
Abstract
Ceracris kiangsu (Orthoptera: Arcypteridae), is greatly affected by climatic factors and exhibits strong adaptability, posing a serious threat to the ecological environment. Therefore, predicting its potential suitable habitat distribution provides a proactive theoretical basis for pest control. This study using the Biomod2 package of R simulated and predicted the current and future potential distribution, area changes, changes in the center points of suitable habitats, and niche shifts of C. kiangsu under two different greenhouse gas emission scenarios, SSP1-26 and SSP5-85. The results show that: (1) Currently, the high suitability areas for C. kiangsu are mainly distributed in Yunnan, Jiangxi, Hunan provinces in southern China and phongsaly province in northern Laos. In the future, the center of the suitable habitat distribution pattern of C. kiangsu will remain unchanged, primarily expanding outward from medium and high suitability areas. Additionally, significant suitable habitats for C. kiangsu were discovered in Southeast Asian countries without previous pest records. (2) Compared to the present, the overall suitable habitat area for C. kiangsu is expected to expand, particularly under the SSP5-85 climate change scenario. (3) In the SSP1-26 and SSP5-85 climate scenarios, the geometric center of the suitable habitat generally shows a trend of gradually shifting northeast. (4) Under different climate scenarios, the suitable habitat of C. kiangsu has highly overlapping, indicating that the suitable habitat of C. kiangsu in the invaded areas is broader than in its native regions. In conclusion, the research findings represent a breakthrough in identifying the potential distribution areas of C. kiangsu, which is of great practical significance for the monitoring and control of C. kiangsu pest infestation in China and Southeast Asian countries.
Similar content being viewed by others
Introduction
Global climate change represents one of the foremost challenges facing modern society1. As ectothermic organisms, insects rely entirely on external temperatures for their growth, development, and various physiological activities2. A Warming climate accelerates the developmental rate of insects throughout all life stages in all periods. It has been observed that the developmental period of each life stage of Acanthoscelides obtectus (Coleoptera: Bruchidae) shortens with increasing temperatures, and a positive correlation between developmental rate and temperature has been established3. A study investigating the impact of climatic conditions in China on the flight behaviour of S. gregaria (Orthoptera: Acrididae) found that extreme weather events, such as tropical cyclones, have created favorable conditions for the migration of S. gregaria towards the southern Tibetan border of China, thereby posing significant risks to agricultural and ecological systems4. Luo et al.5 sought to examine changes in the suitable habitats of Calliptamus italicus (Orthoptera: Oedipodidae) and Gomphocerus sibiricus (Orthoptera: Acrididae) under the influence of global climate change in Xinjiang, China5. Climate and geographical distribution data were combined with the CLIMEX model2 to verify that rising temperatures have expanded the suitable habitats of these two locust species, facilitating their invasion and settlement in new territories, with their distributions extending to higher latitudes and altitudes5. In summary, insects exhibit high sensitivity to changes in ambient temperature of their environments2. Consequently, elucidating the causes and mechanisms through which climate change drives the gradual expansion or contraction of suitable habitats for insects is a crucial objective in the field of ecology2.
Ceracris kiangsu (Orthoptera: Arcypteridae) is a polyphagous pest commonly referred to as the bamboo locust6. C. kiangsu has a varied diet, consumes substantial quantities of plant material, and inflicts damage over an extended period. Infestation of this pest primarily occur from late May to September each year, with the egg-laying period extending from late August to early October. The eggs overwinter in the soil7. C. kiangsu primarily damages moso bamboo (Gramineae: Phyllostachys) and other Gramineae plants8. It predominantly feeds on the leaves of the aforementioned plants, creating variously shaped notches. In severe infestations, it can completely consume the leaves, resulting in irreversible damage and causing the plants to appear fire-yellow from a distance (Fig. 1)9.

(a) Geographic distribution records of C. kiangsu; (b,c) scene of locust infestation: Yot Ou County, Phongsaly Province, Laos.
Historical records indicate that since 1368 A.D., southern China has faced infestations of C. kiangsu, resulting in significant economic losses10. Surveys reveal that since 2015, provinces such as Phongsaly and Luang Prabang in Laos have been inundated by C. kiangsu, which has severely impacted the growth of bamboo, Musa basjoo and other Gramineae plants11,12. In Phongsaly Province, the infestation primarily concentrated in Yot Ou, Boun Neua, and Phongsaly counties, affecting an area exceeding 40.4685 km2. The pest density was immeasurable, and the impact was severe. In 2018, C. kiangsu began migrating on a small scale to Jiangcheng County in Yunnan Province. By 2020, a large-scale invasion into the Chinese border commenced13,14. From 2020 to the present, Jiangcheng County in China has observed C. kiangsu migrating from the China-Laos border, resulting in varying degrees of locust infestations in areas such as Menglie Town, Niuluo River Nature Reserve, and Huangjiang Ridge. The cumulative affected area totals 80.319 km2, with direct economic amounting to 19.4 million yuan. The damage initially manifested as localized outbreaks and progressively spread to adjacent areas12.
Species Distribution Models (SDM) employ statistical or machine learning algorithms to predict the spatial probability of species presence or habitat suitability, based on occurrence records and environmental variables15,16,17,18,19. Previous studies have predicted and evaluated the distribution of C. kiangsu using only a single model, the Maximum Entropy model (MaxEnt). These studies primarily focused on China and overlooked the impacts of C. kiangsu in neighboring countries on the ecological environments of both local and Chinese borders11,14. The "Biomod2" package is an integrated platform in R for species distribution modeling20. The platform utilizes multiple modeling techniques to create ensemble models, which effectively reduce the biases inherent in individual models20. Biomod2 predictions are generally more accurate than those produced by MaxEnt21. Therefore, in this study, we expanded the study area and employed Biomod221 to predict the current and potential future geographic distribution, areas of overlap under climate change, and the ecological niche dynamics of C. kiangsu in China and Southeast Asia. This approach offers a theoretical foundation for enhancing our understanding and monitoring of the biological invasion of C. kiangsu, and supports early warning and integrated control measures. It is crucial for disaster prevention and risk assessment.
Materials and methods
Data source and processing
Data source and processing of distribution points of C. kiangsu
The distribution data of C. kiangsu were collected from multiple surveys including field surveys conducted in Jiangcheng County, China, and Yot Ou County, Laos; the Global Biodiversity Information Facility (GBIF, https://doi.org/10.15468/39omei); and scholarly literature17,22,23,24,25,26,27,28,29,30. Using ENMtools, we eliminated duplicate samples as per Wang et al.31, resulting in 284 unique distribution points (Fig. 1) and thereby minimizing clustering effect errors during the modeling process.
Data acquisition and screening of environmental factors
Future climate scenarios encompass four Representative Concentration pathways (RCPs): SSP1-26, SSP2-45, SSP3-70, and SSP5-85. This study predominantly utilized SSP1-26 and SSP5-85, which correspond to low and high greenhouse gas emission scenarios, respectively. Regarding future climate scenarios, most studies have relied on the Coupled Model Intercomparison Project Phase 5 (CMIP5); however, the latest CMIP6 version demonstrates significantly higher climate sensitivity32. The CMIP6 model simulations align more closer with observational data, reducing uncertainty and permitting a more comprehensive exploration of future climatic outcomes. Different scenarios imply varied impacts of socio-economic and technological developments on radiative forcing33. SSP1-26 and SSP5-85 are updated scenarios corresponding to RCP2.6 and RCP8.5, which represent radiative forcing stabilization at 2.6 W/m-2 and 8.5 W/m-2 by the year 2100, respectively32,33. These scenarios are frequently employed to assess species distribution under projected future conditions.
Climate environmental factors for the present, 2050s, and 2090s were sourced from the WorldClim Version 2.0 database (http://www.worldclim.org), from which 19 bioclimatic variables, Bio1 to Bio19, were downloaded. These variables encompass annual mean temperature, mean diurnal range, isothermality, temperature seasonality, maximum temperature of the warmest month, minimum temperature of the coldest month, annual temperature range, mean temperature of the wettest quarter, driest quarter, warmest quarter, coldest quarter, annual precipitation, precipitation of the wettest month, driest month, precipitation seasonality, and precipitation of the wettest, driest, warmest, and coldest quarter.
To eliminate multicollinearity and validate the authenticity of environmental factors, we utilized principal components analysis (PCA) to conduct a correlation analysis of the 19 environmental variables, taking into account the survival conditions of C. kiangsu. This step is crucial because multicollinearity among environmental variables can result in model overfitting and adversely affect prediction outcomes34,35. In the principal component analysis of each indicator, the principal (PCA) of each indicator, principal components were identified based on a cumulative contribution of 80% or higher34,35. Finally, four environmental factors were selected: bio_02 (mean diurnal range of temperature), bio_05 (max temperature of the warmest month), bio_08 (mean temperature of the wettest quarter), and bio_14 (precipitation of the driest month) (Fig. 2).

Principal component analysis(PCA) of environmental factors.
The world administrative map was obtained from the National Earth System Science Data Center, a part of the National Science & Technology Infrastructure of China (http://www.geodata.cn). Finally, the resolution of all environmental variables was standardized to 2.5 arc-minutes.
Biomod2 model construction
SDMs are widely employed to predict potential distributions across various scales, from global species diversity patterns and regional landscape-scale invasive species spread to local population distribution simulations36,37. Commonly applied SDM algorithms include Generalized Linear Models (GLM), Maximum Entropy Model (MaxEnt), Artificial Neural Networks (ANN), Random Forests (RF), among others.21,52,53. These models frequently link species distributions to climate data to elucidate variations in distributions and the critical environmental variables that determine species distribution patterns38. However, due to the multitude of species distribution models, their differing perspectives, and their respective strengths and weaknesses, achieving accurate predictions with a single model in challenging38. Integrating the predictive outputs of various models into an ensemble model can significantly enhance predictive accuracy20,52,53.
The Biomod2 ensemble model integrates ten widely used SDM algorithms, including Generalized Linear Models (GLM), Generalized Additive Models (GAM), Artificial Neural Networks (ANN), Random Forests (RF), and Maximum Entropy Models21,36,37. When obtaining real data is challenging, SDMs use randomly generated pseudoabsence (background or implied absence) data from the study area to extract available environmental information. This method is known as the pseudoabsence allocation method15,16. Biomod2 is an integrated platform for implementing SDM. It leverages modeling and classification techniques, including statistical and machine learning algorithms, to establish relationships between species and environmental variables. This platform is employed to predict the habitat suitability of species in geographical space20,38,39. Compared to single models, Biomod2 results are more objective and repeatable, independent of expert experience. They integrate species niche information, yield more accurate predictions, and can incorporate and analyze a greater number of environmental variables21,40.
Therefore, this study utilized the R package "Biomod2" to predict the current and future distribution of C. kiangsu in Asia, accounting for environmental factors and shifts in geometric center points and ecological niches. The primary operational approach of Biomod2 involves involves initially using the ten models provided by the package for individual model construction. A model demonstrating superior individual performance is selected as the foundational model. Subsequently, Biomod2 is employed to integrate and process this model, yielding a composite model framework41. Ultimately, the optimal combination model algorithm is applied to derive the ensemble model outcomes. Specifically, for each modeling process, 75% of distribution records were randomly selected as the training dataset, and the remaining 25% were designated as the test dataset, with the division between training and testing data repeated five times. Additionally, 1000 pseudo-absence points were randomly selected, repeated three times, resulting in 240 models. After evaluating the accuracy of the models generated by the eight algorithms, those with AUC values > 0.9 and TSS values > 0.8 were selected, including ANN, Classification Tree Analysis (CTA), Flexible Discriminant Analysis (FDA), Gradient Boosting Machine (GBM), and RF41,42,43,44.
Model accuracy
Model performance was evaluated using the area under the curve (AUC) and true skill statistics (TSS) metrics41. Both AUC and TSS range from 0 to 1, with higher values denoting greater accuracy of the model outcomes44. An AUC value between 0.8 and 0.9 indicates a good model prediction; while an AUC > 0.9 suggests high model precision, accurately reflecting the potential distribution of the species11. TSS is recognized for maintaining the advantages of Kappa concerning occurrence rate, sensitivity, and specificity in species distribution predictions, while avoiding its unimodal dependence on occurrence rate41. TSS values between 0.55 and 0.7 suggest average prediction outcomes; values from 0.7 to 0.85 indicate good prediction performance; and TSS > 0.85 signifies high model precision, accurately reflecting the potential distribution of the species41,42,43,44.
Spatial patterns of C. kiangsu in the present and future
Based on the model simulation results, ASCII raster layers were generated using suitability values (S, ranging from 0 to 1) to represent the potential presence of C. kiangsu. According to the species existence probability by IPCC-CMIP5 (IPCC 2014), the suitable habitats were classified into four categories: highly suitable areas (0.7 ≤ P ≤ 1.0), moderately suitable areas (0.5 ≤ P < 0.7), less suitable areas (0.3 ≤ P < 0.5), and unsuitable areas (0.0 ≤ P < 0.3). The areas of these habitat types were subsequently calculated for each period37.
Migration of potential geographic distribution center and overlapping geographic distribution areas
The shifts in the potential geographic distribution center of C. kiangsu under current and future climate scenarios were determined using SDM tools. The SDM toolbox was utilized to analyze the migration of the potential geographic distribution center of C. kiangsu under various climate scenarios31. The "Quick Re-classify to Binary" tool was employed to convert the raster data of suitable and unsuitable areas under various climatic conditions into binary data (1 and 0). The "Centroid Changes (Lines)" tool was employed to calculate the displacement of the geometric center of the predicted distribution across different periods, detecting the overall trend in the distribution area of C. kiangsu45.
Ecological niche comparison
The ecological niches of C. kiangsu in its native and invaded habitats were delineated. Initially, the ecoshot package in RStudio, combined with methodologies form used in related literature37, was utilized to compare ecological niche changes under varying conditions between the native and invaded areas37. A PCAenv analysis was conducted using bioclimatic variables associated with C. kiangsu. Subsequently, the ecospit package was used for 1000 iterations of climate ecological niche similarity testing in both directions (native ↔ invaded areas). If the observed ecological niche similarity value was significantly lower than the niche overlap value (P < 0.05), the null hypothesis of ecological niche similarity was accepted31. Finally, based on these results, the ecological niche differences of C. kiangsu were ascertained, including the extent of niche overlap and the range of overlap, comparing the degree of ecological niche overlap between its native and invaded areas31,45.
Results
Model prediction accuracy
In this study, all six species distribution models demonstrated high predictive performance, with ROG and TSS values exceeding 0.65, indicating that all models exhibited good or better performance. Specifically, the GLM model had the lowest predictive performance, with a ROC value of 0.86 and a TSS value of 0.655. The ANN, FDA, CTA models demonstrated higher predictive performance, each with a ROC value of 0.96 and a TSS value of 0.85, which were superior to the GLM model, though the performance differences among these three models were relatively small. The RF and GBM models exhibited the best prediction performance, with ROC values nearing 0.99. The TSS value for the RF model was 0.94, which is significantly higher than that of the GBM model (0.91). In summary, the SDMs constructed using the six modeling algorithms in this study showed overall good prediction accuracy, though with some differences. The RF model had the best overall performance, followed by the GBM model (0.94), and the GLM model (0.85). The RF model had the best overall performance, followed by the GBM, ANN, CTA, and FDA models, with the GLM model exhibiting the lowest predictive accuracy compared to the above models. This indicates that the ensemble model is more accurate and reliable for predicting the potential distribution of C. kiangsu in China and Southeast Asian countries.
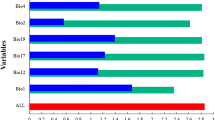
Analysis of the response curves of environmental variables
Using the Biomod2 platform, we calculated the contribution of each environmental factor to the modeling process and observed that different models utilize distinct set of factors. Furthermore, the impact of different factors on the models varies significantly. Using the optimal combination model, we investigated the environmental variables influencing the distribution of C. kiangsu (Fig. 3). The mean diurnal temperature range suitable for locust distribution in between 4 and 19 °C, with the optimal range being between 9 and 13 °C. C. kiangsu prefers areas with larger diurnal temperature variations, which are beneficial for its feeding, reproduction, and other life history activities. The maximum temperature of the warmest month suitable for locust distribution ranges from 15 to 40 °C, with the optimal range between 25 and 35 °C. Locusts can inhabit high-temperature areas due to their high biological tolerance, resulting in a wide distribution. The mean temperature of the wettest quarter suitable for C. kiangsu distribution ranges from 10 to 38 °C, with the optimal range being 21–23 °C. The mean temperature during the wettest quarter exhibits a narrower tolerance range compared to other environmental factors, making it a crucial determinant. Precipitation in the driest month shows a broader suitable range for C. kiangsu distribution, within 0–240 mm, with the highest suitability at 20 mm. In summary, the mean diurnal temperature range, maximum temperature of the warmest month, mean temperature of the wettest quarter, and precipitation of the driest month are critical factors influencing the distribution of C. kiangsu.

Response graph of environmental factors.
Current distribution patterns of C. kiangsu in Asia under current climatic conditions
Based on the model simulation results, under current climatic conditions, the suitable habitats for C. kiangsu in China and Southeast Asia are predominantly concentrated in the southern provinces of China, including Yunnan, Jiangxi, Jiangsu, Zhejiang, Fujian, Guangdong, and Guangxi. High suitability areas are primarily located in the border regions of Yunnan with Laos, as well as in the middle and lower reaches of the Yangtze River in Jiangxi, Hunan, Zhejiang, Anhui, and along the southern coast in Guangdong and Guangxi (Fig. 4). In central Anhui, northern Hubei, western Hunan, and Guangxi, less suitable areas primarily spread outward from medium to high suitability areas. Additionally, significant suitable habitats for C. kiangsu were identified in Southeast Asian countries where no prior infestation records exist. These regions include the China-Vietnam border, China-Thailand border, China-Myanmar border, central Vietnam, northern Myanmar, the Malay Peninsula, and the Indochina Peninsula. The types of suitable habitats in these regions are predominantly medium to low suitability, with a smaller proportion of highly suitable areas. In Laos, areas overrun by C. kiangsu are predominantly concentrated in the northern part of Phongsaly Province, specifically in Yot Ou County, Boun Neua County, and Phongsaly County. Field surveys in Yot Ou County, including villages such as Manla, Malitou, Songkan, Dalishu, and Huangpiao, as well as around the county town, identified 65 sites of C. kiangsu damage across 21 villages. Extensive bamboo forests in Udai Mountain near Yot Ou County were damaged, with the Yot Ou County viewing platform serving as an important migration corridor for C. kiangsu. The most severely affected area is Dalishu Township, with large contiguous bamboo forests damaged, a significant population of C. kiangsu, and close proximity to Jiangcheng County, posing a high risk of migration into the region. Next is the town of Malitou, which borders Jiangcheng County and acts as one of the corridors for C. kiangsu migrating into China. Additionally, Huangpiao Township, near the China-Laos 10–12 boundary markers adjoining Mengla County, serves as another migration corridor into China.

Geographical distribution of Ceracris kiangsu in China and Southeast Asia under current and future climatic conditions.
Prediction of suitable areas for C. kiangsu under future climatic conditions
The potential geographical distribution of C. kiangsu in China and Southeast Asia for the 2050s and 2090s under the SSP1-26 and SSP5-85 climate scenarios is illustrated in Fig. 4. A comparison of the predicted suitable habitats for the 2050s and 2090s reveals significant differences in the response of C. kiangsu to climate change under various scenarios (Table 1).
Overall, the distribution pattern of suitable habitats, centered around highly suitable areas and radiating outward, remains unchanged. Under both greenhouse gas emission scenarios in the 2050s, the total area of suitable habitats slightly decreases. However, by the 2090s, a significant increase in the total area of suitable habitats is observed, with SSP5-85 exhibiting the most pronounced expansion. The overall suitability area expands by 32.96%, from 278.4806 × 104 km2 in the 2050s to 370.2568 × 104 km2 in the 2090s.
It is evident that under the SSP1-26 climate scenario, the response of suitable habitat area C. kiangsu to climate change is less pronounced than under the SSP5-85 scenario. Under the SSP5-85 scenario, the suitable habitat area for C. kiangsu expanded more markedly. The analysis compares the changes in the suitable habitat area for C. kiangsu under different climate scenarios across various time periods. In future periods, the suitable habitat area for C. kiangsu predominantly increases (Table 2). Notably, during different periods (2050s and 2090s), the changes in expansion, loss, and stability within the suitable habitat area under the SSP1-26 scenario are more drastic compared to the SSP5-85 scenario. Under the SSP1-26 scenario, the gain area from the current period to the 2090s is more than three times that from the current period to the 2050s, while under the SSP5-85 scenario, the gain area from the current period to the 2090s is approximately half of that from the current period to the 2050s. Spatially, as shown in Fig. 5, under the SSP1-26 scenario, the most significant expansion of suitable habitat is concentrated in the southeastern part of China, including regions such as Guangxi, Guangdong, Zhejiang, Fujian. The loss areas are primarily concentrated in the southern part of China, such as northeastern Guizhou and Yunnan, while the overall trend in the Yunnan border region remains stable. Under the SSP5-85 scenario, from the current period to the 2050s, changes in the suitable habitat in southern China are more pronounced, with a larger distribution range of losses, but the gain area is greater than that in the same period under the SSP1-26. Overall, the suitable habitat for C. kiangsu changes with climate variation, and the distribution pattern of C. kiangsu under different climate scenarios across various periods consistently responds to climate change.

Spatial changes in the geographical distribution of Ceracris kiangsu under SSP1-26 and SSP5-85 Climate Scenarios in the 2050s and 2090s.
A comparative analysis of the area changes of C. kiangsu under various climate scenarios across different periods (Table 2 and Fig. 5) indicates that the area of suitable habitats for C. kiangsu predominantly increases in future periods. Notably, the changes in the area of highly suitable habitats under the SSP1-26 scenario, including expansion, loss, and stability, are more pronounced than under the SSP5-85 scenario. As shown in Fig. 5, under the SSP1-26 scenario, the most significant expansion of suitable habitats is primarily concentrated in southeastern China, such as Guangxi, Guangdong, Zhejiang, Fujian. The loss areas are primarily in southern China, such as Guizhou Province and the northeastern region of Yunnan Province, with the border areas of Yunnan generally tending towards stability. Under the SSP5-85 scenario, changes in habitat loss are more pronounced, with southern suitable habitats in China showing an overall trend of loss. Overall, the suitable habitats of C. kiangsu change with climate, and the distribution patterns of C. kiangsu under various climate scenarios across different periods consistently respond to climate change.
Center points of suitable areas
This study employs the geometric center to represent the overall spatial position of potential suitable habitats for C. kiangsu under current and future climatic conditions (Fig. 6 and Table 3). The current geometric center of potential suitable habitats for C. kiangsu is located near the Xiaoqikong scenic area in Hechi City, Guangxi Province (107.735° E, 25.2217° N). Under the SSP1-26 climate scenario in the 2050s and 2090s, the centers of distribution for suitable habitats of C. kiangsu are predicted to be in Hechi City (108.1235° E, 25.2028° N) and Liuzhou City, Guangxi (108.798° E, 25.1289° N), respectively, with the predicted geometric centers being 39.06557 km and 107.4012 km away from the current suitable area. Under the SSP5-85 scenario, the distances between the predicted geometric centers of suitable habitats in the 2050s and 2090s and the current habitat are 455.75118 km and 125.90022 km, respectively, primarily located in Hengyang City, Hunan Province (112.484° E, 26.3884° N), and Qiannan Buyei and Miao Autonomous Prefecture, Guizhou Province (107.692° E, 25.6631° N).

Migration of the center of suitable habitats for Ceracris kiangsu under current and future climatic conditions.
However, the accuracy of the future distribution center of suitable habitats for C. kiangsu across different periods may be limited, primarily due to the inherent uncertainty in simulating future climate scenarios. Under the SSP1-26 scenario, the distribution center in the 2090s exhibits significant changes, migrating northeastward by 455.75118 km, while the center for the same period under the SSP5-85 scenario remains relatively stable compared to the current period. From another perspective, the substantial migration distance of the distribution center under the SSP1-26 scenario is attributed to significant changes in the spatial distribution of non-suitable areas under future climate models, leading to increased uncertainty in the fluctuation of the distribution center due to the fragmentation of suitable habitat distribution.
Regarding migration trends, C. kiangsu exhibits varying displacement tendencies under different climate scenarios. Under the SSP1-26 scenario, the geometric center of the suitable habitat gradually shifts northeastward over time. In contrast, under the SSP5-85 scenario, the center of the suitable habitat in the 2050s tends to move eastward, but the displacement is not significant. By the 2090s, the suitable habitat shifts eastward from the current center. C. kiangsu prefers high temperature and high humidity areas, and the locations of the suitable habitat center in each period exhibit these characteristics. The high suitability areas for C. kiangsu under future climate scenarios align with the current average annual temperature conditions in China and Southeast Asia.
Ecological niche comparison
The geographical distribution areas of C. kiangsu are partially determined by its fundamental ecological niche, defined as a set of environmental conditions essential for its survival. Based on the comparison of climate ecological niche spaces between native and invaded areas, Fig. 7 illustrates the ecological niche of C. kiangsu in Asia. The figure indicates that the ecological niche overlap of C. kiangsu is substantial in each period under various climate scenarios. The ecological niche of C. kiangsu in the invaded areas is broader than in the native areas but does not entirely encompass the ecological niche of the native area. Therefore, the ecological niches of C. kiangsu in its native and invaded areas are not identical but are more similar than would be expected by chance in the invasion process.

Ecological niches of Ceracris kiangsu in the 2050s and 2090s under SSP1-26 and SSP5-85 Climate Scenarios (purple indicates ecological niche overlap, green indicates unoccupied areas, and red indicates expansion. Red arrows represent the centroids of realized ecological niches of each species).
Discussion
Potential distribution of C. kiangsu and its influencing factors
Wen11 utilized the MaxEnt model to predict the current suitable habitat distribution of C. kiangsu in China, indicating that its suitable habitats are primarily in provinces such as Hunan, Jiangxi, Yunnan, and Fujian. In this study, the current high suitability areas of C. kiangsu are primarily distributed in southern provinces such as Yunnan, and the predicted future expansion of suitable areas is mainly in southeastern coastal provinces like Guangdong and Fujian. The results of this study are consistent with those of Wen et al. and align with existing reports of C. kiangsu infestations, indicating the accuracy of the model outputs11. This study also identified suitable habitats for C. kiangsu in countries such as Laos, Thailand, Vietnam, and the Philippines, with high suitability areas primarily concentrated in northern Phongsaly Province, Laos. However, field studies by Yang et al.46 found that besides Phongsaly Province, provinces such as Luang Prabang, Houaphanh, Xiangkhouang, and Oudomxay in Laos also experienced severe infestations and crop damage. The limited distribution of suitable habitats to northern Phongsaly Province, with less emphasis on other provinces, is presumably due to missing species distribution data. In previous studies, only a single model, the MaxEnt model, was used to predict and evaluate the distribution of C. kiangsu in China11,14. In this study, we used the "Biomod2" package, an integrated platform for species distribution modeling in R, to analyze and evaluate the potential distribution areas of C. kiangsu in depth. Additionally, we expanded our study area and analyzed the changes in the current and future suitability zones of C. kiangsu in China and Southeast Asia, thereby filling the research gap regarding its potential distribution. This is of great significance for locust infestation prevention and monitoring.
Environmental principal component analysis revealed that mean diurnal range of temperature, maximum temperature of the warmest month, mean temperature of the wettest quarter, and precipitation of the driest month are crucial to the distribution of C. kiangsu. The biological characteristics of C. kiangsu indicate that June to August is a critical period for adult C. kiangsu, with the risk of water loss is exacerbated by insufficient precipitation47. C. kiangsu overwinters as eggs, and the warmer climate and milder winters create favorable climatic conditions for its safe overwintering31. The increase in global temperatures due to the massive emission of greenhouse gases directly impacts the development rate and population dynamics of C. kiangsu. The rise in temperature creates favorable conditions for C. kiangsu to hatch and reproduce. This, combined with persistent hot and dry weather during the summer, lays the groundwork for locust outbreaks47.
However, the factors influencing the distribution of C. kiangsu include not only climatic factors but also non-climatic factors. Sun confirmed that C. kiangsu often prefers to lay eggs on north-facing sunny slopes or bamboo slopes2. Li in their study on the causes and control measures of C. kiangsu outbreaks in China, affirmed that besides climate, soil moisture is a critical habitat factor affecting the occurrence of C. kiangsu, with lower soil moisture being more conducive to the survival of its eggs43. Wen et al.11 confirmed that as a host-specific folivorous pest, the host species significantly influences the growth and development of C. kiangsu11. Environmental factors directly or indirectly influence the distribution of C. kiangsu. Direct effects pertain to its suitable environmental range, while indirect effects involve environmental factors influencing plant distribution, thereby affecting its feeding and other life history aspects. Wang et al.31 indicate that under similar temperature conditions, nymphs of C. kiangsu feeding on wheat seedlings exhibit higher survival rates, shorter development times, and higher adult emergence rates compared to nymphs consuming other food sources31. Additionally, compared to glutinous maize and sweet maize, the nymphs of C. kiangsu exhibit a relatively slower decline in survival rates on wheat and rice at different temperatures. Moreover, human activities may also significant impact the gene flow of C. kiangsu. These activities stimulate the population to generate local diversity and influence its distribution27. Furthermore, the migration and spread of C. kiangsu are closely related to wind direction. Therefore, factors such as topography, slope, aspect, host plants, and wind direction are critical in influencing the distribution of C. kiangsu. In this study, only climatic factors such as temperature and precipitation were considered, without accounting for non-climatic factors. Therefore, future research should comprehensively consider the impact of these factors on the suitability of C. kiangsu. By conducting an in-depth analysis of its potential suitable habitats, the accuracy of predictions can be significantly enhanced.
Predictions from different models and ensemble modeling
In terms of the precision of model predictions, Biomod2 integrates multiple models, thus providing better predictions with fewer input distribution points21, which strongly aligns with the relevant conclusions of this study. The reasons are as follows: Firstly, we chose the current mainstream and recognized high-performance algorithms, which exhibit high robustness and generalization ability overall, providing a good strong model foundation for predicting biological species in this study. Secondly, the model integration algorithm provided by Biomod2 combines multiple models efficiently to improve prediction accuracy. It reduces errors by evaluating uncertainties and the importance of variables in the modeling process, and enhances accuracy by eliminating poorly performing single models in the integrated model48. Consequently, the overall modeling precision and the accuracy of species distribution in the study area were effectively improved.
Classical single model algorithms possess distinct advantages and disadvantages. specifically, the Maximum Entropy Model (MaxEnt) accurately reflect the response of species to environmental variables and habitat selectivity but is less precise in simulating the boundaries of species distribution40. However, integrating multiple high-performance single algorithms can effectively overcome the limitations of individual algorithms, thus enhancing the overall robustness of the model. Meanwhile, integrating multiple models can also lead to overfitting and partial defects in the selection of indicators to some extent48. Ensemble algorithms like the Generalized Additive Model (GAM) and Random Forests (RF) enhance overall prediction accuracy by increasing model complexity. However, overly complex models risk overfitting, necessitating cautious selection through methods like cross-validation21. Overall, compared to single algorithms, the Biomod2 ensemble model provides more precise and robust predictions. In terms of the overall accuracy and robustness of various SDM algorithms constructed in this study, significant difference exist between the accuracy of each single model and the prediction model integrated by multiple algorithms. The integrated model can greatly improve the accuracy and efficiency of prediction, and has higher accuracy and reliability.
The critical premise for a SDM to accurately predict species distribution points under various environmental conditions is to evaluate the reliability and accuracy of the prediction model, which is essential before making predictions49. Therefore, appropriate quantitative evaluation methods for are used to assess the accuracy and robustness of the model. It is necessary and significant to study the impact of various data and species attributes on the accuracy of the generated prediction results50. Hence, we selected the current mainstream and widely used important indicators that can evaluate the reliability of probabilistic prediction modeling for accuracy evaluation51. Specifically, this study used two common assessment metrics, AUC and TSS, to evaluate the performance of the model. The average AUC value reached 0.96, and the average TSS value was 0.8, both meeting excellent standards and demonstrating higher precision and reliability than single models like MaxEnt and GLM.
Impact of global climate change on the distribution of C. kiangsu
In recent years, global climate change and the excessive exploitation of natural resources by humans have caused ecological damage, creating favorable conditions for locust breeding and exacerbating the frequency and severity of locust plagues. According to the results of this study, from the present to the 2090s, the high suitability areas for C. kiangsu in Asia exhibit an expanding trend. In China, the high suitability areas for C. kiangsu are primarily concentrated in regions such as Yunnan, Hunan, Jiangxi, Zhejiang, and Fujian, consistent with the results of Yang et al.14, who predicted the suitable habitats of three locust species in China using the MaxEnt model. In other Asian countries, such as Laos, Vietnam, the Philippines, Thailand, and the southern Malay Peninsula, the high suitability areas for C. kiangsu appear to remain stable. Overall, compared to the present, the future habitat areas of C. kiangsu are expected to expand, particularly under the SSP5-85 scenario, with shows a greater increase. The centroid points of suitable habitats for C. kiangsu across different periods all exhibit a trend of moving northeast, and the ecological niche in the invaded areas also tends to expand further than in the native areas.
Role of SDMs in biological invasion and control of C. kiangsu
Based on the prediction results of local suitable areas in Southwest China and Southeast Asia, as well as the practice and relevant research of biological disaster prevention and control in China, Laos, and other countries, this study posits that the cross-border pest C. kiangsu on the China-Laos border and the outbreak of biohazards within a certain range and time are the results of multiple factors. These factors include the suitability of the natural environment, favorable climate conditions, social governance experience, and economic and social development. Specifically, in China, on one hand, due to the relatively complete monitoring network and prediction and evaluation system for various biological disasters, including C. kiangsu, China has a mature prediction ability for the outbreak of biological disasters. On the other hand, the control technology of C. kiangsu is relatively advanced, allowing for the effective reduction of pest density through active elimination. Regarding the incident of C. kiangsu entering China from the China-Laos border, this study posits that the surge in C. kiangsu numbers, leading to the large-scale invasion of the Chinese border, is due to the suitable environmental conditions for regular large-scale outbreaks of C. kiangsu and the imperfect biological disaster prevention and control system in Laos.
Conclusions
Ceracris kiangsu is highly responsive to climate change, particularly temperature, and the sharp increase in its population size is closely linked to these change. Understanding the current and future distribution of suitable areas for C. kiangsu is crucial for the surveillance, prevention, and control of locusts. According to the predictions of this study, under different climatic conditions, the current and future high suitability areas for C. kiangsu are primarily concentrated in the southern provinces of China such as Yunnan, Jiangxi, and Hunan, as well as in northern Phongsaly Province, Laos. Additionally, we found large numbers of suitable habitats for C. kiangsu in several Southeast Asian countries where locusts had not been previously recorded. In the coming period, the area of suitable habitat for C. kiangsu is expected to show an upward trend. In summary, given the unique geographical location of Yunnan Province, bordering countries like Laos, Vietnam, Thailand, and Myanmar, it is imperative for the Chinese border to enhance the construction of plant protection and epidemic prevention systems along the border. Additionally, establishing pest and disease monitoring and early warning systems in border areas, and strengthening the monitoring of the distribution dynamics of C. kiangsu in these regions are crucial. Proactively responding to the United Nations 2030 Sustainable Development Goals and establishing a multi-national disaster monitoring collaboration mechanism for data sharing will provide valuable references for strengthening control and risk monitoring in relevant countries and regions, contributing positively to ensuring the ecological safety of multiple countries (Supplementary Information S1).
Data availability
Data can be obtained by contacting the corresponding author.
References
Zhang, G. P. Analysis of forestry pest prevention and control technology under climate change. Chin. Vice Spec. Lin. 4, 80–81. https://doi.org/10.13268/j.cnki.fbsic.2023.04.032 (2023).
Sun, Y. C. & Guo, H. Insect response and adaptation to global climate change. Chin. J. Appl. Entomol. 54(04), 539–552 (2017).
Yao, J. et al. Effects of temperature on the development and reproduction of Soybean elephant. Acta Entomol. Sin. 59(07), 1. https://doi.org/10.16380/j.kcxb.2016.07.006 (2016).
Feng, S. Q. et al. Analysis of the route of desert locust invasion in Tibet border area in China. Plant Protect. J. 49(06), 1. https://doi.org/10.13802/j.cnki.zwbhxb.2022.2021224 (2022).
Luo, D. Biogenic analysis and molecular mechanism of high temperature tolerance in Italian and Siberian Locust under the background of climate warming. Xinjiang Normal University. https://doi.org/10.27432/d.cnki.gxsfu.2021.000673 (2021).
Han, W. J. et al. A preliminary study on the occurrence regularity of Phylloacris flavus in Yunnan Province in 2020. China Plant Protect. Guide. 42(01), 98–100 (2012).
Zhong, W. H., Lian, Y. M. & Zhang, X. K. Biological characteristics and natural enemy protection and utilization of Bamboo Locust. Hunan For. Sci. Technol. 37(05), 57–59 (2010).
Wang, Y. F. et al. Analysis of feeding characteristics of Phylloacris flavus on banana and banana. Trop. Agric. Sci. 41(07), 68–74 (2019).
Chen, S. Y. Invasion, diffusion, spread and control of Bamboo Locust. Yunnan Agric. 02, 59–61 (2022).
Cheng, J., Pan, Y. Z. & Shu, J. P. Current research status and progress of the Yellow-Spined Bamboo Locust. Shandong For. Sci. Technol. 40(02), 116–119 (2010).
Wen, X. Y. et al. Predicting suitable habitats for the yellow-spined bamboo locust in China based on the MaxEnt Model. J. Environ. Entomol. 43(06), 1427–1434 (2021).
Ding, G. H. Control measures and effectiveness of the yellow-spined Bamboo Locust in Jiangcheng County. Yunnan. Agric. Eng. Technol. 41(11), 38–39. https://doi.org/10.16815/j.cnki.11-5436/s.2021.11.022 (2021).
Ze, S. Z. et al. Occurrence and control of the yellow-spined bamboo locust in Yunnan Province in 2020. Forest Pest Dis. China. 40(04), 41–43. https://doi.org/10.19688/j.cnki.issn1671-0886.20210011 (2021).
Yang, X. L. et al. Occurrence and control of the yellow-spined Bamboo Locust in Jiangcheng Yunnan. China Plant Protect. Guide. 42(06), 53–55 (2022).
Guisan, A. & Thuiller, W. Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 8(9), 993–1009. https://doi.org/10.1111/j.1461-0248.2005.00792.x (2005).
Elith, J. & Leathwick, J. R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 40(1), 677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159 (2009).
Xiao, J. D. et al. Current status and recommendations for the control of the Yellow-Spined Bamboo Locust in Anhua County. Hunan For. Sci. Technol. 03, 37–39 (2008).
Bosso, L. et al. Integrating citizen science and spatial ecology to inform management and conservation of the Italian seahorses. Ecol. Inform. 79, 102402. https://doi.org/10.1016/j.ecoinf.2023.102402 (2024).
Abebe, H., Desta, A. B. & Dejene, S. W. Modeling the distribution of Aloe ankoberensis and A. debrana under different climate change scenarios in North Shewa Zone, Amhara National Regional State Ethiopia. Ecol. Process. 13(1), 39. https://doi.org/10.1186/s13717-024-00511-x (2024).
Araújo, M. B. & New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 22(1), 42–47. https://doi.org/10.1016/j.tree.2006.09.010 (2007).
Zhao, J. Y. et al. Analysis of potential distribution and disturbance intensity of the Plateau Pika in the Source Area of the yellow river based on the BIOMOD2 ensemble model. J. Ecol. 43(04), 1192–1201. https://doi.org/10.13292/j.1000-4890.202403.011 (2024).
Zhang, Z. Q. et al. Study on monitoring methods and control effectiveness of the yellow-spined bamboo locust in Eastern Hunan. Green Technol. 11, 6–7 (2012).
Zhou, J. G. et al. Investigation and control of pests and diseases of phyllostachys Pubescens in Guangdong. Pract. For. Technol. 7, 47–49. https://doi.org/10.13456/j.cnki.lykt.2014.07.016 (2014).
Liu, B. C. Analysis of the causes and control strategies for the severe outbreak of the yellow-spined bamboo locust in Ziyuan County. Guangxi Plant Protect. 30(04), 51–52 (2017).
Wang, W. T. Occurrence and control measures of the yellow-spined bamboo locust in Shucheng County. Mod. Agric. Sci. Technol. 12, 93–96 (2019).
Xia, Z. H. Control methods for the yellow-spined bamboo locust in Dingcheng District Changde City. For. Ecol. 07, 40–41. https://doi.org/10.13552/j.cnki.lyyst.2020.07.025 (2020).
Fan, Y. Chemical control experiment of yellow-spined bamboo locust in Huashan District Ma’anshan City. Jiangsu For. Sci. Technol. 47(05), 37–39 (2020).
Li, Z. H. & Luo, G. Practices and suggestions for prevention and control of yellow-spined bamboo locust disasters in Ning’er County. Yunnan Agric. Sci. Technol. 02, 49–50 (2021).
Ye, X. Y. & Ou, W. S. Spatial distribution and occurrence trends of yellow-spined Bamboo Locust in Nanping City. For. Survey Des. 42(04), 10–12 (2022).
Xu, J. W. & Jin, G. Causes and control recommendations for the occurrence of yellow-spined bamboo locust in Chibi. Hubei Hubei Plant Protect. 06, 64–65 (2022).
Wang, Y. et al. Prediction of potential distribution areas and priority protected areas of Agastache rugosa based on Maxent model and Marxan model. Front. Plant Sci. 14, 1200796. https://doi.org/10.3389/fpls.2023.1200796 (2023).
Xin, X. G. et al. Introduction to the BCC model and its CMIP6 experiments. Adv. Clim. Chang. Res. 15(05), 533–539 (2019).
Shi, X. et al. Climate sensitivity and feedbacks of BCC-CSM to idealized CO2 forcing from CMIP5 to CMIP6. J. Meteorol. Res. 34(4), 865–878. https://doi.org/10.1007/s13351-020-9204-9 (2020).
Graham, M. H. Confronting multicollinearity in ecological multiple regression. Ecology. 84(11), 2809–2815. https://doi.org/10.1890/02-3114 (2003).
De Marco, P. & Nóbrega, C. C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PloS One. 13(9), e0202403. https://doi.org/10.1371/journal.pone.0202403 (2018).
Feng, X. et al. A checklist for maximizing reproducibility of ecological niche models. Nat. Ecol. Evol. 3(10), 1382–1395. https://doi.org/10.1038/s41559-019-0972-5 (2019).
Xiao, Q. X., Hao, X. Z. & Rui, W. Climate change has increased the global threats posed by three ragweeds (Ambrosia L.) in the Anthropocene. Sci. Total Environ. 859(2), 9697. https://doi.org/10.1016/j.scitotenv.2022.160252 (2023).
Segurado, P. & Araujo, M. B. An evaluation of methods for modelling species distributions. J. Biogeogr. 31(10), 1555–1568. https://doi.org/10.1111/j.1365-2699.2004.01076.x (2004).
Thuiller, W. Generalized models versus classification tree analysis: A comparative study for predicting spatial distributions of plant species at different scalesJ. J. Veg. Sci. 14, 669–680 (2003).
Luo, M., Wang, H. & Lu, Z. Evaluation of Biomod2 and MaxEnt distribution prediction models using giant panda data. Chin. J. Appl. Ecol. 28(12), 4001–4006. https://doi.org/10.13287/j.1001-9332.201712.011 (2017).
Gao, M. L. et al. Study on potential distribution area of Aspen in China based on Biomod2 combination model. J. Nanjing For. Univ. (Natural Science Edition). 48(02), 247–255 (2024).
Tang, Q. H. et al. Potential threat of weeds and ragweed in farmland under global climate change and its driving factors. Chin. J. Ecol. 43(04), 1130–1140. https://doi.org/10.13292/j.1000-4890.202403.010 (2019).
Li, S. et al. Predicting the potential suitable distribution area of Emeia pseudosauteri in Zhejiang Province based on the MaxEnt model. Sci. Rep. 13(1), 1806. https://doi.org/10.1038/s41598-023-29009-w (2023).
Bi, Y. Q. et al. Distribution of suitable habitats of wild Paeonia lactiflora in China based on Biomod2 combination model. Chin. J. Tradit. Chinese Med. 47(02), 376–384. https://doi.org/10.19540/j.cnki.cjcmm.20211023.101 (2022).
Zhao, G. H. et al. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 132, 1470–2160. https://doi.org/10.1016/j.ecolind.2021.108256 (2021).
Yang, H. et al. Occurrence status and control strategies of Phyllothecus flavus in Laos. J. China Plant Protect. Guide 37(09), 50–54 (2017).
Ye, M. et al. The causes serious occurrence of jiangxi yellow ridge bamboo locust and sustainable control countermeasures. J. Jiangxi For. Sci. Technol. 3, 38–40. https://doi.org/10.16259/j.carolcarrollnki.36-1342/s.2007.03.016 (2007).
Hao, T. et al. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography. 43(4), 549–558. https://doi.org/10.1111/ecog.04890 (2020).
Barry, S. & Elith, J. Error and uncertainty in habitat models. J. Appl. Ecol. 43(3), 413–423. https://doi.org/10.1111/j.1365-2664.2006.01136.x (2006).
Shabani, F., Kumar, L., & Ahmadi, M. Assessing accuracy methods of species distribution models: AUC, specificity, sensitivity and the true skill statistic. Glob. J. Hum. Soc. Sci. B Geogr. Geo-Sci. Environ. Sci. Disaster Manag. 18(1), 1 (2018).
Ali, F. et al. Species distribution modelling of Monotheca buxifolia (Falc.) A. DC.: Present distribution and impacts of potential climate change. Heliyon. 9(2), 1. https://doi.org/10.1016/j.heliyon.2023.e13417 (2023).
Di Febbraro, M. et al. Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Glob. Chang. Biol. 29(19), 5509–5523. https://doi.org/10.1111/gcb.16901 (2023) (Epub 2023 Aug 7).
Rodriguez, C. S. et al. High potential for Brassica tournefortii spread in North American introduced range, despite highly conserved niche. Biol. Invas. 26, 337–351. https://doi.org/10.1007/s10530-023-03176-3 (2024).
Acknowledgements
I would like to express my gratitude to the Jiangcheng County Forestry and Grassland Bureau for their strong support and assistance in this research.
Funding
This research was funded by the National Natural Science Foundation of China “Multi frequency SAR polarized interferometric data for forest tree height inversion" (Grant No. 42061072); Forestry Innovation Programs of Southwest Forestry University of Key Laboratory of National Forestry and Grassland Administration on Biodiversity Conservation in Southwest China(Grant No:LXXK-2023Z06 ); and Yunnan Province Agricultural Basic Research Joint Special Project “Estimation of aboveground biomass of Simao pinus forest based on multisource remote sensing data"( Grant No. 202401BD070001-117).
Author information
Authors and Affiliations
Contributions
C.L. and G. L. designed experiments, completed data analysis, and wrote papers; L.Y. and W.Z.M. provided important help for experimental data acquisition. C.Y. Funding acquisition, guidance for research method, experimental design, and writing of the paper and C.P. paper writing. Z.L.Z, Y.S and D.Y.F assist in the development of the charts. All authors have read and agree to the published manuscript versions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, C., Luo, G., Yue, C. et al. Distribution patterns and potential suitable habitat prediction of Ceracris kiangsu (Orthoptera: Arcypteridae) under climate change- a case study of China and Southeast Asia. Sci Rep 14, 20580 (2024). https://doi.org/10.1038/s41598-024-69897-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-69897-0
