
Abstract
Nuclear clearance and cytoplasmic aggregation of TDP-43, initially identified in ALS-FTD, are hallmark pathological features observed across a spectrum of neurodegenerative diseases. We previously found that TDP-43 loss-of-function leads to transcriptome-wide inclusion of deleterious cryptic exons, a signature detected in presymptomatic biofluids and postmortem ALS-FTD brain tissue, but the upstream mechanisms that lead to TDP-43 dysregulation remain unclear. Here, we developed a web-based resource (SnapMine) to determine the levels of TDP-43 cryptic exon inclusion across hundreds of thousands of publicly available RNA sequencing datasets. We established cryptic exon inclusion levels across a variety of human cells and tissues to provide ground truth references for future studies on TDP-43 dysregulation. We then explored studies that were entirely unrelated to TDP-43 or neurodegeneration and found that ciclopirox olamine (CPX), an FDA-approved antifungal, can trigger the inclusion of TDP-43-associated cryptic exons in a variety of mouse and human primary cells. CPX induction of cryptic exons arises from heavy metal toxicity and oxidative stress, suggesting that similar vulnerabilities could play a role in neurodegeneration. Our work demonstrates how diverse datasets can be linked through common biological features and underscores how public archives of sequencing data remain a vastly underutilized resource with tremendous potential for uncovering novel insights into complex biological mechanisms and diseases.
Similar content being viewed by others

Introduction
RNA splicing is an essential process in eukaryotic cells that maintains the integrity of the transcriptome and proteome. The spliceosome plays a pivotal role in this process in which a single gene gives rise to multiple distinct mRNAs through precise joining of different combinations of exons during transcription. The resulting protein isoforms often exhibit tissue- or developmental stage-specific expression patterns, highlighting the role of alternative splicing in regulating gene expression and protein structure1,2,3,4,5,6,7. Defects in RNA splicing can lead to significant alterations in the transcriptome and proteome which contribute to cellular dysfunction and disease states8,9,10. Certain RNA binding proteins (RBPs) help maintain transcriptome fidelity by acting as intronic splicing repressors. These splicing repressor RBPs recognize and bind to specific motifs located near aberrant splicing sites for regions such as cryptic exons, thereby repressing their incorporation into mature mRNA during the splicing process11,12,13,14,15,16,17,18,19,20.
TDP-43 performs its role in cryptic exon repression by preferentially binding to uracil-guanine repeats ([UG]n) in RNA11,21,22. Inclusion of cryptic exons often leads to premature termination codons and nonsense-mediated decay23, while cryptic exons in the 5’ or 3’ untranslated regions can impact processes such as mRNA stability and translation efficiency24. Although TDP-43 may have multiple functions, recent studies have converged on the importance of cryptic exon repression in neurodegenerative disorders, particularly amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Over 97% of ALS and 45% of FTD cases exhibit TDP-43 pathology23,25,26,27, which is characterized by nuclear clearance and cytoplasmic aggregation of TDP-43 protein. Recent studies have also associated TDP-43 pathology with other neurodegenerative disorders, such as Alzheimer’s disease and Huntington’s disease28,29,30,31,32,33,34,35,36,37,38,39,40,41. While significant research has focused on the cytoplasmic aggregation of TDP-43, the nuclear clearance of the protein has emerged as a critical aspect of its pathophysiological role. Nuclear loss of TDP-43 leads to the inclusion of nonconserved cryptic exons11,17,22,23,42,43,44, some of which are unique to specific cell types42. These cryptic exons can be identified in both experimental models and patient samples using RNA-sequencing (RNA-Seq) technologies.
While most cryptic exons are out-of-frame, approximately 3% of TDP-43 cryptic exons introduce in-frame insertions into mRNA coding sequences42. For example, the translated human cryptic exon in HDGFL2 produces a neoepitope that can be detected in patients presymptomatically and can potentially be used as a fluid biomarker for TDP-43 loss-of-function45,46,47. Other cryptic exon peptides, such as those in MYO18A and G3BP1, have also been detected in post-mortem samples using mass spectrometry, indicating translation of these aberrant mRNAs47. The feasibility of using cryptic exon-encoded peptides as biomarkers relies on their exclusivity to affected tissues. However, establishing baseline levels of cryptic exon inclusion across human tissues can be challenging to accomplish computationally without pre-processed resources tailored for this purpose. For instance, although the Genotype-Tissue Expression (GTEx) project provides a wealth of RNA-Seq data across multiple human tissues, the GTEx web portal lacks a user-friendly way to perform custom splicing queries on a sample-by-sample basis. This limitation makes it difficult for users to establish comprehensive baseline splicing profiles and assess the specificity of cryptic exon inclusion levels.
Prior work has compiled and processed large collections of public RNA-Seq archives to provide resources to investigate splicing patterns in samples, such as Snaptron48, Recount349,50, and ASCOT51. The first two offer databases of pre-processed RNA-Seq data that can be mined to extract information on cryptic exon inclusion levels. However, the ability to effectively utilize these tools requires a certain level of bioinformatics expertise, making them less accessible to many biologists. We previously developed ASCOT to perform comprehensive, annotation-free identification of binary splicing events using a computationally intensive junction-walking method. Due to its complexity, ASCOT’s analyses are limited to precomputed compilations (GTEx, ENCODE, and two manually curated mouse datasets) and cannot directly query new events or rapidly extend analyses to large-scale archives like TCGA or SRA. To address this challenge, we have developed a user-friendly, web-based resource (SnapMine) that streamlines the process of querying Snaptron and extracting relevant splicing information. SnapMine employs a complementary approach to ASCOT, where users with a predefined splice junction query can instantly access splicing information across hundreds of thousands of diverse public RNA-Seq datasets. Our web-based tool enables biologists to rapidly profile splicing and gene expression data across the public archive without the need for bioinformatics experience. SnapMine aims to facilitate exploring the role of alternative splicing in health and disease by providing a more accessible interface to query under-utilized RNA-Seq resources.
SnapMine is available at https://snapmine.idies.jhu.edu/ and can be used to mine human and mouse RNA-Seq datasets from The Cancer Genome Atlas (TCGA), GTEx, the Sequence Read Archive (SRA), and shRNA-Seq in HepG2 and K562 from the ENCODE project52,53,54,55,56. To demonstrate SnapMine’s utility, we employed TDP-43 cryptic exons as a representative splicing phenotype and mined these RNA-Seq archives for their inclusion patterns. By leveraging SnapMine to comprehensively profile cryptic exon inclusion across hundreds of thousands of datasets, we established essential baseline levels in diverse non-disease tissues and cell types. This reference will support the development of robust biomarkers of TDP-43 loss-of-function and empower future studies on the role of cryptic exons in disease.
SnapMine makes it simple and intuitive to uncover hidden connections across disparate datasets. As a proof-of-principle, we harnessed SnapMine to explore the upstream processes that can regulate TDP-43 by identifying cryptic exons in studies that had no prior connection to TDP-43 or neurodegeneration. By detecting splicing patterns through this integrated and unbiased approach, SnapMine identified datasets that could shed light on potential disease pathways that contribute to the initiation or progression of TDP-43 pathology. This represents a shift from traditional methods that focus on mining pre-selected datasets already known for their relevance and significance. Instead, searches can be performed across diverse datasets to find hidden links that no other approach could reveal. SnapMine provides a new, effective way to understand complex biological processes by harnessing the underutilized potential of large-scale public data archives.
Results
Mining the Snaptron database for alternative splicing events of interest
The Snaptron database48 includes compiled exon-exon splice junction information across samples from four human RNA-Seq sample databases: SRA52, GTEx53, TCGA54, and ENCODE56. Mouse splice junction information has also been extracted from the SRA. We queried the previously described known TDP-43-associated cryptic exon targets identified from human and mouse TDP-43 knockdown datasets11,42,57,58,59,60. Junction counts were extracted from the Snaptron database, and the percent spliced-in (PSI) ratio was calculated using counts from each sample (Fig. 1a). For each sample, the PSI value ranged from 0–100% depending on the rate of cryptic junction inclusion as compared to the canonical junction. Metadata regarding the samples, such as SRA accession number or tissue type, was also extracted to give context to alternative splicing events.

a Schematic of SnapMine algorithm. SnapMine mines Snaptron for exon-exon junction counts of samples in the Sequencing Read Archive (SRA), the Cancer Genome Atlas (TCGA), and the Genotype-Tissue Expression (GTEx) project. The junction counts are extracted, as represented by the blue and red columns for the exclusion and inclusion junctions, respectively, in each sample. The percent spliced-in (PSI) value is then calculated for each inclusion junction of interest. b SnapMine interface with example usage to find results associated with one junction of the HDGFL2 cryptic exon in the Sequencing Read Archive (SRA). The graph tab is minimized. c SRA samples including the HDGFL2 cryptic exon (CE) (left) subset to those with an average inclusion rate higher than 5% (right). Most samples with avgPSI > 5% for the HDGFL2 CE are from ALS-associated or TDP-43-knockdown (KD)/mutation (mut.) studies. Data are presented as box plots with median and interquartile range, along with whiskers from minimum to maximum values. Source data are provided as a Source Data file.
R scripts were initially used to query Snaptron singularly and in bulk for multiple cryptic exons. These scripts are available on GitHub and an application to query the databases for up to five junctions simultaneously has been made publicly available at https://snapmine.idies.jhu.edu/ (Fig. 1b). Users of the application can input inclusion and exclusion junction coordinates to calculate sample PSI values in one of the databases. The output of the query is a table detailing the PSI values of the inclusion junction for each sample with select metadata. If multiple junctions are queried, the average of the PSI values is also calculated. The table can be downloaded by the user and graphs of the data are generated and displayed to provide a quick visual regarding junction. Additionally, the application can tabulate gene expression counts within each sample using the pre-analyzed information in Snaptron. If desired, gene counts can be normalized to a control of choice. The example included in the application refers to EDF1, a gene with relatively constant levels of RNA expression levels across human tissues51. Some common cryptic exon coordinates and the associated inclusion and exclusion junctions are available in Table S1.
HDGFL2 as a biomarker for TDP-43 loss of function
We first tested the capabilities of querying Snaptron by investigating the inclusion of the HDGFL2 cryptic exon within the 316,000 human RNA-Seq samples in the SRA compilation. We calculated the PSI for the junctions on the 5’ and 3’ sides of the cryptic exon compared to the canonical junction that skips the cryptic exon. In the 29,200 human samples in which the HDGFL2 junction could be detected, only 55 samples (0.19%) had an average PSI for both cryptic junctions greater than 5% (Fig. 1c). Of the 55 samples, 18 were directly associated with TDP-43 depletion. Another was from a C9-ALS case, although the TDP-43 pathology level was not available. Finally, nine other samples were associated with cancer samples, while the remaining 26 samples were not as easily categorized. Of uncategorized samples, conditions included viral and immune manipulations, along with other neurodegenerative diseases. Overall, we found that the HDGFL2 cryptic exon was included almost exclusively in cases of TDP-43 manipulation, dysregulation, or depletion and thus would be a relatively specific biomarker for cases of TDP-43 loss-of-function.
Identifying baseline cryptic exon inclusion levels for potential biomarkers
Due to the recent interest in using TDP-43-associated cryptic exons as biomarkers, we also investigated the baseline PSI values for other identified cryptic exons. Of the many identified cryptic exons, we chose to test the application using those in AGRN, ATG4B, G3BP1, HDGFL2, MYO18A, PFKP, RANBP1, STMN2, UNC13A, and UNC13B due to their biomarker and therapeutic relevance45,47,57,58,59,60,61,62. We queried these cryptic exons in the GTEx database, which includes RNA sequenced from many postmortem human organs expected to be free of disease processes63 (Fig. 2a, Fig. S1a, b). Since some donors may have had undiagnosed TDP-43 pathologies, we excluded 62 individuals with multiple cryptic exons in a single tissue (Fig. S2a–c). These individuals expressed high levels of cryptic exons across multiple tissues, with particular enrichment in the brain (Fig. S2b). For many of the cryptic exons, inclusion rates in these normal tissues are below 20%, but the cryptic exon in UNC13B is a notable outlier. This cryptic exon is included at relatively high rates in most sequenced GTEx samples.

a, b Average percent spliced-in (PSI) of cryptic exons (CEs) in AGRN, ATG4B, G3BP1, HDGFL2, MYO18A, PFKP, RANBP1, STMN2, UNC13A, and UNC13B. Data are presented as box plots with median and interquartile range, along with whiskers from minimum to maximum values. Each point represents a sample in Genotype-Tissue Expression (GTEx) project dataset (n = 19803) (a) All samples. Average PSI is not always 0 and the CE in UNC13B has particularly high inclusion rates. b Tissue-specific expression indicates CE inclusion differs by tissue. Some cryptic exons have generally low inclusion in all samples while others have tissue-specific enrichment. Points are colored in a gradient from blue (PSI = 0%) to red (PSI ≥ 20%). The following tissues are sampled: adipose (n = 28446), adrenal gland (n = 6028), bladder (n = 462), blood (n = 23056), blood vessel (n = 30756), bone marrow (n = 4488), brain (n = 64482), breast (n = 10604), cervix uteri (n = 418), colon (n = 18084), esophagus (n = 34694), fallopian tube (n = 198), heart (n = 20724), kidney (n = 2156), liver (n = 5522), lung (n = 14410), muscle (n = 19382), nerve (n = 14498), ovary (n = 4290), pancreas (n = 7920), pituitary (n = 6622), prostate (n = 5786), salivary gland (n = 3916), skin (n = 42680), small intestine (n = 4246), spleen (n = 5610) stomach (n = 8448), testis (n = 9020), thyroid (n = 15532), uterus (n = 3498), vagina (n = 3806). c Heatmap representation of percentage GTEx samples positive for each cryptic exon in each tissue. Samples are considered positive if avgPSI > 5% for the cryptic exon. ARHGAP32, PFKP, RANBP1, and UNC13B have tissue-specific enrichment. A gradient from white (% positive = 0%) to deep red (% positive ≥ 20%) is used to color the tiles. d UCSC Genome Browser visualization of the STMN2 cryptic exon in untreated neuroblastoma cell lines. Blue arrow indicates the cryptic exon. e UCSC Genome Browser visualization of HDGFL2, PFKP, and TRRAP cryptic exons in MDA-MD-231 cells after TDP-43 and/or SRSF3 knockdown. Blue arrow indicates the cryptic exon. Lanes with red-colored reads indicate a TDP-43 knockdown. Lanes with black-colored reads represent samples without TDP-43 knockdown. The PSI of HDGFL2, PFKP, and TRRAP cryptic exons appears to increase with co-knockdown of SRSF3 compared to a TDP-43 only knockdown. SRSF3 knockdown appears sufficient in these cells for cryptic exon inclusion.
Due to the slight spread in PSI values for the cryptic exons, we investigated if there was any tissue-type specificity to the cryptic exons (Fig. 2b, c, Fig. S1a). Cryptic exons in many targets, such as ACTL6B, AGRN, ATG4B, HDGFL2, TRRAP, and UNC13A were included at very low rates in samples. Elevated levels of some other cryptic exons, such as ARHGAP32, RANBP1, and UNC13B are detected at much higher levels than expected within normal samples.
This implies that TDP-43 loss-of-function may be more common in normal tissues than expected, or that other mechanisms beyond TDP-43 dysfunction can contribute to incorporation of those cryptic exons (Fig. 2c). The cryptic exon in UNC13B is likely an alternative splicing event that occurs in some normal tissues. Further work will be needed to identify conditions for the inclusion of cryptic exons prior to their usage as biomarkers in brain tissue or brain-related biofluids.
As a more comprehensive test of potential targets, we queried the inclusion of cryptic exons in GTEx for CAMK2B, EPB41L4A, GPSM2, IGLON5, KALRN, PXDN, RSF1, SLC24A3, SYN1, SYT7, and TRRAP (Fig. S1a, Supplementary Table 1). PSI values of cryptic exons are generally low across most samples, but certain cryptic exons have elevated baseline splicing inclusion levels in certain tissues. Inclusion of the cryptic exon in EPB41L4A is slightly elevated in the esophagus, PXDN in the brain, KALRN in breast tissue, and SLC24A3 in esophageal tissue. Overall, there are more samples with elevated PSI values for these TDP-43-associated cryptic exons in tissue derived from blood vessels, brain, colon, esophagus, heart, and muscle. Brain and muscle tissue show increased susceptibility to TDP-43-associated cryptic exon exclusion in GTEx tissue. Additionally, there is no significant correlation between overall cryptic exon inclusion levels and age group in the GTEx samples (Fig. S1a, b).
Querying SRA database for novel biological associations
After validating our ability to query databases through Snaptron for cryptic exon-associated junctions, we queried the SRA for multiple cryptic exons associated with TDP-43 depletion to search for new associations or contexts in which the phenotype occurs. Unlike the GTEx database, the SRA contains numerous samples from tissues and cells that are not considered normal or have been subjected to environmental and/or genetic manipulations. Additionally, the databases indexed by Snaptron contain samples from both human and mouse sources. We mined the database for cryptic exons, removed the TDP-43-related samples, and searched for new associations. Samples were considered positive for a cryptic exon if the PSI average for cryptic junctions was greater than or equal to 5%. We considered cryptic exons to have low level basal splicing if at least 1% of all samples in the SRA were positive for the cryptic exon.
First, we searched for the junction associated with the cryptic exon in STMN2, which leads to a polyadenylation event and subsequently reduces gene and protein expression40,57,62,64. Current clinical trials are assessing a STMN2-targeting ASO for ALS treatment, and the identification of non-ALS contexts of this cryptic exon inclusion could increase its potential therapeutic impact64,65.
Our findings demonstrate expression of the STMN2 cryptic exon in select neuroblastoma lines, including SMS-SAN, NLF, IMR-32, COG-N-415, NB-69, CHP-212, NB-1, NBL-S, LA-N-5, and COG-N-519 (Fig. 2d). While all these cell lines originate from pediatric neuroblastomas, there was little consistency regarding mutational profiles. For example, the NB-1 cell line has amplified MYC and ALK genes with wildtype levels of p53 RNA while the COG-N-415 line also has amplified MYC and wildtype p53 RNA levels but also has a F1174L mutation in ALK. Despite varying inclusion levels, it is notable that the STMN2 cryptic exon is present in multiple lines. This result was validated in the IMR-32 line. Of the tested cryptic exons, only STMN2 was present at robust levels without any treatment (Fig. S3l). There were also low levels of the ARHGAP32 cryptic exon. However, other cryptic exons associated with TDP-43 depletion in that cell line were not present. Overall, 4.6% of human SRA samples were positive for the cryptic splice site in STMN2. These results indicate that the expression of this cryptic exon is not limited to ALS-FTD and instead may have low level basal splicing in other cell types and conditions. Additionally, this suggests the STMN2 cryptic exon should not be used as a measure of TDP-43 depletion in these neuroblastoma cell lines, and perhaps others.
Next, we looked further into the inclusion of the cryptic exon in HDGFL2 in SRA samples. As previously mentioned, 26 samples had a PSI of greater than 5% but were not easily categorized into samples associated with cancer, ALS, or TDP-43 (Fig. 1c). Surprisingly, multiple samples corresponded to a knockdown of Serine and Arginine Rich Splicing Factor 3 (SRSF3). These samples came from a dataset in which TDP-43 and SRSF3 were knocked down either singularly or in concert. In MDA-MB231 cells, the two proteins were found to form a complex and the loss of either caused splicing dysregulation66. Analysis of the RNA-Seq reads displayed the inclusion of multiple cryptic exons, including that in HDGFL2, after SRSF3 knockdown only (Fig. 2e). Additionally, a combinatorial knockout of both TDP-43 and SRSF3 led to increased incorporation of the cryptic exons in some genes, including HDGFL2, PFKP, and TRRAP, when compared to the single TDP-43 knockdown. Given SRSF3’s established role in alternative splicing67, this supports the idea of the protein acting synergistically with TDP-43 to repress cryptic exons.
Querying human SRA database for potential biomarker cryptic exon inclusion level
Following the success of querying the SRA database for HDGFL2 and STMN2 cryptic exons, we mined the database for the junctions associated with cryptic exons in ACTL6B, AGRN, ATG4B, G3BP1, MYO18A, PFKP, RANBP1, UNC13A, and UNC13B (Fig. 3a, Fig. S1c). For cassette cryptic exons, we averaged the PSI of both junctions in each sample. We found that the inclusion of cryptic exons with a PSI above 5% occurred in less than 1% of samples for ACTL6B, AGRN, ATG4B, G3BP1, PFKP, RANBP1, and UNC13A. This indicates that, generally, these cryptic exons have not been detected within the samples stored in the SRA, whereas cryptic exons in MYO18A, STMN2, and UNC13B were included with a PSI greater than 5% in more than 1% of samples, potentially indicating greater low level basal splicing. This confirms the results from the GTEx database that also show greater rates of inclusion in normal tissue (Fig. 2b). Cryptic exons in STMN2 and UNC13B are included at greater rates than the other cryptic exons as they had a PSI of greater than 15% in more than 1% of the queried samples. Unsurprisingly, the top 20 samples in the SRA that included high rates of cryptic exon inclusion across the different targets were associated with ALS-FTD tissue or TDP-43 knockdown (Fig. 3b, Supplementary Table 2). The top 20 samples with no association with either TDP-43 or ALS-FTD were more varied and had less consistent expression of the panel of cryptic exons across replicates in the same experiment (Fig. S1d). Many of these samples were specifically identified to have nuclear clearance of TDP-43. This is consistent with the cryptic exons being repressed by TDP-43 normally and included when the protein is depleted or dysfunctional.

a–d SnapMine can be used to calculate the average percent spliced-in (PSI) values of cryptic exons (CEs) in samples from the Sequencing Read Archive (SRA). a, c Data are presented as box plots with median and interquartile range, along with whiskers from minimum to maximum values. a Visualization and quantification of PSI of human-specific CEs. b Heatmap visualization of the top 20 human samples in SRA with high inclusion of TDP-43-associated CEs. A gradient from white (PSI = 0%) to purple (PSI = 100%) is used to color the tiles. Gray tiles indicate the PSI cannot be calculated with certainty. All samples are associated with TDP-43 knockout (KO), knockdown (KD), or nuclear depletion (TDP-43-). c Visualization and quantification of PSI of mouse-specific CEs. d Heatmap visualization of the top 20 mouse samples in SRA with high inclusion of all selected TDP-43-associated CEs. A gradient from white (PSI = 0%) to purple (PSI = 100%) is used to color the tiles. Gray tiles indicate the PSI cannot be calculated with certainty. Although the majority are associated with TDP-43 KD, KO, or mutation (mut.), two samples stand out. The two samples correspond to CD4 + Th1 cells treated with ciclopirox olamine (CPX) in vitro. e UCSC Genome Browser visualization comparing RNA-seq reads of CEs in Adnp2, Synj2bp, Tbc1d1, Tecpr1, and Usp15 in TDP-43 KD and CPX treatment conditions. CD4 + Th1 cells treated with CPX incorporate cryptic exons at levels comparable to mouse neuronal and muscle cells with TDP-43 knocked down. Blue arrow indicates the cryptic exon. Lanes with red-colored reads indicate a TDP-43 KD or the CPX treatment. Lanes with black-colored reads are control samples. f Representative RT-PCR measurements of cryptic exon inclusion of mouse splenocyte culture treated with 20 µM CPX for four hours (n = 3). The black arrow points to the cryptic band. g Representative immunoblot of TDP-43 depletion in mouse splenocyte culture treated with 20 µM CPX for four hours (n = 3). TDP-43 protein levels are normalized to GAPDH protein levels. Bar graph quantifies presented TDP-43 protein levels in representative immunoblot with the black bar indicating the control and red bar indicating the CPX-treated sample. Source data are provided as a Source Data file.
Querying mouse SRA database for potential biomarker cryptic exon inclusion level
TDP-43-associated cryptic exons are nonconserved across species. Therefore, cryptic exons identified in mice cannot be explored in human data. Interestingly, TDP-43 has the same function of splicing repression in mice as it does in humans and still binds UG-rich motifs. Due to the ubiquitous expression of TDP-43 and conserved function, it is possible that upstream regulators of the protein are shared between species. Mouse cells are often subjected to genetic or environmental manipulations that would not be possible in human tissue. As a result, there are a greater variety of conditions available in mouse sequencing datasets. For this reason, we mined the mouse SRA for junctions associated with previously identified cryptic exons in Adnp2, Bud23, Crem, Fam135a, Hdac4, Ift81, Pnpla6, Slc7a6, Smg6, Spata7, Synj2bp, Tbc1d1, Tecpr1, and Unc13a (Fig. 3c). We found less than 1% of samples were positive for the cryptic exons in Bud23, Crem, Fam135a, Ift81, Spata7, Synj2bp, Tbc1d1, and Unc13a. We note that the cryptic exon in mouse Unc13a is different from the human UNC13A cryptic exon. They occur at different regions within the gene, are of different lengths, and have different sequences. The inclusion of cryptic exons with a PSI of 15% occurred in more than 1% of samples for Pnpla6 and Slc7a6, indicating low level basal splicing and the potential for other regulators of those splicing events.
As in the human samples, the majority of the top 20 samples that include cryptic exons are associated with TDP-43 manipulations, but two samples stood out from the rest. Like the TDP-43 depleted samples, they contained multiple cryptic exons with high PSI values. Uniquely, they had nothing to do with neurodegeneration, TDP-43, or splicing. Instead, these samples came from a study which treated mouse CD4 + T helper 1 (Th1) T cells with ciclopirox olamine (CPX), an FDA-approved antifungal medication that has also been studied for its properties as an iron chelator68,69 (Fig. 3d). After the treatment, multiple cryptic exons were spliced into the gene transcripts at exceptionally high rates compared to the control (Fig. 3e). These include the cryptic exons in Adnp2, Synj2bp, Tbc1d1, Tecpr1, and Usp15, which can also be identified in TDP-43 depleted mouse cells.
CPX treatment leads to robust cryptic exon inclusion across species and tissue types
In the original study, Th1 T cells were isolated and activated prior to a four hour treatment with 20 µM CPX68. To validate the treatment effect in immune cells, we simplified the experiment and treated a single-cell suspension generated from mouse splenocytes for four hours with 20 µM CPX. We then harvested the cells and confirmed cryptic exon inclusion using RT-PCR.
Our initial test of the treatment led to measurable inclusions of cryptic exons in mouse splenocyte transcripts of Adnp2, Synj2bp, Tbc1d1, Tecpr1, and Usp15 (Fig. 3f). Using protein immunoblot, we then confirmed that CPX treatment led to a protein level depletion of TDP-43 in the samples (Fig. 3g). This highlights the accuracy of querying the Snaptron databases and using SnapMine to find novel biological associations.
Due to the success in replicating cryptic exon inclusion after CPX treatment in splenocytes, we were also interested in whether TDP-43 protein dysregulation by CPX is a general phenomenon or specific to mouse splenocytes. Since TDP-43 dysregulation in ALS-FTD occurs in the nervous system, we first expanded our experiments to the mouse brain. We treated a single cell suspension of the mouse brain with 20 µM CPX for four hours, similar to the splenocytes. After the treatment, we conducted RT-PCR using single-band primers directed towards the cryptic exons in Ift81 and Unc13a, as well as double-band primers targeting Camk1g. We found that these cryptic exons are spliced into the mRNA transcripts after CPX treatment, indicating that TDP-43 is dysregulated (Fig. 4a). Additionally, treatment of a mouse brain in an ex vivo setting led to increased levels of cryptic exon inclusion in Ift81 and Unc13a when measured using Basescope ISH assays (Fig. 4f), suggesting that the application of CPX in vivo could be used to trigger and model TDP-43 dysregulation.

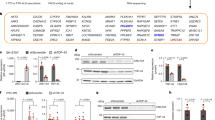
a–c Representative RT-PCR measurements of cryptic exon (CE) inclusion after cell culture treatment with 20 µM CPX for four hours. The black arrow points to the cryptic band. a Mouse brain (n = 2). The primers used to measure Ift81 and Unc13a cryptic exon inclusion target the CE directly, therefore no WT band is measured. b Human primary PBMCs (n = 2). c Human i3N (n = 2). d–f Representative immunoblots of TDP-43 depletion in mouse brain (n = 2), human PBMC (n = 2), and human i3N (n = 2) cultures treated with 20 µM CPX for four hours. TDP-43 protein levels are normalized to GAPDH or alpha-tubulin protein levels. e Representative immunoblot of TDP-43 and alpha-tubulin levels in RIPA-soluble and urea-soluble fractions in i3N. No aggregated, urea-soluble TDP-43 is measured (n = 2). f Bar graph quantifies presented TDP-43 protein levels in representative immunoblots. g Representative images for Basescope measurements of cryptic Ift81 and cryptic Unc13a inclusion of ex vivo treatment of mouse brain with 20 µM CPX for four hours (n = 4, 5 slices/n). Pink dots represent detected CE RNA. h Representative RT-PCR measurements of CE inclusion in human i3N culture after four-hour CPX treatment in combination with different small compound inhibitors (n = 2). Treatment with MG-132, Carfilzomib, or MLN4924 appears to increase cryptic exon inclusion rates while treatment with N-acetylcysteine (N-Ac) attenuates cryptic exon inclusion. i Representative RT-PCR measurements of CE inclusion in human i3N culture after four-hour CPX treatment in combination 5mM N-Ac (n = 2). j Heatmap visualization of average log2 fold change (log2FC) values of selected gene normalized area under the curve (NAUC) values, a measure of expression, between CPX-treated and WT conditions (n = 2 for each condition). A gradient from blue (log2FC of −5) to red (log2FC of 15) is used to color the tiles. I3Neurons and PBMCs, cell types that exhibit cryptic exons after treatment with CPX, exhibit strong gene upregulation of metallothioneins. Further analysis of genes upregulated in i3Neurons (Supplementary Fig. 4) confirms that heat shock response and oxidative stress pathways are upregulated, suggesting that heavy metal toxicity may be mediating CPX’s effect on TDP-43. Source data are provided as a Source Data file.
We then expanded our search to investigate whether the mechanism of TDP-43 dysregulation by CPX is conserved in humans. Due to more robust incorporation of cryptic exons in the mouse spleen as compared to the brain, we decided to test samples containing human immune cells first and treated human peripheral blood monocytes (PBMCs) with 20 µM CPX for four hours. After the treatment, we found incorporation of cryptic exons in PFKP, EPB41L4A, and AGRN (Fig. 4b). This conservation of the effect across species led us to then to treat iPSC-derived i3Neurons (i3N). We found that treated i3N incorporated cryptic exons, including those in PFKP, HDGFL2, EPB41L4A, IGLON5, UNC13A, and STMN2 (Fig. 4c). Additionally, we found that the STMN2 cryptic exon was sometimes incorporated at low levels in untreated i3N. These data indicated that the mechanism of TDP-43 dysregulation due to CPX treatment is not specific to mouse or immune cells and could be associated with pathways affected in ALS-FTD or other TDP-43 pathologies.
Our results confirmed CPX’s effect leading to TDP-43 depletion at a 20 µM concentration for four hours in a cell-type specific manner (Fig. S3a−c). Increasing the dosage and treatment time of CPX to 50 µM for 24 or 48 hours in human dermal fibroblasts was also sufficient to induce TDP-43 depletion and cryptic exons (Fig. S3d, e), although 48-hour treatments also greatly increased cell death of the samples. 100 µM treatment appears not to impact TDP-43 protein levels while still inducing cryptic exons. We hypothesize that treatment led to initial cell death of saturated cells while those surviving after 24 or 48 hours had a survivable dosage of the drug and TDP-43 depletion (Fig. S3d, e). Additionally, CPX had the same effect in IMR-32 cells (Fig. S3l−k).
These results confirmed the effects of CPX in both cell-type and dosage-dependent manner, but there were still several potential mechanisms leading to its impact. During ALS-FTD, TDP-43 is normally depleted from the nucleus and sequestered in cytoplasmic aggregates. Immunoblot measurements of TDP-43 protein indicated CPX led to protein level depletion in these additional cell types although the extent of depletion varied by tissue type (Fig. 4d). However, TDP-43 is not found in the urea-soluble fraction of protein lysate after CPX treatment, suggesting aggregation is not occurring (Fig. 4e). These results indicated CPX treatment leads to increased degradation of TDP-43 protein.
Next, we aimed to determine the potential mechanism impacted by CPX, likely upstream of TDP-43 degradation. Initially, we pre-incubated mouse splenocyte samples with inhibitors of proteolysis or protein modification such as EST, a cysteine protease inhibitor, MG-132, a proteasome inhibitor, and PYR-41, a ubiquitin E1 inhibitor. We found that PYR-41 treatment reduced the level of cryptic exon inclusion, but further analysis found that it appears to have reduced the expression of many proteins in the cell, likely impacting many other pathways beyond TDP-43 (Fig. S3f, g). To test a more varied set of inhibitors in a human-relevant context, we pre-incubated the i3N with various inhibitors to determine whether TDP-43 function could be restored or diminished to a lesser level in the treated cells.
Using cryptic exon inclusion in HDGFL2, EPB41L4A, PFKP, and UNC13A as a readout of TDP-43 function, we found that certain inhibitors of the proteasomal degradation system, MG-132, Carfilzomib, and MLN4924, consistently led to an increase in cryptic exon inclusion although they were not sufficient on their own to induce cryptic exons (Fig. 4g, Fig. S2c). MG-132 and Carfilzomib directly inhibit the proteasome while MLN4924 impacts neddylation, another process involved in regulating proteasomal degradation of proteins70. Additionally, pre-treatment with N-acetylcysteine (N-Ac), a reactive oxygen species (ROS) inhibitor, led to a noticeable decrease in cryptic exon inclusion in both i3N and IMR-32 cells (Fig. 4g, h, Fig. S3h, k). These results are consistent with an upstream mechanism of TDP-43 regulation that is partially dependent on proteasome and neddylation-associated degradation. Overall, our data suggest proteasome inhibitors, in conjunction with CPX, increase TDP-43 depletion, whereas ROS inhibitors have the opposite effect, partially rescuing the CPX-induced depletion.
We performed RNA-Seq on i3N, PBMCs, microglia, and fibroblasts to study gene expression changes between CPX-treated and untreated cells. Among these human primary cells, CPX induces cryptic exons only in i3N and PBMCs, while microglia and fibroblasts appear to be resistant. Our RNA-Seq data (Fig. 4j) revealed significant upregulation of gene signatures specific to i3Neurons and PBMCs. Notably, these signatures did not appear in microglia and fibroblasts, which did not show cryptic exon inclusion after CPX treatment. Among the upregulated genes, metallothioneins were prominently upregulated in both i3N and PBMCs and a STRING network analysis71 of genes upregulated in i3N (Fig. S4) confirms the activation of heat shock response and oxidative stress pathways. These results suggest that cellular and oxidative stress responses to heavy metals might be an underlying mechanism through which CPX exerts its effects on TDP-43, opening new avenues for understanding and potentially mitigating TDP-43 pathology in ALS-FTD and related disorders.
Finally, to further investigate the connection between iron dysregulation and TDP-43 pathology, we conducted superpixel analysis of immunohistochemical staining for ferritin and TDP-43 aptamer (TDP-43APT)27,72,73 in ALS cases and controls. Aptamers are artificial oligonucleotides capable of to binding with high specificity to a target, similar to antibodies. This RNA aptamer is highly sensitive and can be used to identify pathological TDP-43 aggregation events, including in pre-symptomatic cases74. Figure S5 provides compelling evidence of iron dysregulation in ALS. For gray matter, histological iron burden detection is well represented by immunostaining with a ferritin antibody. The analysis reveals that ferritin staining is markedly elevated in ALS cases compared to controls (Fig. S5a−e), indicating significant iron accumulation that is absent in the control samples. Ferritin staining is low in controls, indicative of normal iron regulation (Fig. S5d), while ALS cases exhibit high ferritin levels, suggesting iron dysregulation and increased non-circulating iron burden (Fig. S5e). Furthermore, GPX4 levels are slightly decreased in IMR-32 cells by CPX treatment and restored by N-Ac treatment (Fig. S4k). GPX4 protein inhibits lipid peroxidation (Fig. S5f). Together, the induction of TDP-43 cryptic exons by CPX and the detection of abnormal ferritin staining suggest that iron homeostasis may play a critical role in ALS pathogenesis.
Discussion
In this work, we have created an application that can be used to analyze thousands of samples in GTEx, TCGA, ENCODE, and SRA compilations for alternative splicing patterns of interest. The R script used to query for multiple cryptic exons simultaneously is available on GitHub and can be used by researchers interested in customizing the script. The application queries the Snaptron database of exon-exon junctions of interest and calculates the exon PSI values in each sample available. The output table includes metadata for each sample as annotated by Snaptron and can be downloaded as a csv file. Using the application, we were able to query the inclusion of TDP-43-associated cryptic exons in these databases and better understand baseline inclusions for consideration of each as a suitable biomarker.
To validate our application, we queried the inclusion of TDP-43-associated cryptic exons in these data compilations. GTEx includes biospecimens from donors collected within 24 h of death and eligibility criteria is broad. It is therefore reasonable to expect that many of the donor organs will not be impacted by disease. Using this data, we determined that some cryptic exons can be found within organs without a TDP-43-associated deficit. Additionally, cryptic exon enrichment can differ on a tissue-to-tissue basis. Moving forward, the application can be used to determine baseline levels of cryptic exons and screen them in a tissue-specific manner for use as biomarkers or therapeutic targets. By doing so, biomarker specificity can be tested, and off-target effects of therapeutics can be avoided. Other researchers will also be able to use the application to mine public data for other novel biological contexts causing splicing abnormalities.
Our preliminary analysis of the initial dataset of TDP-43-associated cryptic exon inclusion implicated CPX, an FDA-approved anti-fungal, as capable of causing TDP-43 depletion in a tissue-specific manner in as little as four hours. However, as the original dataset used CPX as an iron chelator, there were no explicit connections to TDP-43-associated pathology. Currently, CPX is available in prescription shampoo and cream formulations and off-brand uses include testing as an anti-cancer agent75,76,77,78,79,80,81. Using the application, we were able to clearly identify TDP-43-associated cryptic exons in these CPX-treated samples and conduct further testing to explore the mechanism CPX targets (Fig. 5). Our in vitro experiments indicate the importance of caution when considering alternative applications of CPX, as it may impact proteins such as TDP-43.

RNA is depicted with blue and red bars representing canonical and cryptic exons, respectively. The black line connecting the exons represents the introns. (top) Snaptron Sequencing Read Archive (SRA), The Cancer Genome Atlas (TCGA), and the Genotype-Tissue Expression (GTEx) project data compilations were queried for TDP-43-associated cryptic exons. Ciclopirox olamine (CPX) was identified as a candidate for TDP-43 modulation. (bottom) CPX was validated to induce TDP-43 degradation, cryptic exons, and increased oxidative stress and metallothionein upregulation in various cell-types. N-Acetylcysteine (N-Ac) was found to inhibit this effect.
Since we screened datasets using the average PSI value of multiple cryptic exons, there are likely other contexts in which single cryptic events occur. This provides the opportunity to find other upstream regulators of splice events. While we focused on TDP-43, it would be interesting to look at the targets of other RBPs, such as PTBP1 or MATRIN312,16,17,19,82,83,84. Another potential future use would be to identify novel contexts of known alternative splicing and then re-analyze the RNA-Seq data from implicated samples for new, context-specific RNA targets of the RBPs.
Our validation results with CPX also suggest that heavy metal toxicity and oxidative stress may be sufficient to induce TDP-43 dysregulation. Given that acute four hour treatments with CPX caused TDP-43 protein depletion, upstream mechanisms are likely related to protein modification or degradation process that occurs over short timespans. Additionally, we were able to show the process is conserved across multiple types of primary cell types beyond mouse Th1 T cells. Although not all cell types had TDP-43 dysregulation after 20 µM CPX treatment, its efficacy in human PBMCs and i3Neurons suggests that CPX treatment could be used in the future to create models of TDP-43 depletion. It is also possible that some cell types are more resistant to CPX-induced TDP-43 dysregulation and require higher levels or longer treatment, as is seen with the HDFs.
Our work also supports prior research indicating increased TDP-43 dysfunction following proteasomal inhibition by MG-13285. While much of the previous research with TDP-43 and MG-132 focuses on ubiquitination and insolubility, 24 h treatment with MG-132 also leads to the inclusion of cryptic STMN2 in human motor neurons, which is indicative of nuclear depletion57. While we did not find the four-hour treatment of MG-132 sufficient for cryptic exon inclusion in i3Ns, combined treatment with both CPX and MG-132 led to increased cryptic exon inclusion when compared to only CPX. Supporting this hypothesis is the observation that treatment with Carfilzomib, a proteasome inhibitor used therapeutically, also increases cryptic exon inclusion when supplemented with CPX. This suggests proteasomal pathways play a role in the stability of TDP-43 protein but are not the sole mechanism through which TDP-43 is degraded.
Since proteasomal inhibition led to worsened TDP-43 pathology, we hypothesize that CPX targets a different pathway which influences TDP-43 stability. If so, the combined treatment of CPX and proteasome inhibition would inhibit two different regulatory pathways of TDP-43 and increase its dysregulation. Previous treatment of cells with both niclosamide and a stressor found that treatment reduced levels of oxidative stress and TDP-43 pathology85. Others have found that N-Ac treatment to reduce ROS has also reduced arsenite-induced stress granule formation and TDP-43 pathology86,87. Our findings suggest CPX is another stressor that could lead to increased ROS, given that N-Ac treatment in combination with CPX partially alleviates cryptic exon burden. Some prior studies have also linked oxidative stress to TDP-43 aggregation and various neurodegenerative deficits88,89,90. Thus, the observation that oxidative stress might contribute to nuclear clearance of the protein is compelling and points towards a common mechanism for TDP-43 pathology. Further investigation of the pathway through which CPX affects TDP-43 could elucidate information regarding endogenous regulators of TDP-43 and pave the way for the development of therapies that modulate these pathways in ALS-FTD and related dementias.
Superpixel analysis of ferritin and TDP-43APT staining supports the idea that iron dysregulation may occur in ALS91,92,93,94,95. Elevated ferritin levels in ALS cases indicate significant iron accumulation, which is not observed in control tissues. This iron dysregulation aligns with the potential mechanism of CPX, which upregulates ferric permeases and reductases, increasing Fe2+ levels69,96,97,98. Fe2+ acts as a cofactor in lipid oxidation via LOX and contributes to oxidative stress through Fenton’s reaction99,100,101, processes that can lead to TDP-43 dysregulation. Furthermore, past studies have similarly identified iron dysregulation in ALS cases92,93,102,103,104,105,106,107,108. Thus, it is possible that iron dysregulation may directly contribute to the initiation and progression of TDP-43-related neurodegeneration. Our results underscore the need for further research into the critical role of oxidative stress and iron homeostasis in ALS. Understanding this link opens new avenues for exploring therapeutic strategies aimed at mitigating TDP-43-associated neurodegeneration.
Methods
Animal handling
All the animal experiments were conducted as per the regulations of the Animal Care and Use Committee at Johns Hopkins University School of Medicine in accordance with the laws of the State of Maryland and the United States of America. C57BL/6 J mice, aged 4–8 weeks, were used for experiments. Mice (4-5/cage) were maintained in a facility with a 12-h light/dark cycle. Facility temperature and humidity are maintained between 68–79 F and 30–70%, respectively. Food and water were provided ad libitum.
Analysis of published RNA-Seq Data
FASTQ files were downloaded from the NCBI’s Sequence Read Archive and aligned to the GRCh38 human genome assembly using STAR (v.2.7.10a) with default parameters109. Megadepth was used to convert the output BAM files to BigWig files, and the data were visualized on the UCSC Genome Browser (http://genome.ucsc.edu/)110,111,112. Computational resources from the Advanced Research Computing core facility at Johns Hopkins University were used to conduct RNA-Seq analysis.
RNA-Sequencing and differential gene expression analysis
Libraries for RNA sequencing were prepared using a Stranded mRNA library preparation kit (Illumina) following the manufacturer’s protocol. Sequencing was performed using an Illumina NovaSeq S4 300 to generate 150 bp paired-end reads. Reads were aligned to the GRCh38 human genome assembly using STAR (v.2.7.10a) with default parameters109. Similar to past publications48,49,51, gene expression levels were quantified using featureCounts113 and differential gene expression by calculating log2 fold change of each measured gene’s normalized area under the curve (NAUC) between conditions. NAUC was filtered so only genes with a value greater than 0.05 in both conditions for each cell type were plotted.
Analysis of Snaptron data
All analysis was conducted using R version 4.3.2 on RStudio (R Core Team 2023). Inclusion and exclusion junction hg38 or mm10 coordinates were identified and then Snaptron junction counts for each sample were downloaded. Due to the nonconserved nature of cryptic exons, they differ between human and mouse queries. PSI was calculated for each sample using the equation:
If the total number of junction counts, or the denominator of the above fraction, was less than 15, the sample was excluded from the analysis. Metadata downloaded from Snaptron was merged with the output PSI table. The tidyverse package was used to manipulate the data tables for analysis114.
GEX counts were extracted for each sample from Snaptron using gene hg38 or mm10 gene coordinates and Ensembl gene ID of the queried gene. If normalization was desired, the counts were also extracted for the gene that is used for normalization. Relative quantitative gene count was calculated as follows. Normalization factor was calculated by determining the highest expression count of the normalization gene measured within a single study. For each sample, the normalization factor was the expression count of the normalization gene of the sample divided by the maximum value measured within the relevant study. The normalized count for the gene of interest was the gene expression count for the sample divided by the normalization factor for the sample. If there was no measurement of the normalization gene in a sample or study group, those sample(s) were marked as not normalized. This is depicted below.
Samples in which the cryptic exon PSI was greater than or equal to 5% were considered positive for a cryptic exon. Individuals with greater than five unique cryptic exons in a single tissue type were considered individuals with potential TDP-43 pathology in the GTEx database.
Visualization of Snaptron data was completed using the ggplot2 and viridis packages in R115. Scripts for visualization are available at https://github.com/irikas/SnapMine_Paper. Final figures were made in Adobe Illustrator.
SnapMine application
Usage of the above code for junction or gene expression count extraction and analysis from Snaptron is available at https://snapmine.idies.jhu.edu.
Users can use the web application to query the TCGA, SRA, GTEx, and ENCODE shRNA compilations for junctions of interest in available samples. These data were processed during the creation of recount3 and include sequencing data from TCGA v3.0, GTEx v8, SRA human datasets as of 10/06/2019, and SRA mouse datasets as of 01/08/202048,49. The ENCODE shRNA-Seq data were processed by Snaptron and uploaded 06/201851.
The results are displayed in an interactive table where data can be further filtered based on user criteria. The results can also be downloaded into a CSV file. The metadata files used are the original files on Snaptron. Code used to generate the results on SnapMine can be found at https://github.com/ssec-jhu/snaptron-query. SnapMine is developed by SSEC engineers and hosted at IDIES at JHU. It is maintained by F.A., I.R.S., and SSEC. Issues can be submitted to GitHub.
Cell culture
To isolate mouse organs, the mouse was euthanized using carbon dioxide and perfused with PBS. Mouse spleen and brain were harvested as needed. To create a mouse splenocyte culture, the spleen was filtered through a 70 µm cell strainer, spun down, and red blood cells were removed using ACK Lysis buffer (Gibco #A1049201). Cells were resuspended in RPMI supplemented with 10% FBS and 1X Penicillin-Streptomycin (Gibco #15140122). To create a mouse brain cell culture, the brain was enzymatically dissociated (Miltenyi #130-107-677). Cells were resuspended in DMEM (Gibco #11960044) supplemented with 1X GlutaMAX (Gibco #35050061), 10% FBS, and 1X Penicillin-Streptomycin (Gibco #15140122). Cells were plated for half an hour and then treated.
The i3Neuron (i3N) iPSCs were cultured, grown, and differentiated as previously detailed116,117. Neurons were treated on day 7 post-differentiation in cortical media. Preliminary human PBMC data was from PBMCs harvested from a healthy donor and dissociated by and Viral Immune Pathogenesis Center (VIPC) at Johns Hopkins University. Primary Peripheral Blood Mononuclear Cells (PBMC), Normal, Human (ATCC #PCS-800-01, Lot #80104236) were thawed as recommended and resuspended in RPMI 1640 medium (Gibco #11875093) with 1X GlutaMAX (Gibco #35050061), 10% FBS, and 1X Penicillin-Streptomycin (Gibco #15140122). They were plated for half an hour and then treated. Human dermal fibroblasts (HDFs) were obtained from multiple sources. First, sequenced HDFs were from a healthy, 32 M subject (Coriell Institute #2003-071-056) from the Baltimore Longitudinal Study of Aging. HDF for further testing (ATCC # PCS-201-012, Lot #8405992, #80512242) were from healthy 31 F and 51 F individuals. HDF were cultured in DMEM, high glucose, HEPES medium (Gibco #12430054) with 10% FBS and 1X Penicillin-Streptomycin (Gibco #15140122). iPSC-derived microglia (Fujifilm iCell Microglia #01279, Lot # 106714) were thawed and cultured as per manufacturer recommendations. IMR-32 cells (ATCC # CCL-127, Lot #70058104) were thawed as recommended and resuspended and maintained in DMEM medium (Gibco #11875093) with DMEM (Gibco #11960044) supplemented with 1X GlutaMAX (Gibco #35050061), 10% FBS, and 1X Penicillin-Streptomycin (Gibco #15140122). All cells were incubated at 37 °C and 5% CO2 during treatment and incubation/recovery steps.
CPX treatment
Ciclopirox olamine, CPX, (Millipore Sigma #1134030-125MG) was solubilized in ethanol to 100 mM and 10 mM. Plated cells were treated with 20 µM CPX, or other concentration as indicated, in media for four hours in a 37 °C tissue culture incubator. Control cells were treated with an equal volume of 200 proof ethanol in media. Cells were harvested after the treatment period and spun down. The supernatant was aspirated and the cells were flash-frozen on dry ice for downstream analysis.
Inhibitor treatment
Plated i3N were incubated with inhibitors at given concentrations for 30 minutes. After the pre-treatment period, cells were incubated with or without CPX for four additional hours prior to harvest. The inhibitors, solvents, and concentrations tested are given in Supplementary Table 3.
RT-PCR analysis
RNA was extracted from samples using the Monarch® Total RNA Miniprep Kit (New England Biolabs, #T2010S), and cDNA was synthesized using the ProtoScript II First Strand cDNA Synthesis Kit (New England Biolabs, #E6560L). cDNA was amplified using primer pairs designed to target either two wildtype sequences flanking the cryptic exon or one wildtype sequence and another sequence directly within the cryptic exon. Target sequence amplification was performed using Phusion Plus Green PCR Master Mix (Thermo Scientific, #F632L) and a modified touchdown PCR protocol118, described in Supplementary Table 4. Amplified targets were then separated by 1.5% agarose gel electrophoresis and visualized by ethidium bromide or GelRed (Biotium #41001) staining.
We used two types of primer pairs to amplify targets (Supplementary Table 5). The first type comprises a primer targeting the cryptic exon and another targeting an adjacent canonical exon, which we term single-band primers. Consequently, RT-PCR amplification would occur only when transcripts containing cryptic exons are in the sample. This leads to the amplification of cryptic exon-containing RNA at low levels. In the second type, both primers bind canonical mRNA flanking the cryptic exon, which we term double-band primers. Accordingly, two potential transcripts are amplified through RT-PCR: the wildtype and the heavier cryptic exon-including one.
Protein immunoblot and analysis
Samples were collected from wells by scraping and suspension in PBS. The cell suspensions was centrifuged at 1600 x g for 10 min to pellet the cells. PBS was aspirated, and the cell pellets were then resuspended in RIPA Lysis and Extraction Buffer (Thermo Scientific, #89900) with Halt Protease Inhibitor Cocktail (Thermo Scientific, #78430). After 30 min rotating at 4 C, the extracts were centrifuged for 30 min at 4 °C and 20,000 x g. The supernatants were collected as the RIPA-soluble fraction. To detect urea-soluble proteins, the remaining cell-pellets were resuspended in urea buffer (30 mM Tris, pH 8.8, 7 M urea, 2 M thiourea, and 4% CHAPS) and sonicated. The lysates were then centrifuged at 100,000 x g for 30 min at 22 °C in an ultracentrifuge. The supernatants were collected as the urea-soluble fraction. The Pierce BCA Protein Assay kit (Thermo Scientific, #23225) was used to quantify protein lysate concentration.
Protein blot analysis was performed following electrophoresis of reduced samples using NuPAGE 4 to 12% Bis-Tris Mini Polyacrylamide Gels (Thermo Scientific, #NP0322BOX). Proteins were then transferred to PVDF membranes using the iBlot 2 Dry Blotting System from Invitrogen. The membrane was then used for western blot detection in one of two ways.
-
1.
The membrane was then incubated in blocking buffer, 5% dry nonfat milk in Tris-buffered saline with 0.1% Tween (TBS-T). Membranes were probed with primary antibody (Supplementary Table 6) in blocking buffer either overnight at 4 °C or for two hours at room temperature with rocking. Secondary conjugated to HRP in blocking buffer was applied the next day for one hour at room temperature for antibodies not pre-conjugated to HRP.
-
2.
Alternatively, the membrane was pre-activated in methanol, soaked in iBind™ Flex Solution (ThermoScientific, #SLF2020), and probed using the iBind™ Flex Western Device (ThermoScientific, #SLF2000) according to the manufacturer’s protocol.
Immobilon® ECL UltraPlus Western HRP Substrate (Millipore Sigma, #WBULP-100ML) was used for detection in both cases. The BioRad ChemiDoc Imaging System was used to image the blots. To detect other targets, membranes were stripped using Restore™ PLUS Western Blot Stripping Buffer (Thermo Scientific, #46430) and re-probed. Detection occurred as previously described.
To detect N-terminal TDP-43, antibody 10782-2-AP (ProteinTech) was pre-conjugated with HRP using the HRP Conjugation Kit - Lightning-Link (abcam ab102890).
Superpixel analysis of TDP-43APT and ferritin immunohistochemistry
Brain tissue from standardized Brodmann areas (BA) was post-mortem fixed in 10% formalin for at least 48 h, dehydrated, and embedded in paraffin. The formalin-fixed paraffin-embedded (FFPE) tissue was sectioned into 4 μm slices, dried, and immunostained using the Novolink Polymer detection system with anti-Ferritin antibody (Supplementary Table 6) and TDP-43APT staining as described by standard protocols27,72. Slides were scanned at 20X magnification, and three regions of interest (ROI) per slide were analyzed using QuPath119 software for superpixel analysis. The script for analysis is available at https://github.com/irikas/SnapMine_Paper. The analysis involved setting image type and color deconvolution stains, selecting annotations and detections, and running plugins for superpixel and intensity feature extraction. Pixel classification based on mean pixel intensity identified 1+, 2+, and 3+ detections corresponding to one, two, and three standard deviations above the mean, respectively. Digital Burden Score was calculated.
BaseScope-ISH and co-detection assay
RNA in-situ hybridization was performed using BaseScope Detection Reagent v2-RED Assay Kit (Advanced Cell Diagnostics, Inc. #323900), following manufacturer’s instructions. Expression of transcripts containing cryptic exon splice sites in Unc13a and Ift81 transcripts was detected using 3zz custom BaseScope probes (BA-Mm-Unc13a-O1-2EJ-C and BA-Mm-Ift81-E17-intron17-NJ, respectively). Both positive and negative control probes were employed to assess the RNA quality (BA-Mm-Ppib and BA-DapB). Briefly, consecutive tissue sections of 10 μm thickness underwent de-paraffinization followed by pre-treatment with hydrogen peroxide, target retrieval buffer, and protease IV. Subsequently, these sections were subject to hybridization with target probes in a HybEZII oven (Advanced Cell Diagnostics, Inc.) for 2 hours at 40 °C. The signals were amplified, and the slides were counterstained with hematoxylin. Images were acquired using a Zeiss Apotome Inverted Brightfield Microscope (Zeiss, Germany), and RNA puncta for each cryptic exon were manually quantified using ImageJ software.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The application to query junction inclusion data is hosted at https://snapmine.idies.jhu.edu. RNA-Seq data have been deposited in the NCBI Sequence Read Archive. They are associated with BioProject accession PRJNA1242253. The RNA-Seq data generated in this study have been deposited in the NCBI Sequence Read Archive under SRA study number SRP573260 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP573260]. The publicly deposited RNA-Seq data used this study are available on the NCBI Sequence Read Archive under SRA study numbers SRP16628257 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP166282], SRP05781911 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP057819], SRP092413120 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP092413], SRP10440566 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP104405], SRP194266121 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP194266], SRP367696122 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP367696], and SRP07923668 [https://trace.ncbi.nlm.nih.gov/Traces/study/?acc=SRP079236]. The NAUC data used in this study are available on ASCOT51. Source data are provided with this paper.
Code availability
Early SnapMine interface and visualization scripts are available in the following GitHub repository: https://github.com/irikas/SnapMine_Paper. Code for the SnapMine web application is available at https://github.com/ssec-jhu/snaptron-query and on Zenodo123.
References
Chen, M. & Manley, J. L. Mechanisms of alternative splicing regulation: insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 10, 741–754 (2009).
Wahl, M. C., Will, C. L. & Lührmann, R. The spliceosome: design principles of a dynamic RNP machine. Cell 136, 701–718 (2009).
Wang, E. T. et al. Alternative isoform regulation in human tissue transcriptomes. Nature 456, 470–476 (2008).
Matlin, A. J., Clark, F. & Smith, C. W. J. Understanding alternative splicing: towards a cellular code. Nat. Rev. Mol. Cell Biol. 6, 386–398 (2005).
Pan, Q., Shai, O., Lee, L. J., Frey, B. J. & Blencowe, B. J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 40, 1413–1415 (2008).
Yang, X. et al. Widespread expansion of protein interaction capabilities by alternative splicing. Cell 164, 805–817 (2016).
Raj, B. & Blencowe, B. J. Alternative splicing in the mammalian nervous system: recent insights into mechanisms and functional roles. Neuron 87, 14–27 (2015).
Nikom, D. & Zheng, S. Alternative splicing in neurodegenerative disease and the promise of RNA therapies. Nat. Rev. Neurosci. 24, 457–473 (2023).
Wright, C. J., Smith, C. W. J. & Jiggins, C. D. Alternative splicing as a source of phenotypic diversity. Nat. Rev. Genet. 23, 697–710 (2022).
Sibley, C. R., Blazquez, L. & Ule, J. Lessons from non-canonical splicing. Nat. Rev. Genet. 17, 407–421 (2016).
Ling, J. P., Pletnikova, O., Troncoso, J. C. & Wong, P. C. TDP-43 repression of nonconserved cryptic exons is compromised in ALS-FTD. Science 349, 650–655 (2015).
Ling, J. P. et al. PTBP1 and PTBP2 repress nonconserved cryptic exons. Cell Rep. 17, 104–113 (2016).
McClory, S. P., Lynch, K. W. & Ling, J. P. HnRNP L represses cryptic exons. RNA 24, 761–768 (2018).
Zarnack, K. et al. Direct competition between hnRNP C and U2AF65 protects the transcriptome from the exonization of alu elements. Cell 152, 453–466 (2013).
Ehrmann, I. et al. An ancient germ cell-specific RNA-binding protein protects the germline from cryptic splice site poisoning. eLife 8, e39304 (2019).
Khan, M. et al. MATR3 pathogenic variants differentially impair its cryptic splicing repression function. FEBS Lett. 598, 415–436 (2024).
Tan, Q. et al. Extensive cryptic splicing upon loss of RBM17 and TDP43 in neurodegeneration models. Hum. Mol. Genet. 25, ddw337 (2016).
Carmen-Orozco, R. P. et al. Elevated nuclear TDP-43 induces constitutive exon skipping. Mol. Neurodegener. 18, 45 (2024).
Eom, T. et al. NOVA-dependent regulation of cryptic NMD exons controls synaptic protein levels after seizure. eLife 2, e00178 (2013).
Jangi, M., Boutz, P. L., Paul, P. & Sharp, P. A. Rbfox2 controls autoregulation in RNA-binding protein networks. Genes Dev. 28, 637–651 (2014).
Lukavsky, P. J. et al. Molecular basis of UG-rich RNA recognition by the human splicing factor TDP-43. Nat. Struct. Mol. Biol. 20, 1443–1449 (2013).
Humphrey, J., Emmett, W., Fratta, P., Isaacs, A. M. & Plagnol, V. Quantitative analysis of cryptic splicing associated with TDP-43 depletion. BMC Med. Genomics 10, 38 (2017).
Mehta, P. R., Brown, A.-L., Ward, M. E. & Fratta, P. The era of cryptic exons: implications for ALS-FTD. Mol. Neurodegener. 18, 16 (2023).
Hong, D. & Jeong, S. 3’UTR diversity: expanding repertoire of RNA alterations in human mRNAs. Mol. Cells 46, 48–56 (2023).
Neumann, M. et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314, 130–133 (2006).
Ling, S.-C., Polymenidou, M. & Cleveland, D. W. Converging mechanisms in ALS and FTD: disrupted RNA and protein homeostasis. Neuron 79, 416–438 (2013).
Spence, H. et al. RNA aptamer reveals nuclear TDP-43 pathology is an early aggregation event that coincides with STMN-2 cryptic splicing and precedes clinical manifestation in ALS. Acta Neuropathol. 147, 50 (2024).
Chang, K. et al. Loss of TDP-43 splicing repression occurs early in the aging population and is associated with Alzheimer’s disease neuropathologic changes and cognitive decline. Acta Neuropathol. 147, 4 (2024).
Boer, E. M. J. et al. TDP-43 proteinopathies: a new wave of neurodegenerative diseases. J. Neurol. Neurosurg. Psychiatry 92, 86–95 (2021).
Masaki, K. et al. RNA-binding protein altered expression and mislocalization in MS. Neurol. Neuroimmunol. Neuroinflammation 7, e704 (2020).
Nelson, P. T. et al. Limbic-predominant age-related TDP-43 encephalopathy (LATE): consensus working group report. Brain 142, 1503–1527 (2019).
McKee, A. C. et al. TDP-43 proteinopathy and motor neuron disease in chronic traumatic encephalopathy. J. Neuropathol. Exp. Neurol. 69, 918–929 (2010).
Higashi, S. et al. Concurrence of TDP-43, tau, and α-synuclein pathology in brains of Alzheimer’s disease and dementia with Lewy bodies. Brain Res. 1184, 284–294 (2007).
Ayuso, V. E. et al. TDP-43-regulated cryptic RNAs accumulate in Alzheimer’s disease brains. Mol. Neurodegener. 18, 57 (2023).
Josephs, K. A. et al. Staging TDP-43 pathology in Alzheimer’s disease. Acta Neuropathol. 127, 441–450 (2014).
Josephs, K. A. et al. Updated TDP-43 in Alzheimer’s disease staging scheme. Acta Neuropathol. 131, 571–585 (2016).
Weihl, C. C. et al. TDP-43 accumulation in inclusion body myopathy muscle suggests a common pathogenic mechanism with frontotemporal dementia. J. Neurol. Neurosurg. Psychiatry 79, 1186 (2008).
LaClair, K. D. et al. Depletion of TDP-43 decreases fibril and plaque β-amyloid and exacerbates neurodegeneration in an Alzheimer’s mouse model. Acta Neuropathol. 132, 859–873 (2016).
Meneses, A. et al. TDP-43 pathology in Alzheimer’s disease. Mol. Neurodegener. 16, 84 (2021).
Quadros, A. R. A. A. et al. Cryptic splicing of stathmin-2 and UNC13A mRNAs is a pathological hallmark of TDP-43-associated Alzheimer’s disease. Acta Neuropathol. 147, 9 (2024).
Pickles, S. R. et al. TDP‐43 cryptic RNAs in perry syndrome: differences across brain regions and TDP‐43 proteinopathies. Mov. Disord. 40, 662–671 (2025).
Jeong, Y. H. et al. Tdp-43 cryptic exons are highly variable between cell types. Mol. Neurodegener. 12, 13 (2017).
Šušnjar, U. et al. Cell environment shapes TDP-43 function with implications in neuronal and muscle disease. Commun. Biol. 5, 314 (2022).
Cao, M. C. et al. A panel of TDP-43-regulated splicing events verifies loss of TDP-43 function in amyotrophic lateral sclerosis brain tissue. Neurobiol. Dis. 185, 106245 (2023).
Irwin, K. E. et al. A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS–FTD. Nat. Med. 30, 382–393 (2024).
Irwin, K. E., Sheth, U., Wong, P. C. & Gendron, T. F. Fluid biomarkers for amyotrophic lateral sclerosis: a review. Mol. Neurodegener. 19, 9 (2024).
Seddighi, S. et al. Mis-spliced transcripts generate de novo proteins in TDP-43-related ALS/FTD. Sci. Transl. Med. 16, eadg7162 (2024).
Wilks, C., Gaddipati, P., Nellore, A. & Langmead, B. Snaptron: querying splicing patterns across tens of thousands of RNA-seq samples. Bioinformatics 34, 114–116 (2018).
Wilks, C. et al. recount3: summaries and queries for large-scale RNA-seq expression and splicing. Genome Biol. 22, 323 (2021).
Nellore, A. et al. Human splicing diversity and the extent of unannotated splice junctions across human RNA-seq samples on the Sequence Read Archive. Genome Biol. 17, 266 (2016).
Ling, J. P. et al. ASCOT identifies key regulators of neuronal subtype-specific splicing. Nat. Commun. 11, 137 (2020).
Leinonen, R., Sugawara, H., Shumway, M. & Collaboration, I. N. S. D. The sequence read archive. Nucleic Acids Res 39, D19–D21 (2011).
Consortium, T. Gte. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 369, 1318–1330 (2020).
Chang, K. et al. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet 45, 1113–1120 (2013).
Sundararaman, B. et al. Resources for the comprehensive discovery of functional RNA elements. Mol. Cell 61, 903–913 (2016).
Sloan, C. A. et al. ENCODE data at the ENCODE portal. Nucleic Acids Res 44, D726–D732 (2016).
Klim, J. R. et al. ALS-implicated protein TDP-43 sustains levels of STMN2, a mediator of motor neuron growth and repair. Nat. Neurosci. 22, 167–179 (2019).
Melamed, Z. et al. Premature polyadenylation-mediated loss of stathmin-2 is a hallmark of TDP-43-dependent neurodegeneration. Nat. Neurosci. 22, 180–190 (2019).
Ma, X. R. et al. TDP-43 represses cryptic exon inclusion in the FTD–ALS gene UNC13A. Nature 603, 124–130 (2022).
Brown, A.-L. et al. TDP-43 loss and ALS-risk SNPs drive mis-splicing and depletion of UNC13A. Nature 603, 131–137 (2022).
Prudencio, M. et al. Truncated stathmin-2 is a marker of TDP-43 pathology in frontotemporal dementia. J. Clin. Investig. 130, 6080–6092 (2020).
Baughn, M. W. et al. Mechanism of STMN2 cryptic splice-polyadenylation and its correction for TDP-43 proteinopathies. Science 379, 1140–1149 (2023).
Lonsdale, J. et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 45, 580–585 (2013).
López-Erauskin, J. et al. Stathmin-2 loss leads to neurofilament-dependent axonal collapse, driving motor and sensory denervation. Nat. Neurosci. 27, 34–47 (2024).
Daele, S. H. V., Masrori, P., Damme, P. V. & Bosch, L. V. D. The sense of antisense therapies in ALS. Trends Mol. Med. 30, 252–262 (2024).
Ke, H. et al. Loss of TDP43 inhibits the progression of triple-negative breast cancer in coordination with SRSF3. Proc. Natl Acad. Sci. 115, E3426–E3435 (2018).
Corbo, C., Orrù, S. & Salvatore, F. SRp20: an overview of its role in human diseases. Biochem. Biophys. Res. Commun. 436, 1–5 (2013).
Wang, Z. et al. Iron drives T helper cell pathogenicity by promoting RNA-binding protein PCBP1-mediated proinflammatory cytokine production. Immunity 49, 80–92.e7 (2018).
Eberhard, Y. et al. Chelation of intracellular iron with the antifungal agent ciclopirox olamine induces cell death in leukemia and myeloma cells. Blood 114, 3064–3073 (2009).
Zhou, L., Jiang, Y., Luo, Q., Li, L. & Jia, L. Neddylation: a novel modulator of the tumor microenvironment. Mol. Cancer 18, 77 (2019).
Szklarczyk, D. et al. The STRING database in 2021: customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res 49, gkaa1074 (2020).
Zacco, E. et al. Probing TDP-43 condensation using an in silico designed aptamer. Nat. Commun. 13, 3306 (2022).
Waldron, F. M. et al. Improved detection of pre-symptomatic, non-central nervous system TDP-43 pathology in amyotrophic lateral sclerosis using RNA aptamer. bioRxiv, https://doi.org/10.1101/2025.04.10.648122 (2025).
Thiviyanathan, V. & Gorenstein, D. G. Aptamers and the next generation of diagnostic reagents. Proteom. Clin. Appl. 6, 563–573 (2012).
Abrams, B. B., Hänel, H. & Hoehler, T. Ciclopirox olamine: a hydroxypyridone antifungal agent. Clin. Dermatol. 9, 471–477 (1991).
Sonthalia, S., Agrawal, M. & Sehgal, V. N. Topical ciclopirox olamine 1%: revisiting a unique antifungal. Indian Dermatol. Online J. 10, 481–485 (2019).
Gupta, A. K. Ciclopirox: an overview. Int. J. Dermatol 40, 305–310 (2001).
Minden, M. D. et al. Oral ciclopirox olamine displays biological activity in a phase I study in patients with advanced hematologic malignancies. Am. J. Hematol. 89, 363–368 (2014).
Qi, J. et al. Ciclopirox activates PERK-dependent endoplasmic reticulum stress to drive cell death in colorectal cancer. Cell Death Dis. 11, 582 (2020).
Su, Z. et al. Ciclopirox and bortezomib synergistically inhibits glioblastoma multiforme growth via simultaneously enhancing JNK/p38 MAPK and NF-κB signaling. Cell Death Dis. 12, 251 (2021).
Kang, J.-A. et al. Ciclopirox inhibits Hepatitis B Virus secretion by blocking capsid assembly. Nat. Commun. 10, 2184 (2019).
Yap, K., Lim, Z. Q., Khandelia, P., Friedman, B. & Makeyev, E. V. Coordinated regulation of neuronal mRNA steady-state levels through developmentally controlled intron retention. Genes Dev. 26, 1209–1223 (2012).
Zhao, M., Kim, J. R., Van Bruggen, R. & Park, J. RNA-binding proteins in amyotrophic lateral sclerosis. Mol. Cells 41, 818–829 (2018).
Coelho, M. B. et al. Nuclear matrix protein Matrin3 regulates alternative splicing and forms overlapping regulatory networks with PTB. EMBO J. 34, 653–668 (2015).
Kato, Y. & Sakamoto, K. Niclosamide affects intracellular TDP-43 distribution in motor neurons, activates mitophagy, and attenuates morphological changes under stress. J. Biosci. Bioeng. 132, 640–650 (2021).
Hans, F., Glasebach, H. & Kahle, P. J. Multiple distinct pathways lead to hyperubiquitylated insoluble TDP-43 protein independent of its translocation into stress granules. J. Biol. Chem. 295, 673–689 (2019).
Cohen, T. J., Hwang, A. W., Unger, T., Trojanowski, J. Q. & Lee, V. M. Redox signalling directly regulates TDP-43 via cysteine oxidation and disulphide cross-linking. EMBO J. 31, 1241–1252 (2012).
Zuo, X. et al. TDP-43 aggregation induced by oxidative stress causes global mitochondrial imbalance in ALS. Nat. Struct. Mol. Biol. 28, 132–142 (2021).
Gautam, M. et al. Mitochondria, ER, and nuclear membrane defects reveal early mechanisms for upper motor neuron vulnerability with respect to TDP-43 pathology. Acta Neuropathol. 137, 47–69 (2019).
Chang, H.-Y., Hou, S.-C., Way, T.-D., Wong, C.-H. & Wang, I.-F. Heat-shock protein dysregulation is associated with functional and pathological TDP-43 aggregation. Nat. Commun. 4, 2757 (2013).
Bhattarai, A. et al. Serial assessment of iron in the motor cortex in limb-onset amyotrophic lateral sclerosis using quantitative susceptibility mapping. Quant. Imaging Med. Surg. 10, 1465–1476 (2020).
Kwan, J. Y. et al. Iron accumulation in deep cortical layers accounts for MRI signal abnormalities in ALS: correlating 7 Tesla MRI and pathology. PLoS ONE 7, e35241 (2012).
Gao, J., Okolo, O., Siedlak, S. L., Friedland, R. P. & Wang, X. Ferritin is closely associated with microglia in amyotrophic lateral sclerosis. J. Neuropathol. Exp. Neurol. nlae074 https://doi.org/10.1093/jnen/nlae074 (2024).
Ghaderi, S., Fatehi, F., Kalra, S., Mohammadi, S. & Batouli, S. A. H. Quantitative susceptibility mapping in amyotrophic lateral sclerosis: automatic quantification of the magnetic susceptibility in the subcortical nuclei. Amyotroph. Lateral Scler. Front. Degener. ahead-of-print, 1–12 (2024).
Ravanfar, P. et al. Systematic review: Quantitative Susceptibility Mapping (QSM) of brain iron profile in neurodegenerative diseases. Front. Neurosci. 15, 618435 (2021).
Yin, J. et al. Ciclopirox olamine exerts tumor-suppressor effects via topoisomerase II alpha in lung adenocarcinoma. Front. Oncol. 12, 791916 (2022).
Belenky, P., Camacho, D. & Collins, J. J. Fungicidal drugs induce a common oxidative-damage cellular death pathway. Cell Rep. 3, 350–358 (2013).
Tang, S. et al. The role of iron, its metabolism and ferroptosis in traumatic brain injury. Front. Cell. Neurosci. 14, 590789 (2020).
Costa, I. et al. Molecular mechanisms of ferroptosis and their involvement in brain diseases. Pharmacol. Ther. 244, 108373 (2023).
Sanghai, N. & Tranmer, G. K. Biochemical and molecular pathways in neurodegenerative diseases: an integrated view. Cells 12, 2318 (2023).
Sun, Y. et al. Mechanisms of ferroptosis and emerging links to the pathology of neurodegenerative diseases. Front. Aging Neurosci. 14, 904152 (2022).
Oba, H. et al. Amyotrophic lateral sclerosis: T2 shortening in motor cortex at MR imaging. Radiology 189, 843–846 (1993).
Imon, Y. et al. Low intensity areas observed on T2-weighted magnetic resonance imaging of the cerebral cortex in various neurological diseases. J. Neurol. Sci. 134, 27–32 (1995).
Berg, D., Becker, G., Riederer, P. & Rieß, O. Iron in neurodegenerative disorders. Neurotox. Res. 4, 637–653 (2002).
Goodall, E. F., Haque, M. S. & Morrison, K. E. Increased serum ferritin levels in amyotrophic lateral sclerosis (ALS) patients. J. Neurol. 255, 1652–1656 (2008).
Veyrat-Durebex, C. et al. Iron metabolism disturbance in a french cohort of ALS patients. BioMed. Res. Int. 2014, 485723 (2014).
Zheng, Y., Gao, L., Wang, D. & Zang, D. Elevated levels of ferritin in the cerebrospinal fluid of amyotrophic lateral sclerosis patients. Acta Neurol. Scand. 136, 145–150 (2017).
Mitani, T. T. et al. Amyotrophic lateral sclerosis with speech apraxia, predominant upper motor neuron signs, and prominent iron accumulation in the frontal operculum and precentral gyrus. Neuropathology 41, 324–331 (2021).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Kent, W. J. et al. The human genome browser at UCSC. Genome Res 12, 996–1006 (2002).
Consortium, I. H. G. S. et al. Initial sequencing and analysis of the human genome. Nature 409, 860–921 (2001).
Raney, B. J. et al. The UCSC Genome Browser database: 2024 update. Nucleic Acids Res 52, D1082–D1088 (2023).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Wickham, H. et al. Welcome to the Tidyverse. J. Open Source Softw. 4, 1686 (2019).
Wickham, H. ggplot2, Elegant Graphics for Data Analysis. Use R! 89–107 https://doi.org/10.1007/978-3-319-24277-4_5 (2016).
Tian, R. et al. CRISPR interference-based platform for multimodal genetic screens in human iPSC-derived neurons. Neuron 104, 239–255.e12 (2019).
Wang, C. et al. Scalable production of iPSC-derived human neurons to identify tau-lowering compounds by high-content screening. Stem Cell Rep. 9, 1221–1233 (2017).
Korbie, D. J. & Mattick, J. S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 3, 1452–1456 (2008).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Harenza, J. L. et al. Transcriptomic profiling of 39 commonly-used neuroblastoma cell lines. Sci. Data 4, 170033 (2017).
Song, X. et al. SRSF3-regulated RNA alternative splicing promotes glioblastoma tumorigenicity by affecting multiple cellular processes. Cancer Res 79, 5288–5301 (2019).
Karginov, T. A., Ménoret, A. & Vella, A. T. Optimal CD8+ T cell effector function requires costimulation-induced RNA-binding proteins that reprogram the transcript isoform landscape. Nat. Commun. 13, 3540 (2022).
Abbasinejad et al. SnapMine. Zenodo https://doi.org/10.5281/zenodo.15624203 (2025).
Acknowledgments
This work was supported by the Alzheimer’s Association (to J.P.L.), the Institute for Data-Intensive Engineering and Science (to J.P.L.), the NIH (R01NS095969, UH3NS115608 and R33NS115161 to P.C.W.; R01NS127186 to J.M.G.), the Robert Packard Center for ALS Research at Johns Hopkins (to P.C.W.), the Target ALS Foundation (to P.C.W. and J.M.G.), ALS Finding a Cure (to P.C.W.), the ALS Association (to P.C.W.), the US Food and Drug Administration (no. 1U01FD008129 to P.C.W. and J.D.B.), and the JXTX + CSHL Biological Data Science 2024 Scholarship (to I.R.S.). This material is based upon work supported by the National Science Foundation Graduate Research Fellowship Program under Grant No. DGE2139757 (to I.R.S.). Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation. In part, computational work was carried out at the Advanced Research Computing at Hopkins (ARCH) core facility (rockfish.jhu.edu), which is supported by the NSF (no. OAC 1920103). Funding for SSEC comes from Schmidt Sciences. We thank Dr. Ashani Weeraratna for generously providing fibroblast line 2003-071-056. We thank the Viral Immune Pathogenesis Center at Johns Hopkins University for human PBMCs used in preliminary testing. We thank Jeffrey J. Li, Abigail E. Bush, and Anya A. Kim for technical support.
Author information
Authors and Affiliations
Contributions
I.R.S., P.C.W., and J.P.L. conceptualized the study. I.R.S. and J.P.L. wrote the manuscript, and all other authors edited and approved it. I.R.S., P.S.S., H.S., G.D.B., A.P.M., K.E.I., A.F.C., V.W., S.R.M., J.L.R., and J.M.G., performed experiments and/or analyzed data. F.A. and I.R.S. designed and maintain the application. I.R.S., B.L., and J.P.L. developed application methodology.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Jack Humphrey, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sinha, I.R., Sandal, P.S., Spence, H. et al. Large-scale RNA-Seq mining reveals ciclopirox olamine induces TDP-43 cryptic exons. Nat Commun 16, 6878 (2025). https://doi.org/10.1038/s41467-025-62004-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-62004-5
