
Abstract
Red Sea elasmobranch populations are facing alarming declines. Effective conservation efforts require management strategies informed by extensive datasets and by developing an understanding of distribution patterns within the basin, which is currently lacking. This study introduces CERSE (Central and Eastern Red Sea Elasmobranchs), a comprehensive compilation of elasmobranch observations in the central and eastern Red Sea basin following the route of the Red Sea Decade Expedition. The data set was curated from surveys conducted during two scientific cruises, a time-series survey within Red Sea Global project areas (Tabuk province), and observations extracted from peer-reviewed literature. Through the use of diverse methodologies, we provide a holistic view of elasmobranchs distribution in the region, for a total of 2,847 sightings recorded. By consolidating dispersed information from across the region, this dataset forms a comprehensive foundation for future research and monitoring endeavors focused on elasmobranch fauna, and will serve as a cornerstone for informing targeted conservation initiatives aimed at safeguarding endangered elasmobranch species in this region.
Similar content being viewed by others

Background & Summary
Global elasmobranch populations face severe threats mainly from overfishing and unregulated fisheries, leading to demographic changes and reduced abundance1,2,3. Sharks and rays, as keystone species, play a pivotal role in maintaining the health and balance of marine ecosystems, sustain many global fisheries4,5, and provide economic benefits such as ecotourism activities6,7,8,9. The decline of elasmobranch populations poses significant repercussions for marine ecosystems, impacting not only the intricate web of ocean life but also posing a threat to humans relying on these resources.
The Red Sea, known as a biodiversity hotspot, is highlighted as a critical risk area for these species. It hosts an array of different habitats, including mangroves, seagrass beds, salt marshes, and extensive geophysically diverse coral reefs along its coastlines and beyond10. Its uniqueness is further underscored by the pronounced latitudinal gradients in temperature, salinity, and nutrient levels from North to South11,12,13, significantly influencing the local fauna and flora14,15,16. However, this area is affected by chronical overfishing and illegal fishing activities persisting well after the Saudi Arabia’s 2008 shark-fishing ban, leading to declines in local abundances17,18. Moreover, data on Red Sea sharks and rays remain scattered, lacking cohesion with limited information available for most species.
There is, therefore, an urgent need for an extensive dataset encompassing observations across wide spatial and temporal scales to inform future management strategies, that is accessible to science and policy-makers in a comprehensive repository. These data are indispensable for identifying focal points of elasmobranch abundance and distribution patterns, laying the groundwork for subsequent studies and ongoing monitoring efforts within the region. Effective protection measures depend on understanding the conservation status of target species from both ecological and distributional perspectives. Integrating diverse data sources and survey methods is essential for monitoring elasmobranch status and ensuring successful strategies to aid the recovery of threatened populations19,20,21, aligning with the Saudi Arabian Vision 203022 and, on a broader scale, with the scope of the Kunming-Montreal Global Biodiversity Framework’s Goal A, Goal B, Target 4 and 5.
We present elasmobranch sighting data from two scientific cruises in the central and eastern Red Sea, a time-series survey at the Red Sea Project area (Tabuk Province, Saudi Arabia), and a comprehensive review of published literature. Out of the total, 49% of our dataset is derived from literature records. The dataset spans the entire route of the Red Sea Decade Expedition—the most extensive marine research cruise conducted to date along the eastern Red Sea.
The information reported was gathered through different survey approaches, spanning from aerial to manned submersible surveys, and represents the current state of knowledge on elasmobranch observations from the central and eastern Red Sea.
Political and national borders often pose significant challenges to the scientific community to integrate data at the basin level. This is the case also for research on elasmobranchs, which frequently cross exclusive economic boundaries. In the Red Sea, the western sector remains understudied. In the case of elasmobranchs, we are only aware of 4 studies that would have the requirements to be included in our dataset23,24,25,26, and further research efforts are being disrupted by conflict in some of the coastal nations. Hence, it is not yet possible to integrate data at the basin level, provided the huge observational gaps in the western margin of the basin. The data set provided here is intended to lay the groundwork for future collaborative efforts with more nations bordering the Red Sea. There is, however, evidence that elasmobranch populations in the western sector of the Red Sea may be in better status than those reported here for the easter sector. For instance, shark sighting was a major driver of tourism in Sudanese waters prior to the current civil war23. We hope that the data set reported here would contribute to encourage collaboration whenever the conditions allow to deploy collaborative research efforts. Indeed, the Regional Organization for the Conservation of the Environment of the Red Sea and Gulf of Aden (PERSGA) was created with the aim of improving collaboration toward a shared stewardship of the Red Sea and coordinate research efforts focused on marine conservation. The current study provides valuable insight, yet the inclusion of novel data from surrounding areas would significantly enhance our understanding of the broader Red Sea region. Furthermore, establishing a more extensive dataset would lay the groundwork for long-term monitoring efforts, more robust regional assessments and enable better prediction models. As this project evolves, collaborations with other research initiatives and institutions could play a crucial role in ensuring that future datasets are both comprehensive and standardized, facilitating comparative studies across different ecosystems and geographical settings. Ultimately, this broader approach aligns with the growing recognition of the need for large-scale, integrative datasets to address complex environmental challenges effectively.
Methods
Red Sea Decade Expedition: study area and duration of the cruise
Observations of elasmobranchs were gathered during the Red Sea Decade Expedition (RSDE) from February to June 2022. The cruise was divided into four separate legs, which covered the entire eastern Red Sea coast of Saudi Arabia. Two vessels, R/V OceanXplorer (OceanX) and R/V Al Azizi, were each being used to explore deep and shallow waters of the Saudi Red Sea: R/V OceanXplorer surveyed deeper waters, while R/V Al Azizi focused on shallow coastal domains (<30 m). Each leg was further divided into three sections: Phase 1 covered deep stations (from 500 m to the limit of the Exclusive Economic Zone), Phase 2 covered shallower stations (from 500 to 10 m), and a Discovery Phase dedicated to exploring specific seafloor features (e.g., brine pools). Manned submersibles and helicopters were deployed only during Phase 2 and the Discovery phases of the RSDE.
RSDE ROV & submersible surveys
R/V OceanXplorer was equipped with deep-sea vehicles, including a remotely operated vehicle (ROV) Chimera (Argus Mariner XL model) featuring 360-degree benthic cameras, as well as two Triton manned submersibles, Neptune (designed for scientific sampling operations) and Nadir (designed for media operations). Cameras and media equipment on the media submersible and ROV are listed in details in Table 1, while the scientific submersible was equipped only with Mini Cam Arctic Rays EagleRay 4 K. The duration of ROV and submersible dives were limited by weather and sea conditions, with an average duration of 5.3 hours per dive.
The ROV was outfitted with RBRmaestro3 CTD profilers (RBR), each featuring an RBRduo3 sensor measuring conductivity and temperature, an RBR Oxyguard sensor for measuring oxygen, and an RBR depth (pressure) sensor. The CTD underwent factory calibration prior to each expedition at the RBR laboratory. Footage and video clips of elasmobranch sighting were recorded and stored in an external hard drive (Western Digital, 2TB). Depth (m), time (UTC), habitat type, oxygen concentration (mg L−1), temperature (C°), salinity (PSU) and coordinates of sighting were recorded for each obervation.
RSDE drop cameras
To obtain video data, we deployed customized deep baited remote underwater lander systems (dBRUV27,28) to record mobile teleost and elasmobranch fishes in deep waters along the Saudi Arabian coast, across water depths ranging from 376 m to 1,500 m, totalling ~33 hours of drop camera deployments. The dBRUV systems consisted of a vertical carbon-fiber frame with pressure-tolerant flotation (G2200, McLane Research Laboratories Inc., East Falmouth, MA, USA) and an acoustic release system (CAT, EdgeTech, MA, USA). On each system, a single GoPro camera (GoPro Hero 5/6/7) was housed within a protective deepwater casing (GoBenthic, GroupB Incorporated, USA) and recorded video footage (1080p – 2.7 K video at 60 frames per second). The camera was attached to the frame ~1.5 m above the bottom of the unit. For each unit, two LED lights were used to illuminate the seafloor (SiteLite, Juice Robotics, Rhode Island, USA), with all electronics powered by a custom lithium-ion battery pack. Approximately 500 g of bait (e.g., skipjack tunas) was placed in a bait cage mounted on a carbon-fiber pole in front of the camera. After 4–7 hours of recording, the acoustic release system was manually triggered to release the drop weight and recover the system. We monitored temperature and depth using a calibrated Starmon TD stand-alone logger (Star Oddi, Iceland) or LAT1400 temperature and depth loggers (LOTEK). A depth-rated radio transmitter (F1845B, Advanced Telemetry Systems, Minnesota, USA) and float equipped with an orange flag were attached to facilitate surface retrieval with a YAGI antenna and VHF radio receiver (R410, Advanced Telemetry Systems, Minnesota, USA). Upon retrieval of the dBRUVs, videos were downloaded from micro-SD cards and reviewed at regular speed, noting elasmobranchs presence and maximum number (MaxN) of of individuals observed within a single video frame during the recording, starting from the moment the dBRUV touched the sea bottom until retrieval.
RSDE aerial surveys
R/V OceanXplorer is equipped with an onboard helicopter (Airbus H125). Flights were designed to follow a series of predetermined U-shaped flight plans that spanned along the entire coast of Saudi Arabia. Individual legs were separated by approximately ~3.7 km to avoid double counting individual sightings. A reference marker pole extending from either side of the helicopter allowed for a standardised 400 m belt transect along the flight path. Flights were operated at an altitude of ~200 m at a speed of ~130 km/h, with course and altitude held by autopilot except in instances where observer effort was discontinued to circle back for species identification. Flight durations was typically limited to ~3 h due to fuel limitations. Total flight time for the expedition was 131 hours covering a distance of 6,026 km with transect coverage of 4,248.5 km2.
RSDE SCUBA operations
SCUBA diving operations throughout the RSDE were undertaken with 2 main goals, media dives focused on underwater photography and underwater videography, and benthic habitat surveys conducted from the R/V Al Azizi. During the latter, 20 m transects (between 3 and 5 individual surveys per site) were conducted to study coral reefs and seagrass meadows. During dives elasmobranch observations were recorded opportunistically. The dives lasted an average of 45 minutes, reaching a maximum depth of 20 m. When possible, pictures of elasmobranch species were gathered for further analysis and identification (camera models Olympus TG5 and TG6), together with data on the surrounding environment and depth of the observation.
Southern Red Sea Expedition
The Southern Red Sea Expedition (SRSE) was a collaborative scientific cruise undertaken by the Japan Agency for Science and Technology (JAMSTEC) and King Abdullah University of Science and Technology (KAUST). The cruise was part of an integrated project entitled “Biodiversity and related geological controls of the southern deep Red Sea”. The cruise took place from 4 to 13 May 2023 onboard the vessel R/V Thuwal, in the southern Saudi Arabian Red Sea. The elasmobranch sightings from these surveys were opportunistic as the ROV was primarily deployed to survey the seafloor and targeted potential hydrothermal vent sites. The ROV survey started from the desired location and moved across a range of several meters to tens of meters from the targeted coordinate. At each station, the video recordings began at the water surface (ROV deployment) and ended when the ROV was retrieved onboard.
The videos were inspected, and the presence and depth of elasmobranchs was recorded. A sighting was defined as the observation of an individual in the field of view of the forward-facing ROV camera. A separate CTD deployment was carried out at each station. The CTD was a Seabird SBE-9, equipped with SBE11plus temperature, conductivity and pressure sensor, SBE-43 oxygen sensor, WET Labs ECO AFL/FL fluorometer, WET Labs ECO NTU turbidiy meter, PAR sensor and SUNA nitrate sensor.
Red Sea Global dataset
The Red Sea Global (RSG) dataset was generated with data gathered during monitoring efforts at project sites under the control of RSG including The Red Sea Project (TRSP) and Amaala, between Umluj and Duba (Tabuk Province, KSA). Different survey methods were used: underwater fish surveys (all sharks and rays observed in the course of fish survey dives, typically of around 40 minutes, on complex reef habitats in 20 m or less), manta tows operations, island surveys (observation of sharks and rays made by teams visiting island for bird or turtle nesting surveys), and opportunistic sightings (observations made at any other time by survey teams, typically traveling between survey sites). The data were collected from November 2020 to November 2022 and include shallow sightings (at the surface) down to 40 m depth.
Published datasets review
Additional data were gathered from a comprehensive literature review performed using Google Scholar (last accession date 10/08/2024). The search retrieved previously published reports of elasmobranch sightings in the eastern and central Red Sea. The search terms used include: elasmobranchs, Red Sea, sharks, Selachii, Batoidea, Saudi Arabia. We selected only papers presenting specific coordinates and depth of sightings. Data related to surveys which did not report the specific location or depth of recordings were excluded from our analysis. We extracted data from 14 published articles (Table 2). Coordinates for Rhincodon typus individuals presented in the study by Cochran et al. 2019 and Berumen et al. 2014 were retrieved from Sharkbook - wildbook for sharks (https://www.sharkbook.ai).
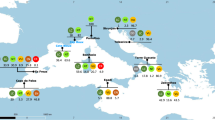
Figure 1 provides an overview of the principal sites discussed in this manuscript (Fig. 1A), along with maps depicting the spatial distribution of Selachii (Fig. 1B) and Batoidea (Fig. 1C), as recorded using various survey methods.

(A) Principal sites mentioned in the manuscript. NRS (Northern Red Sea), CRS (Central Red Sea), SRS (Southern Red Sea), this division follows the approach proposed by Wang et al.50. (B) Spatial distribution of Selachii sightings and (C) spatial distribution of Batoidea sightings, with different colors representing various survey methods. Map created using Esri ArcGIS Pro version 2.6.0 (Esri, 2020).
Traits of the dataset
The dataset includes 2,847 elasmobranch sightings, consisting of 897 batoid records and 1,950 shark records. The Selachii data represent 20 identified species from 11 genera across 8 families (Fig. 2A,B, Table 3). The Batoidea data include observations from 14 genera across 8 families, with 13 species identified (Fig. 3A,B, Table 4). Among Selachii genera within our dataset, Iago was the most abundant, with sightings exceeding 1,000 individuals (1,327 in total). Following the genus Iago, Carcharhinus, Triaenodon, and Rhincodon were the most abundant, with 210, 176, and 155 individual recordings, respectively. Sphyrna encompassed 28 sightings, Nebrius 15, Alopias 14, Mustelus 10, and Galeocerdo only 7 sightings, same as Stegostoma. Negaprion had the lowest numbers, with only 1 record.

Taxonomic distribution of Selachii recorded in our dataset across different families (A) and different genera (B).

Taxonomic distribution of Batoidea recorded in our dataset across different families (A) and different genera (B).
Among the Batoidea recordings, Rhinoptera exhibited high abundance, due to the presence of a huge aggregation of 125 individuals recorded during helicopter surveys in the southern Red Sea, 16.99279 N, 41.72041 S. The genera Taeniura, Aetobatus, and Glaucostegus recorded 235, 180, and 168 individuals, respectively. Himantura accounts for 71 individual sightings, while Pastinachus and Urogymnus had 42 and 23 sightings, respectively. The genus Mobula exhibited 39 sightings in total.
Low numbers of sightings were recorded for the genera Taeniurops with 6 observations and Rhinobatus with 4 observations. The least abundant genera are Rhynchobatus, Maculabatis, and Torpedo, with only 1 single record each.
In respect to the novel observations not derived from literature, a team of experts conducted the identification of various individuals down to the lowest taxonomic level possible, using species guides and references specific to the Red Sea. The World Register of Marine Species (WoRMS) served as the primary source for taxonomic classification. Elasmobranch species were visually identified based on key distinguishing features, such as body shape, coloration patterns, and the position and size of fins. Guidebooks and dichotomous keys were consulted throughout this process29,30,31,32,33.
Table 5 summarizes the number of sightings recorded using each survey method. A total of 302 sightings were documented through aerial surveys, with batoids representing the majority. The deployment of Baited Remote Underwater Video Systems (BRUVs and dBRUVs) accounted for 96 sightings, showing a relatively balanced distribution between Selachii and batoids. In contrast, fishing gear mostly reported shark sightings.
Island surveys offered insight into nearshore habitats with 337 encounters, although the effort predominately identified batoids. Manta tow operations contributed 110 sightings, with Selachii and Batoidea accounting for similar numbers. We also included 69 additional elasmobranch sightings referred as “Other”, considering records coming from incidental observations (e.g., boat observations).
ROVs and submersibles registered 257 sightings, primarily of Selachii, with only seven batoids identified. SCUBA operations, encompassing Underwater Visual Census (UVC) and off-transect data, recorded 308 sightings, highlighting its effectiveness in studying both sharks and batoids within their natural habitats. Although limited, snorkeling operations recorded higher batoid presence in shallower waters and few shark observations. Finally, tagging operations yielded 148 sightings, predominantly of sharks.
A total of 293 new observations were recorded from aerial surveys, supplemented by 9 additional observations from the literature. BRUV observations included 22 new RSDE records and 74 from the literature. All 1,194 fishing gear records were sourced exclusively from the literature. Island surveys contributed 336 records from the RSG dataset and 1 from the literature. Manta tow surveys yielded 24 records from the RSG dataset and 86 from the literature. For the “Other” category, 9 records were derived from the literature, while 60 were newly reported. ROV/SUB surveys included 33 records from the literature and 224 newly reported observations. Scuba surveys comprised 59 records from the literature and 249 new reports. All 26 snorkel survey records were sourced from the literature. Lastly, 148 tag dataset records were drawn entirely from the literature.
Data Record
The dataset is publicly accessible in Dryad repository34. The CERSE Dataset provides comprehensive information on all observations gathered from various datasets analyzed in this study. It encompasses taxonomic details at different levels, such as group, family, genus, and species, as well as the depth and coordinates of records, pelagic habitat, survey methods, and environmental factors, when available. Key data fields include the date of recording in the format dd/mm/yy, and the time of sighting in UTC. Species identification is provided at the species level when possible. Latitude and longitude are recorded in decimal degrees (DD), and the water depth of the observation is noted in meters.
The dataset classifies observations as Batoidea (rays) or Selachii (sharks). The specific pelagic zone where each observation was recorded is also detailed, along with the method used to gather the observation. The dataset sources include the Red Sea Decade Expedition (RSDE), Literature (a review of published literature), the Red Sea Global Dataset (RSG, within The Red Sea Project area), and the Southern Red Sea Expedition (SRSE). Habitat specifics describe the characteristics of the habitat where sightings were recorded, although this information is not available for all sightings. Additionally, the dataset provides temperature in degrees Celsius, salinity in practical salinity units (PSU), and oxygen values in mg L−1 at the time and depth of each observation, when available. Finally, the column titled ‘Video Reference’ indicates the name of the video or picture used for identifying the species observed, when available.
Technical Validation
Identification of Elasmobranchs
The nomenclature source used for taxonomic assignment is WoRMS (World Register of Marine Species https://www.marinespecies.org/). Sightings, photos and videos collected from all survey methods during the RSDE and SRSE, as well as from RSG area, were reviewed in the same manner.
Visual identification of elasmobranch species was conducted using distinguishing characters, such as body shape, coloration/patterns, as well as the position and size of fins. Guide books and dichotomous keys were referenced in all instances29,30,31,32. For sharks of the genus Carcharhinus we also followed Identification guidelines from Voigt and Weber33.
Oxygen data
Throughout the campaign, discrete water samples were taken at various depths (N = 110, from 42.8 – 2,268.7 m water depth) to verify dissolved oxygen concentrations measured by the RBR Oxygaurd sensor, according to ref. 35. Seawater was syphoned from the Niskin bottles using a sterilized drawing tube into 120 mL borosilicate flasks with glass stoppers, which were pre-calibrated for volume. The seawater was overflowed at least two volume equivalents (i.e., >240 mL), sealed, and fixed with 1 mL Manganese sulfate monohydrate (H2MnO5S) and 1 mL sodium hydroxide (NaOH)/sodium iodide (NaI) solution before being stored at 21 °C in darkness for at least 2 hours. After two hours, 1 mL sulfuric acid (H2SO4, 10 N) was added and the measurements were performed using sodium thiosulfate (Na2S2O3) with a T50 Titrator (Mettler Toledo) configured with an InMotion Pro autosampler (Mettler Toledo; after36,37).
The data from the RBR Oxygaurd sensor and Winkler titrations showed high agreement (R2 = 0.975), but with a negative offset of −16.03, −24.82, −33.61, and −38.48 µmol O2 L−1 for legs 1 – 4 in the RBR Oxygaurd measurements, respectively. Acknowledging this apparent sensor drift, we elected to align the RBR Oxygaurd with the calculated average offset for each leg. After the corrections were applied, the RBR Oxygaurd and Winkler titrations showed higher agreement with an R2 > 0.982. Data were processed using ‘respirometry’ package in R (R version 4.3.2 2023-10-31).
Code availability
The authors declare that no custom code was used to generate or process the data described in the manuscript.
References
Almojil, D. Local ecological knowledge of fisheries charts decline of sharks in data-poor regions. Mar Policy 132, (2021).
Ferretti, F., Myers, R. A., Serena, F. & Lotze, H. K. Loss of Large Predatory Sharks from the Mediterranean Sea. Conservation Biology 22, 952–964 (2008).
Pacoureau, N. et al. Half a century of global decline in oceanic sharks and rays. Nature 589, (2021).
Yagnesh, M., Durga, F., Raj, D., Rehanavaz, M. & Poojaben, T. Importance of sharks in ocean ecosystem. ~ 611 ~ Journal of Entomology and Zoology Studies 8, (2020).
Hammerschlag, N. et al. Ecosystem Function and Services of Aquatic Predators in the Anthropocene. Trends Ecol Evol 34, 369–383 (2019).
Anderson, R. C., Adam, M. S., Kitchen-Wheeler, A. M. & Stevens, G. Extent and economic value of manta ray watching in maldives. Tourism in Marine Environments 7, 15–27 (2011).
Gallagher, A. J. & Hammerschlag, N. Global shark currency: the distribution, frequency, and economic value of shark ecotourism. 14, 797–812 https://doi.org/10.1080/13683500.2011.585227 (2011).
Vianna, G. M. S., Meekan, M. G., Pannell, D. J., Marsh, S. P. & Meeuwig, J. J. Socio-economic value and community benefits from shark-diving tourism in Palau: A sustainable use of reef shark populations. Biol Conserv 145, 267–277 (2012).
Mustika, P. L. K., Ichsan, M. & Booth, H. The Economic Value of Shark and Ray Tourism in Indonesia and Its Role in Delivering Conservation Outcomes. Front Mar Sci 7, 261 (2020).
Jawad, L. The Arabian Seas: Biodiversity, Environmental Challenges and Conservation Measures. https://doi.org/10.1007/978-3-030-51506-5 (2021).
Sofianos, S. S. & Johns, W. E. An Oceanic General Circulation Model (OGCM) investigation of the Red Sea circulation, 1. Exchange between the Red Sea and the Indian Ocean. J Geophys Res Oceans 107, 17–1 (2002).
Raitsos, D. E. et al. Monsoon oscillations regulate fertility of the Red Sea. Geophys Res Lett 42, 855–862 (2015).
Chaidez, V., Dreano, D., Agusti, S., Duarte, C. M. & Hoteit, I. Decadal trends in Red Sea maximum surface temperature. Scientific Reports 2017 7:1 7, 1–8 (2017).
Voolstra, C. R. & Berumen, M. L. Coral Reefs of the Red Sea. Springer, Cham 11, (2019).
Racault, M. F. et al. Phytoplankton phenology indices in coral reef ecosystems: Application to ocean-color observations in the Red Sea. Remote Sens Environ 160, 222–234 (2015).
Sofianos, S. S. & Johns, W. E. An Oceanic General Circulation Model (OGCM) investigation of the Red Sea circulation: 2. Three-dimensional circulation in the Red Sea. J Geophys Res Oceans 108, 3066 (2003).
Spaet, J. L. Y. & Berumen, M. L. Fish market surveys indicate unsustainable elasmobranch fisheries in the Saudi Arabian Red Sea. Fish Res 161, 356–364 (2015).
Spaet, J. L. Y., Nanninga, G. B. & Berumen, M. L. Ongoing decline of shark populations in the Eastern Red Sea. Biol Conserv 201, 20–28 (2016).
Chevis, M. G., Graham, R. T., Chevis, M. G. & Graham, R. T. Insights into elasmobranch composition, abundance, and distribution in the Bocas del Toro Archipelago, Panama using fisheries-independent monitoring. Lat Am J Aquat Res 50, 492–506 (2022).
Rabaoui, L. et al. Diversity, Distribution, and Density of Marine Mammals Along the Saudi Waters of the Arabian Gulf: Update From a Multi-Method Approach. Front Mar Sci 8, 687445 (2021).
Garzon, F. et al. A multi-method characterization of Elasmobranch & Cheloniidae communities of the north-eastern Red Sea and Gulf of Aqaba. PLoS One 17, e0275511 (2022).
Khan, M. K. & Khan, M. B. Research, Innovation and Entrepreneurship in Saudi Arabia: Vision 2030. (Khan, M. K. and Khan, M. B. eds., Routledge, 2020).
Rezzolla, D., Boldrocchi, G. & Storai, T. Evaluation of a low-cost, non-invasive survey technique to assess the relative abundance, diversity and behaviour of sharks on Sudanese reefs (Southern Red Sea). Journal of the Marine Biological Association of the United Kingdom 94, 599–606 (2014).
Boldrocchi, G. et al. Annual Recurrence of the Critically Endangered Bowmouth Guitarfish (Rhina ancylostomus) in Djibouti Waters. Biology 2023, Vol. 12, Page 1302 12, 1302 (2023).
Rowat, D., Meekan, M. G., Engelhardt, U., Pardigon, B. & Vely, M. Aggregations of juvenile whale sharks (Rhincodon typus) in the Gulf of Tadjoura, Djibouti. Environ Biol Fishes 80, 465–472 (2007).
Boldrocchi, G., Omar, M., Azzola, A. & Bettinetti, R. The ecology of the whale shark in Djibouti. Aquat Ecol 54, 535–551 (2020).
Gallagher, A. J. et al. First records of the blurred lantern shark Etmopterus bigelowi from the Cayman Islands, Western Atlantic. Front Mar Sci 10, 1165207 (2023).
Phillips, B. T., Shipley, O. N., Halvorson, J., Sternlicht, J. K. & Gallagher, A. J. First in situ observations of the sharpnose sevengill shark (Heptranchias perlo), from the Tongue of the Ocean, Bahamas. Bahamas. Journal of the Ocean Science Foundation 32, 17–22 (2019).
Compagno, L. J. V. FAO Species Catalogue. v. 4: (2) Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date, Pt. 2: Carcharhiniformes. FAO Fisheries Synopsis 4 (1984).
Ebert, D. A., Dando, M. & Fowler, S. Sharks of The World: A Complete Guide. (2021).
Baranes, A. The Red Sea Sharks: Taxonomy, Biology and Ecology. 244 (2013).
Last, P., Naylor, G., Séret, B., de Carvalho, M. & Stehmann, M. Rays of the World. (CSIRO publishing., 2016).
Voigt, M. & Weber, D. Field guide for sharks of the genus Carcharhinus. 151 (2011).
Frappi, S. et al. CERSE: Central and Eastern Red Sea Elasmobranchs [Dataset]. Dryad https://doi.org/10.5061/dryad.ncjsxkt3z (2024).
Dickson, A. G. Determination of dissolved oxygen in sea water by Winkler titration. WHP Operations and Methods (1994).
Winkler, L. ‘Methods for Measurement of Dissolved Oxygen.’ (Ber Deutsch Chem Ges, 21, p.2843, 1888).
Carpenter, J. H. The Accuracy of the Winkler Method for Dissolved Oxygen Analysis1. Limnol Oceanogr 10, 135–140 (1965).
Goldshmidt, O., Galil, B., Golani, D. & Lazar, J. Deep-sea and extreme shallow-water habitats: affinities and adaptations.-Biosystematics and Ecology Series 11:271–298. Food selection and habitat preferences in deep-sea fishes of the northern Red Sea. download unter www.biologiezentrum.at Uiblein, F., Ott, J., Stachowtisch, M. (Eds): Deep-sea and extreme shallow-water habitats: affinities and adaptations. - Biosystematics and Ecology Series 11:271–298 (1996).
Clarke, C., Lea, J. S. E. & Ormond, R. F. G. Reef-use and residency patterns of a baited population of silky sharks, Carcharhinus falciformis, in the Red Sea. Mar Freshw Res 62, 668–675 (2011).
Clarke, C. R., Lea, J. S. E. & Ormond, R. F. G. Changing relative abundance and behaviour of silky and grey reef sharks baited over 12 years on a Red Sea reef. Mar Freshw Res 64, 909–919 (2013).
Braun, C. D., Skomal, G. B., Thorrold, S. R. & Berumen, M. L. Movements of the reef manta ray (Manta alfredi) in the Red Sea using satellite and acoustic telemetry. Mar Biol 162, 2351–2362 (2015).
Kattan, A., Coker, D. J. & Berumen, M. L. Reef fish communities in the central Red Sea show evidence of asymmetrical fishing pressure. Marine Biodiversity 47, 1227–1238 (2017).
Arostegui, M. C. et al. Vertical movements of a pelagic thresher shark (Alopias pelagicus): insights into the species’ physiological limitations and trophic ecology in the Red Sea. Endanger Species Res 43, 387–394 (2020).
McIvor, A. J., Spaet, J. L. Y., Williams, C. T. & Berumen, M. L. Unoccupied aerial video (UAV) surveys as alternatives to BRUV surveys for monitoring elasmobranch species in coastal waters. ICES Journal of Marine Science 79, 1604–1613 (2022).
Williams, C. T. et al. Low diversity and abundance of predatory fishes in a peripheral coral reef ecosystem. Ecol Evol 14, e10920 (2024).
Cochran, J. E. M. et al. Multi-method assessment of whale shark (Rhincodon typus) residency, distribution, and dispersal behavior at an aggregation site in the Red Sea. PLoS One 14, e0222285 (2019).
Berumen, M. L., Braun, C. D., Cochran, J. E. M., Skomal, G. B. & Thorrold, S. R. Movement Patterns of Juvenile Whale Sharks Tagged at an Aggregation Site in the Red Sea. PLoS One 9, e103536 (2014).
Spaet, J. L. Y., Lam, C. H., Braun, C. D. & Berumen, M. L. Extensive use of mesopelagic waters by a Scalloped hammerhead shark (Sphyrna lewini) in the Red Sea. Animal Biotelemetry 5, 1–12 (2017).
Pearce, J. R., Linley, T. D., Bond, T. & Jamieson, A. J. Depth distribution of the bigeye hound shark Iago omanensis and other deep-sea species observed by baited-camera in the Red Sea. Journal of the Marine Biological Association of the United Kingdom 103, e8 (2023).
Wang, Y., Raitsos, D. E., Krokos, G., Zhan, P. & Hoteit, I. A Lagrangian model-based physical connectivity atlas of the Red Sea coral reefs. Front Mar Sci 9, 925491 (2022).
Acknowledgements
We would like to thank the National Center for Wildlife NCW for the invitation to participate in the Red Sea Decade Expedition and the crew of OceanX and Al Azzizi for all the operational and logistical support for the entire duration of this expedition. We would like to acknowledge the ROV, submersible and media teams for the footage acquired and all the cruise leaders and scientific coordinators for the operations support on board. We thank Dr. Nicolas Pilcher for the valuable input and his constant support, data gathering and identification, as well as for his valuable insights during the RSDE expedition. We also acknowledge the contribution of Eleonora Re, Diego Rivera Rosas and Anastasiia Martynova in storing and securing the elasmobranch sightings data during the RSDE. All the videos, images and audio media obtained during the course of the RSDE are credited to the National Center of Wildlife, Kingdom of Saudi Arabia. We extend our sincere thanks to all KAUST and NCW members for their invaluable collaboration and support throughout the cruise, both on the Al Azizi and OceanX vessels. We gratefully acknowledge Hatim M. Abdulbaqi (Marine Instrumentation Engineer, Marine Engineering) and David Atienza (Oceanographic Specialist, Marine Operations) for their essential roles in obtaining the ROV video footage during the Thuwal-JAMSTEC cruise, as well as the precious contribution of the JAMSTEC team. We also extend our appreciation to the ROV operators, as well as the captain and crew, for their critical support throughout the expedition.
Author information
Authors and Affiliations
Contributions
S.F., I.A.C., A.K., C.W., R.H., S.H., L.S., A.G., T.B., O.S., M.R., S.G.K., S.A. collected the data used in this study. Data analysis and visualization performed by S.F., S.K., I.A.C., A.K., A.S., O.F.D., S.A. S.F. and C.M.D. conceptualized and structured the study. C.M.D. supervised the project. S.F. prepared the first draft of the manuscript, with contributions from C.M.D. The draft was reviewed by all authors. Resources and Founding acquisition: C.M.D., M.A.Q., V.P., B.J. M.A.Q., V.P. and C.M.D. designed the R.S.D.E. expedition.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Frappi, S., Klein, S.G., Arossa, S. et al. An integrated multi-source dataset of elasmobranchs in the Red Sea following the Red Sea Decade Expedition. Sci Data 11, 1425 (2024). https://doi.org/10.1038/s41597-024-04262-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41597-024-04262-4
