
Abstract
The eastern Tropical North Atlantic Oxygen Minimum Zone (ETNA OMZ) plays a critical role in marine ecosystems off northwestern Africa. One of the key controls of the ETNA OMZ is ventilation driven by the subsurface ocean circulation of the Atlantic subtropical gyres. However, how this shallow circulation interacts with changes in the strength of the Atlantic Meridional Overturning Circulation (AMOC) remains unclear. Here, we present a deglacial and high-resolution paleo-oxygenation record (combined bottom and pore water) from the margin of ETNA OMZ (GeoB9512-5, 793 m water depth), which registers more strongly oxygenated periods during the Last Glacial Maximum (LGM), two parts of the Heinrich Stadial 1 (HS1), and during the Younger Dryas (YD). We show that steeper meridional temperature gradients during HS1 and YD associated with AMOC slowdown intensified the subsurface subtropical cell circulation and increased the oxygen supply to the ETNA OMZ.
Similar content being viewed by others

Introduction
Oxygen Minimum Zones (OMZ) are found beneath highly productive upwelling systems1 that sustain unique marine ecosystems in the world oceans2,3,4,5. The diversity of these ecosystems depends, among other factors, on the oxygen supply, which has decreased in the last decades, partly due to global ocean warming associated with anthropogenic greenhouse gas emissions6,7. Another concern in a warming world, are projections that the large-scale deep circulation of the ocean, the Atlantic Meridional Overturning Circulation (AMOC), could possibly slowdown in the foreseeable future8,9,10,11,12. Although a future AMOC slowdown is still under debate, our understanding of its relationship with subsurface ventilation driven by shallow wind-driven subtropical cells (STCs) requires attention.
Since the available instrumental oxygen records at any ocean depth date only from the 1970s13, our knowledge must rely on marine paleo-oxygenation records. One approach comes from detailed benthic foraminifera taxonomic analyses used to infer changes in ocean oxygen concentrations14,15. Changes in this parameter actively modify benthic organisms’ physiological responses and are registered as shifts in biodiversity and distribution patterns of benthic foraminifera5,16,17,18. Benthic foraminifera are a microfossil and extant group that makes up around 50% of the eukaryotic biomass in modern oceans19 and are one of the most diverse microorganisms in the ocean20. Their distribution mostly depends on seafloor food and oxygenation21,22,23,24,25,26, and since the environmental preferences of several species are well documented, environmental models based on benthic foraminifera assemblages hold promise to investigate oceanic oxygen variability, particularly in OMZs27,28,29,30,31.
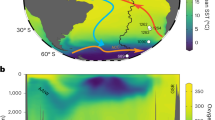
In this study, we used the Enhanced Benthic Foraminifera Oxygen Index15 (EBFOI), supplemented with additional data on benthic foraminifera oxygen preferences14,32,33,34,35 (Supplementary Data 2–4), to reconstruct the deglacial paleo-oxygenation trends of the mid-depth OMZ lower boundary (300−800 m water depth) of the eastern tropical North Atlantic (ETNA OMZ) (300−800 m water depth). This OMZ, which reaches oxygen concentrations as low as 40 µmol/kg in its core36,37,38, forms in the “shadow zone” between the north and south subtropical gyres39, where older water masses with lower renewal rates40 lead to poorly ventilated regions that make up the ETNA OMZ (Fig. 1a). The high productivity of the surface waters off the NW African coast is driven by upwelling induced by the northeast trade winds41, which forms part of a shallow wind-driven overturning circulation: the Subtropical Cell (STC). The upwelled waters are replaced in the subsurface through Ekman downwelling of surface waters in the subtropical convergence40.

a. Geographic location of gravity core GeoB9512-5 (15°20’13.20”N/17°22’1.20”W, 793 m water depth), the Atlantic subtropical gyres and the Eastern Tropical North Atlantic Oxygen Minimum Zone outline at 500 m water depth (shadowed) are located following previous studies37,40,98. Upwelling areas are indicated with the Approximate Chlorophyll a concentration (mg/m3) zones based on Aqua-Modis data (4 km resolution, in November 2019, https://oceancolor.gsfc.nasa.gov/); b. GeoB9512-5 location and modern configuration of the Eastern Tropical North Atlantic Oxygen Minimum Zone at depth (dashed outline). Plotted with Ocean Data View99 using the GLODAP v2.2022 oxygen data base43, interpolated by DIVA.
To reconstruct the combined bottom and pore water oxygen (BPWO) changes of ETNA OMZ, we used the sedimentary record of gravity core GeoB9512-5, retrieved off the coast of Senegal in northwestern Africa42 (15°20’13.20”N, 17°22’1.20”W, 793 m water depth, Fig. 1). The core is situated south of the North Atlantic subtropical gyre in the zone of seasonal (winter) upwelling. Modern dissolved oxygen concentrations at the core site are around 88 µmol/kg43 and correspond to the deep margin of the ETNA OMZ (Fig. 1b), which separates the lower oxygen conditions of the OMZ with the more oxygenated waters below and makes it a sensitive recorder of changes in this OMZ boundary.
The BPWO record presented here is based on a stratigraphic framework modeled with 16 Accelerator Mass Spectrometry radiocarbon dating of planktic foraminifera44 (Fig. 2) and covers the last 27,000 years. This time interval includes periods of AMOC decline, registered in the sedimentary 231Pa/230Th ratios records45,46,47,48 during the Heinrich Stadial 1 (HS1, ~17.6 – 14.7 ka BP) and Younger Dryas ~12.6 – 11.8 ka BP), also seen in carbon isotope records49 (δ13C). Although uncertainties exist in the 231Pa/230Th proxy as it can be overprinted by, for example, opal fluxes50, a large number of available data for the Atlantic allows drawing a reliable record of the large-scale changes seen in AMOC variability51.

Downcore median calendar ages modeled using 16 radiocarbon ages (triangles in the Core Depth axis)44 in the Bacon Package87 with the Marine20 calibration88. The timing of the Heinrich Stadials is supported by elevated Fe/Ca and Ti/Ca ratios100 representing enhanced dust deposition during continental aridity77.
Here, we present a high-resolution continuous BPWO record of the last deglaciation in the eastern tropical North Atlantic to investigate the effect of AMOC-induced circulation changes on the STC during times of documented AMOC shoaling (LGM) and slowdown (HS1 and YD) (Fig. 3).

Graphic Abstract.
Results
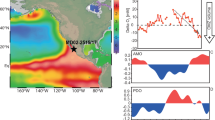
Our record shows bottom/pore water dissolved oxygen concentrations (BPWO in Fig. 4e) of between 92 and 257 µmol/kg (EBFOI between 17.9 and 98.7, Fig. 4e). BPWO reconstruction shows high oxygen conditions (dissolved oxygen concentrations >161–192 µmol/kg, 95% confidence interval-CI) at the ETNA OMZ margin from the onset of the record (~27 ka BP), through most of the last glacial until 18.9 ka BP. This period was interrupted only by one drop in BPWO centered on 25.1 ka BP. Even higher values (on average 180–224 µmol/kg, 95% CI) were recorded between 24 - 23.5 ka BP (Supplementary Data 1), and to a lesser extent during the LGM (169–193 µmol/kg, 95% CI). The deglaciation was characterized by abrupt transitions from oxygenated conditions to lower BPWO periods. After 18.9 ka BP, at the onset of the Heinrich Stadial 1 (HS1), BPWO concentrations decreased to an average 108–137 µmol/kg (95% CI) with lower values of 103–132 µmol/kg (95% CI) around 17.8 ka BP. This was followed by a rapid increase in BPWO during HS1, with two distinctive BPWO peaks (1) of 166–207 µmol/kg (95% CI) around 16.8 - 16.5 ka BP and (2) of 182–231 µmol/kg (95% CI) at 15.5 - 14.8 ka BP, which represents one of the highest BPWO values of the record. Between 16.2 and 15.9 ka BP, BPWO concentrations decreased again (average 113–152 µmol/kg, 95% CI).

a Sedimentary protactinium- thorium ratio (231Pa/230Th) from the Bermuda Rise47 (black and green curves), and deep western North Atlantic45 (red curve), error bars represent 2 s.e.m. b Atlantic Sea Surface Temperature (SST) gradient estimated by subtracting SST at site SU811866 (37.76 N, 10.18 W) from SSTs at site M35003-467 (12.09 N, 61.243 W); NW Africa Upwelling proxies. c Opal (bSiO2 %). d Diatom content during the last deglaciation from site GeoB7926-280 (20°13’N, 18°27’ W), shaded areas within the curve indicate enhanced upwelling periods. e GeoB9512-5 mean bottom and pore water oxygen (BPWO) in µmol/kg (Supplementary Data 1) calculated using the enhanced benthic foraminifera oxygen index (EBFOI, black curve), shaded area corresponds to the 95% confidence interval and gray dots represent the raw data. Key Climate events: Heinrich Stadial 2 (HS2); Last Glacial Maximum (LGM); Heinrich Stadial 1 (HS1); Bølling–Allerød (B-A); Younger Dryas (YD). Triangles in the age axis indicate radiocarbon ages.
BPWO dropped below 122–158 µmol/kg (95% CI) between 14.6 and 13.2 ka BP during the Bølling–Allerød (B-A). A transient BPWO increase (148–179 µmol/kg, 95% CI) was recorded between 13.1 ka and 12.6 ka BP during the YD, which was followed by the highest BPWO values in our record of 154–230 µmol/kg (95% CI) at 12.2 ka BP. This BPWO increase persisted at the onset of the Holocene when the final peak of 170–217 µmol/kg (95% CI) was registered between 11.3 and 10.9 ka BP. The last BPWO increase (on average 126–147 µmol/kg, 95% CI) occurred between 5 and 4.2 ka BP, and was followed by the lowest BPWO concentrations in the record of 97–101 µmol/kg (95% CI) between 1.6 and 0.2 ka BP. These values compare well with the modern measured BPWO concentration of 87.6 µmol/kg at our site43,52.
The paleo-BPWO record presented here is characterized by an alternation of relatively low BPWO intervals and BPWO peaks (Fig. 4e), as seen by the variations of the proportions of oxic, suboxic and dysoxic species used in the EBFOI calculations (Fig. 5b). The relative abundance of stress species (Fig. 5c), which are foraminifera species better adapted to environments of reduced oxygen and increased organic matter availability22, are consistent with these interpretations as they are present in average percentages over 50 % in lower BPWO intervals, and under 50 % in higher BPWO intervals.

a Two-point moving average bottom and pore water oxygen (BPWO µmol/kg, black curve) estimated from the Enhanced Benthic Foraminifera Oxygen Index (EBFOI, green dots). b Relative abundances of Oxic, Suboxic and Dysoxic species. c Relative abundances of Stress Species; and (d) infaunal benthic foraminifera. Key Climate events: Heinrich Stadial 2 (HS2); Last Glacial Maximum (LGM); Heinrich Stadial 1 (HS1); Bølling–Allerød (B-A); Younger Dryas (YD). Triangles in the age axis indicate radiocarbon ages.
In our record, a high abundance of infaunal (Fig. 5d) and stress species (Fig. 5c) indicates eutrophic and mesotrophic seafloor environments (moderate to high organic matter concentrations) throughout the whole record, from the last glacial into modern times (Supplementary Data 1). Although acids released by remineralization of organic matter under oxic conditions can result in calcite dissolution in seafloor sediments53,54, foraminifera tests appeared well preserved throughout the core.
Discussion
We observe an association between oxic intervals at our site (Fig. 4e) and AMOC perturbations (Fig. 4a), that aligns with observations that the major shifts in ocean circulation during these events altered ventilation throughout the whole ocean55. Increased BPWO is recorded during the LGM, when AMOC circulation was shallower and likely stronger than it is today56, while the highest values of the record occur in HS1 and the YD, when AMOC circulation was severely reduced. A slight increase in AMOC strength between 5.5 - 3 ka (Fig. 4e) is also associated with a brief increase in BPWO in our record, suggesting AMOC control on ETNA OMZ oxygenation also under interglacial conditions.
Modern low oxygen conditions of the ETNA OMZ are set by the poorly ventilated Eastern South Atlantic Central Water and the southward flow of better ventilated North Atlantic Central Water37,57 (NACW). On the other hand, cold periods in the North Atlantic are associated with southward shifts of hydrographic and atmospheric frontal systems58,59,60. Southward movement of the front between northern and southern sourced central waters allowed oxygen-rich NACW to penetrate further south along the NW African margin61. At the same time, displacement of the major wind systems and the ITCZ62 lead to the North Atlantic subtropical gyre being positioned several degrees south of its current location during the LGM, HS1 and YD63. This would have exposed the core site to the younger and better ventilated waters of the North Atlantic Subtropical Gyre and is also supported by ΔCO3-2 records of the west Atlantic that show low calcite saturation related to increasing better ventilated northern sourced waters during the HS1 and YD64. During the B-A and Holocene times, as AMOC deepened and strengthened (Fig. 4a), the subtropical gyre moved north, contributing to reduced BPWO conditions registered by benthic foraminifera in our site (Fig. 4e).
The North Atlantic sea level pressure gradient and hence the anticyclonic wind circulation driving the STG is also tightly related to the distribution of sea surface temperature65. To illustrate the relation between North Atlantic Sea Surface Temperature (SST) and OMZ ventilation, we estimated the deglacial SST gradient across the Atlantic by calculating the difference in SST between site SU81-1866 in the northeastern STG and site M35003-467 in the tropical west Atlantic using interpolated datasets (Fig. 4b). Modest surface cooling in the tropical west Atlantic, and greater cooling in the northeast Atlantic during the LGM, increased the temperature gradient compared to the Holocene. Increases in the meridional temperature gradient and wind stress were even more intense during HS1 and YD (Fig. 4b) when reduction in the upper branch of AMOC warmed the sea surface and upper tropical west Atlantic67,68,69,70, while input of glacial meltwater reduced SST in the northeast of the gyre66. Stronger winds would have increased subduction of surface waters into the subtropical cell, hence ventilating the upper ocean.
Such a higher SST gradient coincides with increased BPWO in the GeoB9512-5 record during the last glacial (Fig. 4e). This is related to increased glacial wind stress curl that deepened the Subtropical Gyre circulation, ventilating the subsurface and intermediate Atlantic71. In addition to ventilation by stronger winds, lower nutrient input, due to glacial stratification of the Southern Ocean72, may have reduced oxygen consumption in the upper ocean as a whole. This is supported by increasing BPWO conditions in the LGM compared to the Holocene, recorded in sites of the East Pacific and tropical Atlantic above 1000 m73 (Supplementary Fig. S3.1b in Supplementary Information 3) and the mid-depth/upper Atlantic74,75 (Supplementary Fig. S3.1a in Supplementary Information 3), while the deep ocean was low in dissolved oxygen and high in carbon73,76.
The high temperature gradients across the North Atlantic sea surface registered in times of AMOC slowdown during HS1 and the YD (Fig. 4b) coincide with the highest BPWO values of the GeoB9512-5 record (Fig. 4e). Strong northeasterly winds over NW Africa77 enhanced the upwelling off the African coast which, combined with increased nutrients in the upper Atlantic78, strengthened productivity as shown in diatom and opal flux records (Fig. 4c, d) from the West African coast41,79,80. In our record, BPWO is high in two distinct oxygen peaks during HS1 and during the entire YD. Although high productivity may have led to more oxygen consumption in the water column81, increased ventilation from stronger winds in the formation area of North Atlantic Central Waters was evidently the dominant factor controlling the BWPO at our site.
Increased BPWO during HS1 has also been recorded in the upper West Atlantic32, which in our deglacial BPWO record is observed as a pronounced double peak, a pattern that is absent in the deep 231Pa/230Th record from the Bermuda Rise45,47 (Fig. 4a). A recent compilation of 231Pa/230Th records from the tropical and North Atlantic48 indicates two phases of AMOC slowdown, an early slowdown due to meltwater input and iceberg calving from the Eurasian Ice Sheets from about 19 to 16.5 ka BP, and a later slowdown associated with Laurentide iceberg calving from about 16.5 to 15 ka BP. Two phases of ice rafting and meltwater input are also recorded in Atlantic sediments82. Whether the transition between the two phases of meltwater input is associated with a continuous AMOC decline seems currently unclear. Deglacial radiocarbon records from the Greenland Sea indeed show a brief phase of stronger vertical convection from about 15.9 to 15.2 ka83. We thus suggest that the two phases of AMOC slowdown expressed in ETNA OMZ ventilation as a double peak in BPWO at GeoB9512-5 (Fig. 4e), were likely interrupted by a brief AMOC intensification, which might not be resolved in deglacial core sections, or is below the detection threshold of 231Pa/230Th.
Our data shows that a decline in AMOC strength increased ventilation of the Eastern Tropical Atlantic Oxygen Minimum Zone. This is despite AMOC perturbations being associated with an increase in temperature and salinity of the upper ( < 1500 m) tropical Atlantic63,69,70,84, which would act to reduce oxygen solubility - and, during HS1 and the YD, an increase in productivity off West Africa85 - which would increase oxygen consumption. Our findings imply that the future of the ETNA OMZ is critically dependent on the state of the AMOC, and that any future decline might counteract the current de-oxygenation trend related to global warming.
Methods
Chronology and Age model
The downcore ages from site GeoB9512-5 (Fig. 2) were modeled using 16 Accelerator Mass Spectrometry (AMS) radiocarbon ages from planktic foraminifera (Trilobatus sacculifer and Globigerina bulloides) samples44. The samples were measured at the MICADAS laboratory at the Alfred Wegener Institute (AWI)-Bremerhaven86. A continuous age model was calculated with the R script BACON87 version 2.5.5 and the Marine20 calibration curve88 to obtain a median calendar age and uncertainty for each sampled depth. The resulting age model shows a deglacial sequence deposited in the last 27,000 years with no age reversals and includes the key climatic periods Heinrich Stadial 2 (HS2), Last Glacial Maximum (LGM), Heinrich Stadial 1 (HS1), Bølling–Allerød warming (B-A), Younger Dryas (YD), and the Holocene Climatic Optimum (HCO), that divides the early Holocene from the Late Holocene (Fig. 2).
Oceanographic setting
Water masses in our site were identified using a T-S diagram showing GeoB9512-5 situation (Fig. 6a), along with the existing available references. The studied gravity core GeoB9512-542 (15°20’13.20’N, 17°22’1.20’W, 793 m water depth, Fig. 1), is situated in the “shadow zone” between the north and south Atlantic subtropical gyres (Fig. 1a). At our site, modern low oxygen values of 88 µmol/kg characterize the ETNA OMZ lower margin (Fig. 1b), and bottom water temperatures and salinities are approximately 6.9 °C, and ~34.8 (Fig. 6a) respectively43. Such conditions position GeoB9512-5 in the mixing area of the North Atlantic Central Water (NACW) and Eastern South Atlantic Central Water40 (ESACW, Fig. 6).

a Temperature-Salinity diagram showing the water masses sources in the eastern North Atlantic, adapted from previous studies40,57, dashed lines represent the transition of North and South Atlantic Central Waters (NACW and SACW, respectively) in the eastern North Atlantic85. b Salinity showing the eastern North Atlantic central water masses distribution in the studied area40. c Dissolved oxygen concentration (umol/kg)43 and modern extension at depth of the East North Atlantic Oxygen Minimum Zone (ETNA). Oceanographic sections (a and b) were plotted with Ocean Data View99 using the GLODAP v2.2022 oxygen data base43, interpolated by DIVA.
The South Atlantic Central Water flows northward from the South Atlantic subtropical gyre57, where the coincidence of wind-driven downwelling and seasonal buoyancy drives waters into the thermocline89. In the Cape Verde area, this water mass is present as Western and Eastern South Atlantic Central Waters (Fig. 6b, WSACW and ESACW respectively). The WSACW is differentiated by its formation area in the western South Atlantic, and even though it is mainly concentrated in the western Atlantic it reaches the eastern North Atlantic to some extent57. The ESACW is a nutrient-rich water mass formed at the mixing area of the Agulhas Current and the South Atlantic Current south of Africa40,57. Above the SACW masses, the North Atlantic Central Water (NACW) flows southward and is represented in our area by the Eastern North Atlantic Central Water (ENACW, Fig. 6b, c), bringing –low nutrient, better ventilated waters from its formation area, the inter-gyre region (between 39-48°N)57.
The interaction of these water masses plays an important role in the Eastern Tropical North Atlantic Oxygen Minimum Zone (ETNA OMZ), as the northward flow of poorly ventilated and lower oxygenated waters of the ESACW are counteracted by the southward flow of the oxygenated ENACW37,57. The site studied here (GeoB9512-5) is located in the lower ETNA OMZ in a mixture of central waters, where more than 50% of the lower OMZ is NACW which has retroflected in the gyre, at around 14°N90.
Benthic foraminifera taxonomy and quantitative analyses
Sediment samples were (1) washed through a 63 µm sieve with deionized water; (2) dried in an oven at ~45 °C for no more than 24 hours; and (3) dry sieved through 63, 125, 150, and 250 µm, and stored and labeled in glass vials. The > 150 µm fraction was analyzed to extract benthic foraminifera (for taxonomic and quantitative analyses) and planktic foraminifera (for radiocarbon dating). Paleoenvironmental and paleo-oxygenation results and interpretation are based on the taxonomical identification and the quantitative analyses of at least 200–250 benthic foraminifera from 100 samples from core GeoB9512-591.
Benthic foraminifera were morphologically separated, and the taxonomical identification was made to species level in most cases (Supplementary Data 2). Genera were determined following Loeblich and Tappan (1987)92, and species were identified based on multiple refs. 93,94,95. The updated taxonomy was finally revised using the online database WoRMS96 (Supplementary Data 2). Digital images of the benthic foraminifera extracted during this study were taken at the Microscopy Laboratory at MARUM, using a Keyence VHX 6000 digital microscope with a motorized stage (Supplementary Information 1).
Bottom/Pore Water Oxygenation reconstruction and uncertainties
The BPWO record presented here was based on the Enhanced Benthic Foraminifera Oxygen Index15 (EBFOI). The terms high oxic (>3 ml/l (>131 µmol/kg)), low oxic (1.5 – 3 ml/l (65 – 131 µmol/kg), suboxic (0.3 – 1.5 ml/l (13. – 65 µmol/kg)), dysoxic (0.1 – 0.3 ml/l (4 – 13 µmol/kg)) and anoxic (<0.1 ml/l (<4 µmol/kg)) are used here as previously defined15. EBFOI relies on multiple species, and of the 151 taxonomic units identified at our site, 143 (including all abundant species) have information about their oxygen preference (Supplementary Data 2 and 4). Although the EBFOI is one of the best tools to quantify the dissolved oxygen concentration changes from the whole livable habitat15 (bottom and pore water oxygen concentration), it can be limited by the availability of information about the oxygen preferences of benthic foraminifera species and, has been shown to underestimate oxygen concentrations32. For this study, we used the existing databases of benthic foraminifera oxygen preferences14,15,32 and supplemented this with published information on living assemblages from seafloor-surface samples from the eastern tropical Atlantic33,34,35 (Supplementary Fig. S2.1 in Supplementary Information 2). The methodology is described in detail in Supplementary Information 2 and resulted in an updated compilation (“(1) Total Compilation” in Supplementary Data 3), that was used in the transfer function of Kranner et al.15 to calculate dissolved oxygen concentrations in ml/l.
To facilitate the interpretation of our record in contrast to already published paleo-oxygenation data, and as recent literature includes oxygen descriptions in μmol/kg, we converted our ml/l BPWO record to μmol/kg. Dissolved oxygen concentrations in ml/l for each sample from site GeoB9512-5 (Supplementary Data 1) were converted to μmol/kg by multiplying the calculated oxygen (ml/l) by the molar volume of oxygen (44.66 μmol/ml) and then dividing by the bottom water density of site GeoB9512-5 (1.027 kg/l). The density was computed using the specific bottom water salinity and temperature at our site, estimated from GLODAP version 2.202243 in the seawater state equation97. The universal NOAA/WOD18 notes conversion (1 ml/l of oxygen is equivalent to 43.570 μmol/kg) was also reported for reference.
We also assessed the accuracy of our reconstruction by comparing our paleo-oxygenation record with downcore relative abundances of stress species (Fig. 5c), that are better adapted to oxygen-depleted environments in organic matter rich-environments (Bulimina, Bolivina, Cassidulina, Melonis, Fissurina, Globobobulimna)26.
To estimate the uncertainty associated with our data, we assumed that oceanographic conditions, including AMOC strength, were relatively stable in samples younger than 2.6 ka BP (n = 6), allowing us to estimate reproducibility. Dissolved oxygen calculated from EBFOI for four samples (between 2.5 and 17.5 cm) in gravity core GeoB9512-5 and two samples (2 cm, 8 cm sediment depth) from nearby multicore GeoB9512-442 (Supplementary Data 2) gave a mean value of 96 μmol/kg, close to the modern 88 μmol/kg43, and standard deviation of 7 μmol/kg. Since the distribution of our data is normal (p-value = 0.114 using the Shapiro test), the 95% confidence interval (CI) was estimated considering the age uncertainty and the standard deviation reported here (Fig. 4e1), combined with 10,000 downcore proxy series produced with the Bacon script for R87. The mean oxygen values are detailed in Supplementary Data 1.
Data availability
The benthic foraminifera counts - taxonomical information and Accelerator Mass Spectrometry Carbon 14 data used in this study are available in the PANGAEA® - Data Publisher for Earth & Environmental Science, with the identifiers [https://doi.org/10.1594/PANGAEA.962951] and [https://doi.org/10.1594/PANGAEA.962899]. The data generated in this study have been deposited in the Zenodo repository with the identifier [https://doi.org/10.5281/zenodo.10724771]. The paleoenvironmental information per sample, benthic foraminifera oxygen preference data, and the benthic foraminifera images generated in this study are also provided in Supplementary Data 1, 2 and Supplementary Information 1, respectively.
Code availability
The code used to process and visualize the benthic foraminifera data can be found at Barragán-Montilla (2024) via the identifier https://doi.org/10.5281/zenodo.10724771. The script for the age model is based on Blaauw and Christen (2011) and processing and visualization of the data is available in the same repository.
References
Wyrtki, K. The oxygen minima in relation to ocean circulation. Deep Sea Res. Oceanogr. Abstr. 9, 11–23 (1962).
Bruno, J. F. et al. Climate change threatens the world’s marine protected areas. Nat. Clim. Change 8, 499–503 (2018).
Stramma, L., Johnson, G. C., Sprintall, J. & Mohrholz, V. Expanding oxygen-minimum zones in the tropical Oceans. Science 320, 655–658 (2008).
Stramma, L. et al. Expansion of oxygen minimum zones may reduce available habitat for tropical pelagic fishes. Nat. Clim. Change 2, 33–37 (2012).
Gilly, W. F., Beman, J. M., Litvin, S. Y. & Robison, B. H. Oceanographic and biological effects of shoaling of the oxygen minimum zone. Annu. Rev. Mar. Sci. 5, 393–420 (2013).
Schmidtko, S., Stramma, L. & Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nature 542, 335–339 (2017).
Gilbert, D. Oceans lose oxygen. Nature 542, 303–304 (2017).
Calvin, K. et al.IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (Eds.)]. IPCC, Geneva, Switzerland. (2023).
Liu, W., Fedorov, A. V., Xie, S.-P. & Hu, S. Climate impacts of a weakened Atlantic Meridional Overturning Circulation in a warming climate. Sci. Adv. 6, eaaz4876 (2020).
Zhang, R. et al. A review of the role of the Atlantic meridional overturning circulation in Atlantic multidecadal variability and associated climate impacts. Rev. Geophys. 57, 316–375 (2019).
Caesar, L., Rahmstorf, S., Robinson, A., Feulner, G. & Saba, V. Observed fingerprint of a weakening Atlantic Ocean overturning circulation. Nature 556, 191–196 (2018).
Weijer, W., Cheng, W., Garuba, O. A., Hu, A. & Nadiga, B. T. CMIP6 models predict significant 21st century decline of the Atlantic meridional overturning circulation. Geophys. Res. Lett. 47, e2019GL086075 (2020).
Moore, W. S. Review of the geosecs project. Nucl. Instrum. Methods Phys. Res. 223, 459–465 (1984).
Kaiho, K. Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology 22, 719 (1994).
Kranner, M., Harzhauser, M., Beer, C., Auer, G. & Piller, W. E. Calculating dissolved marine oxygen values based on an enhanced Benthic Foraminifera Oxygen Index. Sci. Rep. 12, 1376 (2022).
Zhang, J. et al. Natural and human-induced hypoxia and consequences for coastal areas: synthesis and future development. Biogeosciences 7, 1443–1467 (2010).
Pörtner, H.-O., Bock, C. & Mark, F. C. Oxygen- and capacity-limited thermal tolerance: bridging ecology and physiology. J. Exp. Biol. 220, 2685–2696 (2017).
Hanz, U. et al. Environmental factors influencing benthic communities in the oxygen minimum zones on the Angolan and Namibian margins. Biogeosciences 16, 4337–4356 (2019).
Gooday, A. J., Levin, L. A., Linke, P. & Heeger, T. inDeep-Sea Food Chains and the Global Carbon Cycle (eds. Rowe, G. T. & Pariente, V.) 63–91 (Springer Netherlands, Dordrecht, 1992).
Sen Gupta, B. K. in Modern Foraminifera 3–6 (Springer, Dordrecht, 1999).
Altenbach, A., Bernhard, J. & Seckbach, J. Anoxia: Evidence for Eukaryote Survival and Paleontological Strategies. (Springer Netherlands, Dordrecht, 2012).
Bernhard, J. M. & Sen Gupta, B. K. in Modern Foraminifera 201–216 (Springer Netherlands, Dordrecht, 1999).
Jorissen, F. J., De Stigter, H. C. & Widmark, J. G. V. A conceptual model explaining benthic foraminiferal microhabitats. Mar. Micropaleontol. 26, 3–15 (1995).
Murray, J. W. Ecology and Applications of Benthic Foraminifera. (Cambridge University Press, 2006).
Murray, J. W. Ecology and Palaeoecology of Benthic Foraminifera. (Longman scientific and technical copublished in the United States with John Wiley and sons, Essex New York, 1991).
Southward, A. J. Advances in Marine Biology. (Academic, San Diego, Calif. London, 2003).
Hoogakker, B. A. A. et al. Reviews and syntheses: Review of proxies for low-oxygen paleoceanographic reconstructions. Biogeosciences 22, 863–957 (2025).
Erdem, Z. et al. Bottom-water deoxygenation at the Peruvian margin during the last deglaciation recorded by benthic foraminifera. Biogeosciences 17, 3165–3182 (2020).
Moffitt, S. E., Hill, T. M., Roopnarine, P. D. & Kennett, J. P. Response of seafloor ecosystems to abrupt global climate change. Proc. Natl. Acad. Sci. USA 112, 4684–4689 (2015).
Tetard, M., Licari, L. & Beaufort, L. Oxygen history off Baja California over the last 80 kyr: A new foraminiferal-based record: Oxygen History in the California Margin. Paleoceanography 32, 246–264 (2017).
Sharon Belanger, C., Du, J. & Mix, A. Reconstructing Paleo-oxygenation for the Last 54,000 Years in the Gulf of Alaska Using Cross-validated Benthic Foraminiferal and Geochemical Records. Paleoceanogr. Paleoclimatol. 36, e2020PA003986 (2021).
Schmiedl, G., Milker, Y. & Mackensen, A. Climate forcing of regional deep-sea biodiversity documented by benthic foraminifera. Earth Sci. Rev. 244, 104540 (2023).
Haake, F. W. Benthische Foraminiferen in Oberflächen-Sedimenten und Kernen des Ostatlantiks vor Senegal/Gambia (Westafrika). (1980).
Licari, L. & Mackensen, A. Assemblage of living benthic foraminifera in sediment core GeoB3715-1. 1304 data points PANGAEA https://doi.org/10.1594/PANGAEA.511360 (2006).
Lutze, G. F. Distribution of benthic foraminifera in surface sediments on the continental margin off North-West Africa. 6 datasets https://doi.org/10.1594/PANGAEA.548469 (1980).
Brandt, P. et al. Changes in the ventilation of the oxygen minimum zone of the tropical North Atlantic. J. Phys. Oceanogr. 40, 1784–1801 (2010).
Brandt, P. et al. On the role of circulation and mixing in the ventilation of oxygen minimum zones with a focus on the eastern tropical North Atlantic. Biogeosciences 12, 489–512 (2015).
Karstensen, J., Stramma, L. & Visbeck, M. Oxygen minimum zones in the eastern tropical Atlantic and Pacific oceans. Prog. Oceanogr. 77, 331–350 (2008).
Sarmiento, J. L. A tritium box model of the North Atlantic thermocline. J. Phys. Oceanogr. 13, 1269–1274 (1983).
Poole, R. & Tomczak, M. Optimum multiparameter analysis of the water mass structure in the Atlantic Ocean thermocline. Deep Sea Res. Part Oceanogr. Res. Pap. 46, 1895–1921 (1999).
Bradtmiller, L. I. et al. Changes in biological productivity along the northwest African margin over the past 20,000 years: African Margin Paleoproductivity. Paleoceanography 31, 185–202 (2016).
Mulitza, S. et al. Report and preliminary results of METEOR Cruise M65/1, Dakar - Dakar, 11.06. - 1.07.2005. (2005).
Lauvset, S. K. et al. GLODAPv2.2022: the latest version of the global interior ocean biogeochemical data product. Earth Syst. Sci. Data 14, 5543–5572 (2022).
Barragán-Montilla, S. & Mulitza, S. Radiocarbon ages of sediment core GeoB9512-1. PANGAEA (2025).
Böhm, E. et al. Strong and deep Atlantic meridional overturning circulation during the last glacial cycle. Nature 517, 73–76 (2015).
Gherardi, J. et al. Evidence from the Northeastern Atlantic basin for variability in the rate of the meridional overturning circulation through the last deglaciation. Earth Planet. Sci. Lett. 240, 710–723 (2005).
McManus, J. F., Francois, R., Gherardi, J.-M., Keigwin, L. D. & Brown-Leger, S. Collapse and rapid resumption of Atlantic meridional circulation linked to deglacial climate changes. Nature 428, 834–837 (2004).
Ng, H. C. et al. Coherent deglacial changes in western Atlantic Ocean circulation. Nat. Commun. 9, 2947 (2018).
Boyle, E. A. & Keigwin, L. North Atlantic thermohaline circulation during the past 20,000 years linked to high-latitude surface temperature. Nature 330, 35–40 (1987).
Lippold, J. et al. Does sedimentary 231 Pa/ 230 Th from the Bermuda Rise monitor past Atlantic Meridional Overturning Circulation? Geophys. Res. Lett. 36, 2009GL038068 (2009)).
Bradtmiller, L. I., McManus, J. F. & Robinson, L. F. 231Pa/230Th evidence for a weakened but persistent Atlantic meridional overturning circulation during Heinrich Stadial 1. Nat. Commun. 5, 5817 (2014).
Key, R. M. et al. Global Ocean Data Analysis Project, Version 2 (GLODAPv2). https://doi.org/10.3334/CDIAC/OTG (2015).
Emerson, S. R. & Bender, M. Carbon fluxes at the sediment-water interface of the deep-sea: Calcium carbonate preservation. 39, J. Mar. Res. https://elischolar.library.yale.edu/journal_of_marine_research/1537 (1981).
Emerson, S. R. & Archer, D. Calcium carbonate preservation in the ocean. Philos. Trans. R. Soc. Lond. Ser. Math. Phys. Sci. 331, 29–40 (1990).
Schmittner, A. et al. Calibration of the carbon isotope composition (δ 13 C) of benthic foraminifera. Paleoceanography 32, 512–530 (2017).
Kurahashi-Nakamura, T., Paul, A. & Losch, M. Dynamical reconstruction of the global ocean state during the Last Glacial Maximum. Paleoceanography 32, 326–350 (2017).
Liu, M. & Tanhua, T. Water masses in the Atlantic Ocean: characteristics and distributions. Ocean Sci. 17, 463–486 (2021).
McGee, D. et al. Hemispherically asymmetric trade wind changes as signatures of past ITCZ shifts. Quat. Sci. Rev. 180, 214–228 (2018).
Shakun, J. D. & Carlson, A. E. A global perspective on Last Glacial Maximum to Holocene climate change. Quat. Sci. Rev. 29, 1801–1816 (2010).
Repschläger, J. et al. Response of the subtropical North Atlantic surface hydrography on deglacial and Holocene AMOC changes. Paleoceanography 30, 456–476 (2015).
Huang, E. et al. Response of eastern tropical Atlantic central waters to Atlantic meridional overturning circulation changes during the Last Glacial Maximum and Heinrich Stadial 1. Paleoceanogr. Paleoclimatol. 27, 2012PA002294 (2012).
Portilho-Ramos, R. C. et al. Coupling of equatorial Atlantic surface stratification to glacial shifts in the tropical rainbelt. Sci. Rep. 7, 1561 (2017).
Reißig, S., Nürnberg, D., Bahr, A., Poggemann, D.-W. & Hoffmann, J. Southward displacement of the North Atlantic subtropical gyre circulation system during North Atlantic cold spells. Paleoceanogr. Paleoclimatol. 34, 866–885 (2019).
Oppo, D. W. et al. Deglacial temperature and carbonate saturation state variability in the tropical Atlantic at Antarctic intermediate water depths. Paleoceanogr. Paleoclimatol. 38, e2023PA004674 (2023).
deMenocal, P. B. & Rind, D. Sensitivity of Asian and African climate to variations in seasonal insolation, glacial ice cover, sea surface temperature, and Asian orography. J. Geophys. Res. Atmos. 98, 7265–7287 (1993).
Bard, E., Rostek, F., Turon, J.-L. & Gendreau, S. Hydrological impact of Heinrich events in the subtropical Northeast Atlantic. Science 289, 1321–1324 (2000).
Rühlemann, C., Mulitza, S., Müller, P. J., Wefer, G. & Zahn, R. Warming of the tropical Atlantic Ocean and slowdown of thermohaline circulation during the last deglaciation. Nature 402, 511–514 (1999).
Came, R. E. et al. in Geophysical Monograph Series (eds. Schmittner, A., Chiang, J. C. H. & Hemming, S. R.) vol. 173 247–263 (American Geophysical Union, Washington, D. C., 2007).
Poggemann, D. et al. Deglacial heat uptake by the Southern Ocean and rapid Northward redistribution via Antarctic intermediate water. Paleoceanogr. Paleoclimatol. 33, 1292–1305 (2018).
Rühlemann, C. et al. Intermediate depth warming in the tropical Atlantic related to weakened thermohaline circulation: Combining paleoclimate data and modeling results for the last deglaciation. Paleoceanography 19, 2003PA000948 (2004).
Wharton, J. H. et al. Deeper and stronger North Atlantic Gyre during the Last Glacial Maximum. Nature 632, 95–100 (2024).
Burke, A. & Robinson, L. F. The Southern Ocean’s role in carbon exchange during the last deglaciation. Science 335, 557–561 (2012).
Wang, Y., Costa, K. M., Lu, W., Hines, S. K. V. & Nielsen, S. G. Global oceanic oxygenation controlled by the Southern Ocean through the last deglaciation. Sci. Adv. 10, eadk2506 (2024).
Skinner, L. C. et al. Atlantic Ocean ventilation changes across the last deglaciation and their carbon cycle implications. Paleoceanogr. Paleoclimatol. 36, e2020PA004074 (2021).
Lynch-Stieglitz, J. et al. A diminished North Atlantic nutrient stream during Younger Dryas climate reversal. Science 384, 693–696 (2024).
Zhou, Y. & McManus, J. F. Authigenic uranium deposition in the glacial North Atlantic: Implications for changes in oxygenation, carbon storage, and deep water-mass geometry. Quat. Sci. Rev. 300, 107914 (2023).
Mulitza, S. et al. Sahel megadroughts triggered by glacial slowdowns of Atlantic meridional overturning. Paleoceanogr. Paleoclimatol. 23, https://doi.org/10.1029/2008PA001637 (2008).
Poggemann, D.-W. et al. Rapid deglacial injection of nutrients into the tropical Atlantic via Antarctic Intermediate Water. Earth Planet. Sci. Lett. 463, 118–126 (2017).
Bouimetarhan, I., Groeneveld, J., Dupont, L. & Zonneveld, K. Low- to high-productivity pattern within Heinrich Stadial 1: Inferences from dinoflagellate cyst records off Senegal. Glob. Planet. Change 106, 64–76 (2013).
Romero, O. E., Kim, J. & Donner, B. Submillennial-to-millennial variability of diatom production off Mauritania, NW Africa, during the last glacial cycle. Paleoceanogr. Paleoclimatol. 23, 2008PA001601 (2008).
Lu, W. et al. I/Ca in epifaunal benthic foraminifera: A semi-quantitative proxy for bottom water oxygen in a multi-proxy compilation for glacial ocean deoxygenation. Earth Planet. Sci. Lett. 533, 116055 (2020).
Hodell, D. A. et al. Anatomy of Heinrich Layer 1 and its role in the last deglaciation: HEINRICH EVENT 1. Paleoceanogr. Paleoclimatol. 32, 284–303 (2017).
Thornalley, D. J. R., Elderfield, H. & McCave, I. N. Reconstructing North Atlantic deglacial surface hydrography and its link to the Atlantic overturning circulation. Glob. Planet. Change 79, 163–175 (2011).
Weldeab, S., Friedrich, T., Timmermann, A. & Schneider, R. R. Strong middepth warming and weak radiocarbon imprints in the equatorial Atlantic during Heinrich 1 and Younger Dryas. Paleoceanogr. Paleoclimatol. 31, 1070–1082 (2016).
Tomczak, M. & Godfrey, J. S. Regional Oceanography: An Introduction. (Pergamon, Oxford, England; New York, 1994).
Mollenhauer, G., Grotheer, H., Gentz, T., Bonk, E. & Hefter, J. Standard operation procedures and performance of the MICADAS radiocarbon laboratory at Alfred Wegener Institute (AWI), Germany. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. 496, 45–51 (2021).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Heaton, T. J. et al. Marine20—The Marine Radiocarbon Age Calibration Curve (0–55,000 cal BP). Radiocarbon 62, 779–820 (2020).
Slowey, N. C. & Curry, W. B. Enhanced ventilation of the North Atlantic subtropical gyre thermocline during the last glaciation. Nature 358, 665–668 (1992).
Peña-Izquierdo, J. et al. Water mass pathways to the N orth A tlantic oxygen minimum zone. J. Geophys. Res. Oceans 120, 3350–3372 (2015).
Barragán-Montilla, S. Benthic Foraminifera counts off NW Africa during the last deglaciation. 45030 data points PANGAEA https://doi.org/10.1594/PANGAEA.962951 (2024).
Loeblich Jr, A. R. & Tappan, H. Foraminiferal Genera and Their Classification. (Springer, New York, NY, 1987).
Jones, R. W. & Brady, H. B. The Challenger Foraminifera. (Oxford University Press, Oxford; New York, 1994).
Holbourn, A. E. L. & Henderson, A. S. Atlas of Benthic Foraminifera. (Natural History Museum, Chichester, West Sussex; Hoboken, NJ, 2013).
Morkhoven, F. P. C. M. van, Berggren, W. A. & Edwards, A. S. Cenozoic Cosmopolitan Deep-Water Benthic Foraminifera. (Elf Aquitaine, Pau, 1986).
WoRMS Editorial Board. World Register of Marine Species. Available from https://www.marinespecies.org at VLIZ. Accessed yyyy-mm-dd. [object Object] https://doi.org/10.14284/170 (2024).
Gill, A. E. International Geophysics Series. Volume 30, Atmosphere-Ocean Dynamics. (Academic Press, New York, 1982).
Pelegrí, J. L. & Peña-Izquierdo, J. Eastern boundary currents off North-West Africa. in Oceanographic and biological features in the Canary Current Large Marine Ecosystem 81–92 (2015).
Schlitzer, R. Ocean Data View, odv.awi.de. https://odv.awi.de/ (2023).
Völpel, R., Mulitza, S., Paul, A., Lynch-Stieglitz, J. & Schulz, M. Water Mass Versus Sea Level Effects on Benthic Foraminiferal Oxygen Isotope Ratios in the Atlantic Ocean During the LGM. Paleoceanogr. Paleoclimatology 34, 98–121 (2019).
Acknowledgements
This research was funded through the Cluster of Excellence “The Ocean Floor – Earth’s Uncharted Interface” (EXC-2077, Project 390741603). The GeoB Core Repository at MARUM provided the sample material used here. The authors thank Master students Jakob Quabeck, who produced some of the microfossil images and Ishani Rathnayake, who helped with sample preparation. This research is also supported by GLOMAR – Bremen International Graduate School for Marine Sciences, University of Bremen.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
S.B.M., H.J.H.J., S.M., D.A.R.M., B.H.A.A. and H.P. contributed to the preparation of the final manuscript. S.B.M contributed with the conceptualization, core sampling, sample preparation, benthic foraminifera extraction, taxonomy and quantitative analyses, data visualization, interpretation and discussion, and wrote the manuscript; H.J.H.J. contributed with the conceptualization, data interpretation and discussion, and manuscript editing; S.M. contributed with the core sampling, conceptualization, age model, discussion, and manuscript editing; D.A.R.M. and B.H.A.A. contributed with the quantitative analyses, oceanographic data revision, and manuscript editing; H.P. contributed with the conceptualization, data discussion and manuscript editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Barragán-Montilla, S., Johnstone, H.J.H., Mulitza, S. et al. Enhanced ventilation of Eastern North Atlantic Oxygen Minimum Zone with deglacial slowdown of Meridional Overturning. Nat Commun 16, 6418 (2025). https://doi.org/10.1038/s41467-025-61177-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-61177-3
