enzymes (two nicotinamide adenine dinucleotide cofactor-dependent oxidoreductases (CmiM1/HitM1 and CmiM3/HitM4), a cytochrome P450 (CmiM4/HitM3), and two putative sugar phosphate isomerase/epimerase type enzymes (CmiM2/HitM2 and CmiM7/HitM5)) in the formation of the common bicyclic structure [33, 34]. Functional analysis of these enzymes will pave the way to understanding the unique mechanism of polyketide backbone modification.

Putative post-PKS modification in cremimycin and hitachimycin biosynthesis
In cremimycin biosynthesis, the glycosylation reaction likely occurs after the formation of the bicyclic structure; this is because the hydroxy group at C10 of cremimycin appears to be introduced during the modification of polyketide backbone, as determined by the domain structures of the PKSs. Glycosyltransferase CmiM5 is likely responsible for glycosylation with dTDP-digitoxose (37), which is presumably biosynthesized by the following enzymes: dTDP-glucose synthase CmiC7, dTDP-glucose 4,6-dehydratase CmiC6, dTDP-4-keto-6-deoxyglucose 2,3-dehydratase CmiC5, dTDP-3,4-diketo-2,6-dideoxyglucose 3-ketoreductase CmiC4, dTDP-4-keto-2,6-dideoxyglucose 3,5-epimerase CmiC3, and dTDP-4-keto-2,6-dideoxyhexose 4-ketoreductase CmiC2 (Fig. 12) [33]. The final step in the biosynthesis of cremimycin is likely O-methylation of the digitoxose moiety by methyltransferase CmiC1.
In hitachimycin biosynthesis, the C10-OH group is anticipated to be O-methylated by the methyltransferase HitM6 in the final step of post-PKS modification.
Future perspectives
The next challenge for the field is to build upon the accumulated knowledge of the biosynthetic mechanisms of β-amino acid macrolactam antibiotics to engineer designer macrolactam compounds. As mentioned earlier, we have successfully produced hitachimycin analogs through mutasynthesis with β-Phe derivatives. However, the scope of the derivatives was limited because the β-amino acid-selective adenylation enzyme HitB did not accept most o- and p-substituted β-Phe derivatives, with the exception of small fluorine-substituted analogs. To address this limitation, the active site of HitB must be engineered to accommodate o- and p-substituted β-Phe derivatives, which would enable the production of a diverse range of hitachimycin analogs. Consequently, the engineering of adenylation enzymes to broaden their substrate scope is a crucial next step.
Moreover, β-amino acid-selective-adenylation enzymes can be exchanged to alter the β-amino acid starter units, leading to the production of new macrolactam compounds. Given the unique PPI revealed by the HitB–HitD cross-linked complex, it is essential to confirm whether the exchanged β-amino acid-selective adenylation enzyme can interact with different ACPs to ligate non-native β-amino acids. To overcome potential challenges in PPIs, designing specific interactions between each adenylation enzyme and the ACP may be necessary to introduce unnatural β-amino acids into the pathway, facilitating the production of designer macrolactams. The engineering of type I PKSs, including the exchange, introduction, and deletion of domains, can also be applied to the biosynthetic machinery of macrolactam antibiotics.
This biosynthetic journey, starting from the discovery of vicenistatin, has spanned three decades. Challenges still lie ahead as we strive to create novel macrolactam compounds with improved biological activities.
References
Kudo F, Miyanaga A, Eguchi T. Biosynthesis of natural products containing β-amino acids. Nat Prod Rep. 2014;31:1056–73.
Shindo K, Kamishohara M, Odagawa A, Matsuoka M, Kawai H. Vicenistatin, a novel 20-membered macrocyclic lactam antitumor antibiotic. J Antibiot. 1993;46:1076–81.
Futamura Y, Sawa R, Umezawa Y, Igarashi M, Nakamura H, Hasegawa K, et al. Discovery of incednine as a potent modulator of the anti-apoptotic function of Bcl-xL from microbial origin. J Am Chem Soc. 2008;130:1822–3.
Igarashi M, Tsuchida T, Kinoshita N, Kamijima M, Sawa R, Sawa T, et al. Cremimycin, a novel 19-membered macrocyclic lactam antibiotic, from Streptomyces sp. J Antibiot. 1998;51:123–9.
Umezawa I, Takeshima H, Komiyama K, Koh Y, Yamamoto H, Kawaguchi M. A new antitumor antibiotic, stubomycin. J Antibiot. 1981;34:259–65.
Ōmura S, Nakagawa A, Shibata K, Sano H. The structure of hitachimycin, a novel macrocyclic lactam involving beta -phenylalanine. Tetrahedron Lett. 1982;23:4713–6.
Naruse N, Tsuno T, Sawada Y, Konishi M, Oki T. Fluvirucins A1, A2, B1, B2, B3, B4 and B5, new antibiotics active against influenza A virus. II. Structure determination. J Antibiot. 1991;44:741–55.
Hegde VR, Patel MG, Gullo VP, Puar MS. Sch 38518 and Sch 39185: two novel macrolactam antifungals. Chem Commun 1991; 810–2 https://pubs.rsc.org/en/content/articlelanding/1991/c3/c39910000810.
Shin YH, Im JH, Kang I, Kim E, Jang SC, Cho E, et al. Genomic and spectroscopic signature-based discovery of natural macrolactams. J Am Chem Soc. 2023;145:1886–96.
Seibel E, Um S, Dayras M, Bodawatta KH, de Kruijff M, Jønsson KA, et al. Genome mining for macrolactam-encoding gene clusters allowed for the network-guided isolation of β-amino acid-containing cyclic derivatives and heterologous production of ciromicin A. Commu Chem. 2023;6:257.
Alvarez R, de Lera AR. Natural polyenic macrolactams and polycyclic derivatives generated by transannular pericyclic reactions: optimized biogenesis challenging chemical synthesis. Nat Prod Rep. 2021;38:1136–220.
Hertweck C. The biosynthetic logic of polyketide diversity. Angew Chem Int Ed. 2009;48:4688–716.
Staunton J, Weissman KJ. Polyketide biosynthesis: a millennium review. Nat Prod Rep. 2001;18:380–416.
Robbins T, Liu YC, Cane DE, Khosla C. Structure and mechanism of assembly line polyketide synthases. Curr Opin Struct Biol. 2016;41:10–18.
Chan YA, Podevels AM, Kevany BM, Thomas MG. Biosynthesis of polyketide synthase extender units. Nat Prod Rep. 2009;26:90–114.
Moore BS, Hertweck C. Biosynthesis and attachment of novel bacterial polyketide synthase starter units. Nat Prod Rep. 2002;19:70–99.
Olano C, Méndez C, Salas JA. Post-PKS tailoring steps in natural product-producing actinomycetes from the perspective of combinatorial biosynthesis. Nat Prod Rep. 2010;27:571–616.
Matsushima Y, Nakayama T, Fujita M, Bhandari R, Eguchi T, Shindo K, et al. Isolation and structure elucidation of vicenistatin M, and importance of the vicenisamine aminosugar for exerting cytotoxicity of vicenistatin. J Antibiot. 2001;54:211–9.
Liang Z, Li J, Ling C, Xu R, Yi X, Ju J, et al. Characterization of the aminosugar biosynthetic and regulatory genes of vicenistatin in Monodonata labio-associated Streptomyces parvus SCSIO Mla-L010. J Nat Prod. 2022;85:256–63.
Otsuka M, Eguchi T, Shindo K, Kakinuma K. Non-fatty acyl polyketide starter in the biosynthesis of vicenistatin, an antitumor macrolactam antibiotic. Tetrahedron Lett. 1998;39:3185–8.
Otsuka M, Fujita M, Matsushima Y, Eguchi T, Shindo K, Kakinuma K. Biosynthetic pathway of macrolactam polyketide glycoside antitumor antibiotic vicenistatins. Tetrahedron. 2000;56:8281–6.
Nishida H, Eguchi T, Kakinuma K. Amino acid starter unit in the biosynthesis of macrolactam polyketide antitumor antibiotic vicenistatin. Tetrahedron. 2001;57:8237–42.
Marsh ENG. Coenzyme-B12-dependent glutamate mutase. Bioorg Chem. 2000;28:176–89.
Ogasawara Y, Katayama K, Minami A, Otsuka M, Eguchi T, Kakinuma K. Cloning, sequencing, and functional analysis of the biosynthetic gene cluster of macrolactam antibiotic vicenistatin in Streptomyces halstedii. Chem Biol. 2004;11:79–86.
Shinohara Y, Kudo F, Eguchi T. A natural protecting group strategy to carry an amino acid starter unit in the biosynthesis of macrolactam polyketide antibiotics. J Am Chem Soc. 2011;133:18134–7.
Kudo F, Kitayama T, Kakinuma K, Eguchi T. Macrolactam formation catalyzed by the thioesterase domain of vicenistatin polyketide synthase. Tetrahedron Lett. 2006;47:1529–32.
Kudo F, Asou Y, Watanabe M, Kitayama T, Eguchi T. Potent oligomerization and macrocyclization activity of the thioesterase domain of vicenistatin polyketide synthase. Synlett. 2012;23:1843–6.
Shinohara Y, Miyanaga A, Kudo F, Eguchi T. The crystal structure of the amidohydrolase VinJ shows a unique hydrophobic tunnel for its interaction with polyketide substrates. FEBS Lett. 2014;588:995–1000.
Hong H, Samborskyy M, Lindner F, Leadlay PF. An amidinohydrolase provides the missing link in the biosynthesis of amino marginolactone antibiotics. Angew Chem Int Ed. 2016;55:1118–23.
Li Y, Llewellyn NM, Giri R, Huang F, Spencer JB. Biosynthesis of the unique amino acid side chain of butirosin: Possible protective-group chemistry in an acyl carrier protein-mediated pathway. Chem Biol. 2005;12:665–75.
Miyanaga A, Kudo F, Eguchi T. Mechanisms of β-amino acid incorporation in polyketide macrolactam biosynthesis. Curr Opin Chem Biol. 2016;35:58–64.
Takaishi M, Kudo F, Eguchi T. Identification of the incednine biosynthetic gene cluster: characterization of novel β-glutamate-β-decarboxylase IdnL3. J Antibiot. 2013;66:691–9.
Amagai K, Takaku R, Kudo F, Eguchi T. A unique amino transfer mechanism for constructing the β-amino fatty acid starter unit in the biosynthesis of the macrolactam antibiotic cremimycin. ChemBioChem. 2013;14:1998–2006.
Kudo F, Kawamura K, Uchino A, Miyanaga A, Numakura M, Takayanagi R, et al. Genome mining of the hitachimycin biosynthetic gene cluster: involvement of a phenylalanine-2,3-aminomutase in biosynthesis. ChemBioChem. 2015;16:909–14.
Miyanaga A, Hayakawa Y, Numakura M, Hashimoto J, Teruya K, Hirano T, et al. Identification of the Fluvirucin B2 (Sch 38518) Biosynthetic gene cluster from Actinomadura fulva subsp. indica ATCC 53714: substrate specificity of the β-amino acid selective adenylating enzyme FlvN. Biosci Biotechnol Biochem. 2016;80:935–41.
Kudo F, Miyanaga A, Eguchi T. Structural basis of the nonribosomal codes for nonproteinogenic amino acid selective adenylation enzymes in the biosynthesis of natural products. J Ind Microbiol Biotechnol. 2019;46:515–36.
Nogawa T, Terai A, Amagai K, Hashimoto J, Futamura Y, Okano A, et al. Heterologous expression of the biosynthetic gene cluster for verticilactam and identification of analogues. J Nat Prod. 2020;83:3598–605.
Schulze CJ, Donia MS, Siqueira-Neto JL, Ray D, Raskatov JA, Green RE, et al. Genome-directed lead discovery: Biosynthesis, structure elucidation, and biological evaluation of two families of polyene macrolactams against Trypanosoma brucei. ACS Chem Biol. 2015;10:2373–81.
Skellam EJ, Stewart AK, Strangman WK, Wright JLC. Identification of micromonolactam, a new polyene macrocyclic lactam from two marine Micromonospora strains using chemical and molecular methods: clarification of the biosynthetic pathway from a glutamate starter unit. J Antibiot. 2013;66:431–41.
Jørgensen H, Degnes KF, Sletta H, Fjærvik E, Dikiy A, Herfindal L, et al. Biosynthesis of macrolactam BE-14106 involves two distinct PKS systems and amino acid processing enzymes for generation of the aminoacyl starter unit. Chem Biol. 2009;16:1109–21.
Zhu Y, Zhang W, Chen Y, Yuan C, Zhang H, Zhang G, et al. Characterization of heronamide biosynthesis reveals a tailoring hydroxylase and indicates migrated double bonds. ChemBioChem. 2015;16:2086–93.
Booth TJ, Alt S, Capon RJ, Wilkinson B. Synchronous intramolecular cycloadditions of the polyene macrolactam polyketide heronamide C. Chem Commun. 2016;52:6383–6.
Jørgensen H, Degnes KF, Dikiy A, Fjærvik E, Klinkenberg G, Zotchev SB. Insights into the evolution of macrolactam biosynthesis through cloning and comparative analysis of the biosynthetic gene cluster for a novel macrocyclic lactam, ML-449. Appl Environ Microbiol. 2010;76:283–93.
Miyanaga A, Cieślak J, Shinohara Y, Kudo F, Eguchi T. The Crystal structure of the adenylation enzyme VinN reveals a unique β-amino acid recognition mechanism. J Biol Chem. 2014;289:31448–57.
Cieślak J, Miyanaga A, Takaku R, Takaishi M, Amagai K, Kudo F, et al. Biochemical characterization and structural insight into aliphatic β-amino acid adenylation enzymes IdnL1 and CmiS6. Proteins. 2017;85:1238–47.
Kudo F, Takahashi S, Miyanaga A, Nakazawa Y, Nishino K, Hayakawa Y, et al. Mutational biosynthesis of hitachimycin analogs controlled by the β-amino acid–selective adenylation enzyme HitB. ACS Chem Biol. 2021;16:539–47.
Miyanaga A, Kurihara S, Chisuga T, Kudo F, Eguchi T. Structural characterization of complex of adenylation domain and carrier protein by using pantetheine cross-linking probe. ACS Chem Biol. 2020;15:1808–12.
Miyanaga A, Kudo F, Eguchi T Cross-linking of the nonribosomal peptide synthetase adenylation domain with a carrier protein using a pantetheine-yype probe. In: Burkart M, Ishikawa F (eds). Non-Ribosomal Peptide Biosynthesis and Engineering: Methods and Protocols, 2023, pp 207–17.
Quadri LEN, Weinreb PH, Lei M, Nakano MM, Zuber P, Walsh CT. Characterization of Sfp, a Bacillus subtilis phosphopantetheinyl transferase for peptidyl carrier protein domains in peptide synthetases. Biochemistry. 1998;37:1585–95.
Miyanaga A, Nagata K, Nakajima J, Chisuga T, Kudo F, Eguchi T. Structural basis of amide-forming adenylation enzyme VinM in vicenistatin biosynthesis. ACS Chem Biol. 2023;18:2343–8.
Miyanaga A, Iwasawa S, Shinohara Y, Kudo F, Eguchi T. Structure-based analysis of the molecular interactions between acyltransferase and acyl carrier protein in vicenistatin biosynthesis. Proc Natl Acad Sci USA 2016;113:1802–7.
Miyanaga A, Ouchi R, Kudo F, Eguchi T. Complex structure of the acyltransferase VinK and the carrier protein VinL with a pantetheine cross-linking probe. Acta Crystallogr F. 2021;77:294–302.
Ogasawara Y, Kakinuma K, Eguchi T. Involvement of glutamate mutase in the biosynthesis of the unique starter unit of the macrolactam polyketide antibiotic vicenistatin. J Antibiot. 2005;58:468–72.
Takaishi M, Kudo F, Eguchi T. Biosynthetic pathway of 24-membered macrolactam glycoside incednine. Tetrahedron. 2008;64:6651–6.
Takaishi M, Kudo F, Eguchi T. A unique pathway for the 3-aminobutyrate starter unit from L-glutamate through β-glutamate during biosynthesis of the 24-membered Mmacrolactam antibiotic, Incednine. Org Lett. 2012;14:4591–3.
Ruzicka FJ, Frey PA. Glutamate 2,3-aminomutase: a new member of the radical SAM superfamily of enzymes. Biochim Biophys Acta. 2007;1774:286–96.
Miyanaga A, Takaku R, Takaishi M, Tashiro E, Kudo F, Eguchi T. Generation of incednine derivatives by mutasynthesis. J Antibiot. 2020;73:794–7.
Amagai K, Kudo F, Eguchi T. Biosynthetic pathway of macrolactam polyketide antibiotic cremimycin. Tetrahedron. 2011;67:8559–63.
Kawasaki D, Miyanaga A, Chisuga T, Kudo F, Eguchi T. Functional and structural analyses of the split-dehydratase domain in the biosynthesis of macrolactam polyketide cremimycin. Biochemistry. 2019;58:4799–803.
Chisuga T, Miyanaga A, Kudo F, Eguchi T. Structural analysis of the dual-function thioesterase SAV606 unravels the mechanism of Michael addition of glycine to an α,β-unsaturated thioester. J Biol Chem. 2017;292:10926–37.
Kawasaki D, Chisuga T, Miyanaga A, Kudo F, Eguchi T. Structural analysis of the glycine oxidase homologue CmiS2 reveals a unique substrate recognition mechanism for formation of a β-amino acid starter unit in cremimycin biosynthesis. Biochemistry. 2019;58:2706–9.
Harris NC, Sato M, Herman NA, Twigg F, Cai W, Liu J, et al. Biosynthesis of isonitrile lipopeptides by conserved nonribosomal peptide synthetase gene clusters in Actinobacteria. Proc Natl Acad Sci USA 2017;114:7025–30.
Harris NC, Born DA, Cai W, Huang Y, Martin J, Khalaf R, et al. Isonitrile formation by a non-heme iron(II)-dependent oxidase/decarboxylase. Angew Chem Int Ed. 2018;57:9707–10.
Chen TY, Chen J, Tang Y, Zhou J, Guo Y, Chang WC. Pathway from N-alkylglycine to alkylisonitrile catalyzed by iron(II) and 2-oxoglutarate-dependent oxygenases. Angew Chem Int Ed. 2020;59:7367–71.
Jonnalagadda R, Del Rio Flores A, Cai W, Mehmood R, Narayanamoorthy M, Ren C, et al. Biochemical and crystallographic investigations into isonitrile formation by a nonheme iron-dependent oxidase/decarboxylase. J Biol Chem. 2021;296:100231.
Del Rio Flores A, Kastner DW, Du Y, Narayanamoorthy M, Shen Y, Cai W, et al. Probing the mechanism of isonitrile formation by a non-heme iron(II)-dependent oxidase/decarboxylase. J Am Chem Soc. 2022;144:5893–901.
Acknowledgements
I would like to express my gratitude to the late Prof. Katsumi Kakinuma, Emeritus Prof. Tadashi Eguchi (Tokyo Institute of Technology), Prof. Akimasa Miyanaga (currently at the University of Tokyo), and all the students who contributed to the research conducted in the same laboratory at the Tokyo Institute of Technology. The research presented in this review was supported in part by the Ministry of Education, Culture, Sports, Science and Technology (MEXT) through a Grant-in-Aid for Scientific Research on Innovative Areas (23108509), Nagase Science Technology Foundation, Takeda Science Foundation, Uehara Memorial Foundation, and the Noda Institute for Scientific Research. I thank Mchem University of York student Rowan Gibbard for editing this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Fumitaka Kudo was awarded the Sumiki-Umezawa Memorial Award from the Japan Antibiotic Research Association in 2023. This review article is partly based on his award-winning research.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kudo, F. Biosynthesis of macrolactam antibiotics with β-amino acid polyketide starter units. J Antibiot 77, 486–498 (2024). https://doi.org/10.1038/s41429-024-00742-2
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41429-024-00742-2
