
Abstract
The time interval between about three and two million years ago is a critical period in human evolution—this is when the genera Homo and Paranthropus first appear in the fossil record and a possible ancestor of these genera, Australopithecus afarensis, disappears. In eastern Africa, attempts to test hypotheses about the adaptive contexts that led to these events are limited by a paucity of fossiliferous exposures that capture this interval. Here we describe the age, geologic context and dental morphology of new hominin fossils recovered from the Ledi-Geraru Research Project area, Ethiopia, which includes sediments from this critically underrepresented period. We report the presence of Homo at 2.78 and 2.59 million years ago and Australopithecus at 2.63 million years ago. Although the Australopithecus specimens cannot yet be identified to species level, their morphology differs from A. afarensis and Australopithecus garhi. These specimens suggest that Australopithecus and early Homo co-existed as two non-robust lineages in the Afar Region before 2.5 million years ago, and that the hominin fossil record is more diverse than previously known. Accordingly, there were as many as four hominin lineages living in eastern Africa between 3.0 and 2.5 million years ago: early Homo1, Paranthropus2, A. garhi3, and the newly discovered Ledi-Geraru Australopithecus.
Similar content being viewed by others

Main
Over the past few decades, the African hominin fossil record predating 3.0 million years ago (Ma) has expanded to include several potential ancestors for later taxa4,5,6,7. Currently, systematic analyses place A. afarensis as the most likely candidate for the middle Pliocene stem taxon from which multiple later hominin genera (that is, Homo and Paranthropus), and possibly other species of Australopithecus, descended8,9,10. A. afarensis was geographically and temporally widespread, and fossil sites with its remains are known from Tanzania to northern Ethiopia and, potentially, Chad. However, temporally, A. afarensis is not known after 2.95 Ma (ref. 11).
Paranthropus and Homo are well-documented in the eastern African fossil record after 2.0 Ma, especially in the Omo-Turkana Basin and at northern Tanzanian localities12,13,14,15, but the hominin fossil record between the last appearance of A. afarensis (around 2.95 Ma) and 2.0 Ma is patchy. For example, no Paranthropus fossils have been recorded from the Afar Region to date, despite its presence in the Omo-Turkana Basin and at Nyayanga, Kenya2 at approximately 2.7 Ma and in the Upper Ndolanya Beds at Laetoli16 at about 2.66 Ma. Additionally, a Homo specimen at Ledi-Geraru, Ethiopia, extends the genus closer in time17 (2.78 Ma) to the last known appearance of A. afarensis1,11. However a simple cladogenic model of A. afarensis, or any other stem taxon, splitting into these daughter genera is complicated by the presence of A. garhi in the Afar3 at approximately 2.5 Ma.
The Ledi-Geraru Research Project (LGRP) area is located towards the northern extent of palaeoanthropological sites in the Afar Region, Ethiopia (Extended Data Fig. 1). New discoveries in Ledi-Geraru suggest that early Homo and Australopithecus were both present in the Afar Region before 2.5 Ma, just as early Homo and Paranthropus are sympatric in the Omo-Turkana Basin and sites to the south18,19 after about 2 Ma. Whether the apparent absence of Paranthropus from the Afar Region reflects the spotty nature of the fossil record or a biogeographical signal is yet to be determined. What is clear is that the Afar Region has currently yielded the only definitive evidence for Australopithecus in eastern Africa after 2.95 Ma—A. garhi and the newly discovered specimens from Ledi-Geraru.
Ledi-Geraru geologic context
The LGRP area contains fossiliferous sediments from the critical 3.0–2.0 Ma time period17. The fossil sites in the Lee Adoyta and Asboli regions of the LGRP area are located west of the Awash River in a region incised by the Mille and Geraru River drainages and their tributaries (Extended Data Fig. 1). The 3.0–2.5 Ma fossiliferous sediments are exposed in fault blocks bounded by post-depositional northeast–southwest and nothwest–southeast trending faults (Fig. 1); age control is provided by many dated and correlated tephra deposits and by magnetostratigraphy17. Detailed 40Ar/39Ar dating methods are described in Methods.

a, Geologic map of the Lee Adoyta basin. Interbedded tuffs are shown as thin coloured lines. The black line indicates the position of the A–A′ cross-section shown in d. b, Stratigraphic sections at hominin localities showing the stratigraphic level of hominin fossil discoveries (yellow bones), fossiliferous horizons (white bones) and marker beds (coloured tuffs). Fossil and stratigraphic section locations are shown in a,c,d. c, Geologic map of the Asboli region superimposed on a hillshade image. Outcrops of multiple tephra deposits are shown as red lines, and these include the Giddi Sands and Lee Adoyta Tuffs that also occur in the Lee Adoyta region. d, South–north geologic cross-section of the Lee Adoyta basin showing the locations of hominin fossil sites. Vertical scale is doubled relative to the horizontal (2×VE).
The distribution and stratigraphy of fossiliferous units and tephra deposits provide context for the hominin fossils described here, which were discovered in sediments cropping out in the Lee Adoyta and Asboli regions (Fig. 1 and Extended Data Fig. 1). The Gurumaha sedimentary package is present in narrow fault-bounded exposures in the central Lee Adoyta basin and in drag-faulted blocks adjacent to basalt ridges bounding the basin to the east (Fig. 1a). The Gurumaha Tuff (GT), a light grey lapilli tuff dated to 2.782 ± 0.006 Ma (1σ; recalculated17,20), provides age control for the unit (Fig. 1b). Stratigraphically younger, the Lee Adoyta sedimentary package is widely exposed in the Lee Adoyta basin and is correlated to exposures in Asboli (Fig. 1). The Lee Adoyta Tuffs (LAT) comprise two thin, geochemically distinct tephras: a yellow altered basaltic ash which occurs approximately 10 cm above a white rhyolitic ash dated to 2.631 ± 0.011 Ma (1σ; recalculated17,20). These tuffs, and an underlying olive-green clay, provide an excellent marker sequence (Fig. 1b and Extended Data Fig. 2a). The Giddi Sands sedimentary package (Tgs; containing the Giddi Sands Tuff (GST)) and the Markaytoli sedimentary package (Tmk; containing the Markaytoli Tuff) crop out in the eastern Lee Adoyta basin (Fig. 1a). The base of the Giddi Sands sedimentary package is an erosional unconformity cutting into the Lee Adoyta sedimentary package in the Lee Adoyta Basin (Fig. 1b). The GST is a 3- to 8-cm-thick laminated, multicoloured (orange, yellow and white), crystal bentonite tuff. 40Ar/39Ar single-crystal incremental heating (SCIH) yielded a weighted-mean age of 2.593 ± 0.006 Ma (1σ) for the GST sampled in Asboli (Methods and Supplementary Figs. 1–3). In Asboli, the Giddi Sands sedimentary package containing the AS 100 fossil site unconformably overlies a sequence of mudstones containing the Asboli Tuffs, which in turn overlies the LAT exposed approximately 500 m to the south (Fig. 1c).
New Ledi-Geraru hominin specimens
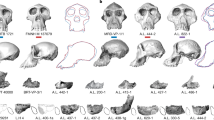
Hominin dental specimens (Fig. 2) were recovered from the three sedimentary packages described above—the Gurumaha, Lee Adoyta, and Giddi Sands (Table 1). Comparative methods and sampling protocols are described in Methods.

Right, from top: LD 302-23 P3, LD 750 P4, AS 100 M1 and AS 100 M2. Left, images show the LD 760 assemblage (top, from left: maxillary molar, I2, I1, maxillary canine; bottom, mandibular molars).
Gurumaha hominin specimen
A mandibular right third premolar, LD 302-23 (Fig. 2 and Extended Data Fig. 3), was recovered approximately 22 m to the southwest and 7 m stratigraphically below the LD 350 early Homo mandible locality1, above the Gurumaha Tuff17 (Fig. 1b). An enamel fragment is missing from the lingual corner, but the crown is otherwise well preserved. The crown is shorter mesiodistally (MD) than buccolingually (BL) (11.5 mm BL × 10.5 mm MD; Extended Data Fig. 3) and the major crown axis is oriented buccolingually. A relatively small metaconid sits well mesial to the protoconid, creating a tiny, centrally placed anterior fovea that is bounded by a low, but continuous, mesial marginal ridge. Occlusal wear makes it difficult to determine if the metaconid would have been completely distinct from the protoconid at the outer enamel surface. The talonid is mesiodistally short and occupies a relatively small portion of the crown area in occlusal view.
LD 302-23 departs strongly in shape and occlusal form from P3s attributed to Australopithecus anamensis and Australopithecus deyiremeda, and unicuspid specimens of A. afarensis (for example, A.L. 128-23 and A.L. 288-1), in which the major axis of an asymmetric crown runs mesiobuccal to distolingual and in having a continuous mesial marginal ridge that creates a ‘closed’ anterior fovea7,21,22 (Extended Data Fig. 3a). Some A. afarensis specimens (such as A.L. 437-2) resemble the crown outline of LD 302-23 (Extended Data Fig. 3a); however, LD 302-23 also differs from those specimens in having a mesially shifted metaconid, which creates an acute angle between the transverse crest and mesial protoconid crest, and in having a highly reduced anterior fovea relative to crown size23. In crown shape index (MD/BL), LD 302-23 falls in the middle range of the A. afarensis distribution (Extended Data Fig. 3b,c). In area (BL × MD), LD 302-23 falls within the range of the A. afarensis size distribution, at the lower limit of the range of Paranthropus boisei, but within the range of values of Paranthropus robustus (Extended Data Fig. 3b,d). Unlike Paranthropus12, the talonid of LD 302-23 is not expansive. We attribute LD 302-23 to Homo for a number of reasons. Although the sample of P3s attributed to early Homo is morphologically diverse (for example, KNM-ER 5431, OMO 75-14, KNM-ER 1802 and OH 7), LD 302-23 is clearly derived relative to pre-3.0 Ma Australopithecus, lacks the suite of derived nonmetric features characteristic of Paranthropus, and is consistent with the size, crown morphology and mesiodistal compression of the LD 350-1 specimen (the LD 350-1 P3 crown is damaged mesiolingually, with only around 65% of the crown remaining), which was found in the same sedimentary package and shares numerous dental and mandibular apomorphies with Homo.
Lee Adoyta hominin specimens
An isolated P4, LD 750-115670 (Fig. 2, Extended Data Fig. 4 and Supplementary Fig. 4), was collected from locality LD 750 at the base of an approximately 8-m-thick fossiliferous exposure of mudstone and sandstone. The stratigraphic position of the locality lies between the 2.63 Ma LAT and the 2.59 Ma GST (Fig. 1b and Extended Data Fig. 2a). The LD 750 P4 is unworn, with salient mesial cusps (Extended Data Fig. 4a). The roots are broken beneath the crown, but the mesial portion preserves a maximum root height of about 2.0 mm. The tooth may not have been fully erupted, as there are no interproximal contact facets and the occlusal surface is unworn. The protoconid and metaconid are shifted relatively mesially on the crown and tightly compressed BL, giving the tooth a slightly ‘puffy’ appearance. The anterior fovea is distinct and deep, but relatively small (reflecting the mesial placement of the cusps), centrally placed and bounded by a narrow mesial marginal ridge. The talonid is relatively large, the distal marginal ridge is low and rounded, and a distinct, but low, distolingual cusp is present, giving the tooth an asymmetric occlusal profile.
With dimensions of 12.4 mm BL × 11.4 mm MD, the tooth area is in the upper size range of A. afarensis, exceeded in area only by A.L. 996-1 (Extended Data Fig. 4b). Morphologically, however, the LD 750 P4 departs from the typical Hadar A. afarensis appearance. For example, the lingual to mesial face transition is smooth and rounded, whereas in Hadar P4s there is a more distinct corner. The occlusal layout differs from that of Hadar P4s in that the metaconid and protoconid are placed more centrally in relation to buccolingual crown breadth and shifted mesially in relation to crown length. Thus, compared with Hadar A. afarensis teeth, the buccal face is more convex occlusocervically and the anterior fovea more confined. The index of protoconid–metaconid apex distance/BL breadth shows the central placement of the LD 750 mesial cusp apices in relation to crown breadth compared with A. afarensis (Extended Data Fig. 4c,d).
Not only is LD 750 large relative to most A. afarensis specimens, it also exceeds the area of all specimens attributed to A. anamensis and all but two specimens of Australopithecus africanus (StW 498 and StW 384). The LD 750 crown area falls within the size distribution of P. robustus and Paranthropus aethiopicus; however, it lacks any of the distinct ‘molarization’ seen in Paranthropus P4s (for example, expanded talonid and distal cusps, thick marginal ridges, symmetrically rounded occlusal profile caused by the filling of the crown distobuccally, and surface complexity in unworn specimens12). We note that the known specimen of A. garhi lacks any mandibular remains3, but LD 750 cannot be excluded from that species on the basis of size alone (see randomization analysis in Extended Data Fig. 4e). In occlusal form and crown outline, LD 750 does resemble OMO 75-14 (Extended Data Fig. 4a), which has been attributed to aff. Homo by Suwa et al.12 and Hlusko et al.24, and KNM-ER 5431, which is also attributed to Homo by Suwa et al.12. However, these attributions are provisional and based mostly on the nonmetric morphology of their associated molars (for example, presence of C7 on M1 and M2 of both specimens). In fact, P4 morphology poorly distinguishes between early Homo and Australopithecus12,24. Thus, the phenetic resemblance between OMO 75-14, KNM-ER 5431 and LD 750, which are all large but lack features typical of Paranthropus, could be most parsimoniously interpreted as symplesiomorphic retentions. Since LD 750-115670 lacks any clear Homo synapomorphies associated with other specimens of early Homo from Koobi Fora (such as talonid reduction in KNM-ER 99225), we provisionally assign it to aff. Australopithecus.
An associated set of five mandibular molars, a maxillary molar fragment, and three maxillary anterior teeth (Fig. 2 and Table 1) representing a single individual were collected at locality LD 760, a relatively flat-lying area dominated by fine-grained sand and silts (Supplementary Video 1). The hominin teeth were all found in close association, in a relatively dense cluster among scattered faunal elements (Extended Data Fig. 2b). The mandibular molars were found eroding from the surface and the maxillary dentition was sieved from the surface silt (approximately 5–10 cm of loose and eroded surface soil) downslope on a gentle gradient over a 7 × 4 m area to the northwest. The LD 760 collection area lies approximately 10 m below the 2.631 ± 0.011 Ma LAT17 mapped in hillslopes surrounding the site (Fig. 1a and Extended Data Fig. 2a,b). Mammalian fossils were also observed eroding from sand units around 3.5 m and 6.5 m above the LAT (Fig. 1b). However, on the basis of the slope and distance to these sand units, as well as the abundance and spread of other fossil specimens, the hominin teeth and other fossils on the LD 760 surface represent a distinct fossiliferous unit below the LAT.
The LD 760 mandibular molars are moderately worn, with dentine exposure visible at buccal cusp apices in the M1 (Fig. 3). They are relatively wide BL compared to their MD length (Extended Data Figs. 5 and 6) and do not taper strongly distally, which gives the crowns, especially M1 and M2, a squarish profile (Fig. 4). The buccal grooves are indistinct, and the buccal profile is convex in occlusal view, in part because a protostylid partially fills in the buccal groove. Further, the buccal crown face slopes occlusocervically, reflecting an internal placement of cusp apices.

Left molars from Ledi-Geraru specimen LD 760 (left) and Hadar specimen A.L. 400-1 (right). Measurements in mm of the LD 760 molars (BL × MD): LM1: 13.3 × 13.4, LM2: 14.5 × 14.6, LM3: 14.0 (estimated) × 15.7, RM1: 13.2 × 13.1, RM2: 14.8 × 15.2 (Supplementary Data 1). Specimens are oriented with their buccal surfaces to the left and mesial surfaces up.

Detail of distal taper (orange lines) and bilobate contour (blue lines) seen in A. afarensis M1 specimen A.L. 400-1 (right) contrasted with the overall more equilateral occlusal profile of the LD 760 M1 (left). The M2s also show this pattern (Fig. 3). Specimens are oriented with their buccal surfaces to the left and mesial surfaces up.
There is no hint of M3 reduction (as seen, for example, in some early Homo such as LD 350-1 and OMO-75-14); instead, the molars follow the plesiomorphic pattern of relative molar size, M1 < M2 < M3. The M1 and M2 are subequally square (Extended Data Figs. 5 and 6), unlike in early Homo which tend to have rectangular crowns that are mesiodistally elongated. Further, the teeth are relatively simple, with few accessory cusps; the M1s show no expression of a C7, unlike LD 350-1 and KNM-ER 1802, and the M2s also lack a C7, which is seen in many specimens attributed to early Homo (for example, OMO-75-14, KNM-ER 5431, KNM-ER 60000 and OH 7). The mesiodistal dimensions of the mandibular molars comfortably fit within the metric variation seen in individual A. afarensis molars (Extended Data Fig. 5), and the buccolingual breadths fall in the upper third of the A. afarensis distribution. However, the sum of the molar areas for the LD 760 individual (614.3 mm2) exceeds all but one (NFR-VP-1/29; Woranso-Mille, Afar Region, Ethiopia) of the 11 A. afarensis individuals that preserve all 3 molars; yet, among Paranthropus, only 1 individual (DNH 7) is smaller in M1–M3 area than LD 760 (Extended Data Fig. 5d). The LD 760 molars depart from the typical feature set of A. afarensis in three ways: (1) the M1s are buccolingually broad relative to their mesiodistsal length (A. afarensis M1s and M2s are buccolingually widest across the mesial cusps, the LD 760 M1 is equally wide at midcrown and the M2 is only slightly narrower at midcrown) (Extended Data Figs. 5 and 6); (2) the M1 and M2 do not taper distally to the extent seen in A. afarensis (Figs. 3 and 4); and (3) the LD 760 molars lack the distinct bilobate buccal contour seen in A. afarensis12 (Fig. 4).
A fragmentary maxillary molar, LD 760-115911, was recovered at the LD 760 locality (Extended Data Fig. 7a). Although the protocone is the only fully preserved cusp, the lingual groove is distinct occlusally and the mesial edge of the hypocone is present. On A. garhi upper molars, the lingual groove is indistinct, but the lingual margin of the crown begins to curve buccally distal to the lingual groove, reflecting the relatively small hypocone and buccolingually narrow distal crown (Extended Data Fig. 7a). This reduction in hypocone area produces a triangular form for the Bouri specimen’s protocone. The LD 760 upper molar fragment shows no similar curvature of the crown margin distal to the lingual groove. Instead, it appears to have had a fully expressed hypocone. However, in terms of absolute molar area (BL × MD), the ratio of LD 760 lower molars to the A. garhi upper molars does not falsify a single-species hypothesis using modern ape comparative data—see resampling analysis in Extended Data Fig. 6b.
A right maxillary canine, a complete left maxillary lateral incisor and a left fragmentary maxillary central incisor were also found at the LD 760 locality (Fig. 2 and Table 1). The canine (LD 760-115979) is well preserved, with all but the root apex present (Fig. 5). The crown has mesial and distal interproximal contact facets. A distal interproximal contact facet is also present on the LI2, which indicates bilateral absence of a canine/I2 diastema; a diastema is present on the BOU-VP-12/130 A. garhi maxilla and is variably present in A. afarensis. The canine crown is worn apically and along the mesial and distal crests. Apical wear makes it difficult to examine the relative placement of the crown shoulders, but the mesial shoulder appears to have been placed apically. Despite the wear, the canine is not small (Extended Data Fig. 8a–c), and the ratio of the canine to molar mesiodistal dimensions puts this specimen in the upper range of Australopithecus and well beyond the observed values for Paranthropus (Extended Data Fig. 8d).

a, Lingual (left) and labial (right) views of the Ledi-Geraru LD 760-115979 canine (left) with Hadar A. afarensis specimens A.L. 763-1 (middle) and A.L. 333x-3 (right). Note that the LD 760 canine is a right canine, whereas the A. afarensis canines are from the left and are mirrored in these images. b–d, LD 760-115979 (b; shown in lingual view) contrasted with Hadar A. afarensis specimen A.L. 199-1 (c; right canine shown; distal to the upper right) and Bouri A. garhi specimen BOU-VP-12/130 (d; left canine, mirrored; distal is to the right). Note the simple mesial–distal chisel-like wear pattern on the LD 760 canine (b) in contrast to the complex multi-faceted wear pattern of A. afarensis (c) and the broad curved basin on the distal side of the A. garhi upper canine (d; this is seen on both left and right canines). Images are oriented differently to emphasize the distinctive relevant morphology. Photos of BOU-VP-12/130 A. garhi holotype fossil by T. White; use courtesy of the Middle Awash research project. Images in b–d are not to scale.
The LD 760 canine departs subtly from the macrowear of A. afarensis (Fig. 5). For example, its wear pattern lacks the characteristic ‘j-pattern’ along the distal crest26; instead, apical wear dominates. In basal size (10.4 mm BL × 10.5 mm MD) the tooth resembles A. afarensis and is narrower than the labio-lingually broad canine of A. garhi (Extended Data Fig. 9). The LD 760 canine also departs morphologically from A. garhi. The A. garhi canine, uniquely among hominins, possesses a broad, shallow distal basin reminiscent of a talon (Fig. 5d); this smooth basin is contiguous with a wide wear furrow that characterizes the entire postcanine dental row (Extended Data Fig. 9a).
The LD 760 individual cannot be attributed to Paranthropus. It lacks the derived reduction of anterior tooth size, which is reflected both in the absolute sizes of the anterior teeth and their size relative to the postcanine dentition (Extended Data Fig. 8d). The mandibular molars do not express relatively large C6s12 and they show a buccolingual wear gradient distinct from the planar pattern typical of Paranthropus12. The LD 760 teeth also lack any derived Homo traits (such as steeply walled molars, narrow M1, C7 presence and M3 reduction). We regard the morphology of LD 760 as inconsistent with known eastern African Australopithecus species because: (1) the lack of any of the derived A. garhi traits, notably the reduced hypocone of the maxillary molars and highly distinctive upper canine ‘talon’ basin; and (2) a morphology derived relative to A. afarensis (that is, lack of bilobate mandibular molar walls and distal taper, and canine wear pattern). Because of the plesiomorphic resemblance to A. afarensis, we make a generic-level attribution and assign the LD 760 assemblage to Australopithecus sp. indet.
Giddi Sands hominin specimens
A partial left M1 crown with lingual cusps only partially preserved (AS 100-1a) and 2 fragments assembled into a complete left M2 crown (AS 100-1b) were discovered eroded from the approximately 1-m-thick Giddi Sands unit in the Asboli region immediately below the GST (Fig. 2, Table 1 and Supplementary Fig. 5). Subsequent screening produced no additional hominin material.
Wear on these teeth is slight, with crown apices salient and no dentine exposed. Interstitial wear is present and the interproximal contact facets are congruent, consistent with identification of AS 100-1a as an M1 and AS 100-1b as an M2 (Fig. 2 and Extended Data Fig. 7). The AS 100-1b M2 is rhomboidal in occlusal outline, with the cusps offset such that the buccal pair sits mesial to the lingual pair, the hypocone is relatively large and projects distolingually, and the lingual profile is straight. A. afarensis M2s range in shape from squarish (for example, A.L. 199-1, A.L. 486-1) to more rhomboidal (for example, A.L. 444-2, A.L. 200-1)27, but A. afarensis specimens invariably show more pronounced lingual occlusocervical sloping, indicating a more internally placed cuspal arrangement than is apparent for AS 100-1b. The lingual cusps of the AS 100 molars are relatively steep vertically and lack the ‘puffy’ contour seen in relatively unworn specimens of A. afarensis (for example, A.L. 200-1). The rhomboidal crown outline, relatively large and projecting hypocone, and cuspal placement of AS 100-1b are closely matched by A.L. 666-1, a Homo specimen from the Busidima Formation at Hadar28 from 2.3 Ma, and MLP-1549, a Homo specimen from Mille-Logya29, 2.5–2.4 Ma. This feature set strongly contrasts with the A. garhi M2 in which the hypocone is relatively small and placed transverse to the metacone, the lingual profile is strongly curved, and the cusps are more internally placed (Extended Data Fig. 7a). In size, the AS 100-1b M2 (14.2 mm BL × 13.9 mm MD) and Homo specimen A.L. 666-1 are nearly identical and both slightly exceed the Mille-Logya M2 in BL breadth; AS 100-1b is smaller in area than the M2 of A. garhi and eastern African Paranthropus (Extended Data Fig. 7b). We attribute AS 100-1 to Homo sp. indet. based on the presence of a derived M2 crown outline (that is, rhomboidal with projecting relatively large hypocone30), a feature shared with other specimens of early Homo.
Taxonomic and phylogenetic implications
The presence of both early Homo and Australopithecus at Ledi-Geraru has implications for hominin taxonomy and diversity in this region in the 3.0–2.0 million year interval. Despite the relative paucity of fossils discovered in this time interval, evidence for multiple non-robust lineages in eastern and southern Africa indicates that taxonomic diversity had already evolved by 2.5 Ma. Here we examine taxonomic and phylogenetic hypotheses for the newly discovered hominin specimens from Ledi-Geraru.
First, although the Asboli sample contains only two molars (Table 1), and they predate Homo specimens A.L. 666-1 and MLP-1549 by more than 150,000 years, we regard the most parsimonious hypothesis to be that these are members of the same species of Homo30. The existence of Homo at Ledi-Geraru by around 2.78 Ma was previously established by the LD 350-1 mandible1; the new dental material from the Asboli area, as well as the LD 302-23 premolar described here from the Gurumaha sedimentary package, provide additional evidence of Homo prior to 2.5 Ma.
Second, LD 750 and LD 760 are separated by 24 m of strata and straddle the LAT. We provisionally regard all teeth recovered from the Lee Adoyta sedimentary package as representatives of a single species of Australopithecus, partially premised on our expectation of the low likelihood of two Australopithecus species existing in such close geographic and temporal proximity. The following are potential phylogenetic scenarios for the LD 750 premolar and the LD 760 teeth:
-
(1)
The Ledi-Geraru australopith teeth represent a late-surviving population of A. afarensis. The Lee Adoyta sample is approximately 350,000 years younger than the last appearance of A. afarensis, which is documented by specimens from the Kada Hadar 2 Submember at Hadar11. Contradicting this hypothesis, the mandibular M1s and M2s are not bilobate and are squarer in crown shape than the majority of A. afarensis; the P4 and molar sizes fall just at the upper limit of size variation in that species; and the pattern of maxillary canine wear in the LD 760 canine is not seen in A. afarensis. Accordingly, if these teeth do represent A. afarensis, they may capture further evolutionary change within the lineage. The resemblances between the Lee Adoyta Australopithecus specimens and A. afarensis are symplesiomorphic. In addition to A. garhi, this is the only other record of the persistence of this genus in eastern Africa after 2.95 Ma, although it has approximate contemporaneity with A. africanus found in South Africa.
-
(2)
The Lee Adoyta Australopithecus teeth represent a population ancestral to later Paranthropus, which is currently undocumented in the Afar Region; this inference could be supported by the large size of the postcanine teeth. However, the appearance of Homo by 2.78 Ma implies a divergence between that clade and Paranthropus by at least that date, and Paranthropus is present as early as 2.66 Ma in the Upper Ndolanya Beds at Laetoli, Tanzania16 and around 2.7 Ma at Nyayanga, Kenya2. Moreover, the Lee Adoyta teeth share no synapomorphies with Paranthropus (such as presence of C6 on the molars, reduced anterior tooth size or planar wear pattern) and megadonty is common in early hominin lineages, including A. garhi, A. africanus and early Homo3. Given these constraints, it is unlikely that the Lee Adoyta Australopithecus is ancestral to Paranthropus.
-
(3)
The Lee Adoyta Australopithecus teeth are early representatives of A. garhi. Accepting this hypothesis would require that A. garhi includes very different canine and maxillary molar forms and postcanine wear patterns on their upper and lower dentitions. This alternative is difficult to evaluate as there are only a few overlapping anatomical elements to compare between the current A. garhi fossil material and the Lee Adoyta Australopithecus sample. However, our assessment is that the lack of similar forms on the few shared elements suggests the Lee Adoyta australopith is not A. garhi.
-
(4)
The Lee Adoyta teeth are a previously unknown species of Australopithecus from the early Pleistocene. This hypothesis is the only alternative that offers no contradictions with the data presented in the previous three hypotheses.
The Ledi-Geraru fossils described here demonstrate that there were at least 3 lineages in the Afar Region between 3.0 and 2.5 Ma: Homo, A. garhi and the newly identified Australopithecus from Ledi-Geraru. The diversity of hominins in the interval around 2.5 Ma demonstrates the ways in which evolution was experimenting with the overall hominin pattern; in addition to these Afar region hominins, A. africanus is present in South Africa and Paranthropus is found in Kenya, Tanzania and southern Ethiopia by this time. The apparent absence of Australopithecus after approximately 2.0 Ma in eastern Africa means that only two hominin genera remained, Homo and Paranthropus, and these were well differentiated in terms of their dietary ecology31. Moreover, the discoveries of Australopithecus and Homo at Ledi-Geraru between 2.78 Ma and 2.59 Ma reveal that a drier, more open habitat was not uniquely associated with the appearance of Homo17,32, raising intriguing questions about niche breadth and landscape use among early hominins—including why Australopithecus was able to survive in the Afar Region alongside Homo until at least 2.5 Ma, and whether Paranthropus was competitively excluded from the Afar Region by late-surviving Australopithecus filling a similar niche.
Methods
40Ar/39Ar dating
Individual phenocrysts of K-feldspar (sanidine to anorthoclase, ~400–1,000 μm) from tuff sample LG-1048-1 of the Giddi Sands Tuff were analysed at the Berkeley Geochronology Center by the SCIH 40Ar/39Ar dating technique17. The feldspar concentrate was irradiated in the Cd-lined CLICIT position of the Oregon State University TRIGA reactor for four hours (BGC irradiation no. 461). Sanidine phenocrysts from the Alder Creek Rhyolite of California were used as the neutron-fluence monitor mineral, with an orbitally referenced age20 of 1.1848 ± 0.0006 Ma. Reactor-induced isotopic production ratios for this irradiation was: (36Ar/37Ar)Ca = 2.65 ± 0.02 × 10−4, (38Ar/37Ar)Ca = 1.96 ± 0.08 × 10−5, (39Ar/37Ar)Ca = 6.95 ± 0.09 × 10−4, (37Ar/39Ar)K = 2.24 ± 0.16 × 10−4, (38Ar/39Ar)K = 1.220 ± 0.003 × 10−2, (40Ar/39Ar)K = 2.5 ± 0.9 × 10−4. Atmospheric 40Ar/36Ar = 298.56 ± 0.31 (ref. 33) and decay constants follow ref. 34.
In total, 26 individual grains were analysed separately by the SCIH approach. One grain was rejected after the initial step as an old xenocryst (27361-08, ~100 Ma), while all others yielded geologically reasonable eruption ages and were carried to conclusion (Supplementary Table 1). All completed experiments demonstrated apparent-age plateaus, typically across the entirety of the 39Ar release spectrum (Supplementary Fig. 1), indicating an absence of complexity to the internal argon systematics, and that these feldspars show no evidence of alteration or inherited argon. A probability density plot of the plateau ages is shown in Supplementary Fig. 2, revealing that the age distribution is symmetrical and gaussian-like. The weighted-mean age of the accepted plateaus is 2.593 ± 0.006 Ma (1σ analytical error). Isochrons (36Ar/40Ar versus 39Ar/40Ar) calculated from the plateau steps yield an indistinguishable weighted-mean age of 2.597 ± 0.006 Ma (Supplementary Fig. 3) but, given the high proportion of radiogenic 40Ar of total 40Ar (typically >90%) in these crystals, we prefer the straightforward plateau weighted mean as the reference age for this sample (2.593 ± 0.006 Ma).
Dental analysis
Hominin specimens discovered in 2015 and 2018 from the LGRP area are described and evaluated taxonomically, using comparative fossil material from eastern and southern Africa including A. anamensis, A. afarensis, A. africanus, A. deyiremeda, A. garhi, Paranthropus aethiopicus, P. boisei, P. robustus, Homo habilis, Homo rudolfensis, and Homo sp. (see Extended Data Table 1 for sites, localities and references for these taxa). Resampling data from Pan troglodytes and Gorilla gorilla metrics are also used to evaluate the comparative relative variation within and between samples. Bootstrapping (resampling with replacement) was performed in R v. 4.4.135. The analyses were conducted using code written by the authors that utilizes the sample function. For each analysis, distributions of values for extant taxa were derived from 10,000 bootstraps. The ratio of the Ledi-Geraru and A. garhi specimens were compared to the bootstrapped distributions for chimpanzees and gorillas; all statistical tests were two-tailed using α = 0.05. All photographed material (including comparative specimens) were examined directly by the authors. Quantitative data came from a variety of sources, including the authors’ and published measurements; both quantitative and qualitative comparative analyses were performed. See Extended Data Table 1 for references and Supplementary Data 1 for all hominin dental metrics.
Survey and collection for each locality
LD 302-23
This mandibular premolar was found 6 February 2015 by K.G.T. during palaeontological survey ~22 m to the southwest and ~7 m stratigraphically below the LD 350 early Homo mandible locality1, above the 2.782 ± 0.006 Ma Gurumaha Tuff17 (Fig. 1b). The fossil was recorded in the LGRP database with GPS coordinates. Numerous non-hominin fossils were also recovered in the area.
LD 750-115670
This mandibular premolar was found by O.A.O. during palaeontological survey on 14 February 2018 (Supplementary Fig. 4). It was located ~9 m stratigraphically above the 2.63 Ma Lee Adoyta Tuff, on the eastern side of the Lee Adoyta Basin (Extended Data Fig. 2a) The fossils were recorded in the database with GPS coordinates and an in situ photograph. The sequence includes fine sandy silts to very fine sand, tuffaceous silts and brown mudstones.
LD 760-115329, LD 760-115685, LD 760-115533, LD 760-115316, LD 760-115323, LD 760-115979, LD 760-115934, LD 760-115884 and LD 760-115911
The first molar, LD 760-115329, an M1, was found by O.A.O. on 14 February 2018, followed by the discovery of four more lower molars by A.L.R. and O.A.O. within ~30 min. These teeth were located ~10 m stratigraphically below the 2.63 Ma Lee Adoyta Tuff on the eastern side of the Lee Adoyta Basin (Extended Data Fig. 2). All fossils surrounding the hominin teeth were flagged and collected during surveying for additional hominin material (Supplementary Video 1). The fossils were recorded in the database with GPS coordinates and in situ photographs. The area was subsequently (19–25 February 2018) screened with 2 mm screen mesh and the maxillary dentition was discovered in the surrounding 4 × 7 m in the upper 5–7 cm of loose surface material. Expanded sieving in 2020 did not produce additional fossils. Fossil survey, collection, and screening are shown in Supplementary Video 1. The sediments below the Lee Adoyta Tuff at the site include fine sands overlain by brown mudstones.
AS 100-1a and AS 100-1b
On 23 February 2015, one partial hominin molar, AS 100-1a (LM1), was found by B.M.A. and a tooth fragment, AS 100-1b, was subsequently found by E.M.L. during palaeontological survey (Supplementary Fig. 5). Further exploration of the small area revealed a second fragment, found by B.M.A., which fit onto AS 100-1b (completing the LM2). All teeth were found within 0.5 m2. All fossils were recorded in the database with GPS coordinates. The area was subsequently screened with 2 mm screen mesh without recovering more fossils. The sequence includes ~1 m thick sand (Giddi Sands) and thin pebble conglomerate units immediately below the Giddi Sands Tuff (Fig. 1).
For all localities, geological data were collected using StraboSpot 2.12.18, Avenza 5.4 and GaiaGPS 2025.2 and integrated in ArcGIS Pro 3.2.2.
Ethics and inclusion
The Ledi-Geraru Research Project has long partnered with local Afar team members to successfully conduct palaeoanthropological research in the Afar Region, Ethiopia. Three Ledi-Geraru Afar team members whose contributions and discoveries were critical for this study are recognized as co-authors. The Ledi-Geraru Research Project is committed to formally acknowledging the contributions of all our collaborators and team members in this and future work. This acknowledgement is one step in our long-term commitment to building research capacity and access for the local Afar community.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data are presented in Supplementary Table 1 (and online at https://tinyurl.com/LediGeraru40Ar39Ar) and Supplementary Data 1 (and online at https://tinyurl.com/LediGeraru).
References
Villmoare, B. et al. Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science 347, 1352–1355 (2015).
Plummer, T. W. et al. Expanded geographic distribution and dietary strategies of the earliest Oldowan hominins and Paranthropus. Science 379, 561–566 (2023).
Asfaw, B. et al. Australopithecus garhi: a new species of early hominid from Ethiopia. Science 284, 629–635 (1999).
Leakey, M. G. et al. New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433–440 (2001).
Leakey, M. G., Feibel, C. S., McDougall, I. & Walker, A. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature 376, 565–571 (1995).
White, T. D. et al. Ardipithecus ramidus and the paleobiology of early hominids. Science 326, 75–86 (2009).
Haile-Selassie, Y. et al. New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature 521, 483–488 (2015).
Strait, D. S., Grine, F. E. & Moniz, M. A. A reappraisal of early hominid phylogeny. J. Hum. Evol. 32, 17–82 (1997).
Strait, D. S. & Grine, F. E. Inferring hominoid and early hominid phylogeny using craniodental characters: the role of fossil taxa. J. Hum. Evol. 47, 399–452 (2004).
Mongle, C. S., Strait, D. S. & Grine, F. E. An updated analysis of hominin phylogeny with an emphasis on re-evaluating the phylogenetic relationships of Australopithecus sediba. J. Hum. Evol. 175, 103311 (2023).
Kimbel, W. H. & Delezene, L. K. ‘Lucy’ redux: a review of research on Australopithecus afarensis. Am. J. Phys. Anthropol. 140, 2–48 (2009).
Suwa, G., White, T. D. & Howell, F. C. Mandibular postcanine dentition from the Shungura Formation, Ethiopia: crown morphology, taxonomic allocations, and Plio-Pleistocene hominid evolution. Am. J. Phys. Anthropol. 101, 247–282 (1996).
Leakey, L. S. B. A new fossil skull from Olduvai. Nature 184, 491–493 (1959).
Leakey, L. S. B., Tobias, P. V. & Napier, J. R. A new species of the genus Homo from Olduvai Gorge. Nature 202, 7–9 (1964).
Leakey, R. E. F. Further evidence of Lower Pleistocene hominids from East Rudolf, North Kenya, 1972. Nature 242, 170–173 (1973).
Harrison, T. in Paleontology and Geology of Laetoli: Human Evolution in Context,Vol. 2 (ed. Harrison, T.) 141–188 (Springer, 2011).
DiMaggio, E. N. et al. Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia. Science 347, 1355–1359 (2015).
Wood, B. A. Koobi Fora Research Project Vol. 4: Hominid Cranial Remains (Clarendon Press, 1991).
Hatala, K. G. et al. Footprint evidence for locomotor diversity and shared habitats among early Pleistocene hominins. Science 386, 1004–1010 (2024).
Niespolo, E. M., Rutte, D., Deino, A. L. & Renne, P. R. Intercalibration and age of the Alder Creek sanidine 40Ar/39Ar standard. Quatern. Geochronol. 39, 205–213 (2017).
Davies, T. W., Delezene, L. K., Gunz, P., Hublin, J.-J. & Skinner, M. M. Endostructural morphology in hominoid mandibular third premolars: discrete traits at the enamel–dentine junction. J. Hum. Evol. 136, 102670 (2019).
Davies, T. W., Delezene, L. K., Gunz, P., Hublin, J.-J. & Skinner, M. M. Endostructural morphology in hominoid mandibular third premolars: geometric morphometric analysis of dentine crown shape. J. Hum. Evol. 133, 198–213 (2019).
Delezene, L. K. & Kimbel, W. H. Evolution of the mandibular third premolar crown in early Australopithecus. J. Hum. Evol. 60, 711–730 (2011).
Hlusko, L. J. et al. Taxonomic assignments for the 3.4 Ma to 1.1 Ma hominin postcanine teeth from the Usno Formation and the Shungura Formation, Lower Omo Valley, Ethiopia. Preprint at PalorXiv https://doi.org/10.31233/osf.io/g7kfx (2024).
Suwa, G. A Comparative Analysis of Hominid Dental Remains from the Shungura and Usno Formations, Omo Valley, Ethiopia. PhD thesis, Univ. California, Berkeley (1990).
Greenfield, L. O. Canine “honing” in Australopithecus afarensis. Am. J. Phys. Anthropol. 82, 135–143 (1990).
Kimbel, W. H., Rak, Y. & Johanson, D. C. The Skull ofAustralopithecus afarensis (Oxford Univ. Press, 2004).
Kimbel, W. H., Johanson, D. C. & Rak, Y. Systematic assessment of a maxilla of Homo from Hadar, Ethiopia. Am. J. Phys. Anthropol. 103, 235–262 (1997).
Alemseged, Z. et al. Fossils from Mille-Logya, Afar, Ethiopia, elucidate the link between Pliocene environmental changes and Homo origins. Nat. Commun. 11, 2480 (2020).
Davies, T. W. et al. Dental morphology in Homo habilis and its implications for the evolution of early Homo. Nat. Commun. 15, 286 (2024).
Teaford, M. F., Ungar, P. S. & Grine, F. E. Changing perspectives on early hominin diets. Proc. Natl Acad. Sci. USA 120, e2201421120 (2023).
Vrba, E. S. Late Pliocene climatic events and hominid evolution. In Evolutionary History of the Robust Australopithecines (ed. Grine, F. E.) 405–426 (Aldine de Gruyter, 1988).
Lee, J.-Y. et al. A redetermination of the isotopic abundances of atmospheric Ar. Geochim. Cosmochim. Acta 70, 4507–4512 (2006).
Min, K., Mundil, R., Renne, P. R. & Ludwig, K. R. A test for systematic errors in 40Ar/39Ar geochronology through comparison with U/Pb analysis of a 1.1-Ga rhyolite. Geochim. Cosmochim. Acta 64, 73–98 (2000).
R Core Team. R: A Language and Environment for Statistical Computing. www.R-project.org/ (R Foundation for Statistical Computing, 2024).
Alemseged, Z. et al. A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context. J. Hum. Evol. 49, 499–514 (2005).
Alemseged, Z. et al. A juvenile early hominin skeleton from Dikika, Ethiopia. Nature 443, 296–301 (2006).
Haile-Selassie, Y. & Ryan, T. M. Comparative description and taxonomy of new hominin juvenile mandibles from the Pliocene of Woranso-Mille (Central Afar, Ethiopia). J. Hum. Evol. 132, 15–31 (2019).
Haile-Selassie, Y. et al. Dentognathic remains of Australopithecus afarensis from Nefuraytu (Woranso-Mille, Ethiopia): Comparative description, geology, and paleoecological context. J. Hum. Evol. 100, 35–53 (2016).
Mbua, E. et al. Kantis: a new Australopithecus site on the shoulders of the Rift Valley near Nairobi, Kenya. J. Hum. Evol. 94, 28–44 (2016).
Melillo, S. M. et al. New Pliocene hominin remains from the Leado Dido’a area of Woranso-Mille, Ethiopia. J. Hum. Evol. 153, 102956 (2021).
White, T. D. New fossil hominids from Laetolil, Tanzania. Am. J. Phys. Anthropol. 46, 197–229 (1977).
White, T. D. Additional fossil hominids from Laetoli, Tanzania: 1976–1979 specimens. Am. J. Phys. Anthropol. 53, 487–504 (1980).
Brown, B., Brown, F. H. & Walker, A. New hominids from the Lake Turkana basin, Kenya. J. Hum. Evol. 41, 29–44 (2001).
Moggi-Cecchi, J., Grine, F. E. & Tobias, P. V. Early hominid dental remains from Members 4 and 5 of the Sterkfontein Formation (1966-1996 excavations): catalogue, individual associations, morphological descriptions and initial metrical analysis. J. Hum. Evol. 50, 239–328 (2006).
Haile-Selassie, Y., Saylor, B. Z., Deino, A., Alene, M. & Latimer, B. M. New hominid fossils from Woranso-Mille (Central Afar, Ethiopia) and taxonomy of early Australopithecus. Am. J. Phys. Anthropol. 141, 406–417 (2010).
Ward, C. V. Taxonomic affinity of the Pliocene hominin fossils from Fejej, Ethiopia. J. Hum. Evol. 73, 98–102 (2014).
Macchiarelli, R. et al. Early pliocene hominid tooth from Galili, Somali Region, Ethiopia. Coll. Antropol. 28, 65–76 (2004).
Ward, C. V., Leakey, M. G. & Walker, A. Morphology of Australopithecus anamensis from Kanapoi and Allia bay, Kenya. J. Hum. Evol. 41, 255–368 (2001).
Ward, C., Manthi, F. & Plavcan, J. New fossils of Australopithecus anamensis from Kanapoi, West Turkana, Kenya (2003–2008). J. Hum. Evol. 65, 501–524 (2013).
White, T. D. et al. Asa Issie, Aramis and the origin of Australopithecus. Nature 440, 883–889 (2006).
Kullmer, O. et al. The first Paranthropus from the Malawi Rift. J. Hum. Evol. 37, 121–127 (1999).
Dominguez-Rodrigo, M. et al. First partial skeleton of a 1.34-million-year-old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. PLoS ONE 8, e80347 (2013).
Leakey, R. & Walker, A. New Australopithecus boisei specimens from east and west Lake Turkana, Kenya. Am. J. Phys. Anthropol. 76, 1–24 (1988).
Prat, S., Brugal, J.-P., Roche, H. & Texier, P.-J. Nouvelles découvertes de dents d’hominidés dans le membre Kaitio de la formation de Nachukui (1, 65–1, 9 Ma), Ouest du lac Turkana (Kenya). C. R. Palevol. 2, 685–693 (2003).
Suwa, G. et al. The first skull of Australopithecus boisei. Nature 389, 489–492 (1997).
Braga, J. & Grine, F. E. New craniodental fossils of Paranthropus robustus from Kromdraai, South Africa (2014–2017 excavations). J. Hum. Evol. 188, 103481 (2024).
Bromage, T. G., Schrenk, F. & Zonneveld, F. W. Paleoanthropology of the Malawi Rift: an early hominid mandible from the Chiwondo Beds, northern Malawi. J. Hum. Evol. 28, 71–108 (1995).
Clarke, R. J. A Homo habilis maxilla and other newly-discovered hominid fossils from Olduvai Gorge, Tanzania. J. Hum. Evol. 63, 418–428 (2012).
Crevecoeur, I. et al. First early hominin from Central Africa (Ishango, Democratic Republic of Congo). PLoS ONE 9, e84652 (2014).
Grine, F. E. et al. Complete permanent mandibular dentition of early Homo from the upper Burgi Member of the Koobi Fora Formation, Ileret, Kenya. J. Hum. Evol. 131, 152–175 (2019).
Johanson, D. C. et al. New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature 327, 205–209 (1987).
Leakey, M. G. et al. New fossils from Koobi Fora in northern Kenya confirm taxonomic diversity in early Homo. Nature 488, 201–204 (2012).
Prat, S. et al. First occurrence of early Homo in the Nachukui formation (West Turkana, Kenya) at 2.3-2.4 Myr. J. Hum. Evol. 49, 230–240 (2005).
Spoor, F. et al. Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature 448, 688–691 (2007).
Tobias, P. V. Olduvai Gorge: The Skulls, Endocasts, and Teeth of Homo habilis (Cambridge Univ. Press, 1991).
Moggi-Cecchi, J., Menter, C., Boccone, S. & Keyser, A. Early hominin dental remains from the Plio-Pleistocene site of Drimolen, South Africa. J. Hum. Evol. 58, 374–405 (2010).
Stratford, D., Heaton, J. L., Pickering, T. R., Caruana, M. V. & Shadrach, K. First hominin fossils from Milner Hall, Sterkfontein, South Africa. J. Hum. Evol. 91, 167–173 (2016).
Acknowledgements
The authors thank the Ethiopian Heritage Authority, especially EHA representatives D. Zerihun in 2018 and G. Tekle in 2020; the Afar National Regional State Tourism Bureau; our 2015 Afar representative A. Mohammed; our 2020 representative A. Yusuf; T. White and the Middle Awash research project for access to and photographs of BOU-VP-12/130; and J. Moggi-Cecchi for A. africanus data. We acknowledge support of NSF Grant EAR-2020044. This research was funded by NSF Grant BCS-1853362, the Institute of Human Origins, and the School of Human Evolution and Social Change at ASU. We share decades of appreciation for Mesfin Mekonnen Bekele and the rest of our field crew. We dedicate this work to the late Charlie Lockwood, who was a founding director of this project and a talented and cherished colleague.
Author information
Authors and Affiliations
Contributions
K.E.R. and J R.A. designed, initiated and managed the overall project. A.L.R., B.V. and L.K.D. outlined the topic. B.V., L.K.D. and W.H.K. analysed and interpreted hominin data. B.M.A., M.A.H., O.A.O., E.M.L., A.L.R., K.E.R., J.R.R., I.E.S., E.S., K.G.T., B.V. and L.W. conducted field surveys, fossil recovery and analysis. J.R.A., C.J.C., D.C., E.N.D., D.A.F. and D.I.G. carried out geological survey, mapping, 3D modelling and analyses. E.N.D. created geology and overview figures and A.L.D. conducted geochronological analyses. B.V., L.K.D., A.L.R., K.E.R., E.N.D., C.J.C., D.A.F. and J R.A. wrote the manuscript with contributions and discussion from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Zeresenay Alemseged, Jacopo Moggi-Cecchi, Elizabeth Niespolo and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer review reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Map of the Ledi-Geraru Research Project area.
a, The Ledi-Geraru Research Project area (yellow star) is located towards the northern extent of palaeontological sites (red circles) in the Afar depression, Afar Region, Ethiopia. b, Within the Ledi-Geraru project area, the Lee Adoyta and Asboli fossil sites are located approximately 12.5 km apart.
Extended Data Fig. 2 Oblique and vertical views of the northern Lee Adoyta basin hominin sites.
a, Geologic mapping draped on orthophotograph created from unmanned aerial vehicle (UAV) images with cm-scale ground resolution, showing LD 750 and LD 760 fossil sites, including traces of the Lee Adoyta Tuffs (2.63 Ma; shown in green) and the Giddi Sands Tuff (2.59 Ma; shown in yellow), as well as NE-SW and NW-SE trending faults. Small offset faults (a few metres) in the Lee Adoyta sedimentary package (light green shading) are present near the fossil sites. b, Hominin specimens recovered from the LD 760 locality are represented by yellow stars and other faunal fossils by red circles. The hominin locality topography is relatively flat-lying, grading downslope gently to the northwest (Supplementary Video S1).
Extended Data Fig. 3 Comparative mandibular third premolar morphology and dimensions.
a, P3 occlusal morphology of eastern African hominins. A.L. 128-23, A.L. 333w-1 (mirrored), KNM-ER 5431, A.L. 437-2, OMO 33-73-5496, and LD 302-23. The crown of LD 302-23 measures 10.5 mm MD x 11.5 mm BL (est.). Mesial is oriented up and lingual is oriented to the right. Scale bar in centimetres; images are to scale. b, Bivariate plot of P3 crown dimensions. The LD 302-23 P3 (magenta star) falls near the upper range of Homo and well within Australopithecus. Authors’ data supplemented by references in Extended Data Table 1. Although LD 302-23 is large for a Homo P3, our interpretation of the occlusal morphology is more consistent with assignment to this genus. c, P3 (MD/BL ratio) shape variation. LD 302-23 falls in the middle range of Australopithecus and lower in the range of Homo, although these patterns show overlap. d, P3 variation in area (mm2). LD 302-23 falls in the upper ranges of Australopithecus and Homo but is small compared to P. boisei.
Extended Data Fig. 4 Comparative fourth premolar morphology and dimensions.
a, Comparison of the Ledi-Geraru LD 750-115670 P4 to other pre-2.0 Ma hominin specimens. All A.L. specimens attributed to A. afarensis. OMO 75-14 and LD 350-1 are attributed to Homo1,2, StW 404 and OMO L51-79 are attributed to Paranthropus1. LD 750-115670 measures 11.4 mm MD x 12.4 mm BL. Mesial is oriented up and lingual is oriented to the right. Scale bar in centimetres; images are to scale. b, Bivariate plot of P4 crown dimensions. The LD 750-115670 premolar (magenta star) falls at the upper range of A. afarensis P4 areas and just outside the range of eastern African early Homo. Authors’ data supplemented by references in Extended Data Table 1. c, Premolar cusp apices are marked in yellow with the outline of anterior fovea to aid identification of trait. d, Protoconid-metaconid cusp apex distance relative to overall BL breadth in unworn Hadar A. afarensis P4 specimens (n = 8 including A.L. 966-1, in D) and LD 750-115670. This specimen falls outside the known range of variation for unworn P4s from Hadar (unpublished data from W.H.K.). e, Ratio of Bouri (A. garhi) P4 area to LD 750 P4 area compared to bootstrapped ratios of upper to lower P4 areas from Pan troglodytes (n = 98P4s/95P4s) and Gorilla gorilla (n = 119P4s/120P4s). The ratio of the Bouri upper premolar to the lower LD 750 specimen could be resampled within the 95% confidence intervals of the randomly selected gorilla (two-way p-value = 0.486) and chimpanzee (two-way p-value = 0.402) samples; although there are no lower A. garhi premolars, the size of LD 750 does not preclude it from being within the appropriate size range for A. garhi based on gorilla and chimp models of upper premolar areas compared to lower premolar areas. The 95% confidence intervals were calculated from the bootstrapped distributions using the percentile method; no adjustments were made for multiple comparisons. Authors’ data supplemented by references in Extended Data Table 1.
Extended Data Fig. 5 Comparative mandibular molar dimensions.
a, Bivariate plot of M1 crown dimensions (mm) for LD 760 and comparative taxa. The LD 760 M1s fall just at the upper edge of Homo but well within Australopithecus. Note that LD 760 M1 crown dimensions have no overlap with P. boisei. b, Bivariate plot of M2 crown dimensions (mm) for LD 760 and comparative taxa. As with the M1s, the LD 760 M2s fall just at the upper edge of Homo but well within Australopithecus. Note the lack of overlap with Paranthropus. c, Bivariate plot of M3 crown dimensions (mm) for LD 760 and comparative taxa. The LD 760 M3 falls just at the upper edge of Homo but well within Australopithecus. Note the lack of overlap with eastern African Paranthropus. d, Area of the LD 760 M1–M3 molar row (mm2 on the y-axis) compared to distributions of eastern and southern African Australopithecus, early Homo, and Paranthropus. The LD 760 molar row area is larger than all but one A. afarensis individual. Authors’ data supplemented by references in Extended Data Table 1.
Extended Data Fig. 6 Molar shape variation.
a, Ratio of MD/BL measurements (mm) on the y-axis as a representation of shape variation for the LD 760 molars compared to Hadar A. afarensis and eastern African early Homo and Paranthropus molars. Authors’ data supplemented by references in Extended Data Table 1. b, Ratio of Bouri (A. garhi) upper molar row area to LD 760 lower molar area compared to bootstrapped ratios of upper to lower molar row areas from Pan troglodytes (n = 74 mandibular molar row areas/71 maxillary molar row areas) and Gorilla gorilla (n = 77 mandibular molar row area/87 maxillary molar row areas). The ratio of the Bouri upper molar row area to the lower LD 760 molar row area could be resampled within the 95% confidence intervals of randomly selected upper and molar rows from these two African apes (Gorilla gorilla two-way p-value = 0.196; Pan troglodytes two-way p-value = 0.269); the LD 760 molar row area size does not preclude it from being within the appropriate size range for A. garhi based on gorilla and chimpanzee variation. The 95% confidence intervals were calculated from the bootstrapped distributions using the percentile method; no adjustments were made for multiple comparisons. Authors’ data supplemented by references in Extended Data Table 1.
Extended Data Fig. 7 Comparative maxillary first and second molar morphology and metrics.
a, Comparison of M1 and M2 Australopithecus and Homo pairs with AS 100-1a and 100-1b. Hadar A. afarensis specimens A.L. 444-2 (opposite side, mirrored – note missing enamel on lingual M1), and A.L. 199-1. Australopithecus garhi specimen BOU-VP-12/130 (mirrored), early Homo maxilla A.L. 666-1, and fragmentary M1 specimen AS 100-1a and M2 specimen AS 100-1b (M2 MD 13.9 ×14.2 BL). Note the relatively small hypocone and expanded BL dimensions of the A. garhi specimen across the mesial cusps. The LD 760-115911 upper right molar (mirrored) preserves the protocone. The mesial edge of the hypocone is visible distal to the lingual groove. In A. garhi, the highly reduced hypocone results in a triangular form for the protocone as well as a generally expanded protocone-paracone axis in molars. The LD 760 upper molar specimen shows no sign of hypocone reduction. Photos of BOU-VP-12/130 A. garhi holotype fossil by T. White; use courtesy of the Middle Awash research project. Mesial is oriented up and buccal is oriented to the right. Scale bar in centimetres; images are to scale. b, Bivariate plot of M2 crown dimensions. Note that AS 100-1b (magenta star) falls within the size distribution of early Homo.
Extended Data Fig. 8 Comparative canine dimensions.
a, Bivariate plot of canine crown dimensions for LD 760 and comparative taxa. The LD 760 specimen falls within the size ranges of early Homo and Australopithecus but not Paranthropus. MD – mesiodistal; LaL – labiolingual. b, Ratio of MD/LaL measurements (mm) on the y-axis as a representation of shape variation for the LD 760 canine and comparative taxa. Authors’ data supplemented by references in Extended Data Table 1. MD – mesiodistal; LaL – labiolingual. c, Maxillary canine area (mm2) on the y-axis as a representation of size variation for the LD 760 canine and comparative taxa. Authors’ data supplemented by references in Extended Data Table 1. d, Upper canine MD length (mm) to lower molar MD length (mm) ratios for individuals preserving the maxillary canine and mandibular M1 and/or M2 for early Homo; Australopithecus (A. afarensis, A. africanus); and Paranthropus (P. boisei and P. robustus). The LD 760 canine/molar ratios do not fall within the Paranthropus range but are within the distribution for Australopithecus and Homo ratios. Authors’ data supplemented by references in Extended Data Table 1.
Extended Data Fig. 9 Comparative wear patterns.
a, The A. garhi BOU-VP-12/130 specimen has a very distinctive wear pattern with a continuous, broad longitudinal furrow from the maxillary canine to the molars, extending mesiodistally through the basin of each tooth, with differential wear on the lingual cusps (pers. obs., BV, LKD). This is different from the pattern on upper postcanine teeth in A. afarensis, A. africanus, P. robustus, and P. boisei, in which wear is more even across the breadth of the teeth (typically with a slight lingual-buccal wear gradient), and the cusps are ultimately reduced to the level of the central basin, producing a relatively flat occlusal profile. In A. garhi, the wear in the longitudinal furrow is sufficient to obliterate the central fissure, even in teeth with relatively unworn cusp apices (for example, contrast the P3s in the two specimens). We see no evidence of any wear pattern on the LD 760 assemblage that would indicate similarity to BOU-VP-12/130 (notably on the preserved fragmentary upper molar). The LD 760 wear pattern is more consistent with A. afarensis. b, In A. afarensis (A.L. 199-1), the canine lacks the large concave talon; the postcanine wear pattern is flatter with more wear on the cusps, ultimately approaching the level of the tooth’s central basin, which creates a flat rather than concave surface (note the visibility of the fissures on the first upper molar of A.L. 199-1 despite considerable cusp wear). Photos of BOU-VP-12/130 A. garhi holotype fossil by T. White; use courtesy of the Middle Awash research project. Scale in b is in centimetres.
Supplementary information
Supplementary Information
Supplementary Figs. 1–5
Supplementary Table 1
40Ar/39Ar analytical data. Data derived from individual grain analysis with the SCIH approach.
Supplementary Data 1
Hominin dental metrics including Ledi-Geraru and comparative specimens.
Supplementary Video 1
Survey, collection, and screening at Lee Adoyta localities.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Villmoare, B., Delezene, L.K., Rector, A.L. et al. New discoveries of Australopithecus and Homo from Ledi-Geraru, Ethiopia. Nature 650, 374–380 (2026). https://doi.org/10.1038/s41586-025-09390-4
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-09390-4
This article is cited by
-
Unlocking Ethiopia’s genomic landscape and its global significance: a call for inclusive genomics research
Human Genomics (2026)
-
Afar fossil shows broad distribution and versatility of Paranthropus
Nature (2026)
-
A variety of early hominin species shared the Afar region of Ethiopia
Nature (2026)
-
First known fossil hand of the hominin Paranthropus boisei
Nature (2025)
-
Early Oldowan technology thrived during Pliocene environmental change in the Turkana Basin, Kenya
Nature Communications (2025)
