
Abstract
Postmitotic neurons have high levels of methylated cytosine and its oxidized intermediates such as 5-hydroxymethylcytosine1. However, the functional relevance of these epigenetic modifications of DNA are poorly understood. Here we show that some cytidine analogues, such as cytarabine, cause DNA double-strand breaks during TET-mediated active 5-methylcytosine demethylation by interrupting TDG-dependent base excision repair. These double-strand breaks are frequently converted into deletions and translocations by DNA ligase 4. In vivo, Purkinje and Golgi cells in the cerebellum are the only neuronal populations that exhibit high levels of DNA damage due to cytarabine. In Purkinje cells, TET targets highly expressed gene bodies marked by enhancer-associated histone modifications. Many of these genes control movement coordination, which explains the long-recognized cerebellar neurotoxicity of cytarabine2. We show that other cytidine analogues, such as gemcitabine, cause only single-strand breaks in neurons, which are repaired by DNA ligase 3 with minimal toxicity. Our findings uncover a mechanistic link between TET-mediated DNA demethylation, base excision repair and gene expression in neurons. The results also provide a rational explanation for the different neurotoxicity profiles of an important class of antineoplastic agents.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
The CRISPRi screen, END-seq, S1-END-seq, RNA-seq, LAM-HTGTS, ATAC–seq, SAR-seq, targeted deep sequencing (amplicon-seq), PS-seq and CUT&RUN data have been deposited into the GEO database under accession number GSE293256. The following data were obtained from publicly available sources: single-nucleus RNA-seq in SCA1 cerebellar tissues (GSE246183); BS-seq and oxBS-seq in Purkinje cells (GSE166423); ATAC–seq in Purkinje and granule cells (GSE174842); ChIP–seq of H3K4me1 in primary cortical neurons (GSE164265); ChIP–seq of H3K4me1 (GSE167259); and dideoxycytosine S1-END-seq and dideoxycytosine END-seq in i3Neurons (GSE210317). The hg19 (human) and mm10 (mouse) genome assemblies were used as references for alignment. Source data are provided with this paper.
References
Kriaucionis, S. & Heintz, N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 324, 929–930 (2009).
Stone, J. B. & DeAngelis, L. M. Cancer-treatment-induced neurotoxicity—focus on newer treatments. Nat. Rev. Clin. Oncol. 13, 92–105 (2016).
Berdis, A. J. Inhibiting DNA polymerases as a therapeutic intervention against cancer. Front. Mol. Biosci. 4, 78 (2017).
Fram, R. J. & Kufe, D. W. DNA strand breaks caused by inhibitors of DNA synthesis: 1-β-d-arabinofuranosylcytosine and aphidicolin. Cancer Res. 42, 4050–4053 (1982).
Huang, P., Chubb, S., Hertel, L. W., Grindey, G. B. & Plunkett, W. Action of 2′,2′-difluorodeoxycytidine on DNA synthesis. Cancer Res. 51, 6110–6117 (1991).
Longley, D. B., Harkin, D. P. & Johnston, P. G. 5-Fluorouracil: mechanisms of action and clinical strategies. Nat. Rev. Cancer 3, 330–338 (2003).
Heinemann, V. et al. Inhibition of ribonucleotide reduction in CCRF-CEM cells by 2′,2′-difluorodeoxycytidine. Mol. Pharmacol. 38, 567–572 (1990).
Gibson, E. M. et al. Methotrexate chemotherapy induces persistent tri-glial dysregulation that underlies chemotherapy-related cognitive impairment. Cell 176, 43–55 (2019).
Jarmolowicz, D. P. et al. 5-Fluorouracil impairs attention and dopamine release in rats. Behav. Brain Res. 362, 319–322 (2019).
Herzig, R. H. et al. Cerebellar toxicity with high-dose cytosine arabinoside. J. Clin. Oncol. 5, 927–932 (1987).
Vaughn, D. J., Jarvik, J. G., Hackney, D., Peters, S. & Stadtmauer, E. A. High-dose cytarabine neurotoxicity: MR findings during the acute phase. Am. J. Neuroradiol. 14, 1014–1016 (1993).
Magge, R. S. & DeAngelis, L. M. The double-edged sword: neurotoxicity of chemotherapy. Blood Rev. 29, 93–100 (2015).
Wang, D. et al. Active DNA demethylation promotes cell fate specification and the DNA damage response. Science 378, 983–989 (2022).
Hussmann, J. A. et al. Mapping the genetic landscape of DNA double-strand break repair. Cell 184, 5653–5669 (2021).
Fernandopulle, M. S. et al. Transcription factor-mediated differentiation of human iPSCs into neurons. Curr. Protoc. Cell Biol. 79, e51 (2018).
Wang, B. et al. Integrative analysis of pooled CRISPR genetic screens using MAGeCKFlute. Nat. Protoc. 14, 756–780 (2019).
Georges, R. O. et al. Acute deletion of TET enzymes results in aneuploidy in mouse embryonic stem cells through decreased expression of Khdc3. Nat. Commun. 13, 6230 (2022).
Ladstatter, S. & Tachibana-Konwalski, K. A surveillance mechanism ensures repair of DNA lesions during zygotic reprogramming. Cell 167, 1774–1787 (2016).
Enokido, Y., Araki, T., Aizawa, S. & Hatanaka, H. p53 involves cytosine arabinoside-induced apoptosis in cultured cerebellar granule neurons. Neurosci. Lett. 203, 1–4 (1996).
Nabet, B. et al. The dTAG system for immediate and target-specific protein degradation. Nat. Chem. Biol. 14, 431–441 (2018).
Yap, E. L. & Greenberg, M. E. Activity-regulated transcription: bridging the gap between neural activity and behavior. Neuron 100, 330–348 (2018).
Dileep, V. et al. Neuronal DNA double-strand breaks lead to genome structural variations and 3D genome disruption in neurodegeneration. Cell 186, 4404–4421 (2023).
Madabhushi, R. et al. Activity-induced DNA breaks govern the expression of neuronal early-response genes. Cell 161, 1592–1605 (2015).
Ramsden, D. A. & Nussenzweig, A. Mechanisms driving chromosomal translocations: lost in time and space. Oncogene 40, 4263–4270 (2021).
Hu, J. et al. Detecting DNA double-stranded breaks in mammalian genomes by linear amplification-mediated high-throughput genome-wide translocation sequencing. Nat. Protoc. 11, 853–871 (2016).
Canela, A. et al. DNA breaks and end resection measured genome-wide by end sequencing. Mol. Cell 63, 898–911 (2016).
Wu, W. et al. Neuronal enhancers are hotspots for DNA single-strand break repair. Nature 593, 440–444 (2021).
Mateos-Gomez, P. A. et al. Mammalian polymerase θ promotes alternative NHEJ and suppresses recombination. Nature 518, 254–257 (2015).
Ceccaldi, R. et al. Homologous-recombination-deficient tumours are dependent on Polθ-mediated repair. Nature 518, 258–262 (2015).
Alexandrov, L. B. et al. The repertoire of mutational signatures in human cancer. Nature 578, 94–101 (2020).
Morton, L. M. et al. Radiation-related genomic profile of papillary thyroid carcinoma after the Chernobyl accident. Science 372, eabg2538 (2021).
Lowenberg, B. et al. Cytarabine dose for acute myeloid leukemia. N. Engl. J. Med. 364, 1027–1036 (2011).
Pinero, J. et al. DisGeNET: a discovery platform for the dynamical exploration of human diseases and their genes. Database 2015, bav028 (2015).
Tejwani, L. et al. Longitudinal single-cell transcriptional dynamics throughout neurodegeneration in SCA1. Neuron 112, 362–383 (2024).
Tan, Y. et al. Signal-induced enhancer activation requires Ku70 to read topoisomerase1–DNA covalent complexes. Nat. Struct. Mol. Biol. 30, 148–158 (2023).
Kwak, Y. D. et al. Chromatin architecture at susceptible gene loci in cerebellar Purkinje cells characterizes DNA damage-induced neurodegeneration. Sci. Adv. 7, eabg6363 (2021).
Stoyanova, E., Riad, M., Rao, A. & Heintz, N. 5-Hydroxymethylcytosine-mediated active demethylation is required for mammalian neuronal differentiation and function. eLife 10, e66973 (2021).
Onodera, A. et al. Roles of TET and TDG in DNA demethylation in proliferating and non-proliferating immune cells. Genome Biol. 22, 186 (2021).
Rahmani, H., Radmehr, M., Hadjibabaie, M. & Solduzian, M. Acute cerebellar toxicity induced by high dose of cytarabine (HiDAC): a case report. J. Oncol. Pharm. Pract. 26, 1492–1494 (2020).
Weber, A. R. et al. Biochemical reconstitution of TET1–TDG–BER-dependent active DNA demethylation reveals a highly coordinated mechanism. Nat. Commun. 7, 10806 (2016).
Prakasha Gowda, A. S., Polizzi, J. M., Eckert, K. A. & Spratt, T. E. Incorporation of gemcitabine and cytarabine into DNA by DNA polymerase β and ligase III/XRCC1. Biochemistry 49, 4833–4840 (2010).
Drenberg, C. D. et al. A high-throughput screen indicates gemcitabine and JAK inhibitors may be useful for treating pediatric AML. Nat. Commun. 10, 2189 (2019).
Luquette, L. J. et al. Single-cell genome sequencing of human neurons identifies somatic point mutation and indel enrichment in regulatory elements. Nat. Genet. 54, 1564–1571 (2022).
Ganz, J. et al. Contrasting somatic mutation patterns in aging human neurons and oligodendrocytes. Cell 187, 1955–1970 (2024).
Shiloh, Y. The cerebellar degeneration in ataxia-telangiectasia: a case for genome instability. DNA Repair 95, 102950 (2020).
Kim, J. et al. Prevalence and mechanisms of somatic deletions in single human neurons during normal aging and in DNA repair disorders. Nat. Commun. 13, 5918 (2022).
Caldecott, K. W. DNA single-strand break repair and human genetic disease. Trends Cell Biol. 32, 733–745 (2022).
Gazestani, V. et al. Early Alzheimer’s disease pathology in human cortex involves transient cell states. Cell 186, 4438–4453 (2023).
Guo, J. U. et al. Neuronal activity modifies the DNA methylation landscape in the adult brain. Nat. Neurosci. 14, 1345–1351 (2011).
Yu, H. et al. Tet3 regulates synaptic transmission and homeostatic plasticity via DNA oxidation and repair. Nat. Neurosci. 18, 836–843 (2015).
Tian, R. et al. CRISPR interference-based platform for multimodal genetic screens in human iPSC-derived neurons. Neuron 104, 239–255 (2019).
Xia, Z., Dudek, H., Miranti, C. K. & Greenberg, M. E. Calcium influx via the NMDA receptor induces immediate early gene transcription by a MAP kinase/ERK-dependent mechanism. J. Neurosci. 16, 5425–5436 (1996).
Li, W. et al. MAGeCK enables robust identification of essential genes from genome-scale CRISPR/Cas9 knockout screens. Genome Biol. 15, 554 (2014).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Shinoda, K. et al. The dystonia gene THAP1 controls DNA double-strand break repair choice. Mol. Cell 81, 2611–2624 (2021).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Langmead, B., Trapnell, C., Pop, M. & Salzberg, S. L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, R25 (2009).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Kent, W. J. et al. The human genome browser at UCSC. Genome Res. 12, 996–1006 (2002).
Kent, W. J., Zweig, A. S., Barber, G., Hinrichs, A. S. & Karolchik, D. BigWig and BigBed: enabling browsing of large distributed datasets. Bioinformatics 26, 2204–2207 (2010).
Ramirez, F. et al. deepTools2: a next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 44, W160–W165 (2016).
Robinson, J. T. et al. Integrative genomics viewer. Nat. Biotechnol. 29, 24–26 (2011).
Xi, Y. & Li, W. BSMAP: whole genome bisulfite sequence MAPping program. BMC Bioinformatics 10, 232 (2009).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Danecek, P. et al. Twelve years of SAMtools and BCFtools. Gigascience 10, giab008 (2021).
Liao, Y., Smyth, G. K. & Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 47, e47 (2019).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation 2, 100141 (2021).
Yu, G., Wang, L. G., Yan, G. R. & He, Q. Y. DOSE: an R/Bioconductor package for disease ontology semantic and enrichment analysis. Bioinformatics 31, 608–609 (2015).
Newman, A. M. et al. Determining cell type abundance and expression from bulk tissues with digital cytometry. Nat. Biotechnol. 37, 773–782 (2019).
Yao, Z. et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature 624, 317–332 (2023).
Barnett, K. R. et al. ATAC-Me captures prolonged DNA methylation of dynamic chromatin accessibility loci during cell fate transitions. Mol. Cell 77, 1350–1364 (2020).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. Preprint at https://doi.org/10.48550/arXiv.1303.3997 (2013).
Bergstrom, E. N. et al. SigProfilerMatrixGenerator: a tool for visualizing and exploring patterns of small mutational events. BMC Genomics 20, 685 (2019).
Vega-Sendino, M. et al. The homeobox transcription factor DUXBL controls exit from totipotency. Nat. Genet. 56, 697–709 (2024).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Hammel, M. et al. An intrinsically disordered APLF links Ku, DNA-PKcs, and XRCC4-DNA ligase IV in an extended flexible non-homologous end joining complex. J. Biol. Chem. 291, 26987–27006 (2016).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2024).
Song, C. X. et al. Genome-wide profiling of 5-formylcytosine reveals its roles in epigenetic priming. Cell 153, 678–691 (2013).
Burger, L., Gaidatzis, D., Schubeler, D. & Stadler, M. B. Identification of active regulatory regions from DNA methylation data. Nucleic Acids Res. 41, e155 (2013).
Gu, T. et al. The disordered N-terminal domain of DNMT3A recognizes H2AK119ub and is required for postnatal development. Nat. Genet. 54, 625–636 (2022).
Acknowledgements
We thank L. Boxer and R. Sakr for help with the isolation of primary mouse cortical neurons; B. Adamson for the AX227 sgRNA library; D. Zong for comments on the manuscript; A. Vogt for purifying the XRCC4–LIG4 complex; M. Greenberg for discussions; staff at the National Cancer Institute/Center for Cancer Research Genomics Core for help with sequencing; staff at the Laboratory Animal Science Program for mouse work; and staff at the Center for Cancer Research/Laboratory of Genome Integrity Flow Cytometry Core for cell sorting. The computational resources of the NIH High-Performance Computing Biowulf cluster were used for data analyses. This work was supported by the Intramural Research Program of the NIH (National Cancer Institute contract HHSN2612015000031 to A.N., F.L. and P.D.A.), an Ellison Medical Foundation Senior Scholar in Aging Award (AG-SS-2633-11 to A.N.), Department of Defense Awards (W81XWH-16-1-599 and W81XWH-19-10652 to A.N.), an International Partnership Program of Chinese Academy of Sciences grant (318GJHZ2023004MI to W.W.), a National Natural Science Foundation of China grant (32370575 to W.W.), the Shanghai Rising-Star Program (24QA2710100 to W.W.) and NIH grants (NS-37956 and NS-123081 to P.J.M., and R01GM144559 and P01CA092584 to Y.H.).
Author information
Authors and Affiliations
Contributions
J.-C.L., D.W., E.C., F.L. and A.N. conceived, designed and analysed the experiments. C.C., W.W. and Y. Song designed the bioinformatic pipelines and performed data analyses. J.-C.L., C.C. and W.W. designed the figures. J.-C.L. and E.C. prepared the i3Neurons, performed CRISPRi screens, gene knockdown and survival experiments. M.E.W. helped with the CRISPRi screen. J.-C.L., E.C. and S.N. performed immunofluorescence imaging. J.-C.L. performed CRISPR knockout, END-seq, S1-END-seq, ATAC–seq, PS-seq, targeted deep sequencing, CUT&RUN and RT–qPCR. J.-C.L. and D.W. prepared cortical neurons and performed RNA-seq and SAR-seq experiments. J.-C.L., D.W., F.L. and E.C. performed FACS. Y. Shang performed LAM-HTGTS and analysed the data, supervised by F.-L.M. G.N.R. generated the virus-like particles used for LAM-HTGTS. R.C. performed cloning experiments for TDGdegron. D.S. performed in vitro reconstitution of BER and analyses, supervised by P.S. Y.H. provided purified the XRCC4–LIG4 complex. N.W. and E.C. performed genotyping of mutant mice. N.W. performed western blotting. Y.Z. assisted in intraperitoneal injections. N.H. and A.R. provided gene knockout mice. P.D.A., N.H., P.J.M. and K.W.C. provided critical insights. J.-C.L., F.L. and A.N. wrote the paper with input from all co-authors. A.N. supervised the study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks the anonymous reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 CRISPR interference screen identifies genes involved in AraC-induced neurotoxicity.
a, Pairwise sample correlations of triplicates collected from different time points (day 0, day 3, and day 9) in iN either treated or not with AraC. The correlation is displayed by a pie graph on the upper right and values are indicated on the bottom left. b, Scatter plot showing the beta score of individual 476 genes in the CRISPRi screen. Top 10 “resistant” genes (red) and top 10 “susceptible” genes (blue) are highlighted. c,d, Interaction network of positive (c) and negative (d) hits with the enriched pathways indicated and color coded. e, Knockdown efficiency of individual sgRNA. Y-axis represents the expression of DNMT1/TDG/UHRF1 in knockdown iN (sgDNMT1/sgTDG/sgUHRF1) relative to that in wild-type iN transfected with a control vector (WT). Data are presented as mean ± SD, n = 3 biologically independent experiments. Statistical significance was determined using two-sided paired t test. f, Western blot analysis of DNMT1/TDG protein levels in wild-type (WT) and DNMT1/TDG knockdown (sgDNMT1/sgTDG) iN. For the gel source data, see Supplementary Fig. 1. g, Images of γ-H2AX foci in wild-type, and TDG/DNMT1/UHRF1 knockdown iN either treated or not with AraC (40 μM) on day 6. 4′,6-diamidino-2-phenylindole (DAPI) (blue); γ-H2AX (red). Scale bar, 20 μm. h, Images of NeuN and γ-H2AX in wild-type, Tdg−/− primary cortical neurons treated with AraC (40 μM) on day 6. 4′,6-diamidino-2-phenylindole (DAPI) (blue); NeuN (green); γ-H2AX (red). 4-Hydroxytamoxifen (4-OHT) was added in vitro to knockout TDG in primary cortical neurons isolated from CreERT2 Tdgfl/fl embryos. Scale bar, 20 μm. i,j, Expression of Tet1/2/3 in wild-type cortical neuron (i) and Purkinje neuron (j). Y-axis represents the expression level measured by RNA-seq. Data are presented as mean ± SD. n = 3 biologically independent samples (cortical neuron), n = 6 mice (Purkinje neuron). Statistical significance was determined using the two-sided paired t test, p value is indicated. k, Images of NeuN and γ-H2AX in wild-type and Tet−/− primary cortical neurons treated with AraC (40 μM) on day 6. 4′,6-diamidino-2-phenylindole (DAPI) (blue); NeuN (green); γ-H2AX (red). 4-Hydroxytamoxifen (4-OHT) was added in vitro to knockout TET1/2/3 in primary cortical neurons isolated from CreERT2 Tet1/2/3fl/fl embryos. Scale bar, 20 μm. l, Survival of wild-type and Tet−/− primary cortical neurons one day after AraC treatment. Y-axis represents viability relative to non-treated cells. Data are presented as mean ± SD, n = 4 biologically independent samples. Statistical significance was determined using the two-sided paired t test, p value is indicated. m, Survival of iPSC with indicated treatments. Y-axis represents viability relative to non-treated cells, X-axis represents days after treatment. Data are presented as mean ± SD, n = 3 biologically independent samples. n, Heatmap of expression of p53-pathway genes in wild-type iN with indicated treatments.
Extended Data Fig. 2 AraC induces double-strand breaks that are TET- and TDG-dependent.
a, Structure of Deoxycytidine (dC), Dideoxycytidine (ddC), Cytarabine (AraC), Gemcitabine (dFdC) and Adenine arabinoside (AraA). b, Representative images of iN with immunofluorescence staining for DAPI (blue) and PARylation (green) in iN treated or not with AraC. Scale bar, 10 μm. c, Heatmaps of AraC-S1-END-seq in TDGdegron iN treated or not with dTAG. Similar results were observed in at least 2 independent experiments. d, Schematic diagram of pyridine borane sequencing (PS-seq). 5-Formylcytosine (5fC) and 5-carboxycytosine (5caC) are converted to dihydrouracil (DHU) by pyridine borane and read as T after PyroMark PCR amplification. e, Bar graph depicting the percentage of (5fC + 5caC)/total C in 25 CpGs at four targeted AraC-induced DSB sites in TDGdegron iN treated overnight or not with dTAG as measured by PS-seq. f, Heatmap of AraC-END-seq in wild-type iN treated or not with DMOG (1 mM). g, Composite DNA sequence motif analysis 4 bp on either side of AraC-END-seq peak summits in iN. Peak fraction and p value are indicated. Statistical significance was determined using the two-sided binomal test.
Extended Data Fig. 3 AraC induces expression of immediate early genes and late response genes in cortical neurons.
a, Genome browser screenshot showing a double-strand break (END-seq) induced by AraC (40 μM) that colocalizes with the enhancer mark H3K4me1 (non-treated) in primary cortical neurons. b, Heatmaps of END-seq (DSB) in non-treated and AraC-treated primary cortical neurons, and ChIP-seq of H3K4me1 in non-treated primary cortical neurons. The peaks are ordered by END-seq intensity and plotted 1 kb on either side of DSB summits. c, Composite DNA sequence motif analysis 4 bp on either side of AraC-END-seq peak summits in primary cortical neurons. Peak fraction and p value are indicated. Statistical significance was determined using the two-sided binomal test. d, Motif analysis by HOMER. Top, the Fos binding motif downloaded from JASPAR motif database. Bottom, the most enriched motif identified from sequences within 200 bp of the AraC-END-seq peak summits in primary cortical neurons. Peak fraction and p value are indicated. Statistical significance was determined using the two-sided binomal test. e, Aggregate plot showing the Fos motif distribution at AraC-END-seq peak summits in primary cortical neurons 1 kb on either side of AraC-END-seq peak summits. f, Volcano plot showing differentially expressed genes upon AraC treatment in primary cortical neurons. Genes whose expression were upregulated and downregulated significantly (log2fold change>1 and p value < 0.01) are indicated in red and blue respectively. The 10 most significantly upregulated genes are indicated. Statistical significance was determined using the two-sided Wald test and adjusted by the Benjamini & Hochberg method. g, Heatmap of immediate early genes (IEGs) and late response genes (LRGs) expression in non-treated and AraC-treated primary cortical neurons. h, Top 15 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways identified from upregulated genes upon AraC treatment in primary cortical neurons. Statistical significance was determined using the one-sided hypergeometric test. i, qRT-PCR measurement of Fos, Fosb, Npas4, Junb, Bdnf and Vgf gene expression in non-treated or AraC/dFdC (40 μM for 16 hr) treated primary cortical neurons. Statistical significance was determined using the two-sided paired t test, p value is indicated. Data are presented as mean ± SD, n = 3 biologically independent samples. j, qRT-PCR measurement of Fos and Npas4 gene expression in non-treated or etoposide (ETO, 50 μM for 16 hr) treated primary cortical neurons. Statistical significance was determined using two-sided paired t test, p value is indicated. Data are presented as mean ± SD, n = 3 biologically independent samples. k, qRT-PCR measurement of Fos, Fosb, Npas4, Junb, Bdnf and Vgf gene expression in wild-type or Tdg−/− primary cortical neurons treated with AraC (40 μM for 16 hr). 4-Hydroxytamoxifen (4-OHT) was added in vitro to deplete TDG in primary cortical neurons isolated from CreERT2 Tdgfl/fl embryos. Statistical significance was determined using two-sided paired t test, p value is indicated. Data are presented as mean ± SD, n = 3 biologically independent samples.
Extended Data Fig. 4 DSBs induced by AraC leads to chromosome translocations and indels.
a, Schematic overview of LAM-HTGTS. b, Aggregate plot of B2M-captured HTGTS signals in wild-type iN treated with (B2M+AraC) or without (B2M) AraC relative to AraC-END-seq summits. Two of the three replicates are plotted, with the remaining shown in Fig. 2g. c, Enhancers (H3K4me1) colocalizing with DNA repair synthesis hotspots (SAR-seq), AraC-induced DSBs (AraC-END-seq) and translocations (AraC-HTGTS) at a prey gene MEX3A and the bait gene NHLH1 on chromosome 1. The translocation hotspot and HTGTS bait are highlighted by orange and red dashed box respectively. d, Schematic overview of targeted deep sequencing. e, Length distribution of deletions induced by AraC. The minimal (1 bp), maximum (89 bp), median (26 bp) and mean (28 bp) deletion lengths are indicated. f, Expression of POLQ in iPSC, day7-iN and day17-iN. Y-axis represent the expression level measured by RNA-seq. Data are presented as the mean, n = 2 biologically independent samples. g, Western blot analysis of LIG4 protein levels in wild-type and lig4 knockout iN. Two independent lig4 knockout clones c13 and c40 were tested. For the gel source data, see Supplementary Fig. 1. h,i, Top, Genome browser screenshot of DSBs induced by AraC on chromosome 1 (h) and 6 (i). Bottom, the deletion profile of the above DSB in wild-type, lig4 knockouts (c13 and c40) and DNA-PKcs inhibitor (NU7441, 5 μM) treated wild-type iN. Y-axis represents the deletion frequency; X-axis represents the chromosome location.
Extended Data Fig. 5 AraC generates DSBs in Golgi and Purkinje cells in the cerebellum.
a, Flow cytometric analysis of nuclei isolated from 1 month old mouse cerebellum after PBS- or AraC treatment. Y- and X-axes indicate the expression of the Purkinje specific marker ITPR1 and the DNA damage marker γ-H2AX respectively. Purkinje cells and an unknown population are marked by the red and green box respectively. For the flow cytometry gating strategy, see Supplementary Fig. 2. b, Cell type composition of sorted Purkinje cells and the unknown population shown in (a) determined by RNA-seq. The estimated fraction is determined using single-cell sequencing data from mouse brain. Nuclei were isolated 3 h after AraC treatment. c,d, Top 10 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways identified from AraC-induced upregulated (c) and downregulated (d) genes in Purkinje cells. Statistical significance was determined using the one-sided hypergeometric test. e, Fos expression in Purkinje cells isolated from wild type mice receiving PBS or AraC. Statistical significance was determined using the two-sided Wald test and adjusted by the Benjamini & Hochberg method, p value = 0.03. Data are presented as mean ± SEM, six and five mice were used for analysis in the PBS and AraC groups, respectively. f, Flow cytometric analysis of nuclei isolated from the whole brain without cerebellum from PBS- or AraC-treated mice. Y- and X-axes represent the expression of the DNA damage marker γ-H2AX and the neuronal marker NeuN respectively. The NeuN and γ-H2AX positive population is highlighted in the red square. For the flow cytometry gating strategy, see Supplementary Fig. 2. g, Cell type composition of sorted γ-H2AX positive population upon AraC treatment shown in (f) determined by RNA-seq. The estimated fraction is determined using single-cell sequencing data from mouse brain. h, Bar graph depicting the percentage of (5fC + 5caC)/total C in 55 CpGs at seven sites in genes Grid2 (six sites) and Itpr1 (one site) in Purkinje nuclei isolated from wild type or Pcp2-Cre TDGfl/fl mice (Tdg−/−) as measured by PS-seq. i, Integrative Genomics Viewer (IGV) Screenshots of H3K27ac and RNA-seq for highly expressed genes in Purkinje cells (Itpr1 and GABBR2), Golgi cells (SGCD and TCF4), and highly expressed genes in both Purkinje and Golgi cells (KCNMA1 and FRMD4A). j, Aggregate END-seq signal at DNA methylation valleys (DMV) within 118 AraC-induced downregulated genes.
Extended Data Fig. 6 The AraC-induced increase in γ-H2AX in Purkinje cells is TET- and TDG-dependent.
a, Representative examples of imaging flow cytometric analysis of Purkinje cell nuclei. BF (bright field), NeuN (green), γ-H2AX (yellow), ITPR-1 (red), DAPI (blue) and Merge (γ-H2AX and ITPR-1). TDG knockout (Tdg−/−) was induced by intraperitoneal injection of tamoxifen for CreERT2 TDGfl/fl mice. Pcp2-Cre Tet1/2/3fl/fl mice were used for Purkinje-specific knockout of Tet1/2/3 (Tet−/−). b, Quantification of γ-H2AX foci from imaging flow cytometric analysis. The genotype and treatment are indicated on the bottom. Data are presented as median with interquartile range, n = 110 and 579 (WT), 256 and 898 (WT with AraC), 101 and 129 (Tdg−/− with AraC), and 352 and 314 (Tet−/− with AraC) nuclei examined in 2 independent experiments. Statistical significance was determined using two-sided Mann-Whitney test, p value is indicated (p < 1e-15). c, Flow cytometric analysis of nuclei isolated from 6-month old mouse cerebellum after PBS or AraC treatment. Y- and X-axes indicate the expression of the Purkinje specific marker ITPR-1 and the DNA damage marker γ-H2AX respectively. γ-H2AX positive Purkinje cells are marked by red. For the flow cytometry gating strategy, see Supplementary Fig. 2. d,e, Quantification of γ-H2AX level by flow cytometry. The genotypes and treatments are indicated. Pcp2-Cre Tet1/2/3fl/fl mice were used for Purkinje-specific knockout of Tet1/2/3 (Tet−/−) in (e). For the flow cytometry gating strategy, see Supplementary Fig. 2.
Extended Data Fig. 7 Adenine arabinoside and Gemcitabine do not cause neurotoxicity.
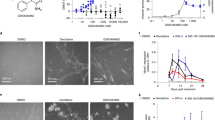
a, Images of γ-H2AX foci in wild-type iN treated with AraC (40 μM) or AraA (40 μM) on day 6. 4′,6-diamidino-2-phenylindole (DAPI) (blue); γ-H2AX (red). Scale bar, 10 μm. Similar results were observed in at least 2 independent experiments. b-d, Survival of iPSC-derived postmitotic neurons (b) and iPSCs (c,d) with indicated treatment. Y-axis represents viability relative to non-treated cells, X-axis represents days after treatment. Data are presented as mean ± SD, n = 3 biologically independent samples. e, Composite DNA sequence motif analysis 4 bp on either side of dFdC-S1-END-seq peak summits in iN. Peak fraction and p value are indicated. Statistical significance was determined using the two-sided binomal test. f, Schematic diagram represents the distance between the summits of dFdC-S1-END-seq and AraC-END-seq (with ‘GG’ motif). Median length: 21 bp. g, Enrichment of GG/CC and G/C motifs at AraC-END-seq peak summits within 25 bp of dFdC-S1-END-seq peak summits. Data are presented as mean ± SEM, n = 27234 (G/C) and 8510 (GG/CC) peaks. Statistical significance was determined using the two-sided Wilcoxon Rank Sum and Signed Rank Tests, p < 2e-16. h, Heatmaps of dFdC-S1-END-seq (SSB) and dFdC-END-seq (DSB) peaks. The peaks are ordered by SAR-seq intensity and plotted 1 kb on either side of SAR-seq peak summits in iN. i, Images of NeuN and γ-H2AX in wild-type primary cortical neurons treated with AraC or dFdC (40 μM) on day 6. 4′,6-diamidino-2-phenylindole (DAPI) (blue); NeuN (green); γ-H2AX (red). Scale bar, 20 μm. Similar results were observed in at least 2 independent experiments. j, In vitro reconstitution of TDG-initiated base excision repair. Top, the structure of DNA substrate (50 nM) containing 5-carboxycytosine (caC) on both strands, the DNA length and fluorescein label (*, red) are indicated. Bottom, native PAGE gel (15%) separating the base excision repair product, the nucleoside triphosphate (50 μM of each dCTP, dFdCTP and AraCTP), enzymes (50 nM of each TDG, APE1, PolB and XRCC1/LIG3), incubation time and product length are indicated. Two of the three replicates are shown. For gel source data, see Supplementary Fig. 1. k,l, In vitro reconstitution of TDG-initiated base excision repair using the same setting as in (j) but either replaced XRCC1/LIG3 by XRCC1/LIG4 (k) or omitting the final ligation step (l). For the gel source data, see Supplementary Fig. 1. m, Knockdown efficiency tested in neurons. X-axis represents the expression of XRCC1 in knockdown iN relative to that in wild-type iN transfected with a control vector (WT). Statistical significance was determined using two-sided paired t test, p = 2.1e-6. n, Western blot analysis of XRCC1 protein levels in wild-type (WT) and XRCC1 knockdown (sgXRCC1) iPSCs. o, Genome browser screenshot showing double-strand breaks (END-seq) detected in dFdC (40 μM) treated wild-type and XRCC1 knockdown iN on day 7. p, Proposed model for active DNA demethylation interrupted by AraC and dFdC at neuronal enhancers. Neurons employ sequential DNA demethylation of symmetrically methylated CpGs to prevent the formation of DSB at enhancers. This mechanism involves the ligation of dFdC by XRCC1/LIG3 during base excision repair, resulting in SSB but minimal DSB formation. However, the unligatable nature of AraC leads to generation of massive SSBs and DSBs at neuronal enhancers.
Extended Data Fig. 8 Proposed Model for AraC-induced cerebellar toxicity.
In Purkinje cells (PCs) of the cerebellum, a subset of highly expressed genes essential for motor coordination (Itpr1, Grid2 and etc.) undergoes active DNA demethylation. These genes are extensively covered with broad H3K27ac domains associated with enhancer marks. Active DNA demethylation involves TET-mediated oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxycytosine (5caC), followed by TDG-mediated excision of 5fC/5caC. AraC can interrupt base excision repair leading to double-strand breaks (DSBs) within gene bodies. AraC-induced DSBs can directly decrease gene expression or be processed into insertion/deletions (indels) and translocations, which can contribute to the development of cerebellar ataxia.
Supplementary information
Supplementary Information
Supplementary Figs. 1 and 2, which contain the uncropped gels and the flow cytometry gating strategy.
Rights and permissions
About this article

Cite this article
Liu, JC., Wang, D., Callen, E. et al. Mechanism of cytarabine-induced neurotoxicity. Nature 643, 1400–1409 (2025). https://doi.org/10.1038/s41586-025-09210-9
Received:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-09210-9
This article is cited by
-
Same same but different
Nature Reviews Cancer (2025)
