
Abstract
Despite the increase in research of hamstring stiffness through the use of ultrasound-based shear wave elastography, the active stiffness of biceps femoris long head (BFlh) and semitendinosus (ST) muscles under fatigue conditions at various contraction intensities has not been sufficiently explored. This study aimed to compare the effects of knee flexor’s isometric contraction until exhaustion performed at 20% vs. 40% of maximal voluntary isometric contraction (MVIC), on the active stiffness responses of BFlh and ST. Eighteen recreationally active males performed two experimental sessions. The knee flexors’ MVIC was assessed before the fatiguing task, which involved a submaximal isometric contraction until failure at 20% or 40% of MVIC. Active muscle stiffness of the BFlh and ST was assessed using shear wave elastography. BFlh active stiffness remained relatively unaltered at 20% of MVIC, while ST active stiffness decreased from ≅ 91% contraction time (55.79 to 44.52 kPa; p < 0.001). No intramuscular stiffness changes were noted in BFlh (36.02 to 41.36 kPa; p > 0.05) or ST (63.62 to 53.54 kPa; p > 0.05) at 40% of MVIC session. Intermuscular active stiffness at 20% of MVIC differed until 64% contraction time (p < 0.05) whereas, at 40% of MVIC, differences were observed until 33% contraction time (p < 0.05). BFlh/ST ratios were not different between intensities (20%=0.75 ± 0.24 ratio vs. 40%=0.72 ± 0.32 ratio; p > 0.05), but a steeper increase in BFlh/ST ratio was found for 20% (0.004 ± 0.003 ratio/%) compared to 40% (0.001 ± 0.003 ratio/%) of MVIC (p = 0.003). These results suggest that contraction duration could play a major role in inducing changes in hamstrings’ mechanical properties during fatigue tasks compared to contraction intensity.
Similar content being viewed by others
Introduction
Skeletal muscle stiffness during contraction (i.e., active stiffness) has been investigated in recent years through the use of ultrasound-based shear wave elastography in both fatigue and non-fatigue conditions1,2,3,4. Such measurements have been valuable to understand several physiological mechanisms5,6, as well as its relevance in sports7, clinical8, and health9 settings. Some research groups have used shear wave measurements to infer the load distribution among agonist muscles crossing a given joint10,11,12. For instance, based on the known force-stiffness relationship13, alterations of localized shear modulus within and/or between muscles for a given joint torque production are suggestive of changes in muscle forces6. Furthermore, the muscle force-stiffness relationship is known to not be modified under fatigue conditions14, unlike the relationship between muscle electrical activity and force15,16. This allows the estimation of relative changes in individual muscle forces during isometric fatiguing contractions, providing relevant information about motor control during the presence of fatigue.
When it comes to the lower-body musculature, previous studies have focused on the hamstring muscles17,18,19,20, due to their relevance in human locomotion21,22 and the high injury incidence (especially for biceps femoris long head, BFlh) in team-sports where sprinting is key23,24. Understanding the mechanical properties of the hamstrings could provide valuable information in developing effective injury prevention and rehabilitation protocols by addressing the hamstrings’ biomechanical load, strain, and coordination in different actions and joint positions25,26. Although the distribution of load sharing among hamstrings appears to be individual-specific27, different coordination patterns have been observed in hamstring-injured limbs compared to healthy28, especially in fatigue conditions3,29. In this regard, the load sharing between semitendinosus (ST) and BFlh has been shown to be dependent on the knee flexor’s isometric contraction intensity1,10. For instance, Mendes et al.1 reported that the ST displays greater active stiffness compared to BFlh during knee flexion isometric contractions at lower intensities, and the BFlh/ST active stiffness ratio increases with the contraction intensity. The same research group also found that the BFlh/ST active stiffness ratio of healthy individuals decreases during a knee flexors’ isometric contraction at 20% of maximal voluntary isometric contraction (MVIC) until exhaustion; mainly due to a drop in ST active stiffness, as BFlh active stiffness tends to remain unaltered along the task. Specifically, the ST active stiffness drop was seen to start from 40% of the time task until reaching values ≆20% below baseline at the end of the task4. Similar responses have also been observed with elite footballers3 but not with healthy males in a study from another research group that used a different fatiguing protocol (i.e., multiple short-duration isometric contractions)30. However, it is important to note that the active stiffness of ST is much higher than BFlh at 20% of MVIC of knee flexors’ isometric contraction (BFlh/ST ratio = 0.42–0.50), but not at 40% of MVIC (BFlh/ST ratio = 0.96–1.03)1. Given that when increasing the knee flexors’ isometric contraction intensity the proportion of BFlh active stiffness compared to ST is expected to increase, and the duration of the task performed until exhaustion should decrease, it is conceivable to speculate that protocols performed at 20% and 40% of MVIC would lead to different load-sharing responses between BFlh and ST. For instance, at 40% MVIC a lower drop of ST active stiffness during the task is expected to occur, although both contraction intensity protocols would lead to knee flexors’ MVIC loss.
Thus, this study aimed to further explore previous findings by comparing the effects of knee flexors’ isometric contraction until exhaustion performed at 20% vs. 40% of MVIC, on the active stiffness responses of BFlh and ST. We hypothesized that with higher knee flexor contraction intensity the drop in ST active stiffness would be lower, and both protocols would lead to similar MVIC loss.
Results
A total of 18 participants were included in the study. Data from one subject was removed from analysis due to failure to perform tests properly.
BFlh and ST passive stiffness, MVIC, fatigue index, and endurance time
Table 1 shows the MVIC and fatigue task performance and muscles’ passive stiffness outcomes. At the start of both testing sessions, BFlh and ST showed similar passive stiffness, with no muscle x session interaction (p = 0.507; η2 = 0.009), or effects for muscle (p = 0.133; η2 = 0.056) and session (p = 0.374; η2 = 0.013) factors. For the knee flexors MVIC, an effect for time was noted (p < 0.001; η2 = 0.644), but not for session (p = 0.708; η2 = 0.001) or time x session interaction (p = 0.067; η2 = 0.017). The fatigue protocols decreased MVIC to a similar extent [average MVIC loss: -18.9% (mean diff.= -30.4 Nm); p < 0.001; ES = 1.11 (0.61 to 1.60)]. However, the fatigue index was significantly higher at 20% of MVIC (mean diff.= -8.7%; p = 0.033; ES= -0.57 [-1.07 to -0.05]). As expected, endurance time at 20% of MVIC was significantly higher than at 40% of MVIC (mean diff.= 382.5 s; p < 0.001; ES = 1.43 [0.73 to 2.10]). Participants maintained torque at the established intensity, both at 20% (p > 0.05) and 40% of MVIC (p > 0.05) sessions (see Figure S1, Supplementary Information).
BFlh and ST active stiffness
Analysis showed significant muscle (p < 0.001), session (p < 0.001), and muscle x session (p = 0.041) interaction effects (see Figure S2, Supplementary Information). Overall, ST showed greater active stiffness than BFlh between 0 and 90% of contraction time (p < 0.001), while muscles had greater stiffness at 40% than 20% of MVIC in most of the contraction time (p < 0.05; see Figure S2, Supplementary Information).
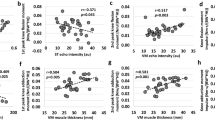
Between muscles analyses showed that ST active stiffness was higher than BFlh from the start until ≅ 64% of contraction time at 20% of MVIC (Fig. 1a and b); whereas at 40% of MVIC this difference was just noted until ≅ 33% of contraction time (Fig. 1e and f). Within muscles analyses revealed that at 20% of MVIC the ST active stiffness decreased from ≅ 91% until the end of contraction time (Fig. 1d), whereas the BFlh active stiffness remained largely unchanged for most of the contraction duration (Fig. 1c). At 40% of MVIC, no changes in both BFlh and ST active stiffness were observed throughout the full contraction time (Fig. 1g and h). Lastly, the active stiffness of BFlh at 20 and 40% of MVIC was almost similar, except at some brief instants from 42 to 45% (p = 0.003) and 46 to 48% (p = 0.017) of contraction time. ST active stiffness was similar between intensities until 72% of contraction, where the ST stiffness increased at 40% over 20% of MVIC at the last stages of the contraction (72 to 93% of contraction time; p < 0.001).

Mean active stiffness of biceps femoris long head (BFlh) and semitendinosus (ST) during the knee flexors submaximal contraction until exhaustion at (a) 20% and (e) 40% of MVIC. Panels b and f show the statistical parametric mapping (SPM) analysis, i.e., t-statistics (SPM{t}), of the differences in active stiffness levels between muscles at 20 and 40% of MVIC, respectively. The SPM analysis of the within muscles differences in active stiffness (BFlh and ST) at 20 (panels c and d) and 40% of MVIC (panels g and h) are also shown. Legend: (i) Data in panels a and e are depicted as mean (solid line) and upper and lower 95% confidence intervals (shaded areas). (ii) In panels b, c, d, f, g, and h, the pink dashed line represents the critical threshold. The darker and lighter grey shaded areas in panels b and f represent the portion of the contraction in which muscles had different active stiffness values. Alpha level was set at p ≤ 0.025 to correct for multiple comparisons. (iii) In panels c, d, g, and h, the darker and lighter gray shaded areas represent the portion of the contraction where significant differences in active stiffness compared to baseline values (i.e., the start of the contraction) were observed. Alpha level was set at p ≤ 0.05.
Regarding individual muscle slopes, significant effects were found for muscle x session interaction (p = 0.009; η2 = 0.063) and muscle (p = 0.001; η2 = 0.268), but not for session (p = 0.261; η2 = 0.022). The negative ST slope at 20% of MVIC was significantly steeper compared to BFlh at the same contraction intensity (mean diff.=0.203 kPa/%; p < 0.001; ES = 1.61 [0.57 to 2.64], but similar slopes between muscles were found at 40% of MVIC (mean diff.=0.071 kPa/%; p = 0.192; ES = 0.56 [-0.36 to 1.48]). Descriptive data of BFlh and ST slopes can be found in Table 1.
Notably, the coefficient of variation of BFlh active stiffness under the 20% (CV = 26.0 ± 3.4%) and 40% (CV = 27.4 ± 2.9%) of MVIC sessions was lower than for the ST (20% of MVIC: CV = 28.8 ± 3.0%; 40% of MVIC: CV = 37.0 ± 2.7%).
BFlh/ST ratio
Regarding the BFlh/ST ratio, no significant session effects were found (Fig. 2a and b). However, a steeper BFlh/ST ratio slope was found for 20% than 40% of MVIC (mean diff.= 0.003 ratio/%; p = 0.003; ES = 0.86 [0.29 to 1.41]) (Table 1).

Mean biceps femoris long head/semitendinosus (BFlh/ST) ratio during the knee flexors submaximal contraction until exhaustion at 20% and 40% of MVIC (panel a). Panel b shows the statistical parametric mapping (SPM) analysis, i.e., F-statistics (SPM{F}), of the session effects on BFlh/ST ratios. Legend: Data in panel a is depicted as mean (solid lines) and upper and lower 95% confidence intervals (shaded areas). The pink dashed line represents the critical threshold. Alpha level was set at p ≤ 0.05.
Discussion
This study examined the effects of a localized knee flexors’ fatigue task performed at two intensities (i.e., at 20 and 40% of MVIC) on knee flexors’ maximal strength and on the BFlh and ST active stiffness in recreationally active individuals. The main results of the current study indicated that, although a lower fatigue task time was observed in the 40% MVIC session (as expected), the ST active stiffness only decreased with fatigue task time in the 20% MVIC session, while BFlh active stiffness remained unchanged in both testing sessions. This led to a greater BFlh/ST ratio increase with fatigue task time at 20% compared to 40% of MVIC. Notably, muscles started with different active stiffness values (i.e., greater for ST) in both conditions but showed similar active stiffness from 64% to 33% contraction time in the 20% and 40% of MVIC testing sessions, respectively.
The findings of the present research also denote that both knee flexors’ fatiguing tasks resulted in reduced MVIC; however, the 20% MVIC session evoked greater knee flexors fatigue index together with load-sharing alteration between BFlh and ST, compared to the 40% MVIC condition. We speculate that the load-sharing alteration at the hamstring observed in the 20% MVIC session (but not in the 40% session) might play a role on the extent of knee flexors’ fatigue, since it is known that muscle stiffness relate to force production6,13, and ST has a high contribution for knee flexors’ torque production1,10. Such muscle stiffness response appears to require greater contraction duration, and thus contraction intensity should be adjusted to allow for a sufficient contraction duration. In this regard, significant decreases in ST active stiffness occurred from ~ 90% of contraction time during the 20% MVIC session (without changes in the 40% MVIC session). Taking into consideration the average task duration, it appears that active stiffness changes would only be expected to occur from contraction durations of 561 s onwards, which could explain why no active stiffness changes occurred in the 40% MVIC condition as its maximum duration was, on average, 153 s. This observation supports previous literature that reported no alteration of the hamstring muscles’ active stiffness following a maximal repeated sprinting task in trained footballers (i.e., 10 trials of 30 m intersped with 30 s rest), although minimal changes were observed in non-athletic individuals31. Evangelidis et al.30 recently explored the effects of 99 knee flexors isometric contractions with 5 s duration (interspersed with 5 s rest, for a total contraction duration of 494 s) at 50% of MVIC in healthy individuals on the hamstring muscles’ active stiffness. The authors found a knee flexors’ MVIC loss of 18.4% with a tendency for ST active stiffness to decrease and BFlh to increase over the fatigue task time. Thus, it would be interesting for future studies to explore the effects of a protocol with intermittent contractions at 40% of the MVIC for the same contraction time of the 20% MVIC condition but using a fatigue protocol. In this sense, it can be hypothesized that, for the same contraction duration, an equal active stiffness ST decrease response might be obtained and the BFlh increase may become evident.
Nevertheless, the reason why contraction duration significantly affects ST stiffness during the fatigue task, while BFlh stiffness appears relatively unaltered is still unknown. We speculate that a combination of morphological, histological, and neural factors could explain, at least in part, these results. For instance, BFlh has been reported to have a greater physiological cross-sectional area32 and a potentially higher proportion of oxidative fibers33 than ST which may predispose BFlh to resist fatigue (contrary to ST) and to produce higher forces with the increase in contraction intensity. Consequently, the central nervous system may increase the neural drive to other knee flexor muscles (including BFlh) to compensate for the decreased ST force with fatigue. Nonetheless, since knee-dominant exercises are thought to preferentially recruit ST over the BFlh29,34,35, it might be interesting to examine, in future research, whether the stiffness responses of ST and BFlh could be different with a combined knee flexion/hip extension fatigue task as previous research has shown higher recruitment of BFlh36.
The active stiffness responses of the ST at 20% of MVIC were found to be consistent with previous studies using a similar methodology3,4. These studies also observed a ~ 20% decrease in ST active stiffness by the end of the task, along with a tendency for BFlh active stiffness to increase. Notably, Freitas et al.3 examined a sample of professional footballers and only noted changes from the 80th percentile of the task, while Mendes et al.4 tested non-athletic individuals (as in the present study) and observed ST changes from the 40th percentile. It should be noted that in Freitas et al.3, individuals had higher knee flexors’ MVIC (i.e., 154–158 Nm) compared to individuals in Mendes et al.4 study (i.e., 123–128 Nm), as well as the individuals of the present study (i.e., 126–131 Nm). This might suggest that the training status may influence the timing where the ST active stiffness alteration occurs. Interestingly, hamstring active stiffness alterations were observed following maximal repeated sprints in non-athletic individuals but not in trained footballers31, which reinforces the importance of training status. In support of this hypothesis, we performed an additional exploratory analysis with the present study data and found a strong correlation between the percentage of ST active stiffness drop at 20% of MVIC and the individuals MVIC baseline values (p < 0.001; r = 0.771), suggesting that the ST stiffness of weaker individuals decreased in a larger degree with fatigue compared to stronger individuals, creating a large interindividual variation. However, individuals from our study maintained ST active stiffness levels even further than the sample of professional footballers of Freitas et al.3 (i.e., ∼90% contraction time vs. 80th percentile of the task, respectively), indicating higher fatigue resistance. These findings, however, should be taken with caution due to the different statistical analysis approaches employed between studies. In this regard, Pataky et al.37 found discrepancies in the statistical outcome when comparing zero- vs. one-dimensional hypothesis testing as zero-dimensional methods inadequately represent the continuous variance observed in one-dimensional trajectories, a feature commonly found in one-dimensional biomechanical datasets. Therefore, future comparisons using both statistical approaches should be made, as conclusions could be different (see Text and Figure S3, Supplementary Information to observe ANOVA outcomes).
Another interesting finding was the loss of heterogeneity of the intermuscular stiffness throughout the contraction in both the 20% and 40% of MVIC conditions, which means that at some point in the fatigue task, the active stiffness between ST and BFlh became similar. We are unaware of the relevance of this mechanical response. However, we speculate that it may affect the mechanical interaction between the BFlh and ST (i.e., two neighboring muscles) by increasing the shear stress in the connective tissue interface, which is commonly prone to injury in athletes who perform repeated sprint efforts. In this regard, interestingly, a previous study by Schuermans et al.29 reported that athletes with previous hamstring injuries had a more homogeneous pattern of T2 relaxation response (assessed using functional magnetic resonance imaging) among the hamstring muscles after a knee flexors’ fatigue protocol, compared to no previously injured individuals. Whether this aspect alters muscle mechanics during contraction and, potentially, increases the susceptibility to injury in this region warrants further investigation.
We would like to note four main study aspects to better interpret its findings. Firstly, the contraction intensities used in this study produced different mechanical and performance responses; however, they did not produce a statistically different BFlh/ST ratio at the beginning of the tasks (20%: BFlh/ST = 0.57 ± 0.22 ratio vs. 40%: BFlh/ST = 0.62 ± 0.25 ratio), which was in contrast with our initial expectation1. Although it is important to note that Mendes et al.1 used a mixed-sex sample and our sample were all males. The high ST active stiffness variability between the healthy individuals under the 40% of MVIC condition (20%: CV = 28.8 ± 3.0% vs. 40%: CV = 37.0 ± 2.7%) may explain why this difference did not reach statistical significance as similar BFlh active stiffness variability was found between the 20% (CV = 26.0 ± 3.4%) and 40% (CV = 27.4 ± 2.9%) MVIC conditions. This means that, when increasing the hamstring contraction intensity, healthy individuals may use different load-sharing strategies, which denotes high individual heterogeneity as seen in Figure S4, Supplementary Information. Nevertheless, the experimental condition provided evidence that different BFlh/ST active stiffness responses are obtained with different contraction intensities. Secondly, the present findings may not be generalized to other experimental conditions (e.g., muscle-tendon lengths, and contraction types used to induce fatigue), and we do not exclude different regional responses along the muscle length (i.e., proximal vs. distal regions) as testing was only performed in the mid-region of both muscles. Thirdly, a stress-relaxation effect could also have occurred in the ST connective tissue, particularly its distal tendon (whose length is considerably high), and thus partly explaining the decrease of the muscle belly active stiffness. Thus, the physiological mechanism underlying the ST active stiffness response with long duration knee flexor isometric contraction deserves to be explored in future research. Moreover, it should be noted that, although local monitoring was conducted to ensure individuals executed the protocol correctly, we noted (after data processing) that some individuals, in some brief instants, had torque deviations below the 5% threshold. Despite this type of behavior, which is implicit to this type of protocol, future research should make efforts to minimize this type of situation. Finally, for a complete understanding of ST and BFlh active stiffness characterization during the fatigue tasks, the full agonist (i.e., semimembranosus, biceps femoris short head, sartorius, gracilis, gastrocnemius, popliteus) and antagonist (i.e., quadriceps heads) crossing the knee joint would need to be assessed, as the torque production depends on the interaction between all muscle actuators acting on the joint38. For instance, it has been suggested that the force production among hamstring heads varies (e.g., with semimembranosus producing ~ 80% of the hamstrings’ total force at intermediate lengths39), and the interplay between these synergistic muscles could explain the high inter-individual variability found in the present study. However, this full muscle assessment is very demanding and methodologically challenging, and thus it was not possible to be performed in the present study. Therefore, we consider that these results should be interpreted with caution and future studies should further characterize the mechanical interaction of this complex muscle group, as well as the potential influence of co-activation of the antagonist muscles (i.e., greater activation of the antagonists with increased contraction intensity)40.
In conclusion, we found evidence showing that muscle contraction duration seems to have greater relevance than contraction intensity in altering the active stiffness of the hamstring muscles during fatiguing tasks, particularly in the ST. On this matter, during a submaximal isometric contraction until exhaustion, changes in stiffness appear to occur after a certain time threshold (i.e., ~ 90% of the contraction duration) in this muscle, while the BFlh remains relatively unchanged. Consequently, a steeper increase of the BFlh to ST active stiffness ratio during the fatigue task with the lower contraction intensity (i.e., 20% MVIC) was observed, which was not the case in the higher contraction intensity (i.e., 40% MVIC). Furthermore, the loss of intermuscular stiffness heterogeneity found at the last (20% of MVIC) and initial (40% of MVIC) stages of the contraction suggests a potential impact on the mechanical interaction between these hamstring muscles.
Methods
Participants
Based on a priori sample size calculation using G*Power software (version 3.1.9.3, Universität Düsseldorf) using the ANOVA: repeated measures within-between interaction test, sixteen participants were required assuming a statistical power of 80%, effect size f of 0.25, and an alpha error of 0.05. Eighteen recreationally active males (age [mean ± SD]: 27.1 ± 6.8 years; height [mean ± SD]: 176.4 ± 6.2 cm; body mass [mean ± SD]: 73.6 ± 8.3 kg) participated in this study. Participants were free from any lower limb musculoskeletal injury in the past 2 years and were asked to cease resistance or flexibility training at least 72 h before the experimental sessions. Before data collection, informed consent was obtained from participants. The study was approved by the local ethics committee (Ethics Council of Faculty of Human Kinetics—CEFMH—approval number: #11/2022) and was conducted under the principles outlined in the Declaration of Helsinki.
Experimental design
The following information provides a concise overview of the methods employed in the study. Participants took part in a randomized crossover study design in which they had to attend two experimental sessions (Fig. 3a). Sessions were separated by 3–7 days and conducted at a similar time of the day to minimize diurnal variations. Two evaluators identified the BFlh and ST muscles in the dominant limb using ultrasound and determined the muscles’ passive stiffness. Participants were then familiarized with the equipment and performed a specific warm-up of 10 submaximal contractions at 50% perceived maximal intensity. Then, the knee flexors’ MVIC of the dominant leg was assessed before the fatiguing task that involved the evaluation of the active stiffness behavior of BFlh and ST during a submaximal isometric contraction until failure at 20 or 40% of MVIC. Afterward, knee flexors MVIC was re-assessed to determine the presence of neuromuscular fatigue. The order of the experimental sessions was randomly selected.

(a) Timeline of the experimental design of the study. (b) Experimental setup used in the study to assess passive and active stiffness of biceps femoris long head (BFlh) and semitendinosus (ST) during a knee flexion isometric contraction at 20 or 40% of maximal voluntary isometric contraction (MVIC). Visual feedback of knee flexion torque production is also shown, with the red lines showing the upper and lower torque limits. Participants’ representative sonograms at the start and the end of the 20% of MVIC knee flexion submaximal isometric contraction from BFlh and ST and the elastogram scale are depicted to improve interpretation.
Procedures
Knee flexor maximal voluntary isometric contraction. Participants were positioned prone, with the hips in a neutral position (secured with a strap), and the knees flexed at ~ 30º in a custom-built equipment as shown in Fig. 3b. The foot was positioned in a foot holder at 90º ankle joint angle in a neutral position to avoid the internal/external rotation of the tibia. The foot holder contained a force transducer (Model STC, Vishay Precision, Malvern, PA) measuring at 1 kHz to collect the linear force perpendicular to the leg orientation. Force was collected (Model UA73.202, Sensor Techniques, Cowbridge, UK), amplified (gain: 1000), digitally converted (USB-230 Series, Measurement Computing Corp., Norton, MA), and recorded using the DAQami software (v.4.1, Measurement Computing Corp., Norton, MA). Knee torque was estimated by multiplying the perpendicular distance between the force transducer center and the femoral lateral condyle. Individuals performed two 3–5 s duration knee flexors’ MVICs with the dominant limb separated by a 60 s rest period. Verbal encouragement was given by the evaluators to ensure maximal effort during the test. If the second MVIC differed 5% from the preceding contraction, more repetitions were performed. The highest peak force value within the 5% difference range was used for analysis.
Fatigue task. After a 5 min rest, participants performed a sustained submaximal isometric knee flexion at 20 or 40% MVIC until failure. The test was stopped when the participant was unable to produce force or when the force produced was below 5% of the established intensity and after encouragement, the participant could not increase the force to the determined level. During the execution of the test, participants had visual feedback of the torque production as well as a visual delimited line that corresponded to their 20% or 40% knee flexors’ MVIC. The evaluators gave verbal encouragement throughout the test to maintain torque at the determined intensity.
Muscle stiffness. Two identical ultrasound systems (Aixplorer, v10; Supersonic Imagine, Aix-en-Provence, France) in B-mode, connected to a linear transducer array (SL10-2, 2–10 MHz, Vermon, Tours, France) were used to identify the location of the largest cross-sectional areas of the BFlh, and ST muscles. To find these regions, the probe was placed, first transversely and then longitudinally, following fascicle orientation at ~ 55% of the distal-to-proximal femur length and during submaximal contraction in each muscle. In regards to ST, care was taken when aligning the probe plane to not capture the ST tendon inscription41. Once the region of interest (ROI) was identified, and to ensure a stable and precise recording of muscles’ passive and active stiffness, a 3D printed plastic cast with the form of the probe was attached to the skin using bi-adhesive tape. The cast was oriented according to the fascicle direction. Then, a 60 s recording in shear wave elastography mode (musculoskeletal preset, penetrate mode, smoothing level 5, persistence off; scale: 0–800 kPa) at a frequency of approximately 1 Hz (range: 0.7–1.1 Hz, depending on the size of the selected ROI) of both muscles was taken to assess passive muscle stiffness. During the passive recording, participants were instructed to remain completely relaxed. Although muscle electromyography was not measured, researchers ensured participants’ relaxation by considering the elastogram map and knee flexor’s torque production values during the assessment. A pedal switch was used to simultaneously start data acquisition from both ultrasound systems. Active stiffness of BFlh and ST was continuously assessed during the fatigue task with the same settings used in the passive measurement. As the maximum recording time for shear wave elastography videos was 60 s, clips were taken continuously until the end of the fatigue task (less than 5 s intervals between clips due to ultrasound processing time).
Data analysis
Torque signal was filtered using a 4th order Butterworth low-pass filter with a cutoff frequency of 10 Hz. The highest peak torque obtained in pre- and post-fatigue knee flexors’ MVIC trials was considered for analysis and used to calculate the fatigue index (i.e., the percentage of MVIC torque loss after the fatigue task relative to baseline knee flexors’ MVIC). Endurance time was calculated as the total time duration from the onset of the submaximal contraction in the fatigue task until exhaustion. Torque data in the fatigue task at 20% and 40% of MVIC was normalized to the respective MVIC of the session and from 0 to 100% contraction time. Lastly, it was interpolated to match the same contraction duration (i.e., from 0 to 100% contraction time, a total of 100 data points).
Shear modulus data from the passive and active assessments were analyzed using customized Matlab® (version R2022a, The Mathworks, Inc., Natick, MA, USA) routines (script code files can be found at https://cimt.uchile.cl/mcerda/). Each video clip was extracted from the ultrasound software and converted into “.avi” format. The Matlab® routine allowed users to manually select the largest rectangular ROI in the elastogram window of the video and convert the pixels containing elastogram measurements into elastic moduli values based on the recorded scale (800 kPa). These values were then averaged to obtain a representative muscle value and exported to individual Excel files. The values displayed in the elastogram windows of the Aixplorer’s ultrasound scanner represent Young’s modulus (E) of the medium measured as: E ≈ 3µ, being (µ) the shear modulus. Therefore, to quantify the shear moduli of the muscles, the values were divided by 3 following the equation: µ = E/3 42. The muscle active stiffness during the initial 5 s after the onset of muscle contraction was discarded from analysis, in order to exclude the period in which the individual adapted to reach the target contraction intensity with a stable elastogram. Customized Matlab® (version R2022a, The Mathworks, Inc., Natick, MA, USA) routines were then used to: (1) detect and remove outliers using the interquartile method; (2) replace missing values (caused by ultrasounds’ processing time between clips) by fitting a 4th order polynomial function; (3) smooth the signal applying a Savitzky-Golay filter; (4) normalize the time series from 0 to 100% contraction time; and (5) interpolate the individuals’ data to match the same contraction duration (i.e., from 0 to 100% contraction time, a total of 100 data points). BFlh/ST ratio values for each data point during the fatigue task were determined, as well as the individuals’ slope of the linear contraction duration-BFlh/ST and ratio relationship, was considered for analysis. The BFlh/ST ratio was calculated by dividing the BFlh active stiffness values at each percentage of the contraction duration with the corresponding ST stiffness values. These variables were selected as indicative of potential load-sharing alterations. Ultimately, to examine the magnitude of inter-individual responses in each muscle and testing condition, the coefficient of variation (CV) of the active stiffness responses of BFlh and ST in the different conditions was calculated.
Statistical analysis
Descriptive data are presented as mean ± SD. Normal distribution of the data was confirmed using the Shapiro-Wilk test. A two-way repeated measures ANOVA [muscle (BFlh, ST) x session (20%, 40%)] was used to analyze muscles’ passive shear modulus and slopes, and knee flexors MVIC [time (PRE, POST) x session (20%, 40%)]. If significant effects were found, a post hoc multiple comparisons test with Holm’s correction was performed. To verify if torque was maintained at the determined intensity levels (i.e., 20% and 40% of MVIC) during the fatigue task, statistical parametric mapping (SPM) one-sample two-tailed t-tests were performed, with 20 and 40% of MVIC as criterion values, respectively.
A two-way repeated measures SPM ANOVA [muscle (BFlh, ST) x session (20%, 40%)] was performed to analyze active stiffness throughout the submaximal contraction. When a significant effect was found for muscle, session, or session x muscle interaction, post hoc SPM t-tests were performed to identify the direction of the differences. SPM paired samples two-tailed t-tests were used to assess between muscle active stiffness differences. Statistical significance was set to p ≤ 0.025 to correct for multiple comparisons accordingly. Then, to analyze within muscle active stiffness changes throughout the contraction at the different sessions, SPM one-sample two-tailed t-tests were performed, with the starting average stiffness values used as baseline comparisons. Furthermore, a one-way repeated measures SPM ANOVA was used to analyze BFlh/ST ratio differences at each contraction intensity.
Lastly, a paired sample t-test was used to analyze the differences in fatigue index, and the BFlh/ST ratio slopes between sessions. Wilcoxon signed-rank test was used to analyze endurance time (as data was non-normal distributed). Eta squared was calculated from the repeated measures ANOVA and categorized as small (0.01–0.06), moderate (0.06–0.14), and large (> 0.14)43. Cohen’s d effect sizes (ES) and its 95% confidence interval were also calculated to describe the standardized effects as follows: trivial (< 0.2), small (0.2–0.59), moderate (0.6–1.19), large (1.2–1.99), very large (2–4), and near perfect (> 4)44. The alpha level was set at p ≤ 0.05. Statistical analysis of discrete variables was performed using JASP (version 0.17.1.0) whereas the analysis of continuous variables was performed in Matlab® (version R2022a, The Mathworks, Inc., Natick, MA, USA) by using the open source SPM code (version M.0.4.10, SPM1D open-source package, spm1d.org)45.
Data availability
The datasets generated analyzed during the current study are available in the figshare repository, https://doi.org/10.6084/m9.figshare.24865479.
References
Mendes, B. et al. Hamstring stiffness pattern during contraction in healthy individuals: Analysis by ultrasound-based shear wave elastography. Eur. J. Appl. Physiol. 118, 2403–2415 (2018).
Bouillard, K., Jubeau, M., Nordez, A. & Hug, F. Effect of vastus lateralis fatigue on load sharing between quadriceps femoris muscles during isometric knee extensions. J. Neurophysiol. 111, 768–776 (2014).
Freitas, S. R. et al. Semitendinosus and biceps femoris long head active stiffness response until failure in professional footballers with vs. without previous hamstring injury. EJSS. 22, 1132–1140 (2022).
Mendes, B. et al. Effects of knee flexor submaximal isometric contraction until exhaustion on semitendinosus and biceps femoris long head shear modulus in healthy individuals. Sci. Rep. 10, 16433 (2020).
Chalchat, E. et al. Muscle shear elastic modulus provides an indication of the protection conferred by the repeated bout effect. Front. Physiol. 13, 877485 (2022).
Bouillard, K., Nordez, A. & Hug, F. Estimation of individual muscle force using elastography. PLoS One. 6, e29261 (2011).
Romer, C. et al. Stiffness of muscles and tendons of the lower limb of professional and semiprofessional athletes using shear wave elastography. J. Ultrasound Med. 41, 3061–3068 (2022).
Davis, L. C., Baumer, T. G., Bey, M. J. & van Holsbeeck, M. Clinical utilization of shear wave elastography in the musculoskeletal system. Ultrasonography (Seoul Korea). 38, 2–12 (2019).
Kim, H. J. et al. Correlation of shear-wave elastography parameters with the molecular subtype and axillary lymph node status in breast cancer. Clin. Imaging. 101, 190–199 (2023).
Evangelidis, P. E. et al. Hamstrings load bearing in different contraction types and intensities: A shear-wave and B-mode ultrasonographic study. PLoS One. 16, e0251939 (2021).
Avrillon, S., Hug, F. & Guilhem, G. Between-muscle differences in coactivation assessed using elastography. J. Electromyogr. Kinesiol. 43, 88–94 (2018).
Hug, F., Tucker, K., Gennisson, J. L., Tanter, M. & Nordez, A. Elastography for muscle biomechanics: Toward the estimation of individual muscle force. Exerc. Sport Sci. Rev. 43, 125–133 (2015).
Ettema, G. J. C. & Huijing, P. A. Skeletal muscle stiffness in static and dynamic contractions. J. Biomech. 27, 1361–1368 (1994).
Bouillard, K., Hug, F., Guével, A. & Nordez, A. Shear elastic modulus can be used to estimate an index of individual muscle force during a submaximal isometric fatiguing contraction. J. Appl. Physiol. 113, 1353–1361 (2012).
Dideriksen, J. L., Enoka, R. M. & Farina, D. Neuromuscular adjustments that constrain submaximal EMG amplitude at task failure of sustained isometric contractions. J. Appl. Physiol. 111, 485–494 (2011).
Edwards, R. G. & Lippold, O. C. The relation between force and integrated electrical activity in fatigued muscle. J. Physiol. 132, 677–681 (1956).
Zhi, L., Miyamoto, N. & Naito, H. Passive muscle stiffness of biceps Femoris is acutely reduced after eccentric knee flexion. J. Sports Sci. Med. 21, 487–492 (2022).
Miyamoto, N. & Hirata, K. Site-specific features of active muscle stiffness and proximal aponeurosis strain in biceps femoris long head. Scand. J. Med. Sci. Sports. https://doi.org/10.1111/sms.13973 (2021).
Goreau, V. et al. Hamstring muscle activation strategies during eccentric contractions is related to the distribution of muscle damage. Scand. J. Med. Sci. Sports. https://doi.org/10.1111/sms.14191 (2022).
Kositsky, A. et al. In vivo assessment of the passive stretching response of the bicompartmental human semitendinosus muscle using shear-wave elastography. J. Appl. Physiol. 132, 438–447 (2022).
Thelen, D. G., Lenz, A. L., Francis, C., Lenhart, R. L. & Hernández, A. Empirical assessment of dynamic hamstring function during human walking. J. Biomech. 46, 1255–1261 (2013).
Howard, R. M., Conway, R. & Harrison, A. J. Muscle activity in sprinting: A review. Sports Biomech. 17, 1–17 (2018).
Maniar, N. et al. Incidence and prevalence of hamstring injuries in field-based team sports: A systematic review and meta-analysis of 5952 injuries from over 7 million exposure hours. Br. J. Sports Med. (2022). bjsports–2021–104936.
Ekstrand, J. et al. Hamstring injury rates have increased during recent seasons and now constitute 24% of all injuries in men’s professional football: The UEFA Elite Club Injury Study from 2001/02 to 2021/22. Br. J. Sports Med. (2022). bjsports–2021–105407.
Li, C. & Liu, Y. Regional differences in behaviors of fascicle and tendinous tissue of the biceps femoris long head during hamstring exercises. J. Electromyogr. Kinesiol. 72, 102812 (2023).
Schache, A. G., Dorn, T. W., Blanch, P. D., Brown, N. A. T. & Pandy, M. G. Mechanics of the human hamstring muscles during sprinting. Med. Sci. Sports Exerc. 44, 647–658 (2012).
Avrillon, S., Guilhem, G., Barthelemy, A. & Hug, F. Coordination of hamstrings is individual specific and is related to motor performance. J. Appl. Physiol. 125, 1069–1079 (2018).
Avrillon, S., Hug, F. & Guilhem, G. Bilateral differences in hamstring coordination in previously injured elite athletes. J. Appl. Physiol. 128, 688–697 (2020).
Schuermans, J., Van Tiggelen, D., Danneels, L. & Witvrouw, E. Biceps femoris and semitendinosus–teammates or competitors? New insights into hamstring injury mechanisms in male football players: A muscle functional MRI study. Br. J. Sports Med. 48, 1599–1606 (2014).
Evangelidis, P. E. et al. Fatigue-induced changes in hamstrings’ active muscle stiffness: Effect of contraction type and implications for strain injuries. Eur. J. Appl. Physiol. https://doi.org/10.1007/s00421-022-05104-0 (2022).
Freitas, S. R., Radaelli, R., Oliveira, R. & Vaz, J. R. Hamstring stiffness and strength responses to repeated sprints in Healthy nonathletes and Soccer players with Versus without previous Injury. Sports Health. 15, 824–834 (2023).
Lieber, R. L. & Fridén, J. Functional and clinical significance of skeletal muscle architecture. Muscle Nerve. 23, 1647–1666 (2000).
Evangelidis, P. E. et al. The functional significance of hamstrings composition: is it really a ‘fast’ muscle group? Scand. J. Med. Sci. Sports. 27, 1181–1189 (2017).
Hegyi, A., Csala, D., Péter, A., Finni, T. & Cronin, N. J. High-density electromyography activity in various hamstring exercises. Scand. J. Med. Sci. Sports. 29, 34–43 (2019).
Bourne, M. N. et al. Impact of exercise selection on hamstring muscle activation. Br. J. Sports Med. 51, 1021–1028 (2017).
Hegyi, A. et al. Superimposing hip extension on knee flexion evokes higher activation in biceps femoris than knee flexion alone. J. Electromyogr. Kinesiol. 58, 102541 (2021).
Pataky, T. C., Vanrenterghem, J. & Robinson, M. A. Zero- vs. one-dimensional, Parametric vs. non-parametric, and confidence interval vs. hypothesis testing procedures in one-dimensional biomechanical trajectory analysis. J. Biomech. 48, 1277–1285 (2015).
Latash, M. L. Biomechanics as a window into the neural control of movement. J. Hum. Kinet. 52, 7–20 (2016).
Kellis, E., Galanis, N., Kapetanos, G. & Natsis, K. Architectural differences between the hamstring muscles. J. Electromyogr. Kinesiol. 22, 520–526 (2012).
Wu, R., Delahunt, E., Ditroilo, M., Lowery, M. M. & DE Vito, G. Effect of knee joint angle and contraction intensity on hamstrings coactivation. Med. Sci. Sports Exerc. 49, 1668–1676 (2017).
Watanabe, K., Otsuki, S., Hisa, T. & Nagaoka, M. Functional difference between the proximal and distal compartments of the semitendinosus muscle. J. Phys. Therapy Sci. 28, 1511–1517 (2016).
Bercoff, J., Tanter, M. & Fink, M. Supersonic shear imaging: A new technique for soft tissue elasticity mapping. IEEE Trans. Ultrason. Ferroelectr. Freq. Control. 51, 396–409 (2004).
Cohen, J. Statistical Power Analysis for the Behavioral Sciences (Routledge, 2013). https://doi.org/10.4324/9780203771587
Hopkins, W. G., Marshall, S. W., Batterham, A. M. & Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 41, 3–13 (2009).
Pataky, T. C. Generalized n-dimensional biomechanical field analysis using statistical parametric mapping. J. Biomech. 43, 1976–1982 (2010).
Acknowledgements
This study was founded by Fundación Séneca (FS), Grant number: 22134/PI/22 (to AMS, TTF, and PEA) and MEC | Fundação para a Ciência e a Tecnologia (FCT), Grant number: PTDC/ SAU-DES/31497/2017 (to RR and SRF). The authors would like to thank Sport Lisboa e Benfica for lending an ultrasound unit, and Professor Carlos Cruz-Montecinos for helping us conduct the statistical parametric mapping analysis.
Author information
Authors and Affiliations
Contributions
Conceived and designed research (SRF, RR, AMS), performed experiments (SRF, RR, AMS), analyzed data (AMS), interpreted results of experiments (SRF, AMS), prepared figures (AMS), drafted manuscript (AMS, SRF, RR, TTF, PEA), edited and revised manuscript (AMS, SRF, RR, TTF, PEA), and approved final version of the manuscript (SRF, RR, TTF, PEA).
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Martínez-Serrano, A., Radaelli, R., Freitas, T.T. et al. Changes in hamstrings’ active stiffness during fatigue tasks are modulated by contraction duration rather than intensity. Sci Rep 14, 24873 (2024). https://doi.org/10.1038/s41598-024-75032-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-75032-w
