
Abstract
Fires alter the stability of organic matter and promote soil erosion which threatens the fundamental coupling of soil biogeochemical cycles. Yet, how soil biogeochemistry and its environmental drivers respond to fire remain virtually unknown globally. Here, we integrate experimental observations and random forest model, and reveal significant divergence in the responses of soil biogeochemical attributes to fire, including soil carbon (C), nitrogen (N), and phosphorus (P) contents worldwide. Fire generally decreases soil C, has non-significant impacts on total N, while it increases the contents of inorganic N and P, with some effects persisting for decades. The impacts of fire are most strongly negative in cold climates, conifer forests, and under wildfires with high intensity and frequency. Our work provides evidence that fire decouples soil biogeochemistry globally and helps to identify high-priority ecosystems where critical components of soil biogeochemistry are especially unbalanced by fire, which is fundamental for the management of ecosystems in a world subjected to more severe, recurrent, and further-reaching wildfires.
Similar content being viewed by others


Introduction
Fires are pervasive events naturally affecting ecosystem development and succession1, and are known to alter multiple aspects of biodiversity and ecosystem functioning worldwide2,3. While natural fires can benefit ecosystems by promoting plant diversity (e.g., reducing competitive exclusion), plant germination, and ecosystem rejuvenation, the rates and severity of anthropogenic fires also represent a serious threat to ecosystem health and human well-being, as they become more severe and cover a greater area on Earth1,2. Fire effects on ecosystem services are expected to be further amplified by the interaction of land-use changes, fuel accumulation, and warming- and drought-associated climatic changes despite human interventions that attempt to manage and prevent fire3,4. As such, more severe and frequent fires are expected to occur over the next decades with still poorly described consequences for terrestrial ecosystems, and the multiple ecosystem services they provide to humankind. We know that carbon (C), nitrogen (N), and phosphorus (P) are closely coupled in natural ecosystems5 (see Supplementary Fig. S1 for an example of this). However, fires can reduce plant biomass, alter organic matter pools, and promote soil erosion2,6, which may cause fundamental changes in biogeochemical coupling of C, N, and P (hereafter decoupling) and, as a consequence, disrupt ecosystem services such as plant production7,8. More importantly, we may have underestimated future responses of ecosystems to fire, because most previous studies focused on fire impacts on soil C and N, while ignoring how soil P responds to and is coupled with responses of soil C and N. Given the foundational importance of soil biogeochemistry for ecosystem sustainability, it is essential to explore the global response pattern of soil biogeochemistry to fire. This knowledge is critical for predicting the future of terrestrial ecosystems and enhance management practices under global change.
Importantly, we know that C, N, and P elements have contrasting origins and availabilities in soil. While C and N are biologically fixed from the atmosphere and often accumulate during ecosystem development, P is derived from heat-induced mineralisation and ash deposition and P availability declines over time during ecosystem development, especially in forests4,9. However, biological and geochemical controls of nutrient cycling are likely to change after fire via declines in plant biomass, shifts in litter input content, and increased soil erosion on the scale of days to decades4,10,11. Therefore, fires, especially frequent and severe wildfires, could have critical consequences for the coupling of soil biogeochemical cycles globally. We predict that globally, fire leads to reductions in the amount of soil C, and increases in the content of mineral elements such as P that are associated with vegetative ash deposition. Despite the importance of biogeochemical cycling for ecosystem functioning and services, no study has evaluated how increases in fire severity and frequency simultaneously impact the contents of soil C, N, and P globally. More importantly, previous meta-analyses investigating fire impacts on soil biogeochemistry showed inconsistent results due to the highly variable number of observations12,13,14,15, which may cause great challenge in providing a clear picture for fire effects on soil biogeochemistry. A comprehensive assessment of the effects of fire on soil biogeochemistry is thus critical to reducing the uncertainty associated with projections of the future of terrestrial ecosystems under global change.
Climate is the major global driver of plant productivity, and a fundamental regulator of soil biogeochemistry and soil microbial biomass16,17,18. Climate also plays a critical role in shifting fire regimes (e.g., severity, frequency), with potentially devastating consequences where drier ecosystems and those accumulating considerable fuel loads are more likely to suffer more severe and frequent fires3,17,19. In contrast, pyrogenic ecosystems are experiencing problems related to climate change and human intervention, where historically-important fires have been halted, thus altering previously stable soil biotic and abiotic conditions20. Yet, despite the interest surrounding fire impacts, most global studies have ignored the role of climate in shaping the responses of soil biogeochemistry to fires across biomes12,21. Similarly, we do not know how the main types of forests, such as those dominated by either angiosperms, conifers, and different mycorrhizal types, regulate the responses of soil biogeochemistry to fires22. Thus, although the fire impacts on certain soil biogeochemical attributes such as soil respiration and soil P have been well documented23,24,25, we still lack a comprehensive investigation of how fire influences soil biochemical coupling of C, N, and P across contrasting climates and plant functional types worldwide.
Here, we conducted a global synthesis with 5460 observations (Fig. 1) to investigate the influence of fire frequency, severity, and type on the balance between C, N, and P contents in topsoils (0–20 cm) across major global climates and plant functional types (e.g., conifers versus angiosperms, arbuscular versus ectomycorrhizal plants). Our global synthesis provides information on the responses of soil C, soil N, and soil P contents to fires, and also how plant biomass and soil microbial biomass as well as soil organic matter (SOM) decomposition respond to fires worldwide. We determined the responses of soil C, N, P, SOM decomposition, and soil microbial biomass to fire using log response ratios (lnRR) comparing the mean value of a particular variable subjected to fire in a given time (year) versus a control (unburned plots from paired reference ecosystems; see Methods for details). Specifically, we hypothesized that: (1) fire would disrupt the biogeochemical coupling between C, N, and P, as fire is expected to remove biologically-driven elements such as C and N while rejuvenating the bedrock and releasing chemically-driven elements such as P; and (2) fire would exhibit greater negative effects on soil biogeochemistry in cold climates and coniferous forests under wildfires with high intensity and frequency because those ecosystems are characterized by slower nutrient cycling and lower microbial activity, making them more vulnerable to disturbance from severe and frequent fires.

Global distribution of the selected experimental sites (A). Colors in the map represent the distribution of current Köppen climate regions. Estimates (±95% CI) of the log response ratio for soil carbon, soil nitrogen, soil phosphorus, soil microbial biomass, and SOM decomposition (B). Estimates (±95% CI) of the log response ratio for soil carbon, soil nitrogen, soil phosphorus, soil microbial biomass, and SOM decomposition under different levels of fire severity (C), fire frequency (D), and fire type (E). The dashed lines are drawn at LnRR = 0. Number values for each point indicate sample size. The error bars indicate the 95% confidence intervals (CI), and the symbol * indicates statistical significance. SOM soil organic matter.
Results and discussion
Our global synthesis provides evidence that, on average, fire significantly decouples soil C, N, and P contents across the globe (Fig. 1, Supplementary Figs. S6, S7). These decoupling impacts are especially strong under more severe and frequent fires, and after wildfire rather than prescribed fire (Fig. 1). Climate and plant functional type played critical roles in explaining the variability of responses of soil elements after fire, with soil elements being more responsive to fire in coniferous and ectomycorrhizal ecosystems compared with angiosperm and arbuscular mycorrhizal ecosystems (Figs. 2 and 3). Our results also indicate that soil C, N, and P contents were more sensitive to fire in cold climate regions compared with arid and tropical/temperate ecosystems (Figs. 2 and 4). Importantly, our analyses also reveal that the impacts of fires on soil biogeochemistry could last longer than half of a century, with important temporal variability for most elements after fire (Fig. 5). Together, these results suggest that fire decouples soil biogeochemistry globally, a response which is predicted to get even stronger with anthropogenic climate change, and may subsequently impact multiple ecosystem services such as plant productivity and C sequestration.

Estimates (±95% CI) of the log response ratio for soil carbon, soil nitrogen, soil phosphorus, microbial biomass, and SOM decomposition under different plant functional types (A), mycorrhizal type (B), and Köppen climate type (C). The dashed lines are drawn at LnRR = 0. Number values for each point indicate sample size. The error bars indicate the 95% confidence intervals (CI), and the symbol * indicates statistical significance. Ecological connectivity network among six soil biogeochemistry attributes in respond to fire (D). Each attribute in the network is a node and connections represent partial correlation coefficients between two variables after conditioning on all other variables. The links with blue and red color indicate positive and negative correlation, respectively. The thickness of the links indicates the partial correlation coefficient. Principal component analysis (PCA) showing the responses (lnRR) of multiple soil biogeochemical attributes to fire (E). SOM soil organic matter, AM Arbuscular mycorrhizal fungi, ECM Ectomycorrizal fungi, SOM soil organic matter.

Multiple ranking regression reveals the relative importance of the most important predictors of fire effects on soil carbon (A), soil nitrogen (B), soil phosphorus (C), soil microbial biomass (D), and SOM decomposition (E). The standardized regression coefficients of the models are shown for each predictor with associated 95% confidence intervals. *P < 0.05, **P < 0.01, ***P < 0.001. Bar graphs show the relative importance of each predictor group (represented by variables on the y-axis), expressed as the percentage of variance explained. Biotic and abiotic predictors of soil biogeochemistry include: climate, space, plant production, soil properties, fire regime and plant functional types. MAT mean annual temperature, MAP mean annual precipitation, GPP gross primary productivity, LAI leaf area index, NDVI normalized difference vegetation index, BD bulk density, SOM soil organic matter.

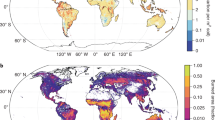
The predicted distribution of fire effects on soil C, soil N, soil P, soil microbial biomass, and SOM decomposition (A–E). Latitudinal distribution of soil biogeochemistry in response to fire (F). The R2 values were determined as predicted vs. observed soil C, soil N, soil P, soil microbial biomass, SOM decomposition were 0.45, 0.41, 0.62, 0.56, and 0.43, respectively. A map including information on model uncertainties is provided Supplementary Fig. S12. We also include model validation performance in Supplementary Fig. S13. Micro biomass, soil microbial biomass; SOM decom, soil organic matter decomposition.

Time since fire-induced change in soil carbon (A), soil nitrogen (B), soil phosphorus (C), soil microbial biomass (D), and SOM decomposition (E). The effects are expressed as the mean response ratios (±95% bootstrapped confidence intervals). Fire effects were considered statistically significant when confidence intervals do not overlap zero. Partial correlation analyses revealed that ecological and environmental factors are significantly correlated with fire-induced changes in soil biogeochemistry even when controlling for time since fire (F). The size of the circles presents correlation coefficients. The green and orange colors represent positive and negative correlations, respectively. MAT mean annual temperature, MAP mean annual precipitation, GPP gross primary productivity, LAI leaf area index, NDVI normalized difference vegetation index, BD bulk density, SOM soil organic matter. Asterisks indicate significance *P < 0.05; **P < 0.01; ***P < 0.001.
Fire decouples soil biogeochemistry worldwide
Consistent with our first hypothesis, our work provides empirical evidence that fires disrupt soil biogeochemical coupling of C, N, and P worldwide (Fig. 1). We showed that fire significantly decreased soil C, had no effect on total N, and increased inorganic N and P, suggesting high levels of nutrient mineralization after fires6,26 (Supplementary Fig. S6). We then cross-validated our results using three independent network analyses, spearman correlation and principal component analysis and still find that fire decouples soil C, N, and P contents (Fig. 2, Supplementary Fig. S8). Given the lack of protection provided by litter or ground vegetation post fire, enhanced soil degradation or erosion would impede substrate acquisition of microorganisms and thereby lower soil C15,27. The fire-induced decrease in plant biomass such as litter mass and root production, is also likely to diminish labile C inputs to microbes in bulk soil and the rhizosphere and its symbionts, leading to a decreased microbial diversity, microbial biomass, as well as the soil C pool (Fig. 1, Supplementary Fig. S8)2,27. Fire-induced reductions in soil C contents further alter the coupling of C with N or P, limiting the capacity of ecosystems to buffer ever-increasing atmospheric CO2 concentrations, and constraining plant and microbial productivity and diversity28,29,30. Our analyses further revealed the importance of fire regime, as the effects of fire on biochemical coupling are notably stronger under severe, frequent, and wildfire conditions compared with milder, rare, or prescribed fires, respectively (Fig. 1). Wildfires greatly altered the balance of biological versus abiotic controls in terrestrial ecosystems by reducing the amount of plant biomass and organic matter inputs and promoting soil erosion and bedrock rejuvenation, creating important regressions in ecosystem development. Fire can also stimulate mechanical rock weathering and ash incorporation into the soil, increasing soil P content through mineral deposit release or direct vegetative ash input, likely promoting mineralization processes10,31. However, a previous meta-analysis suggested that fire decreases both soil C and soil N of terrestrial ecosystems15. This difference may result from the fact that we included soil inorganic N as a part of the soil N component, which shows a positive response to fire due to enhanced soil N mineralization and nitrification32. Moreover, the dataset of the present study is larger than those in previous works, which provides more coverage of different environmental conditions and increases the power of statistical conclusions here. While ecosystem development often results in important soil C and N accumulation as well as P losses in topsoil, we show that fire can have the opposite effect. Together, these results indicate that fires disrupt soil biochemical coupling and may thus potentially undermine the future sustainability of terrestrial ecosystems under global change.
Fire-vulnerability of biogeochemistry peaks in cold and coniferous ecosystems
Our global synthesis further shows that vegetation types and climate are important modulators of the response of soil biogeochemistry to fire across the globe. Coniferous forests, ecosystems dominated by ectomycorrhizal associations, and cold regions exhibited the strongest response of soil biogeochemical metrics to fire, as we hypothesized (Fig. 2, Supplementary Fig. S8). We find that soil biogeochemistry, particularly C-related measures in conifer and ectomycorrhiza-dominated ecosystems are more susceptible to fire-related damage than angiosperm and arbuscular mycorrhizal ecosystems, exhibiting significantly more negative responses of soil C, soil microbial biomass and SOM decomposition. Conifers are well-known to promote high-severity fires as a consequence of more combustible content (e.g., resin), and with greater litter biomass inputs in response to fire33. Similarly, fire may cause greater loss of soil C, soil microbial biomass, and SOM decomposition in ecosystems dominated by ectomycorrhizal (ECM) plant species than in those dominated by plants associated with arbuscular mycorrhizal (AM) fungi, because ECM plants tend to produce more recalcitrant litter and store greater amounts of C in the soil22,34. These results suggest that fires have greater effects on soil biogeochemistry in colder regions than in warmer ones, although conifer forests and ECM plants can be widely found in subtropical areas, because MAT was positively correlated with soil biogeochemical attributes such as soil C content and SOM decomposition after fire (Fig. 2, Supplementary Fig. S9). The higher soil C stocks and soil microbial biomass in colder regions may act as a larger C reservoir35,36, and its combustion during fire might contribute to the more pronounced effects on soil metrics in colder climatic regions than those in tropical/temperate regions37. Importantly, our multiple regression models further confirmed the key role of climate and vegetation type in modulating soil biogeochemistry after fire (Fig. 3, Supplementary Fig. S10). Taken together, our results suggest that fire have stronger effects on soil biogeochemistry in cold and coniferous ecosystems, which should be considered in future land management to better achieve global climate change mitigation targets.
Mapping global fire impacts on soil biogeochemistry
To further identify the reported global divergence in the responses of soil biogeochemistry to fires, we used a random forest model to develop global maps of soil biogeochemical responses to fire (Fig. 4, Supplementary Fig. S11). Our maps account for uncertainty areas with lower sample sizes, wherein predictions are not reported (Supplementary Fig. S12). Model validation indicated that the random forest model performed well in predicting fire impacts on soil biogeochemistry (Supplementary Fig. S13). Fire-induced changes in soil C, soil N, and soil P contents revealed different latitudinal patterns, suggesting that the decoupling among biogeochemistry in response to fire is common across ecosystems. Our maps further indicate that fires have much stronger effects on soil biogeochemistry in cold regions, such as northern Europe and northeast China, than in warmer regions (e.g., sub-tropical and tropical), which is consistent with results reported above (Fig. 2C). Overall, our maps help to identify the climatic zones, ecosystem types, and locations on the planet where fire is likely to have a larger impact on soil biogeochemistry and may significantly disrupt global soil biochemical relationships.
Fire effects on soil biogeochemistry can endure for decades
We investigated the duration that soil biogeochemistry requires to recover to pre-fire or unburned, control levels and how these changes affect the long-term sustainability of ecosystems12,18,38. Our results reveal that the impacts of fires on soil biogeochemistry can last for decades (Fig. 5), which is consistent with findings from Dove et al. 39. Ecosystems after fire seem to transition to new alternative states with reduced levels of soil functionality that may have impacts across ecosystem features (e.g., on plant communities or C sequestration), and induce stabilizing feedbacks such as increasing albedo, reduction of biotic impacts on soil water storage or community composition changes39,40,41. More specifically, our analyses reveal that fire effects on soil C can last up to 55 years, while fire effects on soil N and P dynamics were greater in the short term (less than five years) than those at decadal scales (Fig. 5). The strongly decreased soil C in the short term may be closely associated with the declines in plant C inputs as well as the increasing biodegradation rate of labile C (Supplementary Fig. S8), while increased soil P could be a result of high-P vegetative ash inputs, increases in heat-induced microbial P mineralization, as well as increased biological weathering15,42. By contrast, fire-induced decreases in soil microbial biomass were negatively related with time since fire (Fig. 5), suggesting that soil microbes were more sensitive to fire disturbance than soil C, N, and P contents12. Additional cross-validation with effects of fire on the majority of biogeochemical properties showed similar patterns (Supplementary Fig. S14), providing further evidence that the fire-induced disruptions in global soil biochemical relationships of C, N, and P last for decades. Importantly, our results were consistent after controlling for changes in time since fire when analyzed with both partial correlations between ecological predictors (e.g., climate, soil properties) and soil biogeochemistry and the random forest model (Fig. 5F, Supplementary Fig. S15).
Fire management and implications
We here provide solid evidence that fire significantly disrupts soil biochemical coupling of C, N, and P contents of terrestrial ecosystems, and that fire regimes, vegetation types, as well as climates can modulate fire impacts globally. Prescribed fires have been applied as a land management strategy for centuries, including traditional practices of Australian aborigines or Native American tribes29,30, and our results suggest that the prudent use of prescribed fires can be considered as a practical alternative to active fire suppression in high-risk fire regions to better prevent larger losses of soil C stocks and thereby promote ecosystem sustainability compared with severe wildfires. More importantly, fire tends to exhibit greater impacts on biogeochemical properties in coniferous forests and ecosystems dominated by ectomycorrhizal associations in cold climates. These results highlight the fact that planting forests with mixtures of mycorrhizal types (including AM angiosperm trees) within cold climate regions have a greater potential to resist fire effects and therefore be more effective against changing climates. Together, more active collaboration among scientists, managers, policymakers, and local communities are necessary to promote the establishment of firebreaks, conduct active use of prescribed fires, as well as increasing broadleaf species at the time of stand regeneration, are critical for the long-term sustainability of terrestrial ecosystems.
We note that our global analysis is not exempt from limitations, which should be considered in future experiments to improve our current understanding of ecosystem sustainability under global change. Most fire experiments have been conducted in northern America and western Europe such as Spain and within forest ecosystems (Fig. 1). We also identified the Tropics and Southern Hemisphere regions such as Brazil and Africa as understudied regions, which may trigger uncertainty when assessing the effects of fire on soil biogeochemistry under the respective environmental conditions. These regions should be prioritized when developing new experimental work to provide a more comprehensive picture of soil biogeochemical cycling and facilitate ecosystem sustainability in a world subjected to more severe and recurrent wildfires. Although our study provides insights into the impacts of fire on soil biogeochemistry, we note that the experimental duration of fire effects in most studies was less than 10 years, and long-term responses may be biased towards high-severity biomes (e.g., high-severity wildfires in the Sierra Nevada forests)39, which may raise some uncertainty if we aim to precisely assess multiple ecosystem services that are associated with soil biogeochemistry under global change. Hence, there is a need to undertake long-term studies with more levels of fire severity and across more regions of the globe to better understand the effects of fire on soil biogeochemistry. Moreover, climate may interact with a diversity of other environmental factors such as soil type and vegetation recovery in regulating soil biogeochemistry and its linkages with plant biogeochemistry after fire, while the global patterns still remain virtually unknown43, which poses a challenge for ecosystem management to promote soil health and resilience. Together, our study opens the door to future studies that will deepen our understanding of soil biogeochemistry response to fire.
In summary, understanding the effects of fire on soil biogeochemistry of terrestrial ecosystems is crucial for sustainable ecosystem management11. Although fire is a powerful ecological and evolutionary force that shapes ecosystem services19, our global study provides solid evidence that fire significantly decouples soil biogeochemistry, with significant divergence in the responses of soil biogeochemical properties to fire, which can last for decades. These decoupling effects of fire are consistent across contrasting ecosystem types and climate regions, while the most significant fire impacts were found in coniferous forests, ecosystems dominated by ectomycorrhizal plants, and in cold regions. Most contemporary Earth systems models still consider coupled soil biogeochemical cycles or fail to account for differences in plant functional types and climatic drivers. Our study suggests that next generation Earth System Models should explicitly incorporate the decoupling effects of soil biogeochemistry after fire disturbance to better predict the feedbacks between multiple ecosystem services and global change. Moreover, our study highlights the importance of fire regime for soil biogeochemistry and reveals the decadal impacts of fires on these key ecosystem processes, which should serve as yet another notice that we must act now to mitigate climate change in order to minimize sustained fire impacts on soil processes and promote ecosystem sustainability and human health.
Methods
Global survey of soil biogeochemistry under fire
We searched for journal articles using the ISI Web of Science and the China National Knowledge Infrastructure database (CNKI) with the following key word combinations: (“fire” or “burn” or “wildfire” or “prescribed fire”) and (“soil”) and (“carbon” or “nitrogen” or “phosphorus” or “soil respiration” or “nutrient” or “carbon flux” or “soil organic matter” or “SOM decomposition” or “microbial biomass” or “PLFA” or “bacteria” or “fungi” or “biogeochemistry”) from 1950 to 2023. Soil biogeochemistry were grouped as soil carbon, soil nitrogen, soil phosphorus, soil microbial biomass (i.e., microbial biomass), and soil organic matter (SOM) decomposition (Supplementary Table S1). Each attribute was only included in one category. The following criteria were applied to stringently select fire studies that allow direct comparison of biochemical processes and avoid bias in publication selection: (i) fire experiments were conducted in the field; (ii) initial environmental conditions (e.g., soil type and ecosystem type) and climate in both the control and fire treatments were the same; (iii) at least one of the selected variables above (Supplementary Table S1) was recorded; (iv) the means of selected variables in both control and fire treatments could be extracted directly from associated datasets, tables, or digitized graphs; (v) the selected studies should be carried out with a paired-plot design, and the control treatments were established based on space (unburned plots compared to adjacent burned plots) or time (pre-fire compared to post-fire) comparisons.
Using these five stringent selection criteria and as indicated in the PRISMA diagram in Supplementary Fig. S2, a total of 471 studies with 5460 observations were selected (Fig. 1, Supplementary Fig. S3 and Supplementary Text S1) to evaluate how fire influences soil biogeochemical cycling globally. The majority of current studies were distributed in temperate climate, especially in western Europe and northern America, and most of them come from forest ecosystems (Fig. 1). Among these studies, 50 studies simultaneously include soil carbon (C), nitrogen (N) and phosphorus (P), 217 studies include both soil C and N, 58 studies include both soil C and P, and 78 studies include both soil N and P. Meanwhile, 1545 reported responses of soil C, 1595 reported soil N, 486 reported soil P, 809 reported microbial biomass, 730 reported SOM decomposition, and 295 reported plant biomass. We further repeated our literature search using Google Scholar and ordered the studies by relevance in order to ensure that our search captured all relevant studies. All studies in the top 200 found by Google Scholar that met our criteria were already included in our dataset. Together, this is the most comprehensive synthesis so far on quantifying fire impacts on soil biogeochemistry, which can help to obtain a more generic understanding of future of ecosystem functioning in a world subjected to more severe, recurrent, and further-reaching wildfires.
In parallel, we also compiled the geographic information including latitude and longitude, climate information such as mean annual temperature (MAT) and precipitation (MAP). Both MAT and MAP were taken directly from the original study, or from papers cited in that study. If these data were not presented, they were extracted from the WorldClim database (www.worldclim.com) using geographical information. Annual precipitation over potential evaporation was also obtained from the WorldClim database, which was used to calculate aridity index (AI) as the ratio with annual precipitation. Soil pH, clay, bulk density (BD), and C:N were obtained data from the SoilGrids database (https://www.isric.org/explore/soilgrids). Plant production including Normalized Difference Vegetation Index (NDVI), Gross Primary Productivity (GPP), and Leaf Area Index (LAI) were obtained from the 20-year averages of products from NASA’s Land Processes Distributed Active Archive Center (https://lpdaac.usgs.gov/). We followed Beck et al. 44 to divide the Köppen classification into arid, cold, and tropical-temperate zones (Table S2). Plant functional type were divided into mycorrhizal association and function types. We used the database of Wang and Qiu (2016)45 and Soudzilovskaia et al. 46 to discern the dominant species as either AM (arbuscular mycorrhizal fungi) and ECM (ectomycorrizal fungi) (Supplementary Table S2). Most studies in our dataset were focused on AM and ECM, as biomes with nonmycorrhizal species (NM) were too few to allow for analysis. For ecosystems with mixed mycorrhizal types like savanna with ECM trees and AM grasses, the mycorrhizal status was defined according to the relative abundance of ECM plants compared to AM plants47. Plant functional types were grouped into angiosperms and conifers using information available to test whether the fire effects on soil biogeochemistry depended on tree type. Since the criteria of fire severity, frequency, and type may change with different sites, we directly used information on these factors from the selected references or cited papers to avoid any bias. Specifically, fire severity was divided into low, moderate, and heavy; and fire frequency was divided into “one” and “frequent” (Supplementary Table S2). We classified fire frequency as “frequent”, if the historic number of fires of the selected site was two or more during the past 20 years. The fire type was grouped as “wildfires” or “prescribed fires” based on the fire regime reported from the literature (Supplementary Table S2). We acknowledge that these fairly broad categorizations may represent only part of the complexity of fires occurring in nature, but finer categorization would not have provided sufficient samples for a robust meta-analysis. In addition, we also collected soil taxonomic classification of the selected sites from Harmonized World Soil Database (HWSD, https://gaez.fao.org/pages/hwsd). Twenty four soil types including Luvisols, Cambisols, Leptosols, Podzoluvisols, Gleysols, Arenosols, Acrisols, Solonchaks, Podzols, Phaeozems, Histosols, Regosols, Solonetz, Greyzems, Gypsisols, Fluvisols, Andosols, Kastanozems, Alisols, Vertisols, Chernozems, Nitisols, Ferralsols and Anthrosols were recorded in our dataset.
Influence of fire on soil biogeochemistry
To analyze the direction and magnitude of the changes in soil biogeochemistry under fire disturbance, a meta‐analysis was conducted, which is a quantitative, scientific synthesis tool to provide reliable, broad scale conclusions48. Specifically, the natural logarithm (Ln) of the response ratio (RR) as LnRR= Ln (fire/control) as described by Mooney et al.49 and Crouzeilles et al.50 was used to explore fire effects globally. Here, fire is the mean value of a particular variable subjected to fire disturbance in a given time (year), while control is the value for the unburned controls (i.e., without fire disturbance of nearby natural reference ecosystems). The log response ratio is positive when the value of a particular variable of fire treatment is higher than that in the control treatment. We then calculated the average LnRR values for all variables within soil biogeochemistry, but also for variables grouped as different plant functional types and Köppen classifications. The potential publication bias was first tested by the Egger’s regression test along with inspecting funnel plots51. The Egger’s regression test was applied using the meta-analytic residuals, which consist of sampling errors as well as the effect-size-level effects that are equivalent to normal residuals52. We then cross-validated our results using Rosenthal’s fail-safe number, because it can consider both linear and nonlinear hypotheses and can be widely used for multiple types of meta-analysis53. Both Egger’s regression test and Rosenthal’s fail-safe number were conducted using the metafor package in R54. Egger’s regression test on the meta-analytic residuals showed some potential funnel asymmetry (Supplementary Fig. S4); however, Rosenthal’s fail-safe number suggested that no evidence for publication bias exists (Supplementary Table S3). Taken together, it is likely that publication bias of our global dataset is very limited, and the studies included in the database can been seen as a representative sample of available studies. To be consistent with previous studies, we took the conservative approach of not weighting effect sizes by their variance as described by Mooney et al.49, which provides similar results compared to those using the weighted method50. This approach has a great advantage over other weighted metrics, since it only requires the mean data for the dependent variables for two groups, while other weighted metrics require some measure of variance (e.g., SD) and sample sizes of both control and treatment. This allowed us to analyze a large dataset with >5000 observations. We then created a sub-dataset with variance (i.e., SD) that accounted for more than 90% of the total data to further cross-validate our results using the variance-weighted method15, which confirmed that fire disrupts soil biochemical coupling (Supplementary Fig. S5).
Influence of fire on soil biogeochemistry and size effects
We followed the method of Zhou et al.55 and Eldridge & Delgado-Baquerizo (2017)56 to quantify the average effect of fire on soil biogeochemistry cycling attributes, using linear mixed models in R package lme4 with LnRR as the dependent variable. The significance of these models was tested with likelihood ratio tests. Estimates of resistance were derived from restricted maximum likelihood and 95% confidence intervals for the estimates obtained from the likelihood profile. We examined the spatiotemporal variations of the soil biogeochemistry as affected by MAT, MAP, Aridity, GPP, LAI, NDVI, Soil C:N, pH, BD, Clay, longitude, and latitude.
Multiple regression models
We used multiple regression models to assess the joint effects of climate, plant production, soil properties, fire regime, plant functional types, time and space on soil biogeochemistry attributes as well as plant biomass. This approach based on the variation partition model, allows us to isolate the unique contributions of specific variables (e.g., climate) while considering the effects of other predictors (e.g., soil properties). We included MAT, MAP, and aridity as climatic variables. Plant productivity was represented by GPP, NDVI, and LAI, while soil properties included pH, clay content, bulk density (BD), and soil C:N. Fire regime included fire type (i.e., belonged to wildfire or not) and the time since last fire. Given the importance of plant functional type in our analyses, we also included the effects of ECM and conifer to explore how plant function type, including mycorrhizal association, regulates the fire effects on soil biogeochemistry. In addition, we included latitude and longitude as space factors. All predictors and response variables were standardized before analyses, using the z-score to interpret parameter estimates on a comparable scale. The estimated parameter coefficients for each predictor were computed using the relaimpo package of the R statistical software.
Partial correlation and random forest analyses
Given the strong effect of time, partial correlation analyses were conducted to cross-validate the influence of diverse ecological divers such as MAT, MAP, Latitude, GPP, NDVI, LAI, soil C:N, pH, and Clay on soil biogeochemistry controlling for time since fire. Partial correlation analyses were conducted using packages ggm and psych in R as in Chen et al.57. Correlation coefficients and significance (P < 0.05) were presented as a heat map with contrasting colors representing positive and negative correlations, and circle size depicting the magnitude of the coefficient. Meanwhile, we used random forest analyses using the R package randomForest, rfPermute and A3 to explore the relative importance of diverse predictors, such as soil taxonomic classifications, plant production, soil properties, fire regime, plant functional types, and space in driving soil biogeochemical responses to fire.
Principal component analysis (PCA)
Considering that the nonlinear methods may be over-parameterized on highly diverse data, we conducted a principal component analysis (PCA) on the multivariate space of the diverse soil biogeochemistry (see Zhou et al.55). Specifically, ecosystem attributes were standardized using z-transformation. We then extracted the explained variance of each component (PCA), as well as the loadings of each biogeochemistry measures, demonstrating the contributions of each soil biogeochemical process to the relevant PCA axis.
Associations among diverse soil biogeochemistry attributes in response to fire
Network analysis was conducted to evaluate how diverse soil biogeochemistry attributes associated with each other, assessed which attribute was the most central58, and then identified clusters of attributes with similar responses to fire. Specifically, the pairwise Markov random field network model was used to estimate the Partial correlation networks, which was optimized using the extended Bayesian information criterion (‘EBICglasso’) with the ‘bootnet’ package59. We then computed the centrality by summing the absolute values of partial correlations to obtain the higher values quantified as important attributes to assess the importance of attributes in the network59. The strength of correlation was calculated using the accumulated values of absolute partial coefficients between a focal attribute and all other connected attributes in the network. The strength in the network was standardized by subtracting the mean from the specific values and dividing it by the standard deviation. Larger strength values indicate higher central attributes.
Mapping the global distribution of soil biogeochemistry in response to fire
To predict the extent of global soil biogeochemistry responses to fire, we conducted a Random Forest regression analysis using the variables: mean annual temperature (MAT), mean annual precipitation (MAP) from the WorldClim database (www.worldclim.com); soil properties including clay, BD, and pH, were obtained from the Soilgrids database; plant cover was obtained from the 2010 global plant cover production60; rainfall erosivity from the global Rainfall Erosivity Database (GloREDa)61; elevation, northness, and slope were obtained from the global topography database62; leaf area index (LAI), gross primary productivity (GPP), and normalized difference vegetation index (NDVI) were obtained from the 20-year averages of products from NASA’s Land Processes Distributed Active Archive Center (https://lpdaac.usgs.gov/). Specifically, LAI was obtained from MCD15A2H, GPP was obtained from the average values of products from MOD17A2H and MYD17A2H, and NDVI was obtained from the average values of products from MOD13A2H and MYD13A2H. Since time varied across our global selection of observations, we incorporated the average time since fire as predictor together with other variables such as soil properties and climate into the model. Meanwhile, we used the average value to map the global effect if the same site had multiple soil biogeochemical process measures (e.g., soil C, soil N). All grid data were resampled to a resolution of 0.1° × 0.1°. This model was built by finding the set of covariate combinations that most robustly predict the training samples with 2000 repetitions. We then correlated our global maps of soil biogeochemistry with the attributes of ecosystems in all locations. Our model selection was based on choosing models with a coefficient of determination (R²) greater than 0.3 during the training phase. We also calculated how much the parameter space of the predictors differed from the original datasets to test the accuracy of the predictions calculated from the random forest-based model. The coefficient of variation (CV) of the predicted soil biogeochemistry attributes was calculated by 1000 parallel runs of Random Forest model. We followed the method from Piñeiro et al.63 to validate modeling approach by returning the predicted values (y-axis) vs. the observed values (x-axis). All the gridded data were aggregated to 1° × 1°spatial resolution.
Model validation
Based on the machine learning method, we used 70% of the data as “training” data, and the remaining 30% of the data was treated as “validation” data to validate the concerned variables (e.g., soil C, soil N, soil P, soil microbial biomass, SOM decomposition, and plant biomass) in the global map. Since there is a potential spatial autocorrelation, we employed a five-fold cross-validation approach to validate machine learning models for each variable64. The training dataset was randomly partitioned into five groups, with four groups designated for model training and one group reserved for testing. This random partitioning was conducted in a stratified manner, based on ecosystem types and climate zones, to ensure a comprehensive representation of the diverse range of ecosystems and climatic conditions in both the training and testing datasets. Furthermore, each fire site was exclusively assigned to a single group, allowing for the assessment of model performance in previously unobserved locations. We calculated cross-validated model R2, root mean squared error (RMSE), and median absolute error (MAE) using a leave-one-out cross-validation scheme for each concerned variable. Random forest ensembles were trained after holding out each observation one at a time, and the highest R2 were used to make global predictions.
Data availability
All data generated and analyzed in this study have been deposited inthe figshare database (https://doi.org/10.6084/m9.figshare.28034705)65.
Code availability
Codes for processing the data in this study have been deposited in the figshare database (https://doi.org/10.6084/m9.figshare.28034960)66.
References
Belcher, C. M. Fire Phenomena and the Earth System: An Interdisciplinary Guide to Fire Science (Wiley-Blackwell, 2013).
Pellegrini, A. F. et al. Fire effects on the persistence of soil organic matter and long-term carbon storage. Nat. Geosci. 15, 5–13 (2022).
Feng, X. et al. How deregulation, drought and increasing fire impact Amazonian biodiversity. Nature 597, 516–521 (2021).
Bowd, E. J., Banks, S. C., Strong, C. L. & Lindenmayer, D. B. Long-term impacts of wildfire and logging on forest soils. Nat. Geosci. 12, 113–118 (2019).
Ochoa-Hueso, R. et al. Ecosystem coupling: A unifying framework to understand the functioning and recovery of ecosystems. One Earth 4, 951–966 (2021).
Agbeshie, A., Abugre, S., Atta-Darkwa, T. & Awuah, R. A review of the effects of forest fire on soil properties. J. Forestry Res. 33, 1419–1441 (2022).
Finzi, A. C. et al. Coupled biochemical cycles: responses and feedbacks of coupled biogeochemical cycles to climate change. Examples from terrestrial ecosystems. Front. Ecol. Environ. 9, 61–67 (2011).
Delgado-Baquerizo, M. et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 502, 672–676 (2013).
Wardle, D. A. & Walker, L. R. mBardgett, R. D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 305, 509–513 (2004).
Bauters, M. et al. Fire-derived phosphorus fertilization of African tropical forests. Nat. Commun. 12, 5129 (2021).
Wilkinson, S. L. et al. Wildfire and degradation accelerate northern peatland carbon release. Nat. Clim. Change 13, 456–461 (2023).
Xu, S. et al. Fire frequency and type regulate the response of soil carbon cycling and storage to fire across soil depths and ecosystems: A meta-analysis. Sci. Total Environ. 825, 153921 (2022).
Dooley, S. R. & Treseder, K. K. The effect of fire on microbial biomass: a meta-analysis of field studies. Biogeochemistry 109, 49–61 (2012).
Cheng, Y. et al. Fire effects on soil carbon cycling pools in forest ecosystems: A global meta-analysis. Sci. Total Environ. 895, 165001 (2023).
Li, J. et al. Spatiotemporal variability of fire effects on soil carbon and nitrogen: A global meta‐analysis. Glob. Change Biol. 27, 4196–4206 (2021).
Liang, C. & Balser, T. C. Warming and nitrogen deposition lessen microbial residue contribution to soil carbon pool. Nat. Commun. 3, 1222 (2012).
Tang, R. et al. Increasing terrestrial ecosystem carbon release in response to autumn cooling and warming. Nat. Clim. Change 12, 380–385 (2022).
Gold, Z. J. et al. Herbaceous vegetation responses to experimental fire in savannas and forests depend on biome and climate. Ecol. Lett. 26, 1237–1246 (2023).
McLauchlan, K. K. et al. Fire as a fundamental ecological process: Research advances and frontiers. J. Ecol. 108, 2047–2069 (2020).
Zhou, Y. et al. Limited increases in savanna carbon stocks over decades of fire suppression. Nature 603, 445–449 (2022).
Brianne, P., Rebecca, H. & David, L. The fate of biological soil crusts after fire: a meta-analysis. Glob. Ecol. Conserv 24, e01380 (2020).
Phillips, R. P., Brzostek, E. & Midgley, M. G. The mycorrhizal‐associated nutrient economy: a new framework for predicting carbon–nutrient couplings in temperate forests. N. Phytol. 199, 41–51 (2013).
Zhou, Y., Biro, A., Wong, M. Y., Batterman, S. A. & Staver, A. C. Fire decreases soil enzyme activities and reorganizes microbially mediated nutrient cycles: A meta‐analysis. Ecology 103, e3807 (2022).
Zhou, L. et al. Fire decreases soil respiration and its components in terrestrial ecosystems. Funct. Ecol. 37, 3124–3135 (2023).
Butler, O. M., Elser, J. J., Lewis, T., Mackey, B. & Chen, C. The phosphorus-rich signature of fire in the soil-plant system: a global meta-analysis. Ecol. Lett. 21, 335–344 (2018).
Su, W. Q., Yu, M., Lin, J., Tang, C. & Xu, J. Fire decreases gross mineralization rate but does not alter gross nitrification rate in boreal forest soils. Soil Biol. Biochem. 175, 108838 (2022).
Hu, M., Wang, J., Lu, L., Gui, H. & Wan, S. Global recovery patterns of soil microbes after fire. Soil Biol. Biochem. 183, 109057 (2023).
Vitousek, P. M. Nutrient Cycling and Limitation: Hawai’i as a Model System (Princeton Univ. Press, 2004).
McGill, W. B. & Cole, C. V. Comparative aspects of cycling organic C, N, S and P through soil organic matter. Geoderma 26, 267–286 (1981).
Ryan, K. C., Knapp, E. E. & Varner, J. M. Prescribed fire in North American forests and woodlands: history, current practice, and challenges. Front. Ecol. Environ. 11, e15–e24 (2013).
Lindenmayer, D. B. et al. Logging elevated the probability of high-severity fire in the 2019–20 Australian forest fires. Nat. Ecol. Evol. 6, 533–535 (2022).
Meng, C. et al. Nutrient availability and microbial traits constrained by soil texture modulate the impact of forest fire on gross nitrogen mineralization. For. Ecol. Manag. 541, 121067 (2023).
Bodí, M. B. et al. Wildland fire ash: Production, composition and eco-hydro-geomorphic effects. Earth Sci. Rev. 130, 103–127 (2014).
Popović, Z., Bojović, S., Marković, M. & Cerdà, A. Tree species flammability based on plant traits: A synthesis. Sci. Total Environ. 800, 149625 (2021).
Hawkins, H. J. et al. Mycorrhizal mycelium as a global carbon pool. Curr. Biol. 33, R560–R573 (2023).
Crowther, T. W. et al. The global soil community and its influence on biogeochemistry. Science 365, eaav0550 (2019).
Patoine, G. et al. Drivers and trends of global soil microbial carbon over two decades. Nat. Commun. 13, 4195 (2022).
Chapin, II. F. S., Matson, P. A. & Mooney, H. A. Principles of Terrestrial Ecosystem Ecology (Springer, 2002).
Dove, N. C. et al. High-severity wildfire leads to multi-decadal impacts on soil biogeochemistry in mixed-conifer forests. Ecol. Appl 30, e02072 (2020).
Schmitz, O. J. et al. Trophic rewilding can expand natural climate solutions. Nat. Clim. Change 13, 324–333 (2023).
D’Odorico, P., Okin, G. S. & Bestelmeyer, B. T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 5, 520–530 (2012).
González-Pérez, J. A., González-Vila, F. J., Almendros, G. & Knicker, H. The effect of fire on soil organic matter—a review. Environ. Int. 30, 855–870 (2004).
Dube, O. P. Linking fire and climate: interactions with land use, vegetation, and soil. Curr. Opin. Env Sust. 1, 161–169 (2009).
Beck, H. E. et al. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 5, 1–12 (2018).
Wang, B. & Qiu, Y. L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16, 299–363 (2006).
Soudzilovskaia, N. A. et al. FungalRoot: global online database of plant mycorrhizal associations. N. Phytol. 227, 955–966 (2020).
Luo, S. et al. Higher productivity in forests with mixed mycorrhizal strategies. Nat. Commun. 14, 1377 (2023).
Gurevitch, J., Koricheva, J., Nakagawa, S. & Stewart, G. Meta-analysis and the science of research synthesis. Nature 555, 175–182 (2018).
Mooney, K. A. et al. Interactions among predators and the cascading effects of vertebrate insectivores on arthropod communities and plants. Proc. Natl Acad. Sci. USA 107, 7335–7340 (2010).
Crouzeilles, R. et al. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 3, e1701345 (2017).
Egger, M., Smith, G. D., Schneider, M. & Minder, C. Bias in meta-analysis detected by a simple, graphical test. Brit. Med. J. 315, 629–634 (1997).
Nakagawa, S. & Poulin, R. Meta-analytic insights into evolutionary ecology: an introduction and synthesis. Evol. Ecol. 26, 1085–1099 (2012).
Liu, Z. et al. A global meta-analysis on the drivers of salt marsh planting success and implications for ecosystem services. Nat. Commun. 15, 3643 (2024).
Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 36, 1–48 (2010).
Zhou, G. et al. Temperature and rainfall patterns constrain the multidimensional rewilding of global forests. Adv. Sci. 9, 2201144 (2022).
Eldridge, D. J. & Delgado‐Baquerizo, M. Continental‐scale impacts of livestock grazing on ecosystem supporting and regulating services. Land Degrad. Dev. 28, 1473–1481 (2017).
Chen, L. et al. Regulation of priming efect by soil organic matter stability over a broad geographic scale. Nat. Commun. 10, 5112 (2019).
Poorter, L. et al. Multidimensional tropical forest recovery. Science 374, 1370–1376 (2021).
Epskamp, S. & Fried, E. I. A tutorial on regularized partial correlation networks. Psychol. Methods 23, 617–634 (2016).
Hansen, M. C. et al. High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013).
Amatulli, G. et al. A suite of global, cross-scale topographic variables for environmental and biodiversity modeling. Sci. Data 5, 1–15 (2018).
Panagos, P. et al. Global rainfall erosivity assessment based on high-temporal resolution rainfall records. Sci. Rep. 7, 1–12 (2017).
Piñeiro, G. et al. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Model 216, 316–322 (2008).
Gordó-Vilaseca, C. et al. Future trends of marine fish biomass distributions from the North Sea to the Barents Sea. Nat. Commun. 15, 5637 (2024).
Zhou, G. Database.xls. Figshare, https://doi.org/10.6084/m9.figshare.28034705 (2025).
Zhou, G. Code. Figshare, https://doi.org/10.6084/m9.figshare.28034960.v1 (2025).
Acknowledgements
G.Y.Z. acknowledges the supports from Ramón y Cajal grant (RYC2022-035226-I) funded by the Spanish Ministry of Science and Innovation, the NextGenerationEU program of the European Union (MICIU/AEI/10.13039/501100011033y el FSE+), and AYUDAS DE EXCELENCIA RYC-MAX 2023 project from Spanish National Research Council. N.E acknowledge support by iDiv ([German Research Foundation, DFG]–FZT 118, 202548816), and the DFG (Ei 862/29-1; Ei 862/31-1). The Jena Experiment is funded by the DFG (FOR 5000). M.D.-B. acknowledges support from the Spanish Ministry of Science and Innovation for the I + D + i project PID2020-115813RA-I00 funded by MCIN/AEI/10.13039/501100011033. M.D.-B. is also supported by a project of the Fondo Europeo de Desarrollo Regional (FEDER) and the Consejería de Transformación Económica, Industria, Conocimiento y Universidades of the Junta de Andalucía (FEDER Andalucía 2014-2020 Objetivo temático “01 - Refuerzo de la investigación, el desarrollo tecnológico y la innovación”) associated with the research project P20_00879 (ANDABIOMA). M.B. is supported by a Ramón y Cajal grant from the Spanish Ministry of Science (RYC2021-031797-I). X.H.Z. is supported by National key Research and Development Program of China (2020YFA0608403). D.R. is supported by the MSCA postdoctoral fellowship from Horizon Europe programme (MSCA-PF-2021-101064192-FUNVINE). We thank Nan Li, Yuan Qi, Jingjing Meng, and Jianing Pei for their assistance with data extraction. We thank all the scientists whose data and work were included in this global synthesis.
Author information
Authors and Affiliations
Contributions
M.D.-B. and G.Y.Z. conceived this work. G.Y.Z., Z.G.D. and H.W. developed the methodology. G.Y.Z., M.D.-B., N.E., M.B. wrote the original draft. N.E., M.E.L., K.Y.Z., S.E.L., H.M.D., H.W., D.R., T.S.S., H.C., X.Z. reviewed and edited the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Peng Luo, who co-reviewed with Honglin Li, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhou, G., Eisenhauer, N., Du, Z. et al. Fire-driven disruptions of global soil biochemical relationships. Nat Commun 16, 1190 (2025). https://doi.org/10.1038/s41467-025-56598-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-56598-z
This article is cited by
-
Wildfires under changing climate, and their environmental and health impacts
Journal of Soils and Sediments (2025)
