
Abstract
A major obstacle in knocking down oncogenes for tumor therapy is the efficient delivery of siRNA into the cytosolic spaces of cancer cells. Here, we genetically bioengineer biomimetic nanovesicles with tumor-recognition and enzyme-controlled membrane fusion functions for efficiently delivering small interfering RNA into cancer cells towards gene silencing tumor therapy. The siRNA@eS-BNVs are formulated by encapsulating siRNA inside the core and coating with genetically engineered HEK293TACE2- cell membranes encoded with functional S protein, which can recognize cancer cells and initiate membrane fusion when triggered by the enzyme. The siRNA@eS-BNVs demonstrate better efficacy for cytosolic siRNA delivery and RNA interference than conventional formulations. By intravenous injection, siRNA@eS-BNVs are highly accumulated in tumors and potently inhibited tumor and lung metastasis by simultaneously silencing the epidermal growth factor receptor gene in vivo. The cancer cell-targeting and enzyme-activatable nanovesicles provide a valuable strategy for effective and precise drug delivery.
Similar content being viewed by others

Introduction
Knocking down disease-related genes with synthetic small interfering RNA (siRNA) holds great potential for dealing with a myriad of diseases, such as cancer1,2,3,4. However, naked siRNAs are unable to achieve in vivo RNA interference (RNAi) effects due to the difficulties in reaching diseased tissues and cells, and intracellular trafficking into the cytosol to bind the RNA-induced silencing complex (RISC) for silencing the disease-related genes5,6,7, e.g., oncogenes. Creating delivery systems is the key to protecting siRNA from RNase degradation and systemic clearance, promoting the accumulation in neoplastic tissues and cytosolic siRNA delivery via cellular internalization, and enabling endosome escape for accomplishing effective RNAi and cancer therapy1,4,8,9,10,11. Despite progress in developing siRNA delivery platforms, including cationic polymeric nanoparticles12,13, lipid nanoparticles (LNPs)9, cell membrane-coated nanoparticles (CNPs)14,15,16, and inorganic nanoparticles17, etc., the efficacy for intracellular siRNA delivery is still very low and unsatisfactory8, i.e., only 1-2% of siRNA escaped from endosomes into the cytosol, and most of siRNA was degraded in lysosomes18,19, which impaired the biological outcomes of RNAi. Therefore, developing siRNA delivery vehicles based on well-defined structures and programmable cellular internalization demonstrates high promise for intracellular drug delivery. One recent approach is the rational design and development of nanocarriers with membrane fusion functions to bypass the endosome-lysosome pathway and directly transport drugs (e.g., siRNA) into the cytoplasm of cancer cells for precise drug delivery20,21,22,23. Several fusogen-anchored nanocarriers have been engineered for spontaneous membrane fusion with target cells, demonstrating improved intracellular drug delivery efficacy20,21,22,23. Current membrane fusion approaches rely on the processes that occur naturally between the nanocarriers and target cells. Therefore, we hypothesized that engineering nanocarriers with cancer cell-targeting and controlled membrane fusion functions would further enable precise siRNA delivery24,25.
Here, we genetically bioengineered angiotensin-converting enzyme 2 (ACE2) receptor knock out and mutant spike (SSM, abbreviated as S in this study) protein knock in human embryonic kidney 293 T (eS-HEK293TACE2-) cell membranes, and engineered biomimetic nanovesicles (eS-BNVs) with it for efficient cytosolic siRNA delivery through ligand-receptor-mediated and enzyme-controlled membrane fusion, to inhibit tumor and metastasis. The siRNA@eS-BNVs were formulated by encapsulating siRNA inside the core in the form of siRNA/polyethylenimine (PEI) complex and wrapped with genetically engineered HEK293TACE2- cell membranes encoded with mutated S proteins (Fig. 1a), which could recognize lung cancer cells and specifically initiate membrane fusion when triggered by the thrombin (Thr) enzyme for intracellular siRNA delivery (Fig. 1b). The Thr enzyme-triggered membrane fusion between eS-BNVs and cancer cells has been studied with S, wild-type spike (SWT), and double mutant spike (SDM) protein-anchored nanovesicles. The siRNA@eS-BNVs for cytosolic siRNA delivery through Thr enzyme-triggered membrane fusion, and in vitro and in vivo RNAi were investigated. By intravenous (i.v.) injection, siRNA@eS-BNVs were highly accumulated in tumors, potently inhibited tumor growth and lung metastasis by simultaneously silencing the epidermal growth factor receptor (EGFR) gene in vivo. The cell-targeting and enzyme-controlled membrane fusion biomimetic nanovesicles provide a valuable platform and approach for precise intracellular drug delivery.

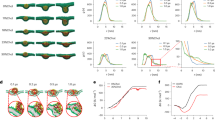
a Scheme shows engineering siRNA@eS-BNVs by genetically programming HEK293T cells via ACE2 receptor knock out and S protein knock in plasmids for obtaining eS-HEK293TACE2- cell membranes to encapsulate siRNA/PEI complexes. b Scheme shows siRNA@eS-BNVs for intracellular siRNA delivery by recognizing ACE2 receptors on cancer cells and enzyme-controlled membrane fusion via Thr-triggered S protein conformation change. c Structures and sequences of S and SWT proteins. Left: simulated structural alignments of S (pink) and SWT (blue, PDB: 6VXX), and S2’ (dark blue) in the S protein. Right: S protein and its cleavage site. SWT is a transmembrane (TM) protein with S1 and S2 domains. S1 contains the native RBD domain for recognition. S2 contains S2’ linker, FP, HR1 and HR2 domains. FP domain is exposed by digestion of S2’ site, then HR1 and HR2 form six helical bundles, which are the main structures for membrane fusion. d Western blot analysis of ACE2 and S expressed on HEK293T and eS-HEK293TACE2- cells. β-actin was an internal reference. e Representative flow cytometry results show higher expression of S protein on eS-HEK293TACE2- cells (blue) than HEK293TACE2- cells (gray). f Diameter of eS-BNVs. g Representative TEM image of eS-BNVs. Scale bar,100 nm. h SDS-PAGE and western blot analysis of S protein in eS-BNVs and No-BNVs nanovesicles. PEI, polyethylenimine; FP, fusion peptide; HR1, heptad repeat 1; HR2, heptad repeat 2; RBD, receptor binding domain; S1, S1 Subunit; S2, S1 Subunit; S1/S2, S1/S2 cleavage site; S2’, S2’ cleavage site; SWT, wild-type spike protein; S, mutant spike protein; Thr, Thrombin enzyme; EGFR, epidermal growth factor receptor; RISC, RNA-induced silencing complex. M, marker. Experiments in (d, g and h) were repeated three times independently with similar results. Source data are provided in a Source Data file.
Results
Genetic engineering S protein overexpressing biomimetic nanovesicles
We genetically programmed ACE2 receptor knock-out and S protein knock in HEK293T cells. Firstly, we genetically knocked out the ACE2 receptor in the HEK293T cells by transfection of short hairpin RNA (shRNA) to obtain HEK293TACE2- cells. Next, to express spike proteins on the membrane of HEK293TACE2- cells, we engineered the S protein plasmid by replacing the native S2’ cleavage site (KPSKR) with Thr (LVPRGS) cleavage site (Fig. 1c). The structure of the S protein, as predicted by SWISS-MODEL, remained unchanged compared to the SWT protein, suggesting that the mutation would not impact its function (Fig. 1c). Subsequently, we encoded the S plasmids for overexpression of S protein on the membranes of HEK293TACE2- cells to obtain eS-HEK293TACE2- cells. The western blot assay confirmed the successful knock out of the ACE2 receptor and resounding knock in of exogenous S protein on the surface of eS-HEK293TACE2- cells (Fig. 1d). The flow cytometry further confirmed significantly high expression of S protein on eS-HEK293TACE2- cells when applied HEK293TACE2- cells as controls (Fig. 1e and Supplementary Fig. 2a, b). These results demonstrated the successful construction of genetically engineered eS-HEK293TACE2- cells.
Subsequently, the eS-HEK293TACE2- and HEK293TACE2- cell membranes were extracted and purified for formulating eS-BNVs and No-BNVs biomimetic nanovesicles with the cell membranes as shells through sonication and mechanical extrusion. The average hydrodynamic diameter of eS-BNVs was 116 nm as measured by dynamic light scatterings (DLS) (Fig. 1f). Transmission electron microscope (TEM) imaging revealed the monodispersity and vesicle morphology of eS-BNVs (Fig. 1g). The SDS-PAGE and western blot assays demonstrated the presence of S protein on the eS-BNVs, appearing as a distinct band with a molecular weight of approximately 180 kDa, consistent with the previous report26, while no S protein was investigated on the No-BNVs (Fig. 1h). Therefore, the S protein overexpressing nanovesicles (i.e., eS-BNVs) were successfully obtained by genetic bioengineering HEK293T cells and formulation.
In vitro interaction between cancer cells and eS-BNVs
To investigate whether the mutation of the S protein affected its binding affinity to the ACE2 receptor, the eSWT-HEK293TACE2- cell membranes were also extracted to engineer the cell membrane-formulated biomimetic nanovesicles, named eSWT-BNVs. The western blot experiment showed a clean band at 180 kDa in eSWT-BNVs (Supplementary Fig. 3a), indicating the successful expression of the SWT protein in the eSWT-HEK293T ACE2- cells. Meanwhile, the ACE2 receptor expressed on the surface of non-small cell lung (i.e., A549ACE2+) cancer cells was observed by confocal laser scanning microscope (CLSM) after labeling with anti-ACE2 antibody (Fig. 2a and Supplementary Fig. 4). To further verify the interaction between S protein and ACE2 receptor, we prepared ACE2 receptor knockout A549 (i.e., A549ACE2-) cancer cells, which have no ACE2 receptor as confirmed by CLSM (Supplementary Fig. 5a). As shown in Fig. 2a, The ACE2 receptor expressed on the membrane of the A549ACE2+ cancer cells is the primary receptor for specifically binding with the spike protein (i.e., S protein)27, which plays an important role in the recognition of eS-BNVs with the A549ACE2+ cancer cells.

a Representative CLSM image shows the ACE2 receptor expressed on A549ACE2+ cells. Scale bar, 10 μm. b eS-BNVsDiI bind to A549ACE2+ cell membrane visualized by CLSM. Red: CM; green: ACE2 receptor; blue: nucleus. CM, cell membrane. Scale bar, 10 μm. c, d Flow cytometry results show higher internalization between A549ACE2+ cells and eS-BNVsDiI or eSWT-BNVsDiI than No-BNVsDiI (n = 3 independent samples). e Western blot analysis of S protein after incubation No-BNVs or eS-BNVs with A549ACE2+ cells, respectively. f, g SRB fluorescence intensity for monitoring Thr-induced membrane fusion between SRB@eSDM-BNVs and NVsACE2+ (f) but failed between SRB@eSWT-BNVs and NVsACE2+ (g) (n = 3 independent samples). Plotted curves with self-quenching SRB fluorescence represent membrane fusion. h Western blot shows cleavage of SDM or SWT after adding Thr to NVsACE2+ and eSDM-BNVs or eSWT-BNVs. i Representative TEM images show Thr enzyme-controlled and S protein-mediated membrane fusion between eS-BNVs and gold nanoparticle-labeled A549ACE2+-BNVs, including recognition of A549ACE2+-BNVs with S protein bind to the ACE2 receptor, fusion pore expanding and full fusion. Red arrows indicate gold nanoparticle markers. Scale bars, 50 nm. j, k Thr-induced strong ensemble content-mixing (membrane fusion) between SRB@eS-BNVs and A549ACE2+ cells with increased SRB fluorescence intensity (j), but Thr did not induce membrane fusion between SRB@eSWT-BNVs and A549ACE2+ cells (k) (n = 3 independent samples). l Western blot shows the cleavage of S or SWT protein after adding Thr to A549ACE2+ cells and eS-BNVs or eSWT-BNVs. The cleavage state of SDM, S and SWT proteins was confirmed by detecting the S1 fragment with anti-RBD antibody. CM, cell membrane; Max, maximum. M, marker. Experiments in (a, b, e, h, i and l) were repeated three times independently with similar results. Data in (d, f, g, j and k) are means ± s.d. p-values were determined by one-way ANOVA test in (d) or two-tailed Student’s t test in (f, g, j, k). ****p < 0.0001, ***p < 0.001, ns, not significant. Source data are provided in a Source Data file.
Subsequently, we evaluated the binding efficiency between the eS-BNVs and A549ACE2+ cancer cells. To visualize the eS-BNVs, No-BNVs and eSWT-BNVs, we labeled the nanovesicles with DiI dye to obtain No-BNVsDiI, eS-BNVsDiI and eSWT-BNVsDiI. As imaged by the CLSM, eS-BNVsDiI were efficiently bound to the membranes of A549ACE2+ cancer cells (Fig. 2b), which would be due to the internalization with the ACE2 receptor. In contrast, eS-BNVsDiI were not detected on the surface of A549ACE2- cancer cells (Supplementary Fig. 5b), which would be due to the disappearance of the internalization between the S protein and ACE2 receptor. Compared to No-BNVsDiI, the eS-BNVsDiI exhibited greater binding efficiency to A549ACE2+ cancer cells as analyzed by flow cytometry, demonstrating the targeting function of eS-BNVs to the A549ACE2+ cancer cells (Fig. 2c, d). However, both No-BNVsDiI and eS-BNVsDiI showed weak binding efficacy to A549ACE2- cancer cells (Supplementary Fig. 5c, d). Of note, the binding affinity of eS-BNVsDiI was similar to that of eSWT-BNVsDiI, demonstrating the mutation of spike protein (i.e., change in the S2’ cleavage site) in eS-BNVs would not affect its binding affinity to the ACE2 receptor. Furthermore, the presence of eS-BNVs on the surface of A549ACE2+ cancer cells was confirmed by western blot analysis of the A549ACE2+ cancer cells and A549ACE2- cancer cells after exposure to the No-BNVs or eS-BNVs (Fig. 2e and Supplementary Fig. 5e). In addition, Calu3 cancer cells are often employed to study the functions of spike protein in membrane fusion between cells28. Therefore, we investigated the binding efficiency between eS-BNVs and Calu3 cancer cells as a control experiment (Supplementary Fig. 7a–d). The binding efficacy of the eS-BNVs to Calu3 cancer cells was considerably lower than that to the A549ACE2+ cancer cells. Thus, the genetically programmed eS-BNVs exhibited stronger binding efficacy to A549ACE2+ cancer cells than to Calu3 cancer cells. Therefore, the eS-BNVs could effectively target the A549ACE2+ cancer cells.
In vitro enzyme-controlled membrane fusion between eS-BNVs and cancer cells
Effective cleavage of the mutant spike protein (i.e., S protein) is an important function and prerequisite for enzyme-controlled membrane fusion29. Therefore, the cleavage of the mutant S protein was evaluated by checking the molecular weight of the protein bands in the western blot after being cleaved via the Thr enzyme. As shown in Fig. 2l, the clean band of the mutant S protein at 180 kDa disappeared upon the treatment by the Thr enzyme (Fig. 2l). However, the Thr enzyme did not cause any changes to the SWT protein (i.e., wild-type spike protein) (Fig. 2l). Therefore, the mutant S protein could be efficiently and specifically cleaved by the Thr enzyme.
To study the Thr enzyme-controlled processes of membrane fusion, we generated another double mutant spike protein (SDM, in which the native S1/S2 cleavage site (PRRAR) and S2’ cleavage site were changed to PreScission (3 C, LEVLFQGP) and Thr enzyme cleavage sites, respectively (Supplementary Fig. 8a, b). In addition, we encoded plasmids to overexpress SDM in HEK293TACE2- cells to obtain eSDM-HEK293TACE2- cells. The membrane of eSDM-HEK293TACE2- cells was extracted to engineer biomimetic nanovesicles, named eSDM-BNVs (Supplementary Fig. 3a). Moreover, we also prepared ACE2 receptor-anchored nanovesicles (NVsACE2+) to mimic the ACE2 receptor-expressing cancer cells. self-quenched sulphorhodamine B (SRB) fluorescent dye-labeled eSDM-BNVs (SRB@eSDM-BNVs). TEM images showed that both NVsACE2+ and SRB@eSDM-BNVs were spheric and monodisperse vesicles (Supplementary Fig. 9a, b). In addition, the successful construction of the ACE2 receptor-anchored NVsACE2+ was verified by SDS-PAGE assay (Supplementary Fig. 3b).
Next, the Thr enzyme-controlled membrane fusion between SRB@eSDM-BNVs or SRB@eSWT-BNVs and NVsACE2+ was assessed through the content-mixing assays. As shown in Fig. 2f, the fluorescence intensity of the mixed SRB@eSDM-BNVs and NVsACE2+ was dramatically increased upon the addition of the Thr enzyme to the mixture system. However, no significant increase in fluorescence intensity was investigated in the SRB@eSDM-BNVs and NVsACE2+ system without adding Thr enzyme, and the SRB@eSWT-BNVs to NVsACE2+ system with or without Thr enzyme (Fig. 2f, g). In addition, in the content mixing test between eSWT-BNVs and NVsACE2+, the intensities were quite low in both presence and absence of Thr enzyme (Fig. 2g). It demonstrated that the cleavage of mutant spike protein (i.e., S protein) is essential for enzyme-controlled membrane fusion, which is consistent with previous reports29,30. Therefore, the cleavage of proteins by the Thr enzyme in the reaction systems between NVsACE2+ and eSDM-BNVs or eSWT-BNVs was verified by western blot. As shown in Fig. 2h, the protein band of the mutant SDM protein at 180 kDa disappeared, and smaller protein bands between 100–150 kDa appeared when treated with Thr enzyme (Fig. 2h, left). However, the Thr enzyme did not cause any changes to the SWT protein (i.e., WT spike protein) (Fig. 2h, right). The results showed that the SDM protein can be specifically cleaved by the Thr enzyme. Those observations suggested that the Thr enzyme could trigger the membrane fusion of nanovesicles, due to the cleavage of the S2’ site in the mutant S protein by Thr. The cleavage of the S2’ site is a critical step in initiating membrane fusion, consistent with previous findings31,32. Moreover, the 3 C enzyme was further added to investigate the membrane fusion between the SRB@eSDM-BNVs and NVsACE2+, as the 3 C enzyme could trigger cleavage of the S1/S2 site in the SDM protein. As shown in Supplementary Fig. 10a, there was no notable difference in the membrane fusion of SRB@eSDM-BNVs and NVsACE2+ when triggered by Thr enzyme alone (i.e., + Thr) or by both Thr and 3 C enzymes (i.e., + Thr and 3 C). Therefore, combining 3 C enzymes did not accelerate the membrane fusion processes of SRB@eSDM-BNVs and NVsACE2+ (Supplementary Fig. 10a). Then, we verified whether Thr and 3 C enzymes can cleave the SDM protein by detecting the protein band changes by western blot. As shown in Supplementary Fig. 10b, a band at the S1 subunit with a molecular weight smaller than the S1’ subunit (cleaved by Thr enzyme) appeared when cleaved by 3 C enzyme at the S1/S2 cleavage site, indicating that both Thr and 3 C enzymes can cleave the SDM protein. In the control experiments, the 3 C enzyme alone did not trigger membrane fusion between SRB@eSDM-BNVs and NVsACE2+, and it also did not trigger membrane fusion of SRB@eSWT-BNVs and NVsACE2+ (Supplementary Fig. 10c). This is apparently due to that Thr and 3 C enzymes were unable to cleave the SWT protein (Supplementary Fig. 10d). Therefore, Thr enzyme specifically induced the cleavage of the SDM protein to initiate the membrane fusion between the eSDM-BNVs and NVsACE2+ rather than the 3 C enzyme.
Furthermore, the efficacy of membrane fusion through spike protein in the presence or absence of ACE2 receptor was verified by content mixing between SRB@eS-BNVs and NVsACE2-. By the content mixing experiments in the presence of Thr enzyme, no membrane fusion phenomenon was investigated between SRB@eS-BNVs and NVsACE2- (Supplementary Fig. 11a). Then, we characterized spike protein by western blot after the experiment, and found that S protein can still be digested by Thr enzyme in the absence of ACE2 receptor (Supplementary Fig. 11b). However, the remaining band of intact spike protein at 180 kDa indicated that the cleavage efficiency of S protein was reduced in the absence of ACE2 receptor, which may be influenced by the interaction between spike protein and ACE2 receptor33. The above experiments indicated that the presence of the ACE2 receptor was critical for the membrane fusion of eS-BNVs.
We further investigated the membrane fusion processes between eS-BNVs and A549ACE2+ cancer cells membrane-made nanovesicles (i.e., A549ACE2+-BNVs) by TEM (Fig. 2i). The A549ACE2+-BNVs were engineered to mimic cancer cells by taking A549ACE2+ cancer cells membrane and extruding it through polycarbonate filters (100 nm pore size), obtaining an average diameter of 127 nm as measured by DLS (Supplementary Fig. 12a, b) and uniform vesicle morphology as characterized by TEM (Supplementary Fig. 12c). Next, we used gold nanoparticles to label the A549ACE2+-BNVs to distinguish from eS-BNVs. As shown in Fig. 2i, the A549ACE2+-BNVs and eS-BNVs were separated initially. Subsequently, upon the recognition of the target A549ACE2+-BNVs by eS-BNVs via the S protein that was cleaved by Thr enzyme, the eS-BNVs and A549ACE2+-BNVs were tightly combined, the membrane fusion ensued, associating with the disappearance of the membrane boundary between the two vesicles and the formation of a fusion pore. Eventually, eS-BNVs and A549ACE2+-BNVs merged into a single vesicle. However, no membrane fusion phenomenon was investigated between the eS-BNVs and A549ACE2+-BNVs without the addition of Thr enzyme (Supplementary Fig. 12d). TEM images provided morphological evidence and illustrated various intermediate states of membrane fusion between A549ACE2+-BNVs and eS-BNVs, including the free state of eS-BNVs, targeted recognition, opening of fusion pores, and completed fusion states, all occurring in the presence of the Thr enzyme. Thus, it provided solid evidence for the Thr enzyme-controlled membrane fusion processes between the eS-BNVs and A549ACE2+-BNVs.
Next, the Thr enzyme-controlled membrane fusion between eS-BNVs or eSWT-BNVs and A549ACE2+ cancer cells was assessed through the content mixing assays (Fig. 2j–l). When the Thr enzyme was introduced to the SRB@eS-BNVs and A549ACE2+ cancer cells, the fluorescence intensity was increased approximately 2.87-fold higher than that without adding Thr enzyme (i.e., -Thr) (Fig. 2j), but failed to induce membrane fusion between SRB@eSWT-BNVs and A549ACE2+ cancer cells (Fig. 2k). The fluorescence intensity in SRB@eS-BNVs and A549ACE2+ cancer cells system was rapidly reached peak within 30 min. The status of S protein and SWT protein after Thr enzyme cleavage was further checked by western blot. As shown in Fig. 2l, the S protein was cleaved by Thr enzyme (Fig. 2l, left), while the SWT protein did not change (Fig. 2l, right). Thus, Thr enzyme can trigger membrane fusion between SRB@eS-BNVs and A549ACE2+ cancer cells, but not between SRB@eSWT-BNVs and A549ACE2+ cancer cells, indicating Thr enzyme is essential for initiating membrane fusion between eS-BNVs and A549ACE2+ cancer cells. Moreover, we verified the necessity of the ACE2 receptor for membrane fusion. By the content mixing assay of SRB@eS-BNVs and A549ACE2- cancer cells, we found that Thr enzyme-induced membrane fusion disappeared (Supplementary Fig. 13a). The western blot further indicated that the cleavage efficacy of S protein by Thr enzyme was lower in the absence of ACE2 receptor (Supplementary Fig. 13b). Therefore, the S protein binding with ACE2 receptor was essential for Thr-triggered membrane fusion. Consistent with the binding capacity, the membrane fusion efficiency of SRB@eS-BNVs and Calu3 cancer cells was lower than that of SRB@eS-BNVs and A549ACE2+ cancer cells (Fig. 2j and Supplementary Fig. 14a), even in the presence Thr enzyme (Supplementary Fig. 14b). Taken together, the Thr enzyme played a critical role in triggering specific and efficient membrane fusion of the eS-BNVs and A549ACE2+ cancer cells.
We further examined the membrane fusion between eS-BNVs and normal cells, including human umbilical vein endothelial cells (HUVEC) and HEK293T cells, by content mixing assay. As shown in Supplementary Fig. 15a, b, there was almost no membrane fusion between SRB@eS-BNVs and HUVEC cells or HEK293T cells, as max intensity was similar to control (Supplementary Fig. 15c). Then, we detected the cellular uptake of SRB@eS-BNVs by HUVEC and HEK293T cells with flow cytometry and CLSM. As shown in Supplementary Fig. 15d, e, there was very lower cellular uptake of SRB@eS-BNVs by HUVEC and HEK293T cells when compared with SRB@eS-BNVs and A549ACE2+ cancer cells. Therefore, eS-BNVs demonstrated high efficacy in membrane fusion and cellular uptake with A549ACE2+ cancer cells rather than normal cells (e.g., HUVEC).
eS-BNVs for intracellular siRNA delivery via enzyme-controlled membrane fusion
We then encapsulated siRNA into the eS-BNVs to obtain siRNA@eS-BNVs for studying intracellular siRNA delivery and RNAi via enzyme-controlled membrane fusion. Considering direct packaging the negatively charged siRNA into eS-BNVs is challenging, we first mixed it with the positively charged PEI to form siRNA/PEI complex and then shelled it with eS-BNVs. To obtain the best siRNA delivery efficiency, we screened the N/P ratios of PEI and siRNA for engineering siRNA@eS-BNVs. As investigated by CLSM, the best PEI and siRNA N/P ratio was 5, which was chosen to formulate siRNA@eS-BNVs in the following experiments, as it has the highest cellular uptake efficacy (Supplementary Fig. 16a, b). As characterized by DLS and TEM, the siRNA@eS-BNVs demonstrated an average hydrodynamic diameter of 143 nm and the zeta potential is about − 11 mV (Fig. 3a and Supplementary Fig. 17), and an obvious spheric core-shell structure (Fig. 3b). To verify the stable loading of siRNA by eS-BNVs, we performed a SYBR Gold assay, which showed that siRNA could be stably encapsulated in eS-BNVs, and the siRNA/PEI complex dissociated in the presence of heparin (Supplementary Fig. 18). The encapsulation efficiency of siRNA by eS-BNVs was 82.44% by gel retardation assay (Supplementary Fig. 18b). In addition, we analyzed that the siRNA was stably loaded inside the eS-BNVs by gel retardation assay (Supplementary Fig. 18c).

a Diameter of siRNA@eS-BNVs. b Representative TEM image of siRNA@eS-BNVs. Scale bar,100 nm. c Single-molecular technique for analyzing the number of encapsulated Cy5-siRNA inside an eS-BNVs. d The frequency distribution of the number of siRNA encapsulated in a single nanovesicle (n = 4 independent samples). e, f CLSM images (e) and statistical analysis (f) show siRNA-Cy5@eS-BNVs for efficient siRNA delivery into A549ACE2+ cancer cells via Thr enzyme-triggered membrane fusion (n = 6 independent samples). Scale bars, 20 μm. g, h CLSM images (g) and statistical analysis (h) show the time-dependent intracellular siRNA delivery by siRNA-Cy5@eS-BNVs to A549ACE2+ cancer cells (n = 5 independent samples). Red: siRNA-Cy5; blue: nucleus. Scale bars, 20 µm. i, j Representative flow cytometry results (i) and statistical analysis (j) show cellular uptake of siRNA-Cy5@eS-BNVs via Thr enzyme-triggered membrane fusion (n = 6 independent samples). k Intracellular tracing of siRNA-Cy5@eS-BNVs and siRNA-Cy5@eS-BNVs+Thr in A549ACE2+ cancer cells. Red: siRNA-Cy5; green: lysosomes; blue: nucleus (n = 6 independent samples). Scale bars, 10 μm. l CLSM images of A549ACE2+-EGFP cancer cells after treating with ICG-labeled eS-BNVs (i.e., ICG@eS-BNVs). Red: ICG; green: A549ACE2+-EGFP cancer cells. Scale bars, 5 μm. Avg, average. Experiment in (b) was repeated three times, in (e, k and l) were repeated six times, and in (g) was repeated five times independently with similar results. Data in (d, f, h, j and k) are the means ± s.d. p-values were determined by one-way ANOVA test in (f, h, j) or two-tailed Student’s t test in (k). ****p < 0.0001, **p < 0.01, *p < 0.05, ns, not significant. Source data are provided in a Source Data file.
We further measured the number of encapsulated siRNA within a single siRNA-Cy5@eS-BNVs by using the single-molecule techniques, which are applied in biophysics to detect single molecules34. The experimental processes are illustrated in Fig. 3c. A single-molecule fluorescence quenching technique was employed to determine the number of siRNA molecules encapsulated in a single biomimetic vesicle. Firstly, the nanovesicles were labeled with biotinylated phosphatidylethanolamine (Biotin-PE) to obtain Biotin-PE-anchored siRNA-Cy5@eS-BNVs (i.e., siRNA-Cy5@eS-BNVsBiotin-PE) and then immobilized on the biotin PEG-coated slide surface by neutravidin. By quenching the fluorescence of siRNA-Cy5 upon strong fluorescence irradiation, the number of fluorescence quenching events that occurred represented the number of encapsulated siRNA within one eS-BNVs (Supplementary Fig. 19). Single-molecule experiments were obtained by a TIR fluorescence microscope (Ti2-E, Nikon). The data were recorded by NIS-elements AR and processed bysmCamera2. According to the statistics of the fluorescence quenching events occurring in a single nanovesicle, a single siRNA@eS-BNVs had an average of 2–5 siRNA molecules and a maximum of 9 siRNA molecules (Fig. 3d). Of note, this is a practical study to measure how many siRNA molecules can be packed into a single biomimetic cell membrane nanovesicles. The results effectively showcased the encapsulation efficiency of siRNA.
Next, the cellular uptake of siRNA@eS-BNVs by A549ACE2+ cancer cells was investigated by CLSM, while Cy5-labeled siRNA was also applied to trace siRNA location. As shown in Fig. 3e, f and Supplementary Fig. 20, siRNA-Cy5@eS-BNVs plus Thr enzyme (i.e., siRNA-Cy5@eS-BNVs + Thr) demonstrated the highest cellular uptake compared to the other groups, such as siRNA-Cy5/PEI, siRNA-Cy5@eS-BNVs and siRNA-Cy5@No-BNVs, etc. siRNA-Cy5@eS-BNVs+Thr taken up by cancer cells was 2.87-fold and 1.95-fold higher than that of siRNA-Cy5@No-BNVs and siRNA-Cy5@eS-BNVs, respectively (Fig. 3e, f), which promoted intracellular siRNA delivery than conventional nanocarriers such as siRNA-Cy5/PEI and siRNA-Cy5@No-BNVs, as well as nanocarriers with cancer cell-targeting ability but without membrane fusion function, such as siRNA-Cy5@eS-BNVs. The cellular uptake efficiency of siRNA-Cy5@eS-BNVs + Thr was 1.84-fold higher than that of siRNA-Cy5@eSWT-BNVs + Thr (Fig. 3f), indicating SWT protein did not induce membrane fusion for intracellular drug delivery. Moreover, there was no significant change in cellular uptake among the siRNA-Cy5@eS-BNVs, siRNA-Cy5@eSWT-BNVs and siRNA-Cy5@eSWT-BNVs + Thr groups. This suggested that both SWT and S proteins can target A549ACE2+ cancer cells, but eSWT-BNVs did not trigger membrane fusion for promoting intracellular gene delivery. We observed the cellular uptake of siRNA-Cy5@eS-BNVs + Thr by A549ACE2+ cancer cells at different times, showing a rapid cellular uptake occurred within 30 min (Fig. 3g, h and Supplementary Fig. 21). In addition, the cellular uptake of siRNA and nanocarriers was further evaluated by flow cytometry. As shown in Fig. 3i, j, it demonstrated that Thr-induced siRNA-Cy5@eS-BNVs vectors were able to provide the highest cellular uptake of siRNA-Cy5 than the other nanocarriers, e.g., siRNA-Cy5@eS-BNVs or siRNA-Cy5@eSWT-BNVs (Fig. 3i, j). There was a significant difference in the uptake of siRNA-Cy5 by A549ACE2+ cancer cells between the siRNA-Cy5@eS-BNVs + Thr and siRNA-Cy5@eSWT-BNVs + Thr groups (Fig. 3i, j), indicating eSWT-BNVs did not undergo membrane fusion to increase intracellular siRNA delivery. These results demonstrated that the eS-BNVs could efficiently deliver siRNA into cancer cells through the cancer cell-targeting and Thr enzyme-triggered membrane fusion functions.
Subsequently, we observed the pathway and state of siRNA uptake by A549ACE2+ cancer cells via membrane fusion using CLSM by referring to a recent study35. First, we prepared siRNA-Cy5@eS-BNVs with Cy5-labeled siRNA (i.e., siRNA-Cy5). The siRNA-Cy5@eS-BNVs and siRNA-Cy5@eS-BNVs+Thr were incubated with A549ACE2+ cancer cells and investigated by CLSM. As shown in Fig. 3k, the colocalization between lysosome and siRNA-Cy5@eS-BNVs was 57.55%. However, the colocalization between lysosome and siRNA-Cy5@eS-BNVs + Thr was remarkably reduced to 21.39% due to the membrane fusion between nanovesicles and cancer cells initiated by adding the Thr enzyme. Therefore, the majority of siRNA was delivered into cancer cells by eS-BNVs through the membrane fusion pathway. Next, we engineered the fluorescent dye indocyanine green (ICG)-labeled eS-BNVs (ICG@eS-BNVs) to track the intracellular trafficking via membrane fusion by CLSM. In the case of nanovesicles with Thr enzyme (ICG@eS-BNVs + Thr), the ICG was dispersed in the cytosolic regions of cancer cells, including the nucleus (Fig. 3l), indicating efficient intracellular drug delivery via Thr-triggered membrane fusion. However, the group without Thr enzyme was different, and almost no ICG dispersed in the nucleus of cancer cells. These results suggest that the eS-BNVs mainly deliver cargos (i.e., siRNA) into cancer cells through the Thr enzyme-triggered membrane fusion pathway.
In vitro and in vivo gene knockdown in cancer cells by siRNA@eS-BNVs
To explore the RNAi effects, the siLuc was incorporated in eS-BNVs to obtain siLuc@eS-BNVs, and the efficiency in silencing luciferase (Luc) protein and messenger RNA (mRNA) in A549ACE2+-Luc cancer cells was investigated. The A549ACE2+-Luc cancer cells were exposed to siLuc@eS-BNVs + Thr, siLuc@eS-BNVs, siLuc@eSWT-BNVs, siCtrl@eS-BNVs + Thr, siLuc@No-BNVs and siLuc/PEI, free siLuc and PBS, and the bioluminescence of A549ACE2+-Luc cancer cells was monitored by the in vivo imaging system (IVIS) spectrum. As shown in Fig. 4a, b, the siLuc@eS-BNVs plus Thr enzyme (siLuc@eS-BNVs + Thr) demonstrated the best gene silencing effects in the mammalian cells, demonstrating the lowest bioluminescence intensity of Luc in A549ACE2+-Luc cancer cells compared to the other samples, e.g., siLuc@No-BNVs, siLuc@eS-BNVs, and siLuc@eSWT-BNVs, etc. Subsequently, we examined the Luc mRNA levels in the A549ACE2+-Luc cancer cells by reverse transcription quantitative PCR (RT-qPCR). As shown in Fig. 4c, siLuc@eS-BNVs + Thr have yielded the lowest relative Luc mRNA level (21.67%) compared to the PBS group. The siLuc@eS-BNVs + Thr group exhibited higher gene silencing efficiency compared to other groups, such as siLuc@No-BNVs (65.90%), siLuc@eS-BNVs (45.43%), etc. This provided convincing evidence that the expression of Luc protein was strongly reduced by the delivery of siLuc to A549ACE2+-Luc cancer cells. Moreover, the gene silence effect of the siLuc@eS-BNVs + Thr group was 1.44-fold greater than that treated by the siLuc@eS-BNVs group and 2.83-fold greater than that treated by the siLuc@No-BNVs group via a multimode microplate reader (Fig. 4d). Of note, the gene silencing efficacy of siLuc@eS-BNVs + Thr was much better than the positive control, siLuc/PEI (Fig. 4a–d). Here, PEI is a cationic polymer and usually applied as a positive control for gene delivery nanocarriers36, as PEI has good endosomal escape effects due to the proton sponge effect. Although PEI has good gene transfection effects, its severe toxicity, such as causing the death of mice, hemolysis and high cytotoxicity as illustrated in this study (Supplementary Fig. 23) and many others, has impaired its direct applications in gene delivery. Thus, modifying PEI with biocompatible biomaterials, such as encapsulating PEI inside eS-BNVs, is a general approach for reliving its toxicity for gene delivery. Together, these findings suggested that recombinant Thr-activated eS-BNVs nanodevices demonstrated excellent RNAi activity in A549ACE2+ cancer cells.

a, b In vitro Luc silencing efficiency in A549ACE2+-Luc cancer cells treated by siLuc@eS-BNVs+Thr and control groups were monitored by IVIS spectrum (a) and quantification (b) of the bioluminescence intensity (n = 6 independent samples). c RT-qPCR analysis of Luc mRNA in A549ACE2+-Luc cancer cells treated by siRNA@eS-BNVs+Thr (n = 6 independent samples). d Relative bioluminescence of A549ACE2+-Luc cancer cells treated by siLuc@eS-BNVs+Thr (n = 6 independent samples). e, f Images by IVIS spectrum (e) and bioluminescence intensity (f) show in vivo Luc knockdown in A549ACE2+-Luc tumors in animal models at 0, 24, 36 and 48 h after i.v. injection of siLuc@eS-BNVs + Thr (n = 4 independent samples). gLuc mRNA levels expressed in A549ACE2+-Luc tumors examined by RT-qPCR at 48 h after treating with siLuc@eS-BNVs + Thr and control groups (n = 4 independent samples). Avg, average. Data in (b, c, d, f and g) are the mean ± s.d. p-values were determined by one-way ANOVA test in (b, c, d, g) or two-way ANOVA test in (f). ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05, ns, not significant. Source data are provided in a Source Data file.
Next, the in vivo RNAi effects were investigated by i.v. injection of PBS, free siLuc, siLuc/PEI, siCtrl@eS-BNVs + Thr, siLuc@No-BNVs, siLuc@eSWT-BNVs, siLuc@eS-BNVs and siRNA@eS-BNVs+Thr into A549ACE2+-Luc tumor-bearing BALB/c nude mice and measuring the bioluminescence of Luc protein in tumors with IVIS spectrum (Fig. 4e, f). Of note, the Thr enzyme was i.v. injected to initiate membrane fusion (1.5 U per mouse) at 12 h after i.v. administration of siLuc@eS-BNVs. As shown in Fig. 4e, f, the siLuc@eS-BNVs+Thr treated A549ACE2+-Luc tumor-bearing mice showed the lowest bioluminescence than the other groups, such as PBS, free siLuc and siCtrl@eS-BNVs, etc. Although there were no significant differences when comparing siLuc@eS-BNVs + Thr with siLuc@eS-BNVs and siLuc@eSWT-BNVs, probably due to the unavoidable variations in the luminescence intensity in vivo that make it difficult to see the difference, the average luminescence intensities were different. For instance, the Luc expression was the lowest in A549ACE2+-Luc tumors treated with siLuc@eS-BNVs + Thr, decreasing to 52.06%, 32.68% and 27.30% at 24, 36 and 48 h compared to siLuc@eS-BNVs, indicating its efficient and stable RNAi effects in vivo. Although siLuc@eSWT-BNVs and siLuc@eS-BNVs can slightly downregulate Luc expression due to the endocytosis of partial nanovesicles, the Thr enzyme-triggered membrane fusion of siLuc@eS-BNVs (i.e., siLuc@eS-BNVs+Thr) achieved much better silencing effects of Luc and its mRNA (Fig. 4e–g). Of note, the Luc silencing effects of the positive control siLuc/PEI in vivo were weak, similar to other controls. In addition, the A549ACE2+-Luc tumors were extracted from mice, and the mRNA of the Luc gene in the tumors was quantified by RT-qPCR. As shown in Fig. 4g, siLuc@eS-BNVs+Thr effectively decreased the Luc mRNA levels in A549ACE2+-Luc tumors, in which the mRNA level was the lowest compared to other groups, demonstrating its high RNAi efficacy in vivo. In summary, these results demonstrated that Thr-mediated membrane fusion of eS-BNVs can achieve robust gene silencing outcomes both in vitro and in vivo.
In vivo pharmacokinetics and biodistributions of siRNA@eS-BNVs
The tumor accumulation and penetration are crucial requirements for the in vivo application of siRNA@eS-BNVs. Firstly, we confirmed the ACE2 receptor expressed in the A549ACE2+ tumors by immunofluorescence (IF) staining, as it can interact with eS-BNVs for specific gene delivery (Supplementary Fig. 25). Considering Thr enzyme is utilized for initiating membrane fusion of eS-BNVs, its biodistribution in tumors were assessed by i.v. injection of Alexa647-labeled Thr (i.e., ThrAlexa647) to A549ACE2+ tumor-bearing mice. As shown in Supplementary Fig. 24a, b, ThrAlexa647 was efficiently accumulated in tumors at 1 h and 2 h, as monitored by IVIS spectrum. To study the biodistribution, the siRNA-Cy5@eS-BNVs were i.v. injected into the A549ACE2+ tumor-bearing mice and imaged by IVIS spectrum. As shown in Fig. 5a, the siRNA-Cy5@eS-BNVs + Thr were effectively and rapidly accumulated in the A549ACE2+ tumors, which effectively improved the tumor accumulation of siRNA. However, the free siRNA-Cy5 and siRNA-Cy5/PEI was unable to accumulate in tumors. Then, the blood, major organs and tumors were obtained and analyzed by the IVIS spectrum. The siRNA-Cy5@eS-BNVs and siRNA-Cy5@eSWT-BNVs demonstrated higher accumulation in tumors than that of siRNA-Cy5@No-BNVs (Fig. 5b, c), possibly due to the targeting effects of S protein in nanovesicles to the ACE2 receptor expressed in A549ACE2+ tumors. The siRNA-Cy5@eS-BNVs+Thr exhibited even much higher tumor accumulation than siRNA-Cy5@eS-BNVs and siRNA-Cy5@eSWT-BNVs due to efficient membrane fusion with cancer cells. Of note, siRNA-Cy5@eS-BNVs+Thr accumulated in the A549ACE2+ tumors was the highest than that of other organs (e.g., heart, liver and lung, etc.), further indicating its good tumor targeting efficacy (Fig. 5b, c). A more detailed comparison by ex vivo IVIS spectrum images indicated that the siRNA-Cy5@eS-BNVs+Thr group distributed less in the main organs compared with tumors (Fig. 5c), indicating its ligand-receptor-mediated ability. The highest tumor accumulation of siRNA-Cy5@eS-BNVs was observed at 12 h compared to the other samples, such as siRNA-Cy5@eS-BNVs and siRNA-Cy5@eSWT-BNVs (Supplementary Fig. 26a, b). Therefore, we chose to inject Thr enzyme at 12 h after administration of siRNA@eS-BNVs to trigger membrane fusion in the follow-up anti-tumor experiment. Moreover, the in vivo pharmacokinetics of nanovesicles and siRNA were studied (Supplementary Fig. 26c). Free siRNA-Cy5 was rapidly cleared from the blood circulation, while siRNA-Cy5@eS-BNVs demonstrated longer retention time and higher levels in the blood circulation than free siRNA-Cy5 (Supplementary Fig. 26c). Therefore, the siRNA@eS-BNVs remarkably prolonged the blood half-life of siRNA and promoted its accumulation in solid tumors.

a In vivo biodistribution of Cy5-labeled siRNA@eS-BNVs in A549ACE2+ tumor-bearing mice was measured by IVIS spectrum at 2, 12 and 24 h post i.v. injection. b Ex vivo imaging of Cy5-labeled siRNA@eS-BNVs distributed in tumors at 24 h post i.v. administration. c Quantification of Cy5-labeled siRNA@eS-BNVs in main organs and tumors at 24 h after i.v. injection of nanovesicles (n = 3 independent samples). d Extravasation of Cy5-labeled siRNA@eS-BNVs from blood vessels into tumor tissues at 24 h post i.v. injection. Scale bars, 50 μm. e Representative fluorescence images show the distribution of Cy5-labeled siRNA@eS-BNVs across the A549ACE2+ tumors at 24 h after i.v. administration. Green, A549ACE2+-EGFP tumors; red, siRNA-Cy5; white, blood vessel stained by anti-CD31 antibody. Scale bars, 100 μm. Avg, average. Experiments in (d and e) were repeated three times independently with similar results. Data in (c) is the mean ± s.d. p-values were determined by a two-way ANOVA test in (c). ****p < 0.0001. Source data are provided in a Source Data file.
Next, the extravasation and penetration of the siRNA-Cy5 from blood vessels into 549ACE2+-EGFP tumor tissues were investigated by immunofluorescent staining of the blood vessels. The siRNA-Cy5@eS-BNVs, siRNA-Cy5@eSWT-BNVs, and siRNA-Cy5@eS-BNVs + Thr efficiently extravasated from blood vessels into distant tumor tissues, e.g., up to about 200 μm far from blood vessels. However, there was almost no free siRNA-Cy5 and siRNA-Cy5/PEI extravasated from blood vessels into tumors (Fig. 5d). In addition, to visually examine the penetration and microdistribution of siRNA@eS-BNVs in tumors, the A549ACE2+-EGFP tumors were resected at 24 h after i.v. injection of nanovesicles, and were made into slices. As shown in Fig. 5e, the siRNA-Cy5@eS-BNVs + Thr could penetrate deep tumor regions and almost homogenously distribute across whole A549ACE2+ tumors, associating with constant fluorescence intensity across tumor sections (Fig. 5e). However, rare free siRNA-Cy5 or siRNA-Cy5/PEI was penetrated the tumor tissues (Fig. 5e). The CLSM images also showed that the siRNA-Cy5@eS-BNVs + Thr appeared a distinct yellow area, indicating that the siRNA-Cy5@eS-BNVs + Thr could penetrate more into cancer cells compared with siRNA-Cy5@eS-BNVs and siRNA-Cy5@eSWT-BNVs. As further investigated by IntraVital Microscope, the siRNA-Cy5@eS-BNVs also demonstrated efficient extravasation and accumulation in tumor tissues (Supplementary Fig. 27). The efficient extravasation from blood vessels and deep penetration in tumors would further benefit tumor therapy by siRNA@eS-BNVs.
siEGFR@eS-BNVs for effective tumor therapy by silencing EGFR in tumors
EGFR is a critical target for tumor therapy because it is highly expressed in several tumors, especially in lung cancer. Although monoclonal antibodies can target EGFR expressed on cancer cells to inhibit tumor proliferation15,37, there are still many side effects in the clinical applications. Here, we encapsulated siEGFR in the eS-BNVs (i.e., siEGFR@eS-BNVs) for tumor therapy by interfering with EGFR in the A549ACE2+ cancer cells, which also overexpressed EGFR on cancer cell surfaces38. We also engineered siCtrl@eS-BNVs as a control sample. Considering i.v. injection of siLuc/PEI multiple times even at a low dose caused mice death (Supplementary Fig. 23), we did not use siLuc/PEI as the control in antitumor experiments.
The subcutaneous (s.c.) A549ACE2+ tumor-bearing mice were i.v. injected with siEGFR@eS-BNVs and control samples (i.e., PBS, free siEGFR, siEGFR@No-BNVs, and siCtrl@eS-BNVs) on days 10, 12, 14, 16, and 18, while the Thr enzyme (1.5 U per mouse) was i.v. administrated at 12 h after drug administration to give the treatments of siCtrl@eS-BNVs + Thr and siEGFR@eS-BNVs + Thr (Fig. 6a). As shown in Fig. 6b, c and Supplementary Fig. 28, siEGFR@eS-BNVs + Thr significantly inhibited the growth of A549ACE2+ tumors, demonstrating the lowest tumor growth rate and smallest tumor volume at the end point compared to siEGFR@eS-BNVs, siEGFR@No-BNVs and siCtrl@eS-BNVs + Thr, etc. In addition, after i.v. injecting Thr enzyme into A549ACE2+ tumor-bearing mice, the average tumor volume was similar to the PBS group (Supplementary Fig. 24c), indicating that Thr enzyme cannot inhibit tumor growth rate, consistent with the previous studies39,40. There was no significant change in the body weight of the mice (Supplementary Fig. 24d), indicating the safety of the Thr enzyme. Considering the injection of Thr enzyme at this dose did not affect tumor growth and cause safety problems in mice39,40, Thr enzyme was not applied as a control sample. Moreover, the survival rates were monitored, while siEGFR@eS-BNVs + Thr exhibited longer survival time than the control groups, e.g., siCtrl@eS-BNVs + Thr (Fig. 6d). Of note, there was no big change in the body weight of mice almost during the treatments, suggesting the safety of eS-BNVs (Supplementary Fig. 29). These results demonstrated that siEGFR@eS-BNVs with Thr-triggered membrane fusion function could effectively eradicate A549ACE2+ tumors.

a Scheme shows in vivo tumor therapy by i.v. administration of siEGFR@eS-BNVs to A549ACE2+ tumor-bearing mice on days 10, 12, 14, 16 and 18, and then injected with Thr enzyme by i.v. injection (1.5 U per mouse) at 12 h after drug administration. Created in BioRender. Cui, L. (2025) https://BioRender.com/umqxomy. b, c Average tumor growth rates (b) and individual tumor growth rate (c) of A549ACE2+ tumors after treating by siEGFR@eS-BNVs + Thr (n = 6 independent samples). d Survival rates of A549ACE2+ tumor-bearing mice after treating by siEGFR@eS-BNVs + Thr and control samples. e Western blot and statistically analyzing the EGFP receptor expressed in A549ACE2+ tumors were tested on day 16 (n = 6 independent samples). fEGFR mRNA levels expressed in the A549ACE2+ tumors as tested on day 16 (n = 6 independent samples). g Immunofluorescent staining of EGFR expressed in the A549ACE2+ tumors after the above treatments. Red, EGFR stained by anti-EGFR antibody; blue, nucleus stained by DAPI. Scale bars, 500 μm. Experiment in (g) was repeated three times independently with similar results. Data in (b, e and f) are the mean ± s.d. p-values were determined by two-tailed Student’s t test in (b) or one-way ANOVA test in (e, f). ****p < 0.0001, ***p < 0.001, *p < 0.05. Source data are provided in a Source Data file.
Next, we resected the tumor tissues and quantified the EGFR expression in the treated A549ACE2+ tumors. As characterized by western blot, siEGFR@eS-BNVs + Thr significantly downregulated the expression of EGFR in A549ACE2+ tumors (Fig. 6e), while other treatments such as siEGFR@eS-BNVs and siEGFR@No-BNVs showed less effectiveness in silencing the EGFR in the A549ACE2+ tumors. On the contrary, free siEGFR and siCtrl@eS-BNVs + Thr were unable to silence EGFR in the A549ACE2+ tumors. Moreover, the EGFR mRNA levels in A549ACE2+ tumors were measured byRT-qPCR (Fig. 6f), which also demonstrated the lowest EGFR mRNA level downregulated by siEGFR@eS-BNVs + Thr compared to the controls, e.g., siEGFR@eS-BNVs + Thr. In addition, the EGFR in tumors was characterized by immunofluorescence staining. As shown in Fig. 6g, the effects of silencing EGFR by siEGFR can be seen across the whole A549ACE2+ tumor tissues, and siEGFR@eS-BNVs + Thr yielded the least EGFR expression across whole tumors compared to other groups. Therefore, Thr-controlled membrane fusion of siEGFR@eS-BNVs can robustly silence EGFR in solid tumors to facilitate effective tumor therapy.
siEGFR@eS-BNVs for effectively inhibiting lung metastasis
Inspired by the antitumor effects of siEGFR@eS-BNVs in vivo, we further applied it to inhibit lung metastasis of A549ACE2+-Luc cancer cells (Fig. 7). The lung metastasis tumor model was established by i.v. injection of A549ACE2+-Luc cancer cells on day 0 and monitored by IVIS spectrum. The lung metastasis-bearing BALB/c nude mice were i.v. injected with PBS, free siEGFR, siCtrl@eS-BNVs, siEGFR@No-BNVs and siEGFR@eS-BNVs on days 10, 12, 14, 16 and 18, while Thr enzyme (1.5 U per mouse) was i.v. injected into two groups of mice at 12 h to receive the treatments of siCtrl@eS-BNVs+Thr and siEGFR@eS-BNVs+Thr (Fig. 7a). Of note, i.v. injection is commonly used for drug administration, and nanocarriers by i.v. injection have the opportunity to reach both primary and metastatic tumors. As shown in Fig. 7b, c, siEGFR@eS-BNVs + Thr effectively inhibited the development of lung metastasis, exhibiting the lowest bioluminescence intensity in lungs compared to other groups, such as siEGFR@eS-BNVs and siEGFR@No-BNVs, etc. Of note, some mice died in the control groups, but all mice survived in the siEGFR@eS-BNVs + Thr group within 35 days. The survival rates were also recorded, and siEGFR@eS-BNVs + Thr also exhibited longer survival time than the other groups, such as siCtrl@eS-BNVs + Thr and siEGFR@eS-BNVs (Fig. 7d). The survival rates between the siEGFR@eS-BNVs + Thr and siEGFR@eS-BNVs groups were not so significantly different, which may be attributed to the termination of drug administration until day 18, and the effects of the drugs were unable to maintain for long time (e.g., 50–60 days). Because the in vivo half-life of cell membrane-modified nanoparticles was approximately 40 h41 and the RNAi effects generally persist for 3–7 days3. Our additional experiments demonstrated that the RNAi effects of siRNA@eS-BNVs + Thr maintained for approximately 7 days (Supplementary Fig. 30). Of note, there was no big change in the body weight of mice during the treatment period, suggesting that the safety of biomimetic nanovesicle-based treatments (Supplementary Fig. 31).

a Scheme shows the eradication of lung metastasis of A549ACE2+-Luc by i.v. administration of siEGFR@eS-BNVs on days 10, 12, 14, 16 and 18, and i.v. injection of Thr at 12 h after drug administration. Created in BioRender. Cui, L. (2025) https://BioRender.com/umqxomy. b Bioluminescence imaging of lung metastasis of A549ACE2+-Luc in mice by IVIS spectrum. c Bioluminescence intensity of lung metastasis measured in panel (b). (n = 5 independent samples). d Survival rates of lung metastatic tumor-bearing mice after treating with siEGFR@eS-BNVs and control samples. e Western blot and statistical analysis of EGFR levels in lungs with A549ACE2+-Luc metastatic tumors (n = 6 independent samples). f RT-qPCR analysis of EGFR mRNA levels in lungs with A549ACE2+-Luc metastatic tumors (n = 6 independent samples). g Representative photographs of metastatic A549ACE2+-Luc tumors in lungs after treatments. Scale bar, 2 mm. h, i Representative H&E staining of lungs with metastatic tumors (h) and area of metastatic tumors (i). Scale bars, 2 mm. (n = 6 independent samples). j, k Representative bioluminescence images (j) and intensity (k) of lung metastasis. Scale bar, 2 mm. (n = 6 independent samples). l Representative IHC staining of EGFR in A549ACE2+-Luc lung metastatic tumors. Scale bars, 2 mm. Avg, average. Data in (c) is mean ± s.e.m. Data in (e, f, h and k) are mean ± s.d. p-values were determined by one-way ANOVA test in (c, e, f, i, k). ****p < 0.0001, *p < 0.05, ns, no significant. Source data are provided in a Source Data file.
Next, the lungs were resected to characterize the EGFR expression and pathological features of lung metastases. Firstly, metastatic tumors showed remarkably higher expression of ACE2 receptor in the A549ACE2+ tumor regions than that of the normal lung tissues (Supplementary Fig. 32a, b), which could be efficiently targeted by the siEGFR@eS-BNVs. Next, the EGFR expression in lung metastases was assessed by western blot (Fig. 7e), and siEGFR@eS-BNVs + Thr remarkably decreased the EGFR levels compared to other groups, e.g., siEGFR@eS-BNVs, etc. However, the EGFR expression was not reduced in the free siEGFR and siCtrl@eS-BNVs groups compared to the PBS group. Moreover, the EGFR mRNA in lung metastases measured by RT-qPCR also revealed that siEGFR@eS-BNVs + Thr effectively downregulated EGFR, exhibiting the lowest level of EGFR mRNA than the other treatments (Fig. 7f). Thus, those results provided that lung metastases could be effectively inhibited by efficiently silencing EGFR via Thr-controlled delivery of siEGFR by eS-BNVs.
The metastatic tumors in the lungs were checked and imaged. As shown in Fig. 7g, treatment by siEGFR@eS-BNVs + Thr yielded the lowest number of metastatic A549ACE2+-Luc tumors in the lungs compared to the other groups, which had lots of lung metastases, e.g., siEGFR@No-BNVs and siCrtl@eS-BNVs + Thr, etc. Then, the hematoxylin and eosin (H&E) staining of the lungs further clearly indicated that rare metastatic foci were found in the lungs when treated by siEGFR@eS-BNVs + Thr, while many big metastatic tumors were found in the other groups (Fig. 7h, i and Supplementary Fig. 33). Next, the bioluminescence of A549ACE2+-Luc cancer cells in lungs was imaged by IVIS spectrum, exhibiting the lowest bioluminescence intensity in the siEGFR@eS-BNVs + Thr group than the others (Fig. 7j, k). Moreover, the expression of EGFR in the lungs was characterized by immunohistochemistry (IHC) staining, which further demonstrated the lowest EGFR expression in lungs knocked down by the siEGFR@eS-BNVs + Thr treatment than the others (Fig. 7l). Together, siEGFR@eS-BNVs + Thr can effectively inhibit lung metastasis by silencing the EGFR expression through functions of ligand-receptor-mediated and Thr-controlled membrane fusion for intracellular siRNA delivery.
In addition, the safety was evaluated by i.v. injection of siRNA@eS-BNVs into mice. The blood test was conducted, and no abnormal events were found in the indicators of alanine aminotransferase (ALT), aspartate aminotransferase (AST), blood urea nitrogen (BUN), creatinine (CREA) and lactate dehydrogenase (LDH) compared to the PBS control (Supplementary Fig. 34). Moreover, H&E staining of the major organs revealed no damage to those organs caused by treating with siRNA@eS-BNVs and the control samples (Supplementary Fig. 35). There, these results demonstrated the safety of eS-BNVs as a drug delivery system for in vivo applications.
Discussion
Tumor therapy by knocking down oncogenes via RNAi is heavily restricted by the delivery of siRNA to tumors and its trafficking into the cytosolic compartment of cancer cells due to the extracellular excretion by circulation, endocytic uptake and RNase, poor intracellular trafficking, and lysosome degradation42. In this study, we have genetically engineered biomimetic nanovesicles, eS-BNVs, with ligand-receptor-mediated and enzyme-controlled membrane fusion functions for efficiently delivering siRNA into cancer cells to inhibit tumors in animal models effectively. The present eS-BNVs were rationally engineered with several critical functions, which enabled addressing the barriers beyond the successful delivery of siRNA into cancer cells, as well as knocking down specific mRNA and proteins in vitro and animal models.
Firstly, the eS-BNVs were bioengineered with S protein, which can specifically internalize with the ACE2 receptor expressed on cancer cells for ligand-receptor-mediated drug delivery and membrane fusion. Secondarily, this study has systemically illustrated the Thr enzyme-triggered membrane fusion between different S protein-decorated nanovesicles (e.g., eSDM-BNVs, eSWT-BNVs and eS-BNVs) and NVsACE2+, and A549ACE2+ cancer cells (Fig. 2 and Supplementary Fig. 10). Although conventional membrane-made nanovesicles (e.g., No-BNVs) may have weak membrane fusion function, they are unable to achieve cancer cell-targeted membrane fusion43. Therefore, the introduction of tumor cell-targeting fusogen to nanocarriers is an effective approach for initiating membrane fusion with specific cancer cells. This study presented an enzyme-controlled membrane fusion approach by leveraging the S protein in eS-BNVs to target cancer cells and triggering membrane fusion when the S protein was cleaved by the Thr enzyme. Although membrane fusion proteins have been studied before, such as fusogen, there are still areas that need improvement, such as controlling membrane fusion and tumor targeting. Herein, the spike protein can achieve both targeted effects and rapid intracellular delivery by membrane fusion, which is crucial for cytosolic siRNA delivery. Thirdly, eS-BNVs indeed exhibited remarkably higher efficacy for cytosolic siRNA delivery to cancer cells via enzyme-controlled membrane fusion than the conventional nanocarriers, e.g., siRNA-Cy5/PEI and No-BNVs, achieving better RNAi effects (Fig. 4). Fourthly, we delivered siEGFR by eS-BNVs with enzyme-triggered membrane fusion for tumor therapy, as EGFR is an important biomarker and therapeutic target in several cancers, especially lung cancer. The i.v. administration of siEGFR@eS-BNVs effectively eradicated solid tumors (Fig. 6b–d) and lung metastasis (Fig. 7b–d) as a result of silencing EGFR mRNA (Figs. 6f and 7f) and knocking down EGFR (Fig. 6e, g and 7e, l) in the tumors.
Besides this work, other delivery systems such as LNPs are also utilized for siRNA delivery44, but there are differences between LNPs and our system: (1) Different materials, as LNPs generally contain four compositions such as ionizable lipid, helper lipid, PEGylated lipid and cholesterol, while our system uses bioengineered cell membranes as vehicles; (2) Different functions, as LNPs mainly use ionizable lipid for endosomal escape and intracellular siRNA delivery, while our system utilizes membrane fusion for directly intracellular siRNA delivery; (3) Different formulation methods. Moreover, other nanocarriers with membrane fusion function have been reported, such as bioinspired engineering of fusogen and GPC3-targeted single-chain variable fragment (scFv) co-expressing cell-derived nanovesicles CNVs (eFT-CNVs)20, fusion-associated small transmembrane-proteolipid vehicle (FAST-PLV)45, cholesterol-enriched milk-derived exosomes (MEs)35, demonstrating efficient gene or drug delivery efficacy in dealing with diseases. For instance, the eFT-CNVs were prepared using fusogen as a membrane fusion protein and anti-GPC3 scFv as the targeting moiety for efficient intracellular gene delivery via membrane fusion and effective cancer therapy20. Different from those works, our work mainly focuses on bioengineering cell membranes anchored with mutated S proteins for enzyme-controlled membrane fusion. The drug delivery materials, such as mutated S proteins, cargos and specific applications, are also different from the above studies. Besides S proteins, other membrane fusion proteins may are also be genetically engineered on the cell membranes to achieve membrane fusion functions, such as the spike proteins of MERS-CoV, hCoV-229E, hCoV-HKU1, and SARS-CoV-2 Delta, etc. Moreover, this study focuses on using the spike protein RBD domain to target the ACE2 receptor, which may limit their applications for dealing with other types of tumors without the expression of the human ACE2 receptor. For future clinical translation purposes, its interaction with endogenous cells that also express human ACE2 should be carefully considered and investigated. This can be improved by replacing the RBD region with tumor-targeting peptides such as anti-GPC3 scFv20. Furthermore, other stimuli may further be applied for triggering membrane fusion, e.g., modifying the linker in spike protein sensitive to other compounds, such as 3 C protease. Finally, considering people may be affected by SARS-CoV-2 or taken vaccines, which may affect the efficacy of eS-BNVs in those persons. Certain solutions may be applied to solve such kind of application situations: (1) Administration of eS-BNVs after 10 months or later, as studies have reported that IgA antibodies produced by SARS-CoV-2 infection do not last more than 9 months46; (2) Patient selection can be performed before administrating eS-BNVs, which is conventionally applied in clinic for most drugs; (3) Replacing the RBD region with a tumor-targeting peptide.
Taken together, we successfully developed biomimetic eS-BNVs encoding with cancer cell-targeting and enzyme-activatable fusogen protein for efficient cytosolic delivery of siRNA and silencing oncogenes, leading to effective tumor therapy. This work reported here represents the practical case of enzyme-controlled membrane fusion of nanovesicles for drug delivery. The biomimetic eS-BNVs offer a powerful tool for delivering other bioactive compounds, such as mRNA, DNA, proteins, and anticancer drugs to overcome the systemic or cellular barriers ahead of drug delivery and precision medicine7,25,35,45. The cell-targeting and enzyme-activatable nanovesicles not only provide a valuable tool for drug delivery but also a paradigm for developing nanocarriers for precision medicine in the future.
Methods
Cells and animals
A549 (ATCC Number: CCL-185), HEK293T (ATCC Number: CRL-11268), HUVEC (ATCC Number: CRL-4053) and Calu3 (ATCC Number: HTB-55) cell lines were acquired from the American Type Culture Collection (ATCC, USA). A549ACE2+, A549ACE2+-Luc, A549ACE2+-EGFP, A549ACE2-, HEK293TACE2- and eS-HEK293TACE2-, eSWT-HEK293TACE2- and eSDM-HEK293TACE2- were constructed via lentiviruses. HEK293T, HUVEC (single donor) and Calu3 cell lines were cultured in Dulbecco’s Modified Eagle Medium (DMEM, Cat no: 11965092, Gibco). A549 cancer cell lines were cultured in RPMI Medium 1640 basic (Cat no: 11875093, Gibco). Medium was supplemented with 10% fetal bovine serum (FBS, Cat no: FSP500, ExCell) and penicillin-streptomycin (Cat no: C0222, Beyotime) at 37 °C in a humidified atmosphere of 5% CO2. The cells were routinely passed by mycoplasma testing. BALB/c nude mice and C57BL/6 mice (female, 6 weeks) were purchased from the Gempharmatech Co., Ltd. All animal experiments complied with the protocol approved by the Sichuan University Committee on Animal Care Committee (No.: 20230302063).
Plasmid construction and cell lines editing
The codon-optimized full-length spike gene sequence in the pCAGGS vector was kindly provided by Prof. Guangwen Lu at Sichuan University. The codon-optimized human ACE2 receptor gene sequence (UniProt: Q9BYF1) was synthesized by Youkang Biotechnology Co., Ltd (Zhejiang, China). Point mutations were employed to modify the cleavage sites of the spike protein. Spike gene fragments (amino acids 20-1268) and ACE2 gene fragments (amino acids 18-761) were cloned into the EcoRI and BamHI linearized pLVX-Hyg or pLVX-Puro vector. The plasmid map of S protein was shown in Supplementary Fig. 1. Successful plasmid construction was confirmed through gene sequencing. Other plasmids, including ACE2-shRNA-Puro (Cat no: TR306881, Origene), S-pLVX-Hyg, SWT-pLVX-Hyg, SDM-pLVX-Hyg, human-ACE2-pLVX-Puro, EGFP-pLVX-Hyg and Luc-pLVX-Hyg were also constructed using lentiviral transfection approach. Lentiviral plasmid, packaging plasmid psPAX2 and envelope plasmid pMD2.G were transfected into HEK293T cells by Lipo8000 for lentiviral preparation when they reached 70–80% confluence. Transfected cell supernatants were collected at 48 h after transfection and purified using a 0.45 μm filter membrane. The virus was immediately transferred into target HEK293T cells and A549 cancer cells. eS-HEK293TACE2-, eSWT-HEK293TACE2-, eSDM-HEK293TACE2-, HEK293TACE2-, A549ACE2+, A549ACE2+-EGFP and A549ACE2+-Luc cells were generated by further screening with puromycin (Puro, Cat no: ST551, Beyotime) or hygromycin (Hyg, Cat no: ST1389, Beyotime). Finally, cell surface proteins were verified by western blot (TOUCH IMAGER, eBlot), and the corresponding data were processed by eBLOT14. Briefly, the eS-HEK293TACE2- cells and HEK293T cells were lysed using radio immunoprecipitation assay (RIPA, Cat no: P0013C, Beyotime). Cell samples were separated by Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE, PG111, Epizyme). The processed cell membrane sample was added to the loading well of the gel and subjected to electrophoretic separation. The electrophoresed proteins were transferred to a polyvinylidene difluoride membrane (PVDF, Cat no: 1620177, Bio-Rad) on a solid phase carrier. The membranes were blocked with 5% skim milk solution to reduce nonspecific binding. Then, the PVDF membrane were incubated with spike RBD antibody (Cat no: 40592-T62, SinoBiological) that diluted in 5% skin milk (1:1000 dilution) at 4 °C overnight. Then, PVDF membrane were incubated with the second antibody (Cat no: AS014, ABclonal) for 2 h at room temperature. The signal was enhanced by chemiluminescence (Cat no: PD202, Oriscience) and imaged with a western imaging instrument (TOUCH IMAGER, eBlot). Moreover, 1 × 106 eS-HEK293TACE2- cells and HEK293TACE2- cells were identification by flow cytometry. The cells were collected and incubated with spike S1 antibody (Cat no: 945101, BioLegend) that diluted in PBS (1:100 dilution) for 30 min at 4 °C. Then, cells were incubated with FITC anti-rat IgG (Cat no: 405404, BioLegend) diluted in 100 μl PBS (1:100 dilution) for 30 min at 4 °C. The representative gating strategy for the flow cytometry analysis is shown in Supplementary Fig. 2c. Next, cells were performed with flow cytometry (NovoCyte, Agilent) and the corresponding data were processed with FlowJo_V10.
Cell membrane collection and eS-BNVs preparation
eS-HEK293TACE2-, eSWT-HEK293TACE2-, eSDM-HEK293TACE2-, HEK293TACE2-, and A549ACE2+ cell membranes were collected using a modified version of gradient centrifugation method47. Various cells were resuspended in HEPES buffer (20 mM HEPES, 150 mM NaCl, pH 7.4, 1 mM EDTA) containing a proteinase phosphatase inhibitor cocktail and pre-cooling on ice for 30 min. The pellets were disrupted with a homogenizer (KZ-III-96, Servicebio). The cell membranes were purified by differential centrifugation. In brief, the homogenized solution was centrifuged at 20,000 × g at 4 °C for 20 min to remove cellular detritus. Then, the supernatant was collected after ultra-centrifugation at 100,000 × g at 4 °C for 1 h (Sorvall™ WX + , Thermo Fisher). Subsequently, the purified cell membranes were collected and washed with HEPES buffer twice and quantified using the bicinchoninic acid (BCA, Cat no: P0010, Beyotime) assay.
The eS-BNVs, eSWT-BNVs, eSDM-BNVs and No-BNVs were synthesized using eS-HEK293TACE2-, eSWT-HEK293TACE2-, eSDM-HEK293TACE2- and HEK293TACE2- cellular membranes, respectively, with a sonicator bath (KQ-400KDE, Shumei) for 6 min and coextruded through polycarbonate filters (800, 400 and 100 nm pore size, Avanti Polar Lipids) using a mini extruder (Avanti)41.
Preparation and characterization of siRNA@eS-BNVs
The siRNA (Synthesized by Sangon Biotech) dissolved in HEPES buffer (13.3 μg, 4 μM) was mixed with an equal volume of positively charged polymer PEI (9 μg) (Cat no: 765090, 10 kDa, linear, Sigma) dissolved in HEPES buffer at a N/P (amine nitrogen group in PEI, phosphate group in siRNA) mole ratio of 5:1 by vortex, and then incubated for 20 min to form siRNA/PEI complexes. Subsequently, the siRNA/PEI complexes were mixed with eS-BNVs and treated by gentle sonication at 4 °C for 1 h to obtain siRNA-encapsulated nanovesicles (i.e., siRNA@eS-BNVs). Similarly, the siRNA/PEI complexes were mixed with No-BNVs to engineer siRNA@No-BNVs as a control. These nanoparticles were finally purified by passing through a 100-kDa molecular cutoff filter (Millipore) and centrifugation at 2000 × g by HEPES buffer to purify siRNA@eS-BNVs and siRNA@No-BNVs. Whether siRNA could completely form complexes with PEI and whether siRNA/PEI nanoparticles were completely encapsulated by BNVs were verified by SYBR Gold assay. The DLS hydrodynamic diameters and polydispersity index (PDI) of eS-BNVs and No-BNVs were measured by Zetasizer Nano ZS90 (Malvern Instruments), and the corresponding data were processed by ZetasiZer software. The morphology of eS-BNVs and No-BNVs was characterized by TEM at 120 k voltage (JEM-1400, JEOL) after staining the samples with 1.5% (w/v) uranyl acetate. Images were acquired using RADIUS software. The S protein on the surface of nanovesicles was characterized by western blot.
Analyzing siRNA encapsulation efficacy by single-molecule quantified assay
The single-molecule assay was employed to determine the encapsulation efficiency of siRNA encapsulated in siRNA@eS-BNVs according to the previous reports48,49. The polyethylene glycol (PEG)-coated quartz slide flow chambers were coated with PEG and biotin-PEG to obtain Biotin-PEG modified slides. Purified eS-BNVs suspended in HEPES buffer were used to re-resuspended the dried lipid film, added with Biotin-PE and incubated for 30 min at room temperature to prepare eS-BNVsBiotin-PE. The siRNA-Cy5 was encapsulated in eS-BNVsBiotin-PE to obtain siRNA-Cy5@eS-BNVsBiotin-PE. The siRNA-Cy5@eS-BNVsBiotin-PE were immobilized on slides via neutravidin.
The siRNA-Cy5@eS-BNVsBiotin-PE were immobilized on the PEG-coated surface during a 30 min incubation via neutravidin. Then, the free siRNA-Cy5@eS-BNVsBiotin-PE were washed with wash buffer (20 mM HEPES, 100 mM NaCl, pH 7.4). The siRNA-Cy5@eS-BNVsBiotin-PE were immobilized on the slides by the interaction between neutravidin and biotin. Firstly, we find the appropriate region by low light under the microscope field. To quantify the siRNA-Cy5 encapsulation assay, the fluorescence quenching of siRNA-Cy5 was observed after strong fluorescence irradiation, and the images or videos of siRNA-Cy5@eS-BNVsBiotin-PE were obtained from a total internal reflect (TIR) fluorescence microscope (Ti2-E, Nikon)50 and the data were recorded by NIS-elements AR. Then, smCamera was used to analyze the curve of the change of the fluorescence signal value of a single vesicle in the field of view. Each decrease in the signal represented the occurrence of a siRNA-Cy5 fluorescence quenching, and the number of fluorescence quenching events in a vesicle represented the number of siRNA-Cy5 in a vesicle.
Gel retardation assay
To confirm the successful preparation of siRNA/PEI coated with eS-BNVs (i.e., siRNA@eS-BNVs), we performed a gel retardation assay. Briefly, siRNA/PEI and siRNA@eS-BNVs were prepared, then incubated with either heparin or heparin + Triton X-100 for 30 min at room temperature. The complexes were mixed with 6X gel loading buffer (Cat no: B7024S, NEB), separated by 1% agarose gel (Cat no: 1110GR100, BioFroxx) alongside a DNA ladder (Cat no: DM007, Novoprotein), and used SYBR Gold nucleic acid stain (Cat no: S11494, Invitrogen) for visualization. Gel images were acquired using a ChemiDoc gel imager (ChemiDoc XRS +, Bio Rad).
Interaction between eS-BNVs and A549ACE2+ cancer cells
To determine the interaction between eS-BNVs and cancer cells, the A549ACE2+ cancer cells, A549ACE2- cancer cells and Calu3 cancer cells were seeded into 6-well plates (1 × 106 cells per well) for 24 h and then exposed to PBS, No-BNVsDiI, eS-BNVsDiI and eSWT-BNVsDiI for 30 min, respectively. Next, the cells were detached, washed with PBS, and analyzed by flow cytometry (NovoCyte, Agilent). Data analysis and gating strategy were performed by NovoExpress software and FlowJo_V10. The representative gating strategy for the flow cytometry analysis is shown in Supplementary Figs. 6a, b, and 7c. The binding efficiency between A549ACE2+ cancer cells and eS-BNVs or No-BNVs was also investigated by western blot (TOUCH IMAGER, eBlot) and the corresponding data were processed with eBLOT14. Finally, the CLSM (Zeiss LSM 880) was used to observe the localization of eS-BNVsDiI to A549ACE2+ cancer cells and A549ACE2- cancer cells.
Studying the enzyme-triggered membrane fusion between eS-BNVs and A549ACE2+ cancer cells in vitro
An ensemble content-mixing assay was used to assess the effect of the S protein on membrane features. The in vitro fusion studies for A549ACE2+ cancer cells, A549ACE2- cancer cells, Calu3 cancer cells, HUVEC cells, HEK293T cells, NVsACE2-, NVsACE2+ to eS-BNVs, eSDM-BNVs and eSWT-BNVs were studied by measuring real-time fluorescence spectrophotometer (Agilent Cary Eclipse, Agilent) for about 2700–3000 s, and the corresponding data were processed with Cary WinFLR. For the ensemble content-mixing assay, the self-quenched SRB (Cat no: S1307, Invitrogen) molecule served as a content indicator and was encapsulated in eSDM-BNVs, eSWT-BNVs and eS-BNVs to obtain SRB@eSDM-BNVs, SRB@eSWT-BNVs and SRB@eS-BNVs51. SRB@eSDM-BNVs, SRB@eSWT-BNVs and SRB@eS-BNVs were formed in 50 mM SRB by soft ultrasound and dialyzed to remove the excess SRB dye. Content-mixing was measured by the increased fluorescence emission at 585 nm of the SRB dye upon excitation at 532 nm.
The real-time membrane fusion processes between eS-BNVs and A549ACE2+-BNVs was observed by TEM (JEM-1400, JEOL). The size of A549ACE2+-BNVs was controlled by extruding in a 100 nm polycarbonate membrane. Anti-rabbit lgG antibody nanogold (Cat no: 2003, Nanoprobes) in 1.4 nm diameter was conjugated to anti EGFR antibody (Cat no: ET1604-44, Huabio) to label A549ACE2+-BNVs. The labeled A549ACE2+-BNVs were purified by ultracentrifugation. An equal volume of the eS-BNVs and nanogold-labeled A549ACE2+-BNVs were mixed, and the Thr enzyme (Cat no: T4648, Sigma) was added to initiate the membrane fusion process. The samples were stained with uranium acetate and observed by TEM (JEM-1400, JEOL). Images were acquired using RADIUS software.
Cellular uptake of siRNA-Cy5@eS-BNVs by A549ACE2+ cancer cells, HUVEC and HEK 293 T cells
To detect eS-BNVs mediated siRNA delivery in vitro, A549ACE2+ cancer cells (2 × 104 cells per dish, 1 ml) were seeded in 2 cm diameter dishes and cultured in DMEM supplemented with 10% FBS for 24 h. Then, they were exposed to siRNA-Cy5@eS-BNVs+Thr (150 nM, based on siRNA) at 37 °C and imaged at 10 min, 30, 60, 120 min. The same conditions were applied to control experiments with PBS, free siRNA-Cy5, siRNA-Cy5/PEI, siRNA-Cy5@No-BNVs siRNA-Cy5@eSWT-BNVs, siRNA-Cy5@eSWT-BNVs + Thr and siRNA-Cy5@eS-BNVs. Subsequently, A549ACE2+ cancer cells were washed with PBS, fixed with 4% paraformaldehyde phosphate buffer (PFA), and the nucleus was labeled by 4’,6-Diamidino-2’-phenylindole (DAPI, Cat no: C0065, Solarbio). The subcellular localization of siRNA-Cy5 was analyzed by CLSM (Zeiss LSM 880), and the corresponding data were collected by ZEN 3.6 software.
Next, we assessed the cellular uptake of siRNA-Cy5@eS-BNVs by HUVEC and HEK293T normal cells with CLSM. The HUVEC and HEK293T cells (2 × 104 cells per dish) seeded in 2 cm dishes were treated with PBS, siRNA-Cy5@eS-BNVs or siRNA-Cy5@eS-BNVs + Thr (150 nM, based on siRNA) for 2 h at 37 °C. Then, the cells were washed three times with PBS, fixed with 4% PFA, and stained with DAPI to be subsequently analyzed using CLSM (Zeiss LSM 880).
Besides, intracellular drug delivery by siRNA-Cy5@eS-BNVs was investigated by flow cytometry (NovoCyte, Agilent). Briefly, A549ACE2+ cancer cells (1 × 106 cells per well) were seeded in 6-well dishes and cultured in DMEM for 24 h and then exposed to PBS, free siRNA-Cy5, siRNA-Cy5/PEI, siRNA-Cy5@No-BNVs, siRNA-Cy5@eSWT-BNVs, siRNA-Cy5@eSWT-BNVs+Thr, siRNA-Cy5@eS-BNVs and siRNA-Cy5@eS-BNVs+Thr for 120 min. Subsequently, A549ACE2+ cancer cells were washed with PBS and detached for flow cytometry analysis (NovoCyte, Agilent). Data analysis and gating strategy were performed by NovoExpress software and FlowJo_V10. The representative gating strategy for the flow cytometry analysis is shown in Supplementary Fig. 22.
The cellular uptake of siRNA-Cy5@eS-BNVs by HUVEC and HEK293T normal cells was assessed by flow cytometry. HUVECs and HEK293T cells (1 × 106 cells per well) were seeded in 6-well plates and treated with either PBS, siRNA-Cy5@eS-BNVs or siRNA-Cy5@eS-BNVs + Thr (150 nM, based on siRNA) for 2 h at 37 °C. After treatment, cells were washed thrice with PBS and detached for flow cytometry analysis (NovoCyte, Agilent).
To follow the eS-BNVs mediated drugs intracellular tracing, A549ACE2+ cancer cells and A549ACE2+-EGFP cancer cells were imaged with CLSM (Zeiss LSM 880). Briefly, 150 nM siRNA-Cy5@eS-BNVs and siRNA-Cy5@eS-BNVs + Thr were incubated with A549ACE2+ cancer cells at 37 °C for 2 h. Then, the cells were incubated with LysoTracker Green DND-26 (Cat no: L7526, Invitrogen) and Hoechst 33342 (Cat no: C1025, Beyotime), successively. In a separate experiment, the indocyanine green (ICG, Cat no: I107931, aladdin) dye was encapsulated in eS-BNVs to give ICG@eS-BNVs. ICG@eS-BNVs and ICG@eS-BNVs + Thr were incubated with A549ACE2+-EGFP cancer cells. Then the cells were analysed with a CLSM (Zeiss LSM 880).
In vitro gene silencing in A549ACE2+-Luc cancer cells by siLuc@eS-BNVs
To detect the ability of eS-BNVs delivered siRNA to interfere with gene expression in cancer cells, we measured the efficiency of Luc expression in A549ACE2+-Luc cancer cells treated by siLuc@eS-BNVs. The A549ACE2+-Luc cancer cells (1 × 104 cells per well) were seeded in 48-well plates for 24 h. Then, cells were exposed to PBS, free siLuc, siLuc/PEI, siCtrl@eS-BNVs+Thr, siLuc@No-BNVs, siLuc@eSWT-BNVs, siLuc@eS-BNVs and siLuc@eS-BNVs+Thr (150 nM, based on siRNA). Membrane fusion was induced by adding the Thr enzyme (1 μ ml-1) to the system. After 48 h incubation at 37 °C, cells were washed with PBS, and D-Luciferin potassium (150 μg ml-1, Cat no: 115144-35-9, Meilunbio) was added. Then, A549ACE2+-Luc cancer cells were characterized by IVIS spectrum (IVIS Lumina, PerkinElmer) and quantitative analysis of the fluorescence signal via Living Image 4.5 software. In addition, the Luc silencing effect was also investigated by a multimode microplate reader. The A549ACE2+-Luc cancer cells (~ 3500 per well) were seeded in the 96-well plate and cultured for 24 h. Then cells were exposed to different nanoparticles as mentioned above. Then, the cells were incubated with 100 μl Luc detection reagent (Cat no: RG046S, Beyotime) for 5 min. Subsequently, cells were observed by the microplate reader (Infinite E Plex, Tecan). Furthermore, the mRNA level of Luc in A549ACE2+-Luc cancer cells was checked by the RT-qPCR. Total RNAs were extracted and purified using an RNA extraction kit (Cat no: RC112-01, Vazyme) according to the manufacturer’s instructions. Then, the total RNAs were reversely transcribed into cDNA (Cat no: R333-01, Vazyme) and tested its concentration (K5500 Plus, KRIRO). For RT-qPCR, 10 μl SYBR Green RT-qPCR mix (Cat no: RK21205, ABclonal), 1 μl forward primer (1 μM, 5′-ATTACACCCGAGGGGGATGA-3′), 1 μl reverse primer (1 μM, 5′-TCTCACACACAGTTCGCCTC-3′), 2 μl template cDNA and 6 μl ddH2O were mixed evenly in Eppendorf tubes and put into 96-well plate using Archimed (Archimed, ROCGENE). Parameters were set and data were collected through Archimed Analyzer V1.0 software. The 20 μl PCR reaction was initial denaturation 95 °C 3 min, 40 cycles of denaturation at 95 °C for 5 s, annealing at 60 °C for 30 s and extension at 72 °C for 60 s. Luc mRNA expression level was normalized to an endogenous control glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (forward primer: 5′-GACCCCTTCATTGACCTCAAC-3′; reverse primer: 5′-CTTCTCCATGGTGGTGAAGA-3′). All data was processed through the 2-ΔΔCt method.
The long time RNAi effects of siRNA@eS-BNVs were measured. Briefly, A549ACE2+-Luc cancer cells (~ 1000 cells per well) were seeded in a 96-well plate to treat with PBS and siLuc@eS-BNVs + Thr. The bioluminescence of A549ACE2+-Luc cancer cells was monitored daily for seven days by using a microplate reader (Infinite E Plex, Tecan).
In vivo gene silencing in A549ACE2+-Luc tumors by siLuc@eS-BNVs
The s.c. inoculation A549ACE2+-Luc tumor-bearing mice were randomly divided into 8 groups (n = 4) and injected with PBS, free siLuc, siLuc/PEI, siCtrl@eS-BNVs+Thr, siLuc@No-BNVs, siLuc@eSWT-BNVs, siLuc@eS-BNVs, and siLuc@eS-BNVs+Thr (1 mg kg−1 per mouse, total volume of 200 μl) by tail i.v. injection to evaluate the RNAi effect. The siCtrl@eS-BNVs+Thr and siLuc@eS-BNVs+Thr groups were tail i.v. injected Thr enzyme (1.5 U per mouse, total volume of 200 μl) to initiate membrane fusion at 12 h after drug administration, this combination is considered as a treatment. And try to ensure that each group has a similar average bioluminescence intensity at the beginning. Mice were injected intraperitoneally with 200 μl D-Luciferin potassium salt (15 mg ml−1, Cat no: 115144-35-9, Meilunbio) and anesthetized in a chamber with 1-4% isoflurane. Subsequently, the mice were imaged at 0 h, 24 h, 36 h and 48 h post-treatment by an IVIS spectrum (Lumina, PerkinElmer). Finally, the A549ACE2+-Luc tumors were extracted, washed with PBS, and the Luc gene silencing efficiency was determined by RT-qPCR. Details about the sequence of siRNA were presented in Supplementary Table 2.
Safety evaluation of siRNA@eS-BNVs in mice
The cytotoxicity of eS-BNVs was evaluated on C57BL/6 mice (n = 3, 6 weeks) in vivo. Briefly, the mice were tail i.v. injected with PBS, free siRNA, and siRNA@eS-BNVs (1 mg kg−1, based on siRNA) and sacrificed at 72 h. To assess the organ damage, the heart, liver, spleen, lung and kidney were excised for H&E staining. Briefly, tissues were fixed in 4% PFA for more than 24 h. Then, the tissues were were dehydrated with 75, 85, 90 and 95% alcohol for 4, 2, 2 and 1 h, anhydrous ethanol for 30 min, alcohol benzene for 5–10 min, and xylene I for 5–10 min. Then, the tissues were embedded with wax. The fixed tissues were cut into 4 μm thickness slices, and put into the environmental friendly dewaxing transparent liquid I for 20 min, environmental friendly dewaxing transparent liquid II for 20 min, anhydrous ethanol I for 5 min, anhydrous ethanol II for 5 min, 75% ethyl alcohol for 5 min, and then rinsed with PBS. The sections were stained with hematoxylin (Cat no: G1076, Servicebio) for 3–5 min and eosin (Cat no: G1076, Servicebio) for 15 s for histopathological assay, and the slices were photographed by an automatic multispectral imaging system (Pannoramic MIDI, 3DHISTECH). Data were collected using CaseViewer software. The blood was collected and the whole blood specimens were left at room temperature for 2 h and then centrifuged at 4 °C for 20 min at 2000 × g, and the supernatant was removed for ALT, AST, BUN, CREA and LDH assay.
To determine the cytotoxicity of PEI to cells, ~ 3500 HEK293T cells were seeded into a 96-well plate and cultured overnight. Cells were treated with PEI and PEI-loaded in eS-BNVs. A cell counting kit-8 (CCK8) assay (Cat no: CB101, Oriscience) was used, and cell toxicity was determined via a microplate reader (Infinite E Plex, Tecan).
In vivo biodistribution and pharmacokinetic study of siRNA-Cy5@eS-BNVs
To study the biodistribution of eS-BNVs in vivo, free siRNA-Cy5, siLuc/PEI, siRNA-Cy5@No-BNVs, siRNA-Cy5@eSWT-BNVs, siRNA-Cy5@eS-BNVs and siRNA-Cy5@eS-BNVs+Thr were tail i.v. injection in the A549ACE2+ tumor-bearing mice. Briefly, A549ACE2+ cancer cells (2 × 106 cells per mouse, 100 μl, suspended in DMEM) were s.c. inoculated into the left lower abdomen of BALB/c nude mice to prepare s.c. inoculation A549ACE2+ tumor-bearing mice models. When the tumor volume reached 60 ~ 80 mm3, the mice were randomly divided into different groups. Free siRNA-Cy5 or siRNA-Cy5@eS-BNVs (1 mg kg−1 siRNA-Cy5 per mouse, 200 μl) were tail i.v. injected into A549ACE2+ tumor-bearing mice, and then measured using IVIS spectrum at 2, 12 and 24 h. Mice were sacrificed at 24 h, and the A549ACE2+ tumors and major organs, including heart, liver, spleen, lung, kidney, and A549ACE2+ tumors were reseted and imaged using IVIS spectrum (IVIS Lumina, PerkinElmer) and quantitative analysis of the fluorescence signal via Living Image 4.5 software. For the study of pharmacokinetics in vivo, blood was collected and blood was prepared and imaged using the IVIS spectrum. To investigate the distribution of siRNA-Cy5@eS-BNVs and other nanoparticles in ex vivo tumors, the mice were inoculated with A549ACE2+-EGFP tumors. After 24 h, the mice were euthanized. A549ACE2+-EGFP tumors were fixed with 4% PFA and prepared as paraffin sections. The penetration of siRNA-Cy5 was observed by labeling the blood vessels of with CD31 antibody (Cat no: GB113151, Servicebio). The in vivo tumor penetration behavior was also observed in s.c. inoculation A549ACE2+-EGFP tumor-bearing mice injected with the treatment of free siRNA-Cy5 and siRNA-Cy5@eS-BNVs+Thr (1 mg kg−1 siRNA-Cy5 per mouse, 200 μl) using IntraVital Microscope (IVM-CM, IVIM TECHNOLOGY). Data were collected using IVM Studio software. The mice were anesthetized with 5% chloral hydrate and fixed on the thermostatic bench (37 °C) to scan the distribution of siRNA-Cy5 in vivo at 2 h.
Distribution of ThrAlexa647 in tumor-bearing mice
To investigate whether Thr enzyme can reach the tumors, we examined its biodistribution in A549ACE2+ tumor-bearing mice. First, Thr enzyme was fluorescently labeled using Alexa Fluor 647 NHS Ester. Briefly, Thr (2 mg ml-1) was incubated with Alexa Fluor 647 NHS Ester (10 μg ml-1, Cat no: A37573, Thermo Fisher) in sodium bicarbonate buffer (100 mM, pH 8.3) at room temperature overnight. The free dye was removed via ultracentrifugation, and the purified Alexa Fluor 647-labeled Thr (ThrAlexa647) enzyme was collected for subsequent experiments. For biodistribution analysis, A549ACE2+ tumor-bearing BALB/c nude mice were i.v. injected with ThrAlexa647 enzyme (2 μg per mouse) via the tail vein. The A549ACE2+ tumors were harvested at 1 h and 2 h post-injection for ex vivo fluorescence imaging using an IVIS spectrum (IVIS Lumina, PerkinElmer), and the fluorescence intensity was quantified via the Living Image 4.5 software.
siEGFR@eS-BNVs for inhibiting the A549ACE2+ tumors
To evaluate the anti-tumor effect of siEGFR@eS-BNVs in vivo, six-week-old female BALB/c nude mice were received the treatments of PBS, free siEGFR, siCtrl@eS-BNVs + Thr, siEGFR@No-BNVs,siEGFR@eS-BNVs and siEGFR@eS-BNVs + Thr. The s.c. inoculation A549ACE2+ tumor-bearing mice were randomly grouped (n = 6 for each group) and tail i.v. administered with PBS, free siEGFR, siCtrl@eS-BNVs, siEGFR@No-BNVs, siEGFR@eS-BNVs and the second siEGFR@eS-BNVs. The siCtrl@eS-BNVs and the second siEGFR@eS-BNVs (i.e., siEGFR@eS-BNVs + Thr) groups were tail i.v. injected with Thr enzyme (1.5 U per mouse, total volume of 200 μl) to initiate membrane fusion at 12 h after drug administration. The mice received five doses of treatments by PBS or siRNA (1 mg kg−1 per mouse, total volume of 200 μl) of different formulations every 2 days by tail i.v. injection. Tumor volume, body weight and survival rates were recorded. On day 20 of the experiment, the mice were photographed to record the tumor volume. Details about the sequence of siRNA wasere presented in Supplementary Table 2.
To characterize the gene silencing effects, tumors were resected on day 16 after three times administration of siEGFR@eS-BNVs (n = 6). After the third treatment, the mice were euthanized, and the tumors were characterized by IF staining (BZ-X800E, KEYENCE), western blot, and RT-qPCR to verify the EGFR protein and gene expression. The antibodies used in the experiment were organized in Supplementary Table 1.
siEGFR@eS-BNVs for inhibiting the lung metastasis
To further investigate the anti-lung metastasis effect of siEGFR@eS-BNVs in vivo, we carried out tumor therapy experiments in mice with lung metastases. The lung metastasis model was prepared by tail i.v. injection of A549ACE2+-Luc cancer cells (1 × 106 cells per mouse, 100 μl, suspended in DMEM). The mice (n = 5) were randomly grouped and tail i.v. injection the PBS, free siEGFR, siCtrl@eS-BNVs, siEGFR@No-BNVs, siEGFR@eS-BNVs and the second siEGFR@eS-BNVs on days 10, 12, 14, 16 and 18. The siCtrl@eS-BNVs and the second siEGFR@eS-BNVs groups were tail i.v. injected Thr enzyme (1.5 U per mouse, total volume of 200 μl) to initiate membrane fusion at 12 h after drug administration. The mice received five doses of treatments by PBS or siRNA (1 mg kg−1 per mouse, total volume of 200 μl) of different formulations every 2 days by tail i.v. injection. The lung metastases were monitored at days 10, 23 and 35, and the bioluminescence intensity was quantified using the IVIS spectrum (Lumina, PerkinElmer). Body weight and survival rates were monitored. In order to better analyze the differences between the various treatment groups, we re-performed the mouse anti-lung metastasis experiment (n = 6) in the subsequent EGFR characterization analysis. The mice lung metastasis model was reconstructed, and the lungs were excised to be photographed, H&E staining, IHC staining, western blot and RT-qPCR experiments. The method of PCR was specifically referred to above, here, the sequence of EGFR primer was cited as forward 5’-GGACGACGTGGTGGATGCCG-3’ and reverse 5’-GGCGCCTGTGGGGTCTGAGC-3’.
Software and statistical analysis
Softwares used for data collection include eBLOT14, ZetasiZer, Archimed Analyzer V1.0, ZEN 3.6, NovoExpress, Cary WinFLR, i-control 2.0 and Living Image 4.5. Statistical data analysis software includes GraphPad Prism 9.0.0, Microsoft Office 2019, ImageJ 1.53a, and FlowJo_V10.
Unless otherwise specified, experiment data were presented as mean values ± s.d. A two-tailed Student’s t test was performed when comparing the two groups. A one-way analysis of variance (ANOVA) test was used for comparing multiple groups. p < 0.05 of data difference was considered significant. A p-value of 0.05 or higher was considered to indicate non-significance at the 95% confidence level.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The protein structure simulation data generated in this study have been deposited in the ModelArchive under accession numbers ma-9jos9 [https://www.modelarchive.org/doi/10.5452/ma-9jos9]. Other structures used in this study were obtained from the Protein Data Bank (PDB) with accession codes 6VXX [https://www.uniprot.org/uniprotkb/P0DTC2/entry]. Protein sequences used in this study were retrieved from Uniprot with accession ID: P0DTC2 (name spike glycoprotein). Source data is available for Figs. 1d, f, h, 2d, e, f–h, j–l, 3a, d, f, h, j, k, 4b–d, f, g, 5c, 6b–f; 7c–f, i, k and Supplementary Figs. 2b, 3, 5d, 5e, 7b, d, 10, 11, 12a, 13, 14, 15, 16b, 23c, 24b–d; 26b, c, 29, 30, 31, 34 in the associated source data file. All other data supporting the results of this paper are available in the article, supplementary information or source data file. Source data are provided in this paper.
References
Kim, H. J., Kim, A., Miyata, K. & Kataoka, K. Recent progress in development of siRNA delivery vehicles for cancer therapy. Adv. Drug Deliv. Rev. 104, 61–77 (2016).
Fire, A. et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806–811 (1998).
Whitehead, K. A., Langer, R. & Anderson, D. G. Knocking down barriers: advances in siRNA delivery. Nat. Rev. Drug Discov. 8, 129–138 (2009).
Setten, R. L., Rossi, J. J. & Han, S. P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 18, 421–446 (2019).
Rand, T. A., Ginalski, K., Grishin, N. V. & Wang, X. Biochemical identification of Argonaute 2 as the sole protein required for RNA-induced silencing complex activity. Proc. Natl. Acad. Sci. USA 101, 14385–14389 (2004).
Banskota, S. et al. Engineered virus-like particles for efficient in vivo delivery of therapeutic proteins. Cell 185, 250–265 (2022).
Kreitz, J. et al. Programmable protein delivery with a bacterial contractile injection system. Nature 616, 357–364 (2023).
Davis, M. E. et al. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 464, 1067–1070 (2010).
Dammes, N. et al. Conformation-sensitive targeting of lipid nanoparticles for RNA therapeutics. Nat. Nanotechnol. 16, 1030–1038 (2021).
Dowdy, S. F. Overcoming cellular barriers for RNA therapeutics. Nat. Biotechnol. 35, 222–229 (2017).
Wang, J. et al. Imaging-guided delivery of RNAi for anticancer treatment. Adv. Drug Deliv. Rev. 104, 44–60 (2016).
Watanabe, S. et al. In vivo rendezvous of small nucleic acid drugs with charge-matched block catiomers to target cancers. Nat. Commun. 10, 1894 (2019).
Hoogenboezem, E. N. et al. Structural optimization of siRNA conjugates for albumin binding achieves effective MCL1-directed cancer therapy. Nat. Commun. 15, 1581 (2024).
Liu, M., Hu, S., Yan, N., Popowski, K. D. & Cheng, K. Inhalable extracellular vesicle delivery of IL-12 mRNA to treat lung cancer and promote systemic immunity. Nat. Nanotechnol. 19, 565–575 (2024).
Qiu, C. et al. Regulating intracellular fate of siRNA by endoplasmic reticulum membrane-decorated hybrid nanoplexes. Nat. Commun. 10, 2702 (2019).
Fang, R. H., Gao, W. & Zhang, L. Targeting drugs to tumours using cell membrane-coated nanoparticles. Nat. Rev. Clin. Oncol. 20, 33–48 (2023).
Dahlman, J. E. et al. In vivo endothelial siRNA delivery using polymeric nanoparticles with low molecular weight. Nat. Nanotechnol. l9, 648–655 (2014).
Yin, Q. et al. Quantitative imaging of intracellular nanoparticle exposure enables prediction of nanotherapeutic efficacy. Nat. Commun. 12, 2385 (2021).
Gilleron, J. et al. Image-based analysis of lipid nanoparticle-mediated siRNA delivery, intracellular trafficking and endosomal escape. Nat. Biotechnol. 31, 638–646 (2013).
Wang, L. et al. Bioinspired engineering of fusogen and targeting moiety equipped nanovesicles. Nat. Commun. 14, 3366 (2023).
Yang, J. et al. Drug delivery via cell membrane fusion using lipopeptide modified liposomes. Acs. Cent. Sci. 2, 621–630 (2016).
Kim, B. et al. Immunogene therapy with fusogenic nanoparticles modulates macrophage response to Staphylococcus aureus. Nat. Commun. 9, 1969 (2018).
Lin, M. et al. A biomimetic approach for spatially controlled cell membrane engineering using fusogenic spherical nucleic acid. Angew. Chem. Int. Ed. 61, e202111647 (2022).
Mi, P., Cabral, H. & Kataoka, K. Ligand-installed nanocarriers toward precision therapy. Adv. Mater. 32, e1902604 (2020).
Liu, J., Cabral, H. & Mi, P. Nanocarriers address intracellular barriers for efficient drug delivery, overcoming drug resistance, subcellular targeting and controlled release. Adv. Drug Deliv. Rev. 207, 115239 (2024).
Hoffmann, M., Kleine-Weber, H. & Pöhlmann, S. A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells. Mol. Cell 78, 779–784 (2020).
Hoffmann, M. et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280 (2020).
Bai, L. et al. Coinfection with influenza A virus enhances SARS-CoV-2 infectivity. Cell Res. 31, 395–403 (2021).
Jackson, C. B., Farzan, M., Chen, B. & Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 23, 3–20 (2022).
Wu, M. et al. A suitable membrane distance regulated by the RBD_ACE2 interaction is critical for SARS-CoV-2 spike-mediated viral invasion. Adv. Sci. 10, e2301478 (2023).
Yu, S. et al. SARS-CoV-2 spike engagement of ACE2 primes S2′ site cleavage and fusion initiation. Proc. Natl. Acad. Sci. USA 119, e2111199119 (2021).
Lee, B. et al. Furin cleavage of SARS-CoV-2 Spike promotes but is not essential for infection and cell-cell fusion. PLoS Pathog. 17, e1009246 (2021).
Low, J. S. et al. ACE2-binding exposes the SARS-CoV-2 fusion peptide to broadly neutralizing coronavirus antibodies. Science 377, 735–742 (2022).
Diao, J. et al. ATG14 promotes membrane tethering and fusion of autophagosomes to endolysosomes. Nature 520, 563–566 (2015).
Zhuo, Y. et al. Direct cytosolic delivery of siRNA via cell membrane fusion using cholesterol-enriched exosomes. Nat. Nanotechnol. 19, 1858–1868 (2024).
Zhang, H., Liu, J., Chen, Q. & Mi, P. Ligand-installed anti-VEGF genomic nanocarriers for effective gene therapy of primary and metastatic tumors. J. Control Release 320, 314–327 (2020).
Voigt, M. et al. Functional dissection of the epidermal growth factor receptor epitopes targeted by panitumumab and cetuximab. Neoplasia 14, 1023–1031 (2012).
Yu, J. -j et al. TRIB3-EGFR interaction promotes lung cancer progression and defines a therapeutic target. Nat. Commun. 11, 3660 (2020).
Li, S. et al. A DNA nanorobot functions as a cancer therapeutic in response to a molecular trigger in vivo. Nat. Biotechnol. 36, 258–264 (2018).
Li, S. et al. Combination of tumour-infarction therapy and chemotherapy via the co-delivery of doxorubicin and thrombin encapsulated in tumour-targeted nanoparticles. Nat. Biomed. Eng. 4, 732–742 (2020).
Hu, C.-M. J. et al. Erythrocyte membrane-camouflaged polymeric nanoparticles as a biomimetic delivery platform. Proc. Natl. Acad. Sci. USA 108, 10980–10985 (2011).
Kim, S. S., Garg, H., Joshi, A. & Manjunath, N. Strategies for targeted nonviral delivery of siRNAs in vivo. Trends Mol. Med. 15, 491–500 (2009).
Pitchaimani, A., Nguyen, T. D. T. & Aryal, S. Natural killer cell membrane infused biomimetic liposomes for targeted tumor therapy. Biomaterials 160, 124–137 (2018).
Paunovska, K., Loughrey, D. & Dahlman, J. E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 23, 265–280 (2022).
Brown, D. W. et al. Safe and effective in vivo delivery of DNA and RNA using proteolipid vehicles. Cell 187, 5357–5375 e5324 (2024).
Liew, F. et al. SARS-CoV-2-specific nasal IgA wanes 9 months after hospitalisation with COVID-19 and is not induced by subsequent vaccination. EBioMedicine 87, 104402 (2023).
Fang, R. H. et al. Cancer cell membrane-coated nanoparticles for anticancer vaccination and drug delivery. Nano. Lett. 14, 2181–2188 (2014).
Diao, J. et al. A single vesicle-vesicle fusion assay for in vitro studies of SNAREs and accessory proteins. Nat. Protoc. 7, 921–934 (2012).
Du, Y., Lai, Y., Liu, J. Y. & Diao, J. Epigenetic quantification of DNA 5-hydroxymethylcytosine using DNA hybridization-based single-molecule immunofluorescent imaging. Small Methods 5, e2100061 (2021).
Joo, C. et al. Real-time observation of recA filament dynamics with single monomer resolution. Cell 126, 515–527 (2006).
Kyoung, M. et al. In vitro system capable of differentiating fast Ca2+-triggered content mixing from lipid exchange for mechanistic studies of neurotransmitter release. Proc. Natl. Acad. Sci. USA 108, E304–E313 (2011).
Acknowledgements
This work was partially supported by the National Natural Science Foundation of China (No. 32371446 to P.M., No. 32170686 to Y.L., No, 32401162 to J.L.), the Ministry of Science and Technology of China (No. 2022YFC2303700 to Y.L.), the Sichuan Science and Technology Program (No. 2023NSFSC1923 to P.M.), the Natural Science Foundation of Sichuan Province (No. 00402053A2ABG to Y.L.), the China Postdoctoral Science Foundation (No. 2024M762257 to J.L.), the Postdoctoral Fellowship Program of CPSF (No. GZC20241173 to J.L.), the 1.3.5 project (No. ZYYC23019 to P.M., No. ZYYC21007, Z2021JC007, ZYYC24003 to Y.L., No. 2024HXBH085 to J.L.) for disciplines of excellence and the Postdoctor Research Fund of West China Hospital, Sichuan University, and Transformation Foundation of Tianfu Jincheng Laboratory (No. 2025ZH020 to Y.L.).
Author information
Authors and Affiliations
Contributions
P.M. and Y.L. proposed the concept. P.M., Y.L. and L.C. conceived the experimental design. Y.C. and J.L. contributed to animal-related experiments. W.L. and M.W contributed to vesicle-related experiments. P.M., Y.L. and J.L. received funding for this study. P.M., Y.L. and L.C. wrote the manuscript. X.W. provided critical insights. P.M. and Y.L. provided critical insights, supervised and directed this project. All authors commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Shi Yu, and the other anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cui, L., Cui, Y., Liu, J. et al. Bioengineered nanovesicles for efficient siRNA delivery through ligand-receptor-mediated and enzyme-controlled membrane fusion. Nat Commun 16, 6174 (2025). https://doi.org/10.1038/s41467-025-61230-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-61230-1
This article is cited by
-
Biomimetic PD-1-nanovesicles inducing tri-modal sonodynamic-chemo-immunotherapy for breast cancer therapy and lung metastasis inhibition
Journal of Nanobiotechnology (2026)
