
Abstract
Recent studies have highlighted the crucial role of microglia (MG) and their interactions with the gut microbiome in post-stroke neuroinflammation. The activation of immunoregulatory pathways, including the aryl hydrocarbon receptor (AHR) pathway, is influenced by a dynamic balance of ligands derived from both the host and microbiota. This study aimed to investigate the association between stroke-induced dysbiosis and the resultant imbalance in AHR ligand sources (loss of microbiota-derived [indole-based] and increase of host-derived [kynurenine-based]) after stroke. Microbiota-derived AHR ligands decreased in human plasma and remained low for days following an ischemic stroke highlighting the translational significance. Transient-middle-cerebral-artery-occlusion was performed in aged wild-type and germ-free male mice. MG-AHR expression and activity increased in both in vivo and ex vivo stroke models. Germ-free mice showed altered neuroinflammation and antigen presentation while aged mice showed reduced infarct volume and neurological deficits following treatment with microbiota-derived AHR ligands after stroke. Restoring a balanced pool of host- and microbiota-derived AHR ligands may be beneficial after stroke and may represent a therapeutic target.
Similar content being viewed by others


Introduction
Approximately 800,000 individuals experience a new or recurrent stroke annually1. Ischemic stroke, which is the most prevalent, is caused by reduced cerebral blood supply due to thrombosis or embolism2. Post-ischemic stroke survival rates have improved with advancements in acute stroke management such as endovascular thrombectomy and the use of intravenous thrombolytics like alteplase and tenecteplase3. However, these treatment options are available to <10% of patients with stroke due to exclusion criteria related to timing, risk of hemorrhage, and the need for specialized resources4. Moreover, post-stroke poor functional outcomes contribute considerably to reduced quality of life and increase the economic burden of stroke5. Despite breakthroughs in understanding of the immune response after stroke, no therapeutic options are currently available for controlling neuroinflammation following stroke6.
Recent studies have highlighted the crucial role of gut microbiota and its metabolites in post-stroke immune responses via molecular pathways including the aryl hydrocarbon receptor (AHR) pathway7. AHR, a versatile ligand-activated transcription factor, is expressed by various cell types, including T-cells, neutrophils, astrocytes, and microglia (MG). AHR plays a critical role in immune function and inflammation8,9. MG, the resident immune cells of the brain, are crucial for the regulation of post-stroke neuroinflammation and antigen presentation.
Stroke is primarily an age-related disease with outcomes inextricably linked to changes in the function of MG caused by aging10. Ongoing research on aging mechanisms show that gut dysbiosis significantly impacts age-related variations in MG responses7. Thus, both age- and stroke-induced dysbiosis may exacerbate the MG response to stroke, necessitating analysis of the pathways linking aging, gut microbiota, and MG-mediated immune processes post-stroke7.
The AHR pathway is a key regulator of neuroinflammation and a molecular sensor of gut microbiota-derived metabolites9,11. The ligand-specific effects of AHR are observed in several inflammatory diseases, including atherosclerosis and inflammatory bowel disease12,13,14. Exploring the role of MG AHR in age- and stroke-induced dysbiosis may improve the understanding of the complex gut-brain axis mechanisms that underlie post-stroke neuroinflammation.
The AHR pathway is activated by both exogenous and endogenous ligands, including phytochemicals, metabolic byproducts, and host- and microbiota-derived ligands9. Host-derived AHR ligands are primarily produced via the Kynurenine (Kyn) pathway, whereas microbiota-derived AHR ligands are predominantly produced via the indole pathway15,16,17,18. Indoles are found in plant-based foods such as cruciferous vegetables and produced by bacterial metabolism of dietary tryptophan (Trp). Indole-mediated activation of AHR forms a dynamic signaling pathway between the gut microbiota and brain immune compartment in both homeostasis and disease states19. AHR plays a complex role in MG function, mediating both pro- and anti-inflammatory effects depending on the specific ligand profile in the cellular environment20. Thus, studying the effects of post-stroke dysbiosis on MG AHR activation is important for understanding the complex mechanisms underlying post-stroke inflammation and identifying new therapeutic targets. Despite recent advances, the role of microbiota-dependent AHR activation in post-stroke immune responses remains unclear. However, focused research in this area may lead to novel therapeutic strategies for modulating neuroinflammation and improving the outcomes for patients with stroke.
Plasma Kyn concentrations increase within hours of stroke in humans21. In mice, Kyn-dependent AHR activation post-stroke induces inflammation and neurodegeneration, but its inhibition can be neuroprotective and it carries risks from AHR’s diverse physiological roles22. We hypothesized that stroke-induced dysbiosis leads to the loss of essential microbiota-derived AHR ligands, and restoring the balance between host-derived (Kyn-based) and microbiota-derived (indole-based) AHR ligands may provide therapeutic benefits. To test our hypothesis, we subjected aged wild-type (WT) and germ-free (GF) mice to transient middle cerebral artery occlusion (MCAO). We used metabolomics, flow cytometry, and microbiome sequencing techniques to determine whether post-stroke dysbiosis disrupts the supply of microbiota-derived AHR ligands following experimentally induced stroke. AHR expression increased after stroke in both mice and human MG. Metabolomic targets, which were validated in human samples from patients with ischemic stroke, showed a significant reduction in both indolepropionate and 3-formylindole. Restoring a balanced pool of AHR ligands following stroke via the exogenous administration of indoles provided therapeutic benefits in aged WT and GF mice. Our findings suggest that restoring a well-balanced pool of host- and microbiota-derived AHR ligands has considerable therapeutic potential for acute ischemic stroke treatment.
Results
Microglia from postmortem brain tissue samples of patients with stroke show high levels of AHR expression
First, we assessed changes in the MG AHR expression after stroke in humans. The final sample selection was based on radiological and gross histological confirmation of ischemic stroke (Fig. 1a, b). Immunohistochemical analysis of brain tissue samples showed significantly increased expression of AHR that was co-expressed with ionized calcium binding adapter molecule 1 (Iba-1+) cells (an established marker of human MG) after stroke relative to that of age-matched controls without stroke (Fig. 1c). We found significantly increased Iba1+ AHR+ cells per area in the human brain tissue samples from the acute to sub-acute timepoints relative to the chronic timepoints (Fig. 1d). A significant increase in the expression of AHR and its transcriptional activity (assessed by the transcriptional expression of cytochrome P450 1B1 Cyp1b1) was observed after ex vivo oxygen-glucose deprivation and reperfusion (OGD/R) for 2 h followed by reperfusion-like conditions for 4 h in a human MG cell line (HMC3) (Fig. 1e). The patients with stroke (n = 60) had significantly decreased plasma concentrations of microbiota-derived indole-based AHR ligands relative to the controls (n = 64), and this persisted for up to 7 days after the stroke (Fig. 1f). There was no significant correlation between the plasma concentration of indole-based AHR ligands and stroke severity, as assessed by the National Institute of Health (NIH) Stroke Scale23 (Supplementary Fig. S1).

Activation of MG AHR after oxygen-glucose deprivation and reperfusion (OGD/R) using human cell lines, with decreased concentrations of indoles in human plasma samples after stroke. a, b Final sample selection was based on radiological and gross histological confirmation of ischemic stroke. c Representative images of double immunofluorescent labeling for AHR (green) and Iba-1 (pink) in brain sections. They show AHR expression in Iba-1+ cells and quantification of AHR + MG per unit area in the infarct (p < 0.0001) and peri-infarct regions (p < 0.0001) post-stroke (n = 29) relative to the cortical regions from controls (n = 10). Each dot represents a patient. Scale bar: 100 um. d Representative images of double immunofluorescent labeling for AHR (green) and Iba-1 (pink) in brain sections showing AHR expression in Iba-1+ cells in acute and chronic timepoint postmortem brain samples with quantification of AHR expression in Iba-1 positive cells. e RT-PCR demonstrating increased AHR and Cyp1ba expression levels after OGD/R performed on human MG cell line (HMC3) (n = 5/grp). f Plasma concentrations of indole-3-carboxaldehyde (p < 0.0001, exact value shown on figure) and indole-3-propionate (p < 0.0001, exact value shown on figure, pairwise test: Games-Howell, bars shows significant) are significantly decreased 24 h to 7 days after stroke in patients (n = 60) relative to healthy controls (n = 64). Unpaired two-tailed t-test (e) and Tukey’s two-way ANOVA with multiple comparisons (c, d) were used for statistical analyses. All data (with error bars) are presented as mean ± SEM.
Bioavailability of gut microbiota-dependent indole-based AHR ligands was disrupted after stroke
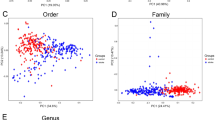
Metabolomic analysis using mass spectrometry of age-matched brain and plasma samples from WT and GF mice revealed the relationships between the brain and plasma concentrations of specific indole-based AHR ligands and presence of microbiota in mice. At least two known microbiota-derived indole-based AHR ligands (indole-3-propionic acid [IPA] and indole-3-aldehyde aka indole-3-carboxyaldehyde [IAld]) were undetectable in the brains of GF mice, and their concentrations were significantly lower in the plasma of the GF mice than in the WT samples (Fig. 2a). In contrast, the concentrations of Kyn and Trp did not differ in the GF and WT brains (Fig. 2b), indicating that they were host- and diet-derived, respectively. We performed 16S rRNA gene sequencing analysis of the large intestinal content to examine variations in microbial diversity after stroke. We found that the major bacterial populations involved in the regulation of AHR ligands and Trp metabolism (such as phylum Actinobacteria predominant genus being Bifidobacteria) were significantly reduced by aging and experimental stroke in aged WT mice (Fig. 2c (1) and Supplementary Fig. S2). The weighted UniFrac distances determined by principal coordinate analysis indicated a significant shift in the beta or inter-sample diversity at 24 h post-stroke relative to the sham controls (Fig. 2d). As expected, we did not observe significant differences in the alpha diversity (measures within sample diversity) and found no differences between the operational taxonomic units (OTUs) (p = 0.7 or 0.5) of the groups (Fig. 2c (2)). As seen in Fig. 2d, there was a significant clustering effect (p ≤ 0.002) along the PC1 axis (29.7% of the variation explained) in the animals with stroke relative to the sham groups, indicating a change in microbial diversity. Consistent with this, the LDA score also indicated changes in the bacterial families in the animals with stroke relative to the sham animals (Fig. 2e). Specific pathobionts (opportunistic pathogens), such as Enterobacteriaceae and Gammaproteobacteria (containing the genera Escherichia, Shigella, Enterococcus, and Erysipelatoclostridium), were significantly increased in mice with stroke24. This indicates that stroke induces gut dysbiosis, which occurs as early as 24 h after stroke (Fig. 2e). Additionally, cladogram visualization showed a clear pathological shift in bacterial diversity in animals with stroke relative to healthy sham animals (Fig. 2f).

a Metabolomics analysis of plasma samples from wild-type (WT) and germ-free (GF) mice. Host-derived metabolites (tryptophan [Trp] and Kynurenine [Kyn]) did not differ between WT and GF plasma samples. However, indole-3-carboxaldehyde and indole-3-propionate concentrations were significantly reduced in GF plasma (n = 5/grp). b Metabolomic analysis of brains revealed specific indole-based AHR ligands that depend on the presence of a microbiota. Indole-3-carboxaldehyde (p = 0.0122) and indole-3-propionate (p < 0.0001) were undetectable in GF brains relative to WT control brains, while Trp and Kyn concentrations did not differ (n = 5/grp). c1 Major bacterial populations involved in the regulation of AHR ligands and Trp metabolism (such as phylum Actinobacteria predominant genus being Bifidobacteria) were significantly (Mann–Whitney test: p < 0.001614, FDR-Adi.p < 0.003766; overall Median: 0.08179) reduced by aging and experimental stroke in aged WT mice. c2 No significant differences (Observed OTUs: p < 0.7 and Shannon: p < 0.54) in alpha diversity (measures within sample diversity) and operational taxonomic units (p = 0.7 or 0.5) between the groups were observed. d Principal component analysis plot of 16S data shows a significant (p = 0.002) clustering effect of stroke animals relative to the sham group. e The LDA score clearly shows changes in bacterial families in stroke versus sham animals. f Cladogram visualization of 16S data shows a clear pathological shift in bacterial diversity in stroke animals relative to healthy sham animals. g Metabolomics analysis of aged WT brain and plasma samples at multiple timepoints post-stroke (n = 5/grp). Kyn concentrations increased while concentrations of indole-3-carboxaldehyde and indole-3-propionate were significantly decreased in both brain and plasma (n = 5/grp). h Metabolomic analysis of aged WT plasma samples shows increased concentrations of Kyn at 3 days post-stroke and no significant difference between groups at 30 days post-stroke (n = 5/control, 6/treated). Concentrations of indole-3-carboxaldehyde and indole-3-propionate (p = 0.0182) were decreased at 3 days post-stroke (n = 5/control, 6/treated). Unpaired two-tailed t-tests were used for statistical analyses (a, b, h). All data (with error bars) are presented as mean ± SEM. All data points (each dot represents) presented are biological replicates.
Given the established ligand specificity of AHR14, we evaluated changes in the host- and microbiota-derived AHR ligands in the brains of aged WT mice at multiple timepoints after stroke. The concentrations of Kyn significantly increased, which was consistent with previous reports18. The concentrations of IAld and IPA decreased in both the plasma and brain 24 h after stroke, indicating a loss of microbiota-derived indole-based AHR ligands (Fig. 2g). Metabolomic analysis of the blood plasma on day 3 after stroke showed a significant loss of IPA relative to that in the age-matched sham controls (Fig. 2h). Consistent with previous reports in the literature, we observed elevated plasma concentrations of Kyn at 3 days after stroke (Fig. 2h).
Host- and microbiota-derived AHR ligands had opposing effects on MG survival after OGD/R
AHR was activated after 2-h ex vivo OGD/R of sorted MG from naive aged WT mice treated with either Kyn or indole cocktail (IAld and IPA) (Fig. 3b). The depletion of Trp from the medium prior to OGD/R prevented AHR activation after OGD/R, indicating that the conversion of Trp to Kyn was the predominant mechanism underlying AHR activation in the absence of indole-based ligands (Fig. 3b). The addition of Kyn or indole cocktail (IAld and IPA) to Trp-depleted media led to a significant increase in the levels of AHR expression after OGD/R, which was consistent with the partial agonistic activity of both classes of ligands. The survival rate of sorted MG cells was significantly lower than that of Trp-depleted controls after OGD/R when Kyn was added. In contrast, adding IAld and IPA cocktail had no significant effect on MG survival after OGD/R (Fig. 3b).

a Schematic showing sorted MG cells from naive aged WT mice treated with indole-based AHR ligands (indole-3-carboxaldehyde and indole-3-propionate) ex vivo. The cells received OGD/R to model stroke conditions. b MG AHR expression and MG survival post-OGD/R differ based on the AHR ligands present. AHR expression was increased after OGD/R and depleting tryptophan (Trp) from OGD/R media reversed this effect (n = 4/group for Vehicle, OGD, Trp Dep OGD (p < 0.0100), and Trp Dep Kyn OGD (p < 0.0001) and n = 8 for Trp Dep Indoles OGD (p < 0.0001) group). The addition of Kynurenine (Kyn) or indole-based AHR ligands (indole-3-carboxaldehyde and indole-3-propionate) led to an increase in AHR expression post-OGD/R. MG survival after OGD/R was significantly lower when Kyn was present relative to when Trp was depleted. Post-OGD/R MG survival increased when indole-based ligands were added (n = 4/group for Vehicle, OGD, Trp Dep OGD, and Trp Dep Kyn OGD and n = 8 for Trp Dep Indoles OGD group). c Schematic showing treatment of mice with indoles (indole-3-carboxaldehyde and indole-3-propionate) in vivo via oral gavage. MG were sorted from indole-treated mice and received OGD/R. d MG survival was increased post-OGD/R in the group treated with indoles relative to the vehicle group (n = 5/grp, p = 0.0249). e Kyn-mediated activation of AHR was detrimental, whereas indole-mediated activation of AHR did not worsen MG survival after OGD/R (n = 6/grp, p = 0.0256). f qPCR-based examination showed that the expression levels of IL-1b, IL−4, and IL-10 are significantly reduced in indole-treated relative to the Kyn-treated group (n = 6/grp, p < 0.0001 for all for all three comparisons). The schematic figures were created in BioRender. Unpaired two-tailed t-test (d) and Tukey’s two-way ANOVA with multiple comparisons (b, e, f) were used for statistical analyses. All data (with error bars) are presented as mean ± SEM. All data points presented are biological replicates.
Cells survived longer after OGD/R in the MG from the mice treated with the indole cocktail than in those from the mice that received the vehicle (Fig. 3c, d). MG cells survived longer after OGD/R of animals treated with indole cocktail (Fig. 3b, d). This suggested that host- and microbiota-derived AHR ligands had differential effects on MG survival after OGD/R (Fig. 3b). Specifically, Kyn-mediated activation of AHR (verified by Cyp1b1 quantitative polymerase chain reaction [qPCR] (Supplementary Fig. S3)) was detrimental, whereas indole-mediated activation of AHR did not reduce the duration of MG survival after OGD/R. In addition, flow and qPCR-based evaluation revealed significantly lower concentrations of interleukin (IL)-4 and IL-10 in the indole-treated groups than in the Kyn-treated groups in primary MG and cultured MG cells (Fig. 3e, f and Supplementary Fig. S4).
Post-stroke treatment with microbiota-derived indole-based AHR ligands regulated MG-mediated neuroinflammation and antigen presentation markers in GF mice
GF animal models that were devoid of bacteria and unable to convert Trp to indole-based molecules were treated with indole-based molecules orally. Metabolomic analysis of their brains and plasma showed increased concentrations of indole-based molecules (Supplementary Fig. S5). This confirmed that peripheral supplementation with indole-based ligands was sufficient to increase brain concentrations of these metabolites.
The brains of GF mice subjected to MCAO were immunophenotyped at 24 h post-stroke using flow cytometry (Fig. 4a). Post-stroke treatment with IAld and IPA 3 h post-reperfusion was associated with increased MG AHR expression (Fig. 4b, d). This increase in AHR expression was associated with higher surface expressions of CD11b, MHC-II, and CD80 by MG in the MCAO group after 24 h relative to the GF MCAO vehicle group (Fig. 4d). Unsupervised analysis of the flow cytometry data of the seven surface parameters of CD45, CD11b, MHC-II, CD80, P2RY12, and Tmem119 detected several subsets of MG, lymphocytes, and infiltrating monocytes (Supplementary Fig. S6). These were visualized as a phonographic comparison of the vehicle and treated GF stroke brains (Fig. 4c). A heatmap of the surface expression levels and fluorescence intensity plots of the cell populations detected by the phonograph algorithm is included as Supplementary Material (Supplementary Figs. S7 and S8).

a Schematics showing timeline of MCAO and indole treatment in GF mice. b Increased expression of MG AHR is associated with increased expression of antigen presenting and co-stimulatory molecules MHC-II and CD80. c Phenographs of non-MG and MG cells of homogenized brains of GF mice post-stroke from vehicle and indole-treated mice. d Surface expressions of AHR, CD11b, MHC-II, and CD80 by MG are significantly increased 24 h after stroke in GF mice treated with indoles at 3 h and 8 h after stroke relative to GF stroke mice receiving vehicle (n = 9/grp). e Surface expressions of MHC-II and CD80 by lymphocytes were significantly increased in mice treated with indoles (n = 9/grp). f Surface expression of MHC-II is significantly decreased and expression of CD80 is increased by monocytes in mice treated with indoles (n = 9/grp). g GF stroke mice treated with indoles had significantly lower reduction in body weight (normalized to pre-stroke bodyweight at 24 h after stroke, lower brain weight (normalized to bodyweight at 24 h post-stroke) and lower brain volume. Neurological deficit scores were not different at 24 h post-stroke in GF mice (n = 10/vehicle, 13/treated). h Expression of microglial Lamp1 (p = 0.0028), IL1B (p = 0.0078), and microglial phagocytosis (% red bead uptake) (p = 0.0256) were decreased in GF mice treated with indoles at 3- and 8-h post-stroke (n = 6/vehicle, 8/treated). Expression of IL10 was increased (p = 0.0825) following treatment with indoles (n = 6/vehicle, 8/treated). i, j Volcano plots and heatmap visualization of Nanostring mRNA expression analysis of homogenized brain tissues from the controls and indole-treated groups of GF stroke mice. The transcriptional profiles of AHR downstream target genes (Cyp1b1, Ifng, Ccl8, Il2, and Ccl20) in the indole-treated group (n = 8) were different. The figures were created in BioRender. Unpaired two-tailed t-test. All data (with error bars) are presented as mean ± SEM. All data points presented are biological replicates.
Examination of non-MG immune populations in the GF brain revealed that lymphocytes, similar to MG, showed higher expressions of MHC-II and CD80 after stroke with post-stroke indole treatment (Fig. 4e). In contrast, monocytes had higher expression of CD80 and lower expression of MHC-II in the treatment group than in the vehicle group (Fig. 4f). Brain weights and volumes were significantly reduced in the GF stroke animals with indole treatment (Fig. 4g). Indole (IPA, IAld) treatment significantly downregulated microglial IL1β and LAMP1+ expressions, which are markers of inflammatory signaling and phagocytic activity, respectively (Fig. 4h). Red bead engulfment by MG obtained from vehicle-treated mice brains after stroke was significantly increased relative to that by MG obtained from indole-treated mice brain at 24 h post-stroke (Fig. 4h). IL-10, an anti-inflammatory cytokine, showed a trend of upregulation with indole cocktail treatment after stroke but this was not significant (p < 0.0825). The mean fluorescence intensity demonstrated a significant increase for red bead engulfment, IL-1β, and LAMP1 in the vehicle-treated microglia relative to the indole-treated microglia. This suggested an anti-inflammatory effect of indole treatment on MG after stroke (Supplementary Fig. S9).
Nano String analysis of over 700 genes, including AHR downstream target genes, significant effects of treatment, as visualized by volcano plot and heatmap of the data (Fig. 4i, j). The Wilcoxon test revealed that Ahr- and Ahr-related genes were significantly upregulated in microglia of MCAO compared to that in the sham group using single-cell RNA sequencing data (Supplementary Fig. S10).
Post-stroke treatment with microbiota-derived indole-based AHR ligands reduced infarct size and neurological deficits in aged WT mice
Finally, we examined the effects of post-stroke treatment with indole-based AHR ligands in aged WT mice (Fig. 5a). The neurological deficit scores (NDS) of the aged WT MCAO mice treated with indole cocktail (IAld and IPA) post-reperfusion were significantly lower than those of the aged WT MCAO vehicle group at 24 h post-stroke (Fig. 5b). Compared with the vehicle group, aged WT MCAO mice treated with IAld and IPA had lower bodyweight loss (normalized to pre-stroke bodyweight) at 24 h after stroke, lower brain weights (normalized to bodyweight at 24 h post-stroke), and lower brain volume, suggesting a reduction in edema (Fig. 5b). Infarct volume analysis of aged WT brains showed that mice treated with the indole-based AHR ligands had significantly smaller total infarct volumes than the vehicle group measured at 24 h after stroke (Fig. 5c).

a Schematic showing the timeline of middle cerebral artery occlusion and indole treatment in aged WT mice. b Aged WT mice treated with indoles had a significant decrease in neurological deficit scores (p = 0.0150), smaller reduction in body weight (normalized to pre-stroke bodyweight, p = 0.0262) at 24 h after stroke, lower brain weight (normalized to bodyweight at 24 h post-stroke, p = 0.0018) and lower brain volume (n = 9/vehicle, 12/treated, p = 0.0003). Whiskers on box plot indicate minimum (2.00) and maximum (3.00). c Quantification of brain infarct volumes as analyzed by 2,3,5-triphenyl-tetrazolium staining in controls and treated mice (n = 9/vehicle, 12/treated). The total, cortical, and striatal infarct sizes suddenly increased in the stroke mice treated with indoles relative to the vehicle group (n = 9/vehicle, 12/treated). Unpaired two-tailed t-test. All data (with error bars) are presented as mean ± SEM. All data points presented are biological replicates. The figures were created in BioRender.
Taken together, these findings strongly support our hypothesis that restoring a balanced pool of host-derived Kynurenine- and microbiota-derived indole-based AHR ligands via post-stroke administration of indole-based AHR ligands can improve stroke outcomes in mice by modulating MG-mediated neuroinflammation and surface molecules involved in antigen presentation and immune co-stimulation.
Discussion
In the present study, we investigated the effects of treatment with microbiota-derived indole-based AHR ligands on MG-mediated neuroinflammation and surface molecules involved in antigen presentation and immune co-stimulation after stroke. We used human brain and plasma samples and GF and aged WT mouse models of ischemic stroke. Our results demonstrate that AHR expression is upregulated following stroke in humans. Post-stroke treatment with indole-based AHR ligands improves MG-mediated antigen processing and co-stimulatory immune functions. Treatment with indole-based compounds reduced infarct size and neurological dysfunction in aged WT mice by restoring a balanced pool of host- and microbiota-derived AHR ligands. A graphical representation of our findings is included as Fig. 6.

The schematics highlights the stroke induced pathological changes of host- and indole-based ligands availability of AHR measured in the circulation and in the ischemic hemisphere of the brain. It emphasizes that the oral treatment with indoles attenuated inflammation in the microglial of the post-stroke brain leading to less severe stroke outcome via AHR dependent pathway. The scheme was created under the BioRender Agreement No: DP27GHBB1A).
AHR is a highly conserved ligand-activated transcription factor that regulates immune differentiation and neuroinflammation9. AHR acts as a sensor that integrates environmental, dietary, metabolic, and microbial cues to regulate immune homeostasis and inflammatory conditions9,25,26. AHR binds to structurally diverse exogenous and endogenous compounds, including environmental toxins such as dioxins, phytochemicals such as flavonoids, host-derived Trp-based molecules such as kynurenines, and microbiota-derived indole-derivatives27. Most AHR ligands are low molecular weight, lipophilic polycyclic planar compounds18 that can easily pass through critical physiological barriers such as the intestinal epithelial barrier and blood-brain barrier (BBB)18,28,29,30,31. A major challenge in investigating AHR immunobiology is that the role of AHR is highly disease-, tissue-, cell-, and ligand-specific9.
Circulating Trp is predominantly albumin-bound, and that prevents its crossing of the BBB32,33. Free Trp is transported from blood across the BBB by a competitive, nonspecific large neutral amino acid transporter34,35. Trp is converted to Kyn by the host enzymes tryptophan 2,3-dioxygenase (TDO) and indoleamine 2,3-dioxygenase (IDO) under the influence of pro-inflammatory cytokines such as IL-1β and IL-636. TDO/IDO concentrations increase after stroke, and pharmacological inhibition of their enzymatic activity reduces AHR activation after stroke22. Kyn pathway metabolites have been detected in GF mice37,38. This was validated in our study, indicating that they were host-derived, even in complete absence of gut microbiota.
Another major class of AHR ligands, primarily regulated by gut microbiota enzymes such as bacterial tryptophanase, are the Trp-derived indole-based AHR ligands28,39,40. Trp is the only amino acid with an indole-containing structure. Indole production is dependent on the microbial catabolic activity of dietary Trp29,41. Indoles are aromatic heterocyclic molecules with planar structures formed by the fusion of a benzene and a nitrogenous pyrrole ring, which makes them structurally distinct from the Trp-derived Kyn-based AHR ligands. Indole-based metabolites are converted to various AHR ligands via hepatic uptake and metabolism9. Most indole-based AHR ligands are partial agonists, meaning that they can serve as competitive antagonists in the presence of other partial agonists. They outcompete the detrimental effects of Kyn and other host-derived AHR ligands42, which is an important premise of the present study.
Generating a global or cell-specific model (Ahr deletion in MG) cannot be used to directly test the main hypothesis of this study. Different AHR ligands share the same set of ligand-binding domains, specifically, the Per-AhR/Arnt-Sim domain of the AHR, which is the ligand-binding domain43. In other words, any knockout model that eliminates the activation of AHR by both Kyn- and indole-based ligands would not allow us to test whether indole-based ligands can reduce the detrimental effects of Kyn-based ligands.
Several endogenous Trp-derived AHR modulators (not exogenous toxins), particularly indole-based molecules, are more active (activation potential) toward human than toward rodent AHR18,19,41. The increased affinity of these endogenous AHR ligands implies an evolutionary adaptation in human host cells, allowing them to be influenced by diverse microbiota. These, in turn, mediate physiological responses, including inflammation. For example, the uremic toxin indoxyl-3-sulfate (I3S) demonstrates 500-fold greater potency in activating human AHR than murine AHR in hepatoma cell lines44, and therapeutic approaches such as AHR antagonism or inhibition of I3S synthesis have been proposed as treatments for chronic kidney disease45,46. Indole is a potent agonist of human AHR but a weak agonist of mouse AHR19. Accumulating data indicate that human AHR has a higher affinity for indole-based ligands than mouse AHR17, and our ex vivo experimental data confirmed the activation of MG AHR after OGD/R. This suggests that there may be greater therapeutic benefits from microbiota-based modulation of AHR in humans than in mice.
Microbiota-regulated Trp-derived ligands activate MG AHR to suppress activation of the nuclear factor kappa-light-chain-enhancer of activated B-cells (NF-κB) pathway. MG AHR activates the transforming growth factor alpha (TGF-α) promoter, which interferes with NF-kB-driven expression of vascular endothelial growth factor (VEGF)-B47. MG TGF-α and VEGF-B suppress and induce astrocyte-mediated inflammation, respectively9. Deletion of MG AHR exacerbates neuroinflammation and increases the recruitment of inflammatory monocytes to the brain47. MG-specific AHR deletion is detrimental in chronic neuroinflammation models such as experimental autoimmune encephalomyelitis47, suggesting that MG AHR is required for the anti-inflammatory effects of indole-based AHR activation. Our results demonstrated that host-derived Kyn-based and microbiota-derived indole-based molecules activate MG AHR, but has different effects on MG survival after OGD/R. Additionally, our results showed that MG antigen presentation and co-stimulation signaling molecules were significantly increased when GF mice were treated with a cocktail of IPA and IAld after stroke, indicating that MG-mediated neuroinflammation can be modulated by the post-stroke administration of indoles.
Kyn expression levels increase rapidly in the brain following stroke, and this promotes deleterious effects through the activation of the AHR. This activation exacerbates inflammation and neurotoxicity, contributing to the overall damage observed in cerebral ischemia22. Pharmacological AHR inhibitors or deletion of the Ahr gene in global knockout models decreases ischemic damage and improves NDS in young male mice16,22. AHR activity increases in the brain after ischemia in mice22. Post-stroke administration of two different pharmacological antagonists of AHR reduced infarct volume at 48 h in a study. Additionally, the administration of Kyn significantly increased the infarct volume, and this effect was not observed with the co-administration of a pharmacological AHR antagonist or in AHR−/− mice22. These previously reported findings confirm the detrimental effects of the Kyn-AHR pathway in acute stroke, and, more importantly, the post-stroke responsiveness of the brain AHR pathway to peripherally sourced molecules (Kyn or pharmacological inhibitors22). Our results show that post-stroke changes in the gut microbiota lead to the loss of microbially derived indole-based AHR ligands. These compounds have a similar role as pharmacological inhibitors of AHR due to their partial agonistic properties. Thus, we hypothesized that increasing the concentration of microbiota-dependent indoles in the brain will ameliorate the deleterious effects of Kyn-based AHR activation by providing a competitive beneficial ligand. As mice with a global deletion of AHR (AHR−/− KO) or mice with an MG-specific deletion of AHR eliminate the cellular receptor for both indole- and Kyn-based AHR ligands, these models are not appropriate for testing our overall hypothesis.
We began by demonstrating significant expression (known to be an indirect measure of activation) of AHR in MG and a significant reduction in the plasma levels of microbiota-derived indole-based AHR ligands after human stroke. Increased gut dysbiosis reduces the levels of microbiota-derived indole derivatives48,49. In contrast with the decreased expressions of indole-based AHR ligands following stroke, increasing brain and plasma concentrations of host-derived Kyn-based AHR ligands have been described22. However, large datasets reporting alterations in both Kyn- and indole-based AHR ligands after human stroke are currently unavailable. This clinical data gap is of high translational relevance in stroke immunology, as multiple studies have reported that indole-based molecules have a significantly higher affinity for human than for mouse AHR at physiological concentrations19,50.
IPA is not detected in GF mouse plasma29, but similar investigations on brain tissue had not been performed before this study. We identified two indole-based AHR ligands (IPA and IAld) that were undetectable in the brain of GF mice. Additionally, we showed that known AHR ligand producers decreased in the gut microbiota with aging and after stroke in aged WT mice, which is consistent with the existing microbiome reports in the literature. For example, the relative abundance of Bifidobacterium significantly decreases in fecal samples collected 24 h after acute ischemic stroke51; Bifidobacterium abundance and diversity decreases with age52,53,54; and Bifidobacterium and Lactobacillus abundance decreases after experimental stroke in rats55.
These findings prompted us to investigate the effects of post-stroke treatment with indole-based AHR ligands in both GF and aged WT mice after stroke. We evaluated multiple time points after stroke and observed a stepwise reduction of IPA and IAld in both plasma and brain, suggesting a loss of the bottom-up supply (produced by the gut microbiota and detected in the plasma and brain) of these microbiota-dependent AHR ligands. This reduction in indole-based AHR ligands is likely due to stroke-induced dysbiosis of the gut microbiota, which has been extensively reported by our group and others56,57,58. Moreover, we substantiated the previously reported22 increase in brain Kyn levels as a representative molecule of host-derived AHR ligands produced via the Kyn pathway16.
Our ex vivo results indicated a ligand-specific effect of Kyn and indole-based AHR ligands on MG survival after OGD/R, suggesting a dichotomy between host-derived Kyn and microbiota-derived indoles in the context of cellular stress such as OGD/R. We examined the beneficial effect of indole-based AHR ligands on MG survival via in vivo administration of indole-based AHR ligands, followed by ex vivo OGD/R of the sorted MG. Our ex vivo data demonstrated the ligand specificity of AHR, which has been extensively reported in other disease conditions9,18. Our results showed that the addition of Kyn to Trp-depleted medium was detrimental, whereas the addition of IPA and IAld cocktail to Trp-depleted medium was not. We hypothesized that indole-based AHR ligands exert their positive effects primarily as partial agonists of AHR, outcompeting the detrimental effects of Kyn and other host-derived AHR ligands. Consequently, in the absence of Trp and Kyn, they may not exhibit an independent beneficial effect. The pool of endogenous AHR ligands can be viewed as a balance between host- and microbiota-derived molecules, disturbing what can be a source of detrimental immune responses after stroke26,59.
As a final proof of principle, we used cohorts of GF and aged WT stroke mice (Figs. 4 and 5). Our results showed that the post-stroke treatment of GF mice with a cocktail of IPA and IAld significantly increased the expression of MG AHR, MHC-II, and CD80. The surface expression of MHC-II and CD80, regulated by the transcriptional activity of AHR9,19, are the direct markers of antigen processing and the co-stimulatory function of MG and other immune cells. Our results indicate concerted regulation of the surface phenotype by MG, brain lymphocytes, and infiltrating monocytes 24 h after stroke in GF stroke mice treated with IPA and IAld relative to GF stroke mice treated with vehicle. Finally, using aged WT mice as a translational preclinical model of stroke, we showed that infarct volume and neurological deficits were significantly reduced by post-stroke treatment with IPA and IAld.
The interplay between host-derived and microbiota-derived ligands and the activation of AHR in microglia and other immune cells is a critical factor in the inflammatory response following stroke. Due to the highly ligand-specific nature of AHR immunobiology and partial agonistic behavior of these endogenous ligands, a cooperative/competitive type response can be expected from AHR in the presence of a balanced pool of ligands. As demonstrated here, the balance of AHR ligands is disturbed by an immediate post-stroke decrease in the microbiota-dependent supply of indoles and an increase in host-derived Kyn, which have detrimental effects after stroke.
Currently, there are no reports in the literature evaluating the regulatory role of specific microbiota or AHR metabolites in aged or GF mice after stroke. Vicentini et al.60 showed that antibiotic-induced behavioral deficits in despair behavior can be restored by microbial recolonization or AHR activation with an AHR agonist in young male mice. Previous studies have used bacterial transgenics to eliminate tryptophanase-positive bacteria to alter AHR ligand profiles and concluded that both tryptophanase-positive and negative bacteria are capable of converting tryptophan to indole-based molecules9. Therefore, the use of antibiotic treatments to specifically deplete biome-derived AHR ligands is not feasible, as no combination of currently available antibiotics can selectively eliminate tryptophan metabolizers. Another approach used commonly in microbiome research is fecal microbiota transplantation (FMT). Wei et al.61 showed that FMT from healthy young mice to recipient mice with colitis alleviated disease severity by altering the intestinal microbiota and metabolite composition and upregulated the expression of AHR. We have previously shown that FMT of young donor biome into aged mice improves outcomes after stroke, if given prior to or several days after stroke58. However, the use of FMT is also not without caveats as there are many potential confounders (such as the introduction of viruses and fungi). Therefore, we chose to use a more selective approach (restoring specific metabolites in GF mice models) to better assess the functional role of metabolites such as indoles to address the central question of ligand specificity of AHR in the context of stroke.
Singh et al. have shown that GF mice have a larger infarct volume after stroke than conventional SPF or Ex-GF mice (GF mice with microbiota recolonization) (J Cereb Blood Flow Metab. 2018 Aug; 38(8): 1293–1298). They did not profile metabolites such as indoles in the same mice. However, this study demonstrated the important interactions between the microbiota and immune cells (primarily T-cells) in stroke. GF mice have compromised immune cell maturation and suboptimal performance across all behavioral tests examined58. Specific bacterial candidates that potentially regulate the AHR pathway will be assessed in future studies using selective recolonization experiments in GF mice. However, our study did not focus on specific bacterial strain identification and replacement but rather the metabolic products produced by the gut microbiota and their effect on AHR activation and stroke outcomes.
We provide robust experimental evidence that restoring microbiome-derived metabolites is beneficial in experimental stroke, there are some limitations that should be recognized. Our results do not exclude the indirect beneficial effects of the indole-derived AHR ligands on post-stroke neuroinflammation. In this investigation, we used GF mice, which have been reported to have larger infarcts than conventional SPF or Ex-GF mice (GF mice with microbiota recolonization). However, the study used female GF mice and a different stroke model (permanent distal MCAO), which produces a much smaller infarct than that in the transient MCAO model used here. Our studies showed a reduction in brain weight and volume in male GF mice but no differences in acute NDS scores. GF mice have poor immune cell maturation, altered responses to inflammatory challenges, and behavioral changes at baseline. GF mice also show sex differences in microglial gene signatures. In adult GF mice, 433 genes were differentially expressed in MG from female GF mice relative to only 26 in male GF mice, and several of the downregulated genes altered in GF female MG were related to inflammation58. In addition, it has been reported that the interactions between AHR and estrogen receptors can occur62. We have also recently found that estradiol has profound effects on the colonic epithelium and the microbiome composition after stroke62. To avoid these confounders in this early work, we restricted our murine studies to male mice. However, understanding sex differences in the response to stroke is an important area, and studies are ongoing to determine sex differences in AHR pathways.
We examined the direct effects of indole supplementation in aged WT mice, a more translationally relevant approach. We found that administration of indole-based AHR ligands (indole-3-carboxaldehyde and indole-3-propionate) reduced neurological deficits and infarct size in aged WT mice 24 h after stroke. Further experiments are needed to investigate the role of immune crosstalk between MG and astrocytes, MG and lymphocytes, and MG and endothelial cells under the influence of microbiota-derived indole-based AHR ligands. Additional studies on long-term stroke outcomes and behavioral differences in stroke mice of both sexes treated with indole-based AHR ligands are warranted. Specific bacterial candidates that potentially regulate the AHR pathway will be investigated in future studies involving recolonization experiments in GF mice.
Our results demonstrate that restoring a balanced pool of host- and microbiota-derived molecules via post-stroke treatment with indole-based AHR ligands to compete with Kyn-based activation of AHR provides a net beneficial effect and improves post-stroke outcomes.
Methods
Ethics and tissue acquisition
The use of postmortem human samples was deemed exempt by the Committee for the Protection of Human Subjects at the University of Texas Health Science Center at Houston (HSC-MS-22-0982 – UTHealth Neuropathology Core). Postmortem human brain tissue was obtained from the Neuropathology Core of the Alzheimer’s Disease Research Center at the University of Pittsburgh.
For the use of human samples for the metabolomics study, the protocol was approved by the Institutional Review Board of the University of Texas Health Science Center at Houston, Houston, Texas, USA (IRB No: HSC-MS-17-0452, HSC-MS-20-0780). All participants provided written informed consent for the study and subsequent use of the stored samples. The exclusion criteria for our study were patients aged <18 years, patients with acute or chronic anemia requiring transfusion of ≥1 unit of packed red blood cells, patients with active cancer, and prisoners.
Animal experiments were performed according to the guidelines of the NIH. All experiments were approved by the Institutional Animal Care and Use Committees of the University of Texas Health Science Center at Houston (UTHSCH) and the Baylor College of Medicine.
All animal procedures were performed at an Association for Assessment and Accreditation of Laboratory Animal Care-accredited (AAALAC-accredited) facility and approved by the Animal Welfare Committee of the University of Texas Health Science Center in Houston, TX, USA.
Immunohistochemistry of human brain tissue samples
Supplementary Table S1 provides the population and clinical characteristics of stroke and control cohorts for postmortem brain tissue samples. Immunohistochemistry of formalin-fixed, paraffin-embedded human brain sections was performed as described previously63,64. After deparaffinization, the sections were subjected to a heat-induced antigen retrieval process; the samples were heated (citrate buffer, pH 6.0; 99 °C; 20 min) and blocked with blocking buffer (5% donkey normal serum, 1% bovine serum albumin with 0.3% Triton X-100 in phosphate buffered saline [PBS]) for 1 h at room temperature. The sections were incubated with primary antibodies followed by detection using Alexa Fluor 488-conjugated donkey anti-rabbit and Alexa Fluor 647-conjugated donkey anti-mouse (1:200, Jackson ImmunoResearch).
Imaging and quantification of MG AHR expression in human brain samples
Imaging was performed with a Leica THUNDER Imager DMi8 using a 40x lens. Five images from both the infarct and peri-infarct regions from patients with stroke and five images from the normal cortical region of normal (control) participants were captured. The infarct and peri-infarct regions were stained using hematoxylin and eosin and confirmed by a neuropathologist. Images were analyzed using the ImageJ software by an investigator blinded to the case characteristics. All images were corrected for background noise, and consistent fluorescence intensity was maintained. Quantities of AHR- and Iba-1-positive cells were obtained using individual regions of interest (ROIs) within green- or red-specific channels, respectively. The number of AHR/Iba-1 double-positive cells was determined using overlay images consisting of both channels, and counts were based on the reselection of double-positive ROIs. Data are expressed as mean values ± standard error of mean.
Human MG cell line
HMC3 cells were maintained in Eagle’s Minimum Essential Medium (EMEM) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin in a humidified incubator at 37 °C and 5% CO2.
OGD/R
For OGD studies of human MG, cells were washed with PBS and incubated with complete media (normoxia condition) or subjected to OGD/R (117 mM NaCl, 5.3 mM KCl, 1.8 mM CaCl2, 0.8 mM MgSO4, 26 mM NaHCO3, and 1.17 mM NaH2PO4, pH 7.4) in a hypoxia incubator chamber for 2 h at 37 °C. The cells were subsequently washed with supplemented EMEM and incubated in the same medium for 4 h65. They were collected in QIZOL reagent for further analysis by qPCR66. Sorted MG from aged (18 months old) male WT mice were seeded into 6-well culture plates at a density of 1 × 104 cells/well. The OGD/R assay was performed as described previously67. Briefly, the culture medium was replaced with glucose-free Dulbecco’s Modified Eagle Medium A1443001 (Thermo Fisher Scientific), and the plates were placed in a sealed chamber, followed by expiration of oxygen for 10 min by flowing a mixture of 95% N2 and 5% CO2 (Airgas-Southwest Inc.) continuously at a low rate. The sealed chamber was transferred to a 37 °C incubator for 4 h to mimic OGD/R after clamping the inlet and outlet. During OGD/R, the O2 levels dropped to <2% after 2 h and <1% at 4 h, as shown by the change in the color of the BD-271051 anaerobic indicator strips from blue (aerobic) to white (anaerobic). The medium was changed to normal Dulbecco’s Modified Eagle Medium 31053036 (Thermo Fisher Scientific), and the cells were reperfused to a normoxic atmosphere by incubation under 95% oxygen and 5% CO2 at 37 °C for 2 h.
Consideration of sex as a biological variable
We restricted our murine studies to male mice. Both male and female samples were included in the human studies, and all human female samples included in this study were from postmenopausal women.
WT mice
C57BL/6 male mice were obtained from the National Institute on Aging (NIA), Charles River, or Jackson Laboratories. Young (3 months) and aged (18 months) mice were used in this study. All mice were procured at least 8 weeks before the experiment date, and they aged in-house to the appropriate study time points to stabilize the microbiome composition.
All animals, except GF mice, were group-housed in Tecniplast individually ventilated cage racks; fed a commercially available irradiated, balanced mouse chow (no. 5058, LabDiet, St Louis, MO, USA); and provided corncob bedding. The animal rooms were maintained at 21–23 °C under a 12–12-h light–dark cycle. All GF animals were maintained under specific pathogen-free conditions.
GF mice
GF C57BL/6-GF male mice (3 months old) were obtained from the Baylor College of Medicine Gnotobiotic Rodent Facility. They were shipped in autoclaved shipping crates to UTHSCH. The crates were assembled, bedded with Alpha-Dri bedding, and autoclaved with lids closed. Mice were removed from the isolator in the gnotobiotic facility using sterile transfer bags, housed in shipping crates in a biosafety cabinet, and immediately transported. The two medical schools are only a few hundred yards apart, which precludes lengthy and stressful travel. To assess the GF status of the mice upon arrival at the UTHSCH, fecal samples were collected to confirm the absence of bacteria in the gut. An internal standard was serially diluted, and the copy number of the 16S rRNA gene in the feces from each transfer crate was analyzed using qPCR. On confirming the absence of 16S rRNA gene in the feces in any of the transfer crates, the mice were labeled as GF. Once the aseptic crates were opened at the UTHSCH, the mice were transferred to autoclaved cages, provided with autoclaved chow and water, and maintained in a conventional animal holding room for surgery on the same day. All GF mice were euthanized 24 h after surgery.
Mouse MCAO
Transient focal ischemia was induced under isoflurane anesthesia in young, aged, or GF mice for 60 min by occlusion of the right middle cerebral artery according to a previously described protocol68. Body temperature was maintained at 37.0 ± 1.0 °C throughout the surgery by an automated temperature control feedback system (TC1000, mouse, CWE Inc., USA). A midline ventral neck incision was made, and unilateral MCAO was performed by inserting a Doccol monofilament (Doccol Corp, Redlands, CA, USA) into the right internal carotid artery. The animals were re-anesthetized 1 h after ischemia, and reperfusion was established by withdrawal of the monofilament. The mice were placed in recovery cages and euthanized 24 h after reperfusion. Sham controls were subjected to the same procedure, except that the sutures were not introduced into the middle cerebral artery. Animals were randomly assigned to the stroke and sham surgery groups, individually housed in their recovery cages for the first 2 h after surgery, and returned to group housing. All analyses were performed by investigators who were blinded to the surgical conditions. Sham and stroke mice were housed separately to minimize the effects of microbiota contamination in the experimental groups. Five mice were excluded from the study because of death during MCAO surgery (n = 2), subarachnoid hemorrhage (n = 1), or no significant intra-ischemic neurological deficits (NDS = 0) after stroke (n = 2).
Mouse NDS
NDS was assessed immediately and 24 h post-stroke. The Bederson score system (0 [normal] to 4 [most severe]) was used56. Briefly, scores were assigned according to the following criteria: 0, no deficit; 1, forelimb weakness and torso turning to the ipsilateral side when the animals were held by the tail; 2, circling to the affected side; 3, inability to bear weight on the affected side; and 4, no spontaneous locomotor activity or barrel rolling. Animals with NDS scores of 0 or 4 after reperfusion were excluded.
Mouse infarct volume assessment
Mice were euthanized 24 h post-MCAO for infarct volume analysis, as previously described69 (with modifications). Briefly, the mice were transcardially perfused with ice-cold PBS. The brains were extracted and submerged in a graduated cylinder containing PBS for precise volumetric measurement. After removing the olfactory bulb and cerebellum, the brains were sectioned into 5 slices of 1.5 mm thickness using a brain matrix. The slices were immersed in 1.5% 2,3,5-triphenyl-tetrazolium in PBS for 10 min at 37 °C and fixed in 4% paraformaldehyde in PBS. The slices were photographed, and the infarct volumes (corrected for edema) were analyzed using Sigma Scan Pro 5 (SPSS Inc.).
Reverse transcription-qPCR
To quantify the relative mRNA expression levels of AHR and cytochrome P450 1B1 (Cyp1b1), mRNA was extracted from HMC3 cultured cells using the miRNeasy® mini kit (QIAGEN) and 2 µg was reverse-transcribed to cDNA using the RevertAid H minus First Strand cDNA Synthesis Kit (Thermo Fisher, USA). Reverse RT-qPCR was performed using a Quant Studio 3 Real-Time PCR System (Applied Biosystems, USA). The RT-PCR reaction mix (adjusted with sterile distilled water to a total volume of 20 μL) contained 1 μL template DNA, 10 μL TaqMan Fast advanced master mix (Thermo Fisher, USA), and 1 μL of the respective primer probes (AHR (Hs00169233_m1), Cyp1b1 (Hs02382916_s1), and glyceraldehyde 3-phosphate dehydrogenase (Hs02758991_g1)). The relative mRNA expression levels of the target gene were normalized to the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase used as a ref. 57,70. The following formula was used:
Treatment with an indole cocktail in mice
IPA and IAld (indole-3-propionic acid and indole-3-aldehyde [indole-3-carboxyaldehyde], respectively; Sigma-Aldrich, Cat #: 220027 and 129445) were dissolved in warm (37 °C) corn oil with <1% DMSO to a final concentration of 0.1 nmol/g of body weight (approximated based on the tissue metabolomics data obtained from WT mice) for oral gavage and were administered as a single dose once at 3 h and at 8 h post-stroke (data included in Figs. 4 and 5). The total volume of dissolved cocktail administered orally was 200 µl/animal. The vehicle solution was identical in content but lacked indole-based molecules.
Single-cell data acquisition and re-processing
The single-cell RNA seq dataset (GSE174574) was retrieved and obtained from the Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/). It included ipsilateral cerebral cortical samples from mice 24 h post-MCAO injury, as well as sham control samples71. This dataset contained three MCAO and three sham cerebral cortical samples of mice. For quality control, three criteria were applied to screen cells and genes in each sample: (1) cells with a minimum of 200 genes profiled were selected for further analysis; (2) to minimize mitochondrial gene contamination, the cells were filtered based on the mitochondrial gene mapping ratio, and a maximum proportion of 20% was applied; and (3) cells with >5000 unique molecular identifiers per cell were excluded. Data integration was performed in the principal component analysis space using SCT normalization and the Harmony algorithm for batch removal (version 0.1.0). The RunUMAP function in the Seurat package was used for dimension reduction72. Microglia and endothelial cells were annotated at the level of cell clusters using the corresponding markers hexosaminidase subunit beta (Hexb) and integral membrane protein 2A (Itm2a). Alterations in the AHR and its downstream gene signaling were visualized using violin plots by comparing the MCAO and sham groups. Significance was determined using the Wilcoxon test in GraphPad Prism.
Flow cytometry
Blood (mouse)
Blood was drawn via a cardiac puncture using heparinized needles. Red blood cell lysis was achieved by two consecutive 10-min incubations with Tris-ammonium chloride (Stem Cell Technologies)73,74.
Brain (mouse)
The mice were transcardially perfused with 60 mL of cold sterile PBS before aseptic removal of the spleen and brain tissues. Brain tissue was placed in complete Roswell Park Memorial Institute medium 1640 (Lonza) and mechanically and enzymatically digested in Collagenase/Dispase (1 mg/mL) and DNase (10 mg/mL; Roche Diagnostics) for 45 min at 37 °C. The cell suspension was filtered through a 70-μm filter. Leukocytes were harvested from the interphase of a 70 to 30% Percoll gradient10,58.
Skull bone marrow (mouse)
To isolate the skull bone marrow, the meninges were gently peeled from the skull cup. The skull was cut into small pieces using sterile scissors and mechanically dissociated in PBS, followed by filtration through a 70-µm cell strainer. After centrifugation at 500 × g for 5 min, red blood cells were removed via the addition of 1 mL of ammonium chloride (Stem Cell Technologies, Cat #07850) lysis buffer for 2 min at room temperature, followed by centrifugation at 500 × g for 5 min and resuspension of the cell pellet in PBS until use.
Spleen (mouse)
Spleen (whole) tissue was removed after PBS perfusion and passed through a 70-μm strainer and ammonium chloride solution (Stem Cell Technologies, Cat #07850) in a 9:1 ratio for red blood cell lysis.
Surface and intracellular staining
Cells were washed and blocked with mouse FcR Block (BioLegend) before staining with antibodies pre-conjugated with the following fluorophores (BioLegend): CD45-eF450 (eBioscience, Cat#: 48-0451-82, Lot: 2005853), CD11b-APC (BioLegend, Cat#: 101212, Lot: B279418), Ly6CPerCP-Cy5.5 (BioLegend, Cat#: 128011, Lot: 292026), Tmem119-PE-Cy7 (eBioscience, Cat#: 25-6119-82, Lot: 2210260), P2RY12-PE (BioLegend, Cat#: 848003, B298459), and MHCII-APC-Fire750 (BioLegend, Cat#: 107652, Lot: B301025) pre-conjugated antibodies and Zombie Aqua (BioLegend, Cat#: 423102, Lot: B300004). For intracellular staining, cells were fixed and permeabilized (BioLegend, Cyto-Fast Fix/Perm kit, Cat#: 750000133) following the manufacturer’s protocol. A pre-conjugated AHR-BV421 antibody (BD Horizon, Cat# 565791) was used for intracellular staining. Data were acquired using a Cytoflex-S cytometer (Beckman Coulter) and analyzed using FlowJo software (Treestar Inc.). No less than 100,000 events were recorded for each sample or tissue. Cell type-matched fluorescence minus one and unstained controls were used as gating strategies. Uniform Manifold Approximation and Projection (UMAP) plots were generated in FlowJo using the DownSample v3 plug-in (3000 Live CD45+ cells per sample), followed by the UMAP v3.1 algorithm on all uncompensated parameters (except viability) using Euclidean distances, 15 nearest neighbors, a minimum distance of 0.5, and two components. Following the UMAP analysis, the Phenograph v3.0 plug-in was used with a K-value of 174 (recommended by FlowJo) to generate 20 clusters of immune populations. Surface expression heatmaps for CD80, MHC-II, CD11b, P2RY12, and Tmem119 for all clusters are provided in Supplementary Fig. S6. The identification markers for each cluster are shown in Supplementary Fig. S7.
Cell sorting
Single-cell suspension and surface staining were performed as described in the preceding section. After viability and singlet selection, MG gated as Live Tmem119+ (verified to be CD45intCD11b+) was sorted under an aseptic hood from a single-cell suspension prepared from naive aged male brains (full brains, n = 8) using BD FACSMelody. Sorting was performed from the tube directly into a 96-well plate for ex vivo experiments to preserve the cell counts. The cells were washed with PBS, stained for surface markers and viability after OGD/R, and analyzed using flow cytometry.
Mouse luminal content collection and 16S rRNA sequencing
Intestinal contents were collected from mice at the same location and time of the day for all groups and stored in sterile tubes at −80 °C until analyzed, as described previously56. The mouse cecum serves as a storage for intestinal contents in which slower and prolonged biochemical reactions take place. Specifically, metabolic reactions of dietary tryptophan and the fermentation activity of indigestible fibers are known to be abundant in the mouse cecum, and microbial AHR ligands are products of microbial metabolism of tryptophan and fermentation75.
The bacterial taxa in each fecal sample were analyzed by amplifying the V4 to V5 hypervariable regions of the 16S ribosomal RNA (rRNA) gene using high-throughput sequence analysis performed by the Alkek Center for Metagenomics and Microbiome Research (Baylor College of Medicine, Houston, TX, USA). The workflow specifications included the following: Nucleic Acid Extraction Protocol: MoBIO PowerSoil v3.4; Amplification Protocols: Illumina 16 Sv4 v1.2; Sequencing Protocol: Illumina MisSeq v2 2 × 250 v1.8 (Illumina, San Diego, CA, USA)76; and Analytics Pipeline: CMMR 16 Sv4 v1.0. Quality-filtered 16S rRNA sequences were clustered into OTUs with 97% similarity by closed-reference picking using the UCLUST algorithm and GreenGenes reference database (v13.5), as implemented in Quantitative Insights into Microbial Ecology (QIIME versions 1.6 and 1.7)76,77,78,79. Sequences were checked for chimeras using ChimeraSlayer with standard options, as implemented in QIIME. Sequences that were not clustered were identified using the Ribosomal Database Project to the lowest possible taxonomic level80.
Human and mouse metabolomics analysis by liquid chromatography coupled with high-resolution mass spectrometry
Plasma samples from the patients with stroke and controls were used for this study. The demographic characteristics of the human plasma samples are included in Supplementary Table S2. To determine the relative concentrations of Trp metabolites in the brain tissue and plasma samples, extracts were prepared and analyzed by LC-HRMS. For tissue samples, 100 mg of tissue was pulverized in liquid nitrogen and homogenized using a Precellys Tissue Homogenizer. Plasma metabolites were extracted from 100 uL of samples using 0.5 mL of ice-cold 50/50 (v/v) methanol/acetonitrile followed by 0.5 mL of 0.1% formic acid in 50/50 (v/v) acetonitrile/water. Extracts were centrifuged at 17,000 × g for 5 min at 4 °C, and supernatants were transferred to clean tubes, followed by drying by evaporation under nitrogen. The samples were reconstituted in 50/50 (v/v) methanol/water, of which, 10 μL was injected into a Thermo Vanquish liquid chromatography (LC) system containing a Waters XSELECT HSS T3 2.1 × 150 mm column with a particle size of 2.5 µm. Mobile phase A (MPA) consisted of 0.1% formic acid in water, and mobile phase B (MPB) was 100% methanol. The flow rate was 200 µL/min (at 35 °C), and the gradient conditions were as follows: initial 5% MPB increased to 95% MPB at 15 min, held at 95% MPB for 5 min, returned to initial conditions, and equilibrated for 5 min. The total run time was 25 min. Data were acquired using a Thermo Orbitrap Fusion Tribrid mass spectrometer in ESI-positive and negative ionization modes at a resolution of 240,000 with full scan mode. Raw data files were imported to the Thermo Trace Finder software for final analysis. The relative concentration of each compound was normalized to the weight of the tissue samples and volume of the plasma samples.
Signals for Indole-3-propionate and Indole-3-carboxylaldehyde (ChemID 100001083 and 100002185) were extracted, merged, batch-corrected, log-normalized, and scaled from the patients with stroke (n = 60) and healthy controls (n = 64) using the R package metabolomicsR81. Violin-box plots were used to visualize differences between these two metabolites in the patients with stroke and age-matched controls without stroke. For patients with stroke, linear regression of the metabolite signals and the NIH stroke scale scores was performed and confidence intervals were obtained using R package ggplot2. R software (version 4.2.2, higher) was used for the metabolic analysis.
Statistical analyses
Statistical analysis was performed using two-way analysis of variance with sex, age, and their interactions as factors. The main effects were tested, followed by a post hoc analysis with all related p values adjusted using Sidak’s method for multiple comparisons. Statistical correlations between the averaged behavioral variables and cytokine concentrations were determined in each experimental scenario using the Spearman method. Statistical significance was considered at p < 0.05. The *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001 conventions were used in the presented figures. All statistical analyses were performed using GraphPad Prism 7.
ROSALIND® nanostring gene expression method and analysis
Data was analyzed by ROSALIND® (https://rosalind.bio/) with a HyperScale architecture developed by ROSALIND, Inc. (San Diego, CA, USA). Read distribution percentages, violin plots, identity heat maps, and sample MDS plots were generated as part of the quality control step. Normalization, fold changes, and p values were calculated using the criteria provided by Nanostring. ROSALIND follows the nCounter Advanced Analysis protocol for dividing counts within a lane by the geometric mean of the normalizer probes from the same lane. Housekeeping probes used for normalization were selected based on the geNorm algorithm, as implemented in the NormqPCR R library82. The abundances of various cell populations were calculated using ROSALIND with the NanoString Cell Type Profiling Module. ROSALIND performs a filtering of Cell Type Profiling results to include those with a p values of ≥0.05.
Differential gene expression analysis was performed using the Student’s t test on log2-transformed and normalized gene expression. p values were adjusted using the Benjamini–Hochberg method to estimate false discovery rates. Differentially expressed genes between groups were visualized using a volcano plot in the R package EnhancedVolcano. The heatmap of the AHR genes was plotted using the R package ComplexHeatmap83. R software (version 4.2.2 and higher) was used for gene expression analysis.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The “minimal data set” necessary to interpret, replicate and build on the findings reported in the paper can be requested at any time by contacting the corresponding authors. Source data for each figure are provided with this paper under the name “source raw data file” and can be accessed using the link https://doi.org/10.6084/m9.figshare.26357611. Metabolomics data set is deposited in Metabolomics workbench under the ID: 4638. Single-cell RNA sequencing data sets (GSE174574) was obtained from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) and reused in Supplementary Fig. 10.
References
Virani, S. S. et al. Heart disease and stroke statistics—2020 update: a report from the American Heart Association. Circulation 141, e139–e596 (2020).
Benjamin, E. J. et al. Heart disease and stroke statistics—2018 update: a report from the American Heart Association. Circulation 137, e67–e492 (2018).
Powers, W. J. et al. Guidelines for the early management of patients with acute ischemic stroke: 2019 update to the 2018 guidelines for the early management of acute ischemic stroke: a guideline for healthcare professionals from the American Heart Association/American Stroke Association. Stroke 50, e344–e418 (2019).
Rinaldo, L. et al. Racial and ethnic disparities in the utilization of thrombectomy for acute stroke. Stroke 50, 2428–2432 (2019).
Murray, C. J. L. et al. Global, regional, and national disability-adjusted life years (DALYs) for 306 diseases and injuries and healthy life expectancy (HALE) for 188 countries, 1990–2013: quantifying the epidemiological transition. Lancet 386, 2145–2191 (2015).
Iadecola, C., Buckwalter, M. S. & Anrather, J. Immune responses to stroke: mechanisms, modulation, and therapeutic potential. J. Clin. Invest. 130, 2777–2788 (2020).
Honarpisheh, P., Bryan, R. M. & McCullough, L. D. Aging microbiota-gut-brain axis in stroke risk and outcome. Circ. Res. 130, 1112–1144 (2022).
Gutiérrez-Vázquez, C. & Quintana, F. J. Regulation of the immune response by the aryl hydrocarbon receptor. Immunity 48, 19–33 (2018).
Rothhammer, V. & Quintana, F. J. The aryl hydrocarbon receptor: an environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 19, 184–197 (2019).
Ritzel, R. M. et al. Aging alters the immunological response to ischemic stroke. Acta Neuropathol. 136, 89–110 (2018).
Koduru, L. et al. Systems biology of gut microbiota-human receptor interactions: toward anti-inflammatory probiotics. Front. Microbiol. 13, 846555 (2022).
Dahlem, C. et al. AHR signaling interacting with nutritional factors regulating the expression of markers in vascular inflammation and atherogenesis. Int. J. Mol. Sci. 21, 8287 (2020).
Paeslack, N. et al. Microbiota-derived tryptophan metabolites in vascular inflammation and cardiovascular disease. Amino Acids 54, 1339–1356 (2022).
Tsuji, A. et al. The tryptophan and kynurenine pathway involved in the development of immune-related diseases. Int. J. Mol. Sci. 24, 5742 (2023).
Kindler, J. et al. Dysregulation of kynurenine metabolism is related to proinflammatory cytokines, attention, and prefrontal cortex volume in schizophrenia. Mol. Psychiatry 25, 2860–2872 (2020).
Colpo, G. D., Venna, V. R., McCullough, L. D. & Teixeira, A. L. Systematic review on the involvement of the kynurenine pathway in stroke: pre-clinical and clinical evidence. Front. Neurol. 10, 778 (2019).
Roager, H. M. & Licht, T. R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 9, 3294 (2018).
Murray, I. A. & Perdew, G. H. How ah receptor ligand specificity became important in understanding its physiological function. Int. J. Mol. Sci. 21, 9614 (2020).
Hubbard, T. D. et al. Adaptation of the human aryl hydrocarbon receptor to sense microbiota-derived indoles. Sci. Rep. 5, 12689 (2015).
Lee, Y. H. et al. Aryl hydrocarbon receptor mediates both proinflammatory and anti‐inflammatory effects in lipopolysaccharide‐activated microglia. Glia 63, 1138–1154 (2015).
Cha, M. H. et al. Metabolomic analysis of clinical plasma from cerebral infarction patients presenting with blood stasis. Evid. Based Complement. Altern. Med. 2015, 453423 (2015).
Cuartero, M. I. et al. L-kynurenine/aryl hydrocarbon receptor pathway mediates brain damage after experimental stroke. Circulation 130, 2040–2051 (2014).
Kasner, S. E. Clinical interpretation and use of stroke scales. Lancet Neurol. 5, 603–612 (2006).
Boopathi, S. et al. Gut Enterobacteriaceae and uraemic toxins—perpetrators for ageing. Exp. Gerontol. 173, 112088 (2023).
Quintana, F. J. & Sherr, D. H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 65, 1148–1161 (2013).
Esser, C. et al. Old receptor, new tricks-the ever-expanding universe of aryl hydrocarbon receptor functions. Report from the 4th AHR Meeting. Int. J. Mol. Sci. 19, 3603 (2018).
Denison, M. S. & Nagy, S. R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 43, 309–334 (2003).
Lee, J.-H., Wood, T. K. & Lee, J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol. 23, 707–718 (2015).
Wikoff, W. R. et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl Acad. Sci. USA 106, 3698–3703 (2009).
Crumeyrolle-Arias, M., Tournaire, M.-C., Rabot, S., Malpaux, B. & Thiéry, J.-C. 5‐hydroxyoxindole, an indole metabolite, is present at high concentrations in brain. J. Neurosci. Res. 86, 202–207 (2008).
Obata, Y. et al. Neuronal programming by microbiota regulates intestinal physiology. Nature 578, 284–289 (2020).
McMenamy, R. H. Binding of indole analogues to human serum albumin. Effects of fatty acids. J. Biol. Chem. 240, 4235–4243 (1965).
Madras, B. K., Cohen, E. L., Messing, R., Munro, H. N. & Wurtman, R. J. Relevance of free tryptophan in serum to tissue tryptophan concentrations. Metabolism 23, 1107–1116 (1974).
Hargreaves, K. M. & Pardridge, W. M. Neutral amino acid transport at the human blood-brain barrier. J. Biol. Chem. 263, 19392–19397 (1988).
Ruddick, J. P. et al. Tryptophan metabolism in the central nervous system: medical implications. Expert Rev. Mol. Med. 8, 1–27 (2006).
Ball, H. J., Jusof, F. F., Bakmiwewa, S. M., Hunt, N. H. & Yuasa, H. J. Tryptophan-catabolizing enzymes—party of three. Front. Immunol. 5, 485 (2014).
Rubio, V. Y., Cagmat, J. G., Wang, G. P., Yost, R. A. & Garrett, T. J. Analysis of tryptophan metabolites in serum using Wide-Isolation Strategies for UHPLC–HRMS/MS. Anal. Chem. 92, 2550–2557 (2020).
Clarke, G. et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 18, 666–673 (2013).
Dvořák, Z., Poulíková, K. & Mani, S. Indole scaffolds as a promising class of the aryl hydrocarbon receptor ligands. Eur. J. Med. Chem. 215, 113231 (2021).
O’Mahony, S. M., Clarke, G., Borre, Y. E., Dinan, T. G. & Cryan, J. F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 277, 32–48 (2015).
Hubbard, T. D., Murray, I. A. & Perdew, G. H. Indole and tryptophan metabolism: endogenous and dietary routes to aH receptor activation. Drug Metab. Dispos. 43, 1522–1535 (2015).
Vyhlídalová, B. et al. Gut microbial catabolites of tryptophan are ligands and agonists of the aryl hydrocarbon receptor: a detailed characterization. Int. J. Mol. Sci. 21, 2614 (2020).
Bisson, W. H. et al. Modeling of the aryl hydrocarbon receptor (AHR) ligand binding domain and its utility in virtual ligand screening to predict new AHR ligands. J. Med. Chem. 52, 5635–5641 (2009).
Schroeder, J. C. et al. The uremic toxin 3-Indoxyl sulfate is a potent endogenous agonist for the human aryl hydrocarbon receptor. Biochemistry 49, 393–400 (2010).
Barreto, F. C. et al. Serum indoxyl sulfate is associated with vascular disease and mortality in chronic kidney disease patients. Clin. J. Am. Soc. Nephrol. 4, 1551–1558 (2009).
Sallée, M. et al. The aryl hydrocarbon receptor-activating effect of uremic toxins from tryptophan metabolism: a new concept to understand cardiovascular complications of chronic kidney disease. Toxins 6, 934–949 (2014).
Rothhammer, V. et al. Microglial control of astrocytes in response to microbial metabolites. Nature 557, 724–728 (2018).
Niu, B. et al. The therapeutic potential of dietary intervention: based on the mechanism of a tryptophan derivative-indole propionic acid on metabolic disorders. Crit. Rev. Food Sci. Nutr. 8, 1–20 (2024).
Konopelski, P. & Mogilnicka, I. Biological effects of Indole-3-Propionic acid, a gut microbiota-derived metabolite, and its precursor tryptophan in mammals’ health and disease. Int. J. Mol. Sci. 23, 1222 (2022).
Dong, F. et al. Intestinal microbiota-derived tryptophan metabolites are predictive of Ah receptor activity. Gut Microbes 12, 1–24 (2020).
Xu, N. et al. Astragaloside IV reversed the autophagy and oxidative stress induced by the intestinal microbiota of AIS in mice. J. Microbiol. 56, 838–846 (2018).
Hopkins, M. J. & Macfarlane, G. T. Changes in predominant bacterial populations in human faeces with age and with Clostridium difficile infection. J. Med. Microbiol. 51, 448–454 (2002).
Claesson, M. J. et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl Acad. Sci. USA 108, 4586–4591 (2011).
Biagi, E. et al. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 5, e10667 (2010).
Wu, W. et al. Integrated 16S rRNA gene sequencing and LC-MS analysis revealed the interplay between gut microbiota and plasma metabolites in rats with ischemic stroke. J. Mol. Neurosci. 71, 2095–2106 (2021).
Spychala, M. S. et al. Age‐related changes in the gut microbiota influence systemic inflammation and stroke outcome. Ann. Neurol. 84, 23–36 (2018).
Blasco, M. P. et al. Age-dependent involvement of gut mast cells and histamine in post-stroke inflammation. J. Neuroinflammation 17, 160 (2020).
Lee, J. et al. Gut microbiota-derived short-chain fatty acids promote poststroke recovery in aged mice. Circ. Res. 127, 453–465 (2020).
Hubbard, T. D. et al. Divergent ah receptor ligand selectivity during hominin evolution. Mol. Biol. Evol. 33, 2648–2658 (2016).
Vicentini, F. A. et al. Behavioural adaptations after antibiotic treatment in male mice are reversed by activation of the aryl hydrocarbon receptor. Brain Behav. Immun. 98, 317–329 (2021).
Wei, Y.-L. et al. Fecal microbiota transplantation ameliorates experimentally induced colitis in mice by upregulating AHR. Front. Microbiol. 9, 1921 (2018).
Chen, L. et al. Dysregulation of intestinal health by environmental pollutants: involvement of the estrogen receptor and aryl hydrocarbon receptor. Environ. Sci. Technol. 52, 2323–2330 (2018).
Carter, A. M. et al. Phosphoprotein-based biomarkers as predictors for cancer therapy. Proc. Natl Acad. Sci. USA 117, 18401–18411 (2020).
Labonté, B. et al. Sex-specific transcriptional signatures in human depression. Nat. Med. 23, 1102–1111 (2017).
Patrizz, A. N. et al. Sex-specific differences in autophagic responses to experimental ischemic stroke. Cells 10, 1825 (2021).
Noh, B. et al. G-quadruplexes stabilization upregulates CCN1 and accelerates aging in cultured cerebral endothelial cells. Front. Aging 2, 797562 (2021).
Ngwa, C. et al. Age and sex differences in primary microglia culture: a comparative study. J. Neurosci. Methods 364, 109359 (2021).
Chauhan, A., Moser, H. & McCullough, L. D. Sex differences in ischaemic stroke: potential cellular mechanisms. Clin. Sci. 131, 533–552 (2017).
Iadecola, C. & Gorelick, P. B. The Janus face of cyclooxygenase-2 in ischemic stroke: shifting toward downstream targets. Stroke 36, 182–185 (2005).
Honarpisheh, P. et al. Dysregulated gut homeostasis observed prior to the accumulation of the brain amyloid-Β in TG2576 mice. Int. J. Mol. Sci. 21, 1711 (2020).
Zheng, K. et al. Single-cell RNA-seq reveals the transcriptional landscape in ischemic stroke. J. Cereb. Blood Flow. Metab. 42, 56–73 (2021).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2023).
Korf, J. M. et al. CD11bhigh B cells increase after stroke and regulate microglia. J. Immunol. 209, 288–300 (2022).
Honarpisheh, P. et al. Potential caveats of putative microglia-specific markers for assessment of age-related cerebrovascular neuroinflammation. J. Neuroinflammation 17, 366 (2020).
Brown, K., Abbott, D. W., Uwiera, R. R. E. & Inglis, G. D. Removal of the cecum affects intestinal fermentation, enteric bacterial community structure, and acute colitis in mice. Gut Microbes 9, 218–235 (2018).
Nelson, M. C., Morrison, H. G., Benjamino, J., Grim, S. L. & Graf, J. Analysis, optimization and verification of Illumina-Generated 16S rRNA gene amplicon surveys. PLoS ONE 9, e94249 (2014).
Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336 (2010).
DeSantis, T. Z. et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072 (2006).
McDonald, D. et al. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 6, 610–618 (2012).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200 (2011).
Han, X. & Liang, L. metabolomicsR: a streamlined workflow to analyze metabolomic data in R. Bioinforma. Adv. 2, vbac067 (2022).
Perkins, J. R. et al. ReadqPCR and NormqPCR: R packages for the reading, quality checking and normalisation of RT-qPCR quantification cycle (Cq) data. BMC Genomics 13, 296 (2012).
Gu, Z., Eils, R. & Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 32, 2847–2849 (2016).
Acknowledgements
The figures were created using BioRender and published under agreement numbers ZX27KC12SA, DP27GHBB1A, KQ23P0DMSF and NE23P0D37X. Metabolomics were performed at the MD Anderson Metabolomics Core Facility and was guided by Philip L Lorenzi, PhD and Lin Tan. This work was supported by the Huffington Foundation and the NIH. NINDS 1F31NS118984-01 (to P.H.), NIH/NIA 1R01AG070934-01 (to B.P.G.), NIH/AG058463 and NINDS R35 NS132265/NINDS (to L.D.M.), and NIA P30 AG066468 (Neuropathology Core, the University of Pittsburgh Alzheimer’s Disease Research Centers to J.K.K.).
Author information
Authors and Affiliations
Contributions
P.P. contributed to the study design, completion of all the experiments, data analysis and interpretation, and manuscript writing and editing. M.P.B.C., A.E., R.K., G.U.G., P.H., E.C.M., A.B., J.M.K., B.E.W., R.M.R., J.F.M.M., and M.E.M. were involved in bench experiments, sample collection, sample storage, and handling. C.T. performed immunohistochemistry on human brain slides. J.A., J.L., JunL., and R.M.B. provided input for the experimental design and manuscript editing. K.A.K., A.C., S.R., and V.R.V. performed MCAO surgeries. L.D.M. and B.P.G. were involved in the study design, data interpretation, and manuscript editing.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Qi Wang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Peesh, P., Blasco-Conesa, M.P., El Hamamy, A. et al. Benefits of equilibrium between microbiota- and host-derived ligands of the aryl hydrocarbon receptor after stroke in aged male mice. Nat Commun 16, 1767 (2025). https://doi.org/10.1038/s41467-025-57014-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-57014-2
This article is cited by
-
Molecular basis of autism spectrum disorders
Molecular Biology Reports (2025)
-
Gut microbiota deficiency reduces neutrophil activation and is protective after ischemic stroke
Journal of Neuroinflammation (2025)
