
Abstract
Background and Aims Cotton-peanut rotation is a sustainable farming practice that enhances land utilization and promotes the sustainable development of agriculture. Crop rotation can reduce the occurrence of pests and diseases, as different crops have varying levels of resistance to such threats. Additionally, by alternating the types of crops grown, the soil environment is changed, which can lead to the elimination of favorable conditions for pathogens and pests, thereby alleviating the impact of these issues. Furthermore, cotton-peanut rotation can improve soil fertility.To investigate the effects of different crop rotation systems on crop yield, soil nutrients, and soil microbial communities. Methods: Using high-throughput sequencing technology, investigate the soil microbial diversity in the root zone after cotton-peanut rotation.Various planting patterns, including cotton continuous cropping (MC), peanut continuous cropping (HC), peanut-cotton-peanut rotation (HR), and fallow (X), were established to assess variations in crop yield, soil nutrients, and soil microbial diversity. Results: Significant differences were observed in crop yield, soil nutrients, and soil microbial community structure among different planting patterns. The HR system significantly increased the output compared with the HC and MC systems. Additionally, HR exhibited significantly lower total nitrogen (N) and basic nitrogen (BN) contents than HC and MC, whereas MC showed lower total potassium (K) and available potassium (AK) contents. HR led to a decrease in soil bacterial diversity but an increase in fungal diversity, with Ascomycota and Mortierellomycota being dominant. Various bacteria (Chloroflexi, Bacteroidota, and Actinobacteriota) associated with organic matter degradation and nutrient cycling were found across different planting systems, enhancing material cycling efficiency. Furthermore, Planctomycetota bacteria related to crop nutrient synthesis and Glomeromycota bacteria aiding plant nutrient absorption were significantly higher in the MC system than in the HR or HC systems. Redundancy analysis indicated a significant negative correlation between crop rotation and soil fungal community, whereas Ascomycota exhibited a significant negative correlation with organic matter.Conclusion: Peanut-cotton rotation can mitigate soil nutrient loss, enhance beneficial microorganism diversity, suppress harmful bacterial populations, stabilize ecosystems, and boost crop yield.
Similar content being viewed by others

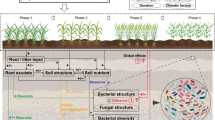

Introduction
Xinjiang, the largest raw cotton(Gossypium spp.)-producing area in China, covering a sowing area of 2.4969 × 106 hm21, contributing to 85% of the total cotton output of China2. Due to year-round cotton cultivation, soil fertility has decreased, and soil-borne diseases have increased3,4. Additionally, peanuts (Arachis hypogaea L.) represent a significant economic crop in China5. The sown area of peanuts in China can reach 4.62 × 106 hm26,7. Xinjiang demonstrates abundant light and heat resources, with annual sunshine hours ranging from 2800 to 3500 h. The region experiences prolonged sunshine, a dry climate, and a low incidence of pests and diseases, which are all favorable conditions for peanut cultivation8,9. However, continuous peanut cropping has severely compromised soil quality and physical and chemical properties, reducing the crop yields and the diversity of soil microbial communities10,11. Therefore, the adoption of peanut cotton crop rotations has attracted considerable attention. Previous studies have explored soybeans and corns12, peanuts-corn-soybeans13, corn-peanuts14, soybeans-wheat15system, corn-wheat16,17, and leguminous-potato18, have clarified that crop rotation can enhance crop yields, practices can mitigate soil nutrient loss.Furthermore, after the crop rotation, there can be a decrease in soil-borne diseases19,20, primarily attributed to the alteration in soil microbial community structure and diversity21,22,23,24. Soil microorganisms play a crucial role in material and energy cycles25,26. The impact on crop growth and development can be categorized into two types: pathogenic bacteria and beneficial bacteria27. Research has indicated that soil microorganisms can decompose soil nutrients, thereby facilitating nutrient absorption by crops. Compared with continuous cropping, crop rotation significantly affects the soil microbial community structure28,29,30, and can be more conducive to maintain microorganism diversity31,32Enhance the microbial decomposition of organic matter in the soil, promote crop nutrient uptake, and increase the utilization efficiency of soil nutrients, in order to address the question. Uzoh et al. observed a significant reduction in beneficial soil microorganisms due to continuous corn and soybean cropping33, whereas research by Zhang Lili et al. have indicated that rotations such as corn-soybean and wheat-corn-soybean can alter the composition of the soil microbial community, increase the abundance of rhizosphere soil bacteria and beneficial fungi, such as nitrogen-fixing bacteria34, and can enhance the ACE, Shannon, and Simpson diversity indices of the rhizosphere soil microbial community35,36.Meriles et al. observed a notable increase in the Trichoderma group within the peanut-corn-soybean rotation system13,37. Li et al. discovered that compared with continuous cropping, soybean rotation significantly enhanced the bacterial and microbial communities in the root system38, with Acidobacteria dominating in continuously cropped soil. After the rotation with corn, the soil microbial community diversity and richness at the phylum level can be changed39,40, reducing fungal community diversity and increasing bacterial diversity41,42,43, particularly dominated by Proteobacteria. Simpson et al. demonstrated that crop rotation planting patterns significantly affected the physical and chemical properties44,45,46. In summary, crop rotation can enhance crop yields, soil nutrients, and the structural diversity of soil microbial communities. After the cotton-peanut rotation, changes in soil physical and chemical properties are essential for improving the soil fertility of continuous cotton fields. The alterations in soil microbial structure and diversity are key factors in alleviating the obstacles associated with cotton field succession. Research and analysis of the changes in soil physical and chemical properties and soil microbial community structure after cotton-peanut rotation have theoretical and practical significance for reducing the occurrence of crop diseases and pests, and for enhancing crop yield and quality under the cotton-peanut rotation cropping system.
With the continuous development of sequencing technology, high-throughput sequencing has become pivotal for studying microbial community composition and structure47,48,49. This technology has promoted the rapid identification of soil microorganisms and provides precise structural information50,51,52,53. In summary, although there are many publications on the effects of crop rotation on crop yields, reports on the impact of cotton-peanut rotation on crop yield, soil physical and chemical properties, and soil rhizosphere microbial structure remains limited. Therefore, this study employed high-throughput sequencing technology and conducted long-term positioning experiments to analyze the changes in crop yields, soil physicochemical properties, and soil rhizosphere microorganisms across cotton-peanut continuous cropping, rotation, and fallow systems. This study aimed to elucidate the effects of cotton-peanut rotation on crop yield, soil physicochemical properties, and soil microbial community structure, thereby contributing to the theoretical foundation for scientific planting practices.
Materials and methods
Overview of the test area
The field experiment was conducted at the Anning Qu Comprehensive Test Site of the Xinjiang Academy of Agricultural Sciences (87° 28’ E, 43° 56’ N, H: 800 m) in Urumqi, Xinjiang, northwest China. This region had a temperate continental climate, characterized by an average annual temperature of 7.5℃. During July and August, the average temperature can reach 25.7℃, indicating high temperatures during this period. The annual rainfall averages 194 mm, and the soil type is arid soils. For spring planting, 1.2 × 104 kg‧hm− 2 of farm fertilizer and 450 kg·hm− 2 of (NH4)2HPO4 were adopted as base fertilizers. According to the local fertilizer recommendations, the cotton plot received 330 kg·hm− 2 of urea, and the peanut plot received 120 kg·hm− 2 of urea (Supplementary Experimental Land Capacity).
Experimental design
In this experiment, cotton variety Xinluzao 50 and peanut variety Huayu 9610 were selected from the Xinjiang agricultural germplasm resource system. The trial crops were cultivated in the Anningqu Comprehensive Test Field of the Xinjiang Academy of Agricultural Sciences from May 2016 to May 2021. Mulch cultivation and subfilm irrigation techniques were employed. The sowing and field management followed the practices established in the local high-yield fields. Annual or rotation cycle fertilizer application remained consistent across the various rotation patterns and treatments. Four treatments were designed according to the research content and objectives of the experiment, including cotton-peanut rotation (HR), cotton continuous cropping (MC), peanut continuous cropping (HC), and fallow (X) as the control treatments, with three replicates for each treatment, using a randomized block design. Each cell measured 12 m × 15 m, totaling an area of 180 m2, with 0.5 m spacing between cells, 1 m spacing between repeats, and 1 m protection lines surrounding the treatment area (Fig. 1).

Field layout of cotton peanut rotation.
Soil sample collection, analysis, and determination
Sample collection and yield data collection
Soil samples were collected after the harvest on September 28, 2021, utilizing the “5-point sampling method” to obtain samples for each treatment. Samples were collected from rhizosphere soil and the impurities were removed. The collected soil samples were thoroughly mixed, divided into 50mL sampling tubes, temporarily stored in dry ice, and promptly transported back to the laboratory, which were stored at − 80 °C for the soil microbial detection. The remaining portion was rapidly dried, ground, and shifted for determination of soil physical and chemical properties. Yield data collection involves measuring the total weight of different planting pattern trial fields at the harvesting stage and recording the data finally.
Analysis and determination of soil physical and chemical properties
The soil physical and chemical properties were determined using Soil Agrochemical Analysis54. Basic nitrogen(BN) and Total nitrogen (TN) were assessed using the Kjeldahl method. Total phosphorus (TP) was determined using the HClO4-H2SO4 cooking method. Total potassium (TK) was analyzed using the NaOH melting and flame photometer method (GB-7854-87). Organic matter (OM) content was measured using the potassium dichromate volumetric method. Available phosphorus (AP) was determined employing 0.5 mol‧L− 1 NaHCO3, and available potassium (AK) was assessed through NH4OAc extraction followed by the flame photometer method.
DNA extraction, PCR amplification, and metagenomic sequencing
DNA extraction
The genomic DNA was extracted from the soil samples and processed using the CTAB method. After the extraction, the purity and concentration of the DNA were assessed using 1% agarose gel electrophoresis. An appropriate quantity of genomic DNA was then aspirated, stored in a centrifuge tube, and subsequently diluted to 1ng‧µL− 1 with sterile water for future applications (Table 1).
PCR amplification
Employing the diluted genomic DNA as the template, the specific primers with Barcode, including 515 F and 806R, were utilized to amplify the V3–V4 hypervariable region of the bacterial 16 S rRNA gene, based on the selection of the target region for sequencing. For fungal ITS gene amplification, the ITS5-1737 F, ITS3-2024 F, ITS2-2043R, and ITS4-2409R primers were applied, testing ITS1 and ITS2 regions. PCR was performed using Phusion® High-Fidelity PCR Master Mix with GC Buffer and a high-efficiency enzyme (New England Biolabs) to ensure accurate and efficient amplification. The PCR reaction system for operation is presented in Supplementary Tables 1 and Table 2.
PCR product purification and metabarcoding sequencing
Equal concentrations of the samples were combined based on the concentration of PCR products and purified using 1×TAE 2% agarose gel electrophoresis. The sequences within the main band size range of 400–450 bp were selected, and the target bands were retrieved by excision with a rubber cutter. The product purification kit utilized was the Thermo Scientific GeneJET glue recovery kit. Appointing Beijing Novogene Co., Ltd. to construct libraries and perform sequencing using the NovaSeq sequencing platform.
Sequence data and bioinformatics analysis
OTU clustering and species annotation
The Uparse algorithm (Uparse v7.0.1001, http://www.drive5.com/uparse/) was applied to all samples to effectively cluster tags. The sequences were clustered into Operational Taxonomic Units (OTUs) with 97% identity, and representative sequences for the OTUs were selected. Following the algorithm principle, the sequences with the highest frequency within the OTUs were selected as the representative sequences of the OTUs. Species annotation of the OTU sequences was conducted using the BLAST method with Qiime software (Version 1.9.1) (http://qiime.org/scripts/assign_taxonomy.html) and the Unite database (https://unite.ut.ee/). The community composition analysis was performed at each classification level (kingdom, phylum, class, family, genus, and species) for each sample.
Sample complexity analysis (Alpha Diversity) and principal coordinate analysis
Qiime software (Version 1.9.1) was used to calculate α diversity indices, including the Chao1 index, Shannon index, ACE index, and species accumulation curves. Tukey’s test was applied for statistical analysis. Graphing conducted using the R software.And utilize principal coordinate analysis to evaluate the target.
Correlation analysis between soil microbial communities and soil nutrients
Data organization and calculations were conducted using Excel software. The data in the chart were presented as the mean ± standard deviation. The Pearson correlation coefficient method was employed to analyze the correlation between the physical and chemical properties of the rhizosphere soil and the composition of the rhizosphere soil microbial communities. Spearman’s correlation between species and environmental factors was calculated using the “corr.test” function of R psych, with significance testing. The redundancy analysis (RDA) between soil physical and chemical properties and rhizosphere soil microbial communities was performed using the R software. All results were plotted using R software.
The functional annotation methods of FUNGuild and FAPROTAX
The analysis mainly focuses on the phylum level of microorganisms, using the following methods.FUNGuild fungal functional annotation method: Collect fungal ITS sequence data and obtain relevant ecological function information from related databases; align the collected fungal ITS sequences to generate a matrix file containing all sequence information; use FUNGuild software to predict the functions of the matrix file, and generate a fungal functional annotation table based on the prediction results; perform statistical analysis on the fungal functional annotation table to understand the functional diversity of the fungal community, providing a reference for further research.
FAPROTAX functional annotation method: Collect bacterial ITS sequence data and obtain relevant ecological function information from related databases; align the collected bacterial ITS sequences to generate a matrix file containing all sequence information; use FAPROTAX software to annotate the functions of the matrix file, and the software will display the prediction results in a visual chart.In the research process, the combined analysis of ITS1 and ITS2 is conducted to summarize the taxonomic门 level. This integration can provide a more comprehensive genetic sequence information, which aids in a more accurate understanding of the relationships between species. Although1 and ITS2 have overlapping sequences, they each possess unique sequence characteristics and patterns of variation. The combined analysis can complement the information between the two, resulting in more thorough and reliable analysis outcomes. By combining the analyses the errors introduced by a single data source can be reduced. The combined analysis can enhance resolution, allowing for more precise differentiation of these relationships, and to some extent, compensate for data loss, providing a more complete genetic information. This improves the accuracy and reliability of the analysis, thereby better addressing the research questions in the study.
Results and analysis
Effects of cotton-peanut rotation on soil nutrients
The cotton-peanut rotation significantly affected the soil nutrients (Table 1). The physical and chemical properties of the soil were assessed after the harvest in 2021. Under the MC system, soil N and BN contents were notably higher than the HC and HR systems, with the BN content reaching the highest level (196.72 mg‧g− 1), a 42.37% increase over the HR system. Additionally, the soil K content in the HC system (10.38 mg‧g− 1) was significantly higher than that in the HR and MC systems. Furthermore, the AK content in the HC system (233.88 mg‧g− 1) surpassed that in the HR system (194.96 mg‧g− 1) and MC system (170.51 mg‧g− 1). The soil AP content was the lowest in the HC, HR, and MC systems, whereas the P content did not exhibit significant differences across planting systems. The soil OM content varied across planting systems: 11.00 mg‧g− 1 in X system, 34.26 mg‧g− 1 in MC system, 20.54 mg‧g− 1 in HC system, and 31.68 mg‧g− 1 in HR system. Notably, after the harvest in 2021, N and BN contents significantly increased compared with pre-sowing data of physical and chemical properties of overwintering soil from 2020 to 2021. The AK, AP, and K contents were more depleted during crop growth. Compared to the HR and HC systems, there was a noticeable difference in the metabolic consumption of AK in the MC system. The OM content increased in the MC and HC systems, while the consumption of HR decreased. However, P had no significant variation across different planting patterns. The physical and chemical properties of the X system remained consistent before sowing in 2021 compared with that after the harvest in 2021.
In summary, this study indicated that soil physical and chemical properties remained stable in the X system, promoting soil self-recovery. The physical and chemical properties of the soil were significantly affected during the crop growth process in the HC and MC systems. Cotton exhibited a preference for K, with substantial AK consumption in the MC system soil. This preference was due to its insensitivity to BN, which consequently contributed to a significant accumulation of BN. The consumption of AK in the HC system was not significantly different from that in the MC system. Hence, cotton-peanut rotation and varying crop planting across years can improve soil physical and chemical properties while reducing the reliance on K and N fertilizers.
Effects of cotton-peanut rotation on crop yield
The results presented significant variations in crop yields owing to the changes in planting patterns (Table 2). Compared to continuous cropping, systemin the rotation system, cotton and peanut yields exhibited consistent annual increases with notable differences. Under the HR system, cotton yields in 2017, 2019, and 2021 were 406.07 kg, 415.84 kg, and 465.47 kg, respectively, obtaining a 14.6% increase. Similarly, peanut yields per mu in 2016, 2018, and 2020 were 405.71 kg, 444.32 kg, and 448.67 kg, respectively, reflecting a 10.6% increase. The rotational production of cotton and peanuts demonstrated consistent year-on-year yield increments. Compared to 2017, cotton output increased by 59.40 kg in 2021. Under the impact of a three-year rotation, peanut production increased by 42.96 kg compared to 2016.
Effects of different planting patterns on soil fungal community composition
The PCoA revealed that the PCoA1 and PCoA2 accounted for 28.32% and 18.15%, respectively, in the first and second principal component analyses. In each planting system, sample aggregation was notably high, with discernible differences observed among HR system, HC system, MC system, and X system (Fig. 2A). The fungal community structure dispersion was significant in the HR and HC systems, with notable differences. The fungal community structure of the X system differed significantly from that of the other three planting systems. Additionally, the MC system exhibited significant differences compared with the HC and X systems, without evident overlap. The correlation coefficient between HR: HC and HC: X is small, indicating a weak correlation (Table 3).
The microbial composition of soil rhizosphere fungi across different planting patterns was analyzed, revealing significant effects at the phylum level (Fig. 2B). Basidiobolomycota, Zoopagomycota, Kickxellomycota, Rozellomycota, Glomeromycota, Olpidiomycota, Basidiomycota, Chytridiomycota, Ascomycota and Mortierellomycetes were the most abundant in all fungal communities, collectively constituting approximately 80% of the top 10 phyla. In the 0–20 cm soil layer, the proportion of Ascomycota in the X system was lower than in the other three planting systems. Specifically, Ascomycota abundance was highest in the MC system (77%), followed by the HR system (75.5%), and HC system (70.7%). This decrease in the proportion of Ascomycota in the HR and HC systems suggested that peanut cultivation may enhance fungal microbial diversity during the long-term cotton cultivation. Additionally, the proportion of Glomeromycota, associated with rhizobium formation, was similar in the HR and HC systems, significantly surpassing that in the MC and X systems. Conversely, the proportion of Mortierellomycota exhibited an opposing trend in the X system compared to the MC, HC, and HR systems, indicating that the fallow state may contribute to microbial community diversity recovery. In the HR and HC systems, the proportion of other bacteria exceeded that in MC system. Overall, crop rotation affected soil microorganism composition, with significant disparities in microbial richness and diversity.
In this experiment, FUNGuild was employed to predict the functional roles of fungal communities across different planting patterns. Functional subclasses (Guilds) were analyzed using the FunGuild functional annotation clustering heat map (Fig. 2C). Endophyte -- Plant -- Pathogen, Animal -- Pathogen, Plant -- Undefined -- Saprotroph, and Wood -- Saprotroph were relatively abundant in the MC system. Endophyte - Undefined - Saprotroph, Animal - Parasite - Fungal - Parasite, Plant - Pathogen-Soil - Saprotroph - Wood - Saprotroph, Leaf_ in HC system Saprotroph − Plant_Pathogen − Undefined_Saprotroph − Wood_Saprotroph is relatively abundant. In HR system, Animal_Endosymbiont − Undefined -- Saprotroph and Arbuscular -- Mycorrhizal are relatively abundant. In fallow system, Lichenized, Fungal_Parasite − Undefined_Saprotroph, Dung - Saprotroph - Undefined - Saprotroph - Wood - Saprotroph, Animal - Pat were formed hogen-Endophyte-Plant - Pathogen-Wood - Saprotroph functional colony.

Soil rhizosphere fungal community structure of different planting systems in 2021. (A) Principal coordinate analysis (PCoA), Reflecting the differences between different planting systems, there was a significant difference between MC and HC in PCoA1; (B) percentage of fungal community structure at the phylum level. In the phylum-level analysis, the fungal community richness was highest for Ascomycota and Mortierellomycota; (C) functional prediction of fungal communities in different planting patterns by FUNGuild. Twenty representative functional groups were predicted and analyzed in total; (D) ACE, Chao1, and Shannon diversity indices. In the analysis of abundance and diversity, the significance of HC was significantly higher than that of the other three planting systems.
Effects of different planting patterns on soil bacterial community composition
The PCoA revealed that the PCoA1 accounted for 30.14% and PCoA2 for 11.33% of the variance in the first and second principal component analyses, respectively. Each planting system exhibited a high degree of sample aggregation, with significant differences observed between the HR, HC, MC, and X systems (Fig. 3A). The correlation coefficient between HR: MC is small, indicating a weak correlation (Table 4).
The macrogene sequencing identified 10 primary phyla of rhizosphere bacteria across different cultivation systems: unidentified phyla Bacteria, Proteobacteria, Acidobacteria, Actinobacteria, Firmicutes, Bacteroidetes, Chloroflexi, Gemmatimonadetes, Planctomycetota, and Crenarchaeota (Fig. 3B). Proteobacteria and Acidobacteriota were dominant, accounting for 15.2% and 14.2% of the top 10 bacteria, respectively. Compared with the X system, there were significant variations in the proportions of bacteria across different phylam in each system. Specifically, the relative abundance of Proteobacteria in the rhizosphere soil of the MC and HC systems was notably higher than that of the HR system. The relative abundances of Acidobacteria and Actinobacteria in the HR system presented no significant difference from MC and HC, while the relative abundance of Acidobacteria in the X system was lower than that in the other three cultivation systems. There was a notable upward trend in the relative abundance of Actinobacteria in rhizosphere soil. In the HR system, the relative abundances of Firmicutes and Bacteroidetes differed significantly between continuous cropping and fallow, at 3.06% and 0.28%, respectively. The relative abundance of Chloroflexi in the HC and MC systems remained relatively stable compared to the X system, while in HR system, it decreased significantly by 0.13% compared to HC, and was lower than that in the X system. In the rhizosphere soil of the HR system, the relative abundances of Gemmatimonadetes and Planctomycetota did not differ significantly from those observed in the HC, MC, and X systems.
The FAPROTAX functional annotation cluster heatmap (Fig. 3C) revealed diverse metabolic pathways in the bacteria across different planting systems. The analysis of the bacterial community functions indicated that the MC planting system harbored a microbial community with relatively high richness. The HC planting system resulted in the highest relative abundance of bacteria involved in chitinolysis. In the X planting system, functional bacteria related to aerobic_chemoheterotrophy and aromatic_compound_degradation were involved. HR system contains the most functional bacterial groups, nitrate_respiration, fermentation, animal_parasites_or_symbionts, chemoheterotrophy, human_pathogens_all, plant_pathogen, human_pathogens_septicemia, human_pathogens_pneumonia, human_pathogens_gastroenteritis, human_gut and mammal_gut, functional abundance were the most obvious. The results offered a theoretical framework for characterizing the soil bacterial microbial diversity and exploring new genetic resources.

Soil rhizosphere bacterial community structure of different planting systems in 2021. (A) Principal coordinate analysis (PCoA). Regarding the PCoA analysis of the bacterial community, in PCoA1, fallow X was significantly different from the other three planting methods; (B) percentage of bacterial community structure at the phylum level. In the detection of bacterial phylum analysis, Proteobacteria and Acidobacteriota had the highest richness; (C) functional prediction of bacterial communities in different planting patterns by FAPROTAX. Twenty representative functional groups were predicted and analyzed in total; (D) ACE, Chao1, and Shannon diversity indices. In the analysis of abundance and diversity, the significance of X was significantly higher than that of the other three planting systems.
Effects of different planting patterns on soil microbial diversity
The abundance-based coverage estimation values of ACE, Chao1, and Shannon were used to represent microbial community richness and diversity. The experimental findings revealed notable discrepancies in the soil microbial community ACE, Chao1, and Shannon indices across the various planting systems. In the X-system soil, bacterial community ACE, Chao1, and Shannon indices significantly surpassed those in the rotation and continuous cropping systems (Fig. 2D), indicating that fallow enhanced soil microbial community resilience and diversity. In the HR planting system, to soil bacterial and microbial community richness and diversity were significantly lower than in the fallow and continuous cropping systems. In the HC planting system, fungal the Shannon diversity index notably higher than that in the HC, HR, and X planting systems(Figs. 3D). Regarding the uniformity measurement, a significant difference in the Shannon index was observed between MC and HC, whereas ACE and Chao1 diversity indices demonstrated no significant variation between the different planting systems. In summary, there were significant differences in the richness and diversity of microbial communities between fallow soil, rotation, and continuous cropping systems.
Correlation analysis between soil microbial communities and soil nutrients
In rhizosphere soil, N, P, OM, and AK were significantly correlated with Acidobacteria and Actinobacteria. The OM was significantly correlated with Acidobacteria, as well as with Actinobacteria, Gemmatimonadetes, and Planctomycetota. P demonstrated significant correlations with Gemmatimonadetes and Firmicutes. AK was notably correlated with Planctomycetota. Additionally, K was significantly correlated with Crenarchaeota. No significant correlation was observed between BN and all major phyla(Fig. 4A).
Significant relationships were observed between N, P, and OM in the rhizosphere soil and Ascomycota and Mortierellomycota. Additionally, a notable relationship was identified between K and Olpidiomycota. However, there was no significant correlation detected between BN and the primary fungal phyla. Furthermore, AP was significantly correlated with Mortierellomycota and Glomeromycota, and AK presented a significant relationship with Mortierellomycota(Fig. 4B).
The RDA of the rhizosphere soil physicochemical properties and soil microbial community composition revealed that at the gate level, the RDA results of the fungal community (Fig. 4C) indicated 54.68% and 32.61% of the total microbial community variation through the first and second axes, respectively. The soil nutrients in the RDA2 direction were positively correlated with Chytridiomycota, Ascomycota, Glomeromycota, Rozellomycota, Kickxellomycota, Zoopagomycota, and Olopidiomycota abundance. In the RDA1 direction, K, AP, and AK were positively correlated with Glomeromycota, Rozellomycota, Kickxellomycota, Zoopagomycota, Olopidiomycota, and Basidiomycota. BN, OM, P, and N were positively correlated with Chytridiomycota, and Ascomycota was the most affected, exhibiting a significant correlation. Similarly, the RDA results of bacteria at the gate level illustrated that 55.32% and 37.22% of the total variation in the microbial community was explained by the first and second axes, respectively (Fig. 4D). Soil nutrients in the RDA1 direction were positively correlated with Acidobacteriota, Planctomycetota, and Firmicutes. In the RDA2 direction, K, AK, N, and BN exhibited positive correlation with Acidobacteriota, Proteobacteria, Chloroflexi, and Gemmatimonadota. P and OM displayed negative correlations with Firmicutes, Crenarchaeota, and Bacteroidota. Acidohacteriota was the most affected by AK, presenting a significant correlation. In summary, K, A and KOM were identified as the primary factors influencing microbial community changes.

Correlation between soil rhizosphere microbial communities and soil nutrients and Redundancy analysis of soil physicochemical properties(A). The correlation between rhizosphere soil nutrients and the rhizosphere bacterial community shows a very significant relationship between Acidobacteria and organic matter (OM); (B) The correlation between rhizosphere soil nutrients and the rhizosphere fungal community shows a very significant relationship between Mortierellomycota and available phosphorus (AP); (C) Underneath the door, the results of fungal community RDA analysis show that the first and second axes explain 54.68% and 32.61% of the total variation in microbial communities, respectively; (D) The results of the RDA analysis at the phylum level for bacteria show that the first and second coordinate axes account for 55.32% and 37.22% of the total variation in the microbial communities, respectively.
Discussion
Studies have indicated that continuous cropping systems not only reduce crop yields and elevate the incidence of soil-borne pathogens55 but also cause soil nutrient imbalances56 and affect the structure of soil microbial communities. Soil microorganisms that are integral to agricultural ecosystems have garnered considerable attention for their pivotal roles57,58,59. Research has indicated that the introduction of microbial inoculants can enhance crop yields60,61,62. However, the artificial addition of these inoculants can disrupt the natural ecological balance, leading to increased environmental strains and ecological pollution. Therefore, the adoption of intercropping and crop rotation can enhance soil microbial diversity, boost crop yields, and ameliorate soil nutrient profiles, thereby exerting a profound impact on the structure of soil microbial communities63,64.
This study indicated a consistent increase in cotton production over five consecutive years in the rotation system. In 2021, cotton production was projected to increase by 74.1 kg compared with 2016, representing an 18.9% year-on-year increase (Table 2). Different planting systems affect crop yields as well as soil nutrients. Specifically, compared with continuous cropping, the N and BN contents in peanut rhizosphere soil under the HR system were lower than those under the HC and MC systems. Previous studies have demonstrated that soil N content significantly decreases in rotations involving legumes and other plants. This is because rhizobia grow within the root system of peanuts during its growth process65,66. Rhizobium performs N fixation, absorption, and synthesis from the soil. Nutrient elements are required for the growth and development. Consequently, soil typically exhibits low N and BN contents67. In this study, the soil AK content was higher in the rotation system than in the continuous cropping system(Table 3), suggesting that peanut and cotton rotation enhanced soil fertility. Cotton, a potassium-loving crop, efficiently absorbed K ions68. Cotton cultivation over an extended period has caused a severe deficiency of K ions in the soil, resulting in nutrient imbalance. Through crop rotation with peanuts, K ions are replenished, consequently reducing the requirement for K fertilizers. Additionally, compared to the HC, HR, and MC systems, the AP content in the soil was the lowest in the MC system, whereas there was no significant difference in P content among the different planting systems. Significant disparities exist in soil OM content among various planting systems, with the X system exhibiting lower levels than the MC, HC, and HR systems. This discrepancy can be attributed to the accumulation of plant leaves and rhizosphere residues in the cultivated soil, which augments the soil OM content. This OM serves as a nutrient source for microbial degradation, thereby elevating soil OM content.
Previous studies have indicated that plant roots play a crucial role in recruiting rhizosphere bacterial communities, effectively inhibiting the invasion of fungal pathogens69,70,71. This study, employing α and β diversity index analysis, revealed that rotating peanuts led to a reduction in the α diversity of bacterial and fungal communities. The fungal community diversity index of the HR system did not significantly differ from that of the X system, while it was lower than those of the MC and HC systems. Notably, in X system, the soil bacterial community exhibited the highest ACE, Chao1, and Shannon indices, indicating that different crop rotations effectively inhibited the increase in microbial communities associated with cotton as the host. Studies have suggested that various vegetation types can influence the restoration of the soil microbial community structure72. Changes in functional microbial communities can significantly accelerate soil nutrient transformation73. Crop rotation facilitates the preservation of soil moisture and temperature in the cultivation layer, effectively alleviating or restoring the effects of long-term farming on soil microorganisms. This provide a conducive environment for microbial life, promoting the growth and reproduction of microorganisms, and enhancing microbial diversity74.
This study revealed a significant impact of crop rotation patterns on the microbial community structure and composition75. Among the bacterial communities, Proteobacteria predominated, comprising 15.2% of the total. During the research process, it was found that the diversity of the Proteobacteria was significantly higher than that of other bacterial phyla, consistent with the results of a previous study by Sudini76. In this study, the Proteobacteria was identified as the most important bacterial phylum in peanut cultivation, capable of regulating the acidity and alkalinity of the rhizosphere soil and promoting microbial diversity in the rhizosphere. Additionally, it was found that the relative abundance of Planctomycetota and Acidobacteria was relatively high in the cotton-peanut rotation system (HR), indicating that rotation may have a significant impact on reducing soil-borne diseases77.Previous studies have indicated a higher abundance of Actinomycetes in the HR mode, with Streptomyces, a key genus known for organic matter decomposition and soil-borne pathogen inhibition78,79,80. Streptomyces is notable for its capacity to generate abundant antibiotics, rendering it a potent biological control agent81,82. Moreover, the HR pattern increased the prevalence of Firmicutes. Bacilli, a significant subgroup of Firmicutes, actively engage in numerous soil biological reactions. Their distinctive capability enables them to optimize the plant rhizosphere soil ecosystem and decompose various harmful substances, such as phenolic acids, phenols, and other toxins83,84. This indicates that cotton-peanut rotation can change the diversity of the soil microbial community and enhance the content of beneficial microorganisms. Additionally, the HR system soil harbored a proportion of Chloroflexi, which can decompose organic residues85. As peanuts are harvested, remnants, such as pod residues and peanut leaves, reintegrate into the soil, promoting the abundance of Chloroflexi.
Compared to bacteria, crop rotation significantly influences fungal microbial communities more than bacterial rotation. Soil fungi exhibit heightened environmental sensitivity owing to their ability to acquire nutrients from soil. Fungi play a pivotal role in nutrient cycling within soil ecosystems by absorbing organic N and P86,87. They are key factors in the decomposition of OM, and the presence and composition of fungal communities profoundly affect decomposition rates and carbon cycle processes88. This study revealed significant disparities in soil fungal community structure across various planting systems. Across all soil samples, the fungal community at the phylum level was primarily composed of Ascomycota, Mortierellomycota, Chytridiomycota, and Basidiomycota. These findings were consistent with those of previous studies on rotational soils involving peanut, pea, and vanilla crops89,90. At the phylum level, the fungal composition was dominated by Ascomycota and Mortierellomycota, accounting for about 80% of the top 10 fungal phyla. Ascomycota had the highest abundance of 77% in the MC system, followed by the HR system with 75.5%, and the lowest in the HC system with 70.7%. This indicates that the structure of the soil fungal community varies under different planting modes.Ascomycota, Basidiomycota, and Chytridiomycota are predominantly saprophytic soil fungi participating in aerobic cellulose degradation91, which can enhance soil fertility but may also increase the incidence of soil-borne diseases during decomposition. Notably, Ascomycota played a significant role in the fungal community of the rhizosphere soil across all treatments and was potentially associated with plant residue degradation. Ascomycota encompasses numerous plant pathogens such as Fusarium, which can cause Fusarium wilt and detrimentally affect plant growth, leading to substantial economic losses62. Meanwhile, a beneficial fungal community, Glomeromycota, was identified in the soil under the cotton-peanut rotation system. These fungi are associated with rhizobial production and play pivotal roles in crop growth. They can facilitate N recruitment and fixation in the crop rhizosphere, thereby enhancing the synthesis of organic nutrients that are crucial for crop development. After rotation, the relative abundance of cotton-peanut root microbial communities significantly increased. Peanut roots form nodules that harbor rhizobia, which form mycorrhizal associations and enhance interactions with other soil microbial communities, promoting the diversity of soil microbial functions. Moreover, when legume plants are rotated, the decomposition of legume residues releases organic nitrogen compounds, increasing the diversity of organic matter in the soil and promoting nitrogen content92, enhancing the accumulation of plant proteins93, and reducing the reliance on nitrogenous fertilizers94. Among different cropping systems, microbial functional groups related to rhizobia are particularly prominent. The increased diversity of rhizosphere microorganisms can significantly reduce the damage from pathogens and improve the overall function of the root system95. The cotton-peanut rotation can effectively alter specific pathogenic microorganisms that thrive due to prolonged monocropping. Because the microbial communities that are adapted to the cotton root environment cannot quickly adapt to the peanut root environment, they are effectively suppressed. Therefore, experimental studies have shown that the HR cropping system can enhance beneficial fungal communities in the root zone while suppressing harmful bacterial communities by increasing root exudates that affect the growth environment of microorganisms in the root zone. Harmful microbial communities that rely on root exudates from cotton roots are influenced by the addition of peanuts in the rotation, as the exudates from peanut roots cannot be absorbed by the original microbial groups, leading to the disruption of their nutritional environment and preventing normal growth.
Soil microorganisms, as key members of agricultural systems, are capable of decomposing soil organic matter, providing absorbable nutrients for crops, and increasing crop yields96. However, the structure of soil microbial communities is influenced by numerous factors, with agricultural planting patterns, soil characteristics, and soil nutrients significantly affecting microbial community structure97. Research has found that the main factors influencing the differences in soil microbial community structure at the phylum level are the positive correlation between Ascomycota and soil organic carbon content, and the positive correlation between Acidobacteria and Actinobacteri with soil nitrogen content98,99. This study found that soil potassium (K), available potassium (AK), and available phosphorus (AP) are positively correlated with Ascomycota. This further suggests that cotton-peanut rotation is beneficial for the growth of subsequent crops.
Moreover, the sampling in this study was conducted after crop harvest, which, to some extent, only reflects the differences in microbial activity during the later stage of crop growth. During the cropping rotation, different crops have different growth and development periods, and microbial communities also undergo changes. Therefore, in subsequent research, it is important to study the soil microbial communities and their functions during the growth process of crops under the rotational system to more scientifically interpret the relationship between microbial community structure research and agricultural production.
Conclusion
Compared with continuous cropping, cotton-peanut rotation has led to consecutive yearly increases in peanut and cotton yields. Notably, peanut production in 2020 increased by 10.59% compared to 2016, and cotton production in 2021 increased by 14.63% compared to 2017. These data indicated crop rotation not only increased cotton and peanut yields, but also augmented the N and K content in the rhizosphere soil, thereby enhancing soil nutrients. This practice also reduced the application of N and K fertilizers, enhancing the bioavailability of N and K in the rhizosphere soil. By utilizing the cotton-peanut rotation system, the different secretions in the rhizosphere effectively improve the rhizosphere soil environment, allowing for a more scientific and environmentally friendly approach to biological pest control. Furthermore, cotton-peanut rotation can significantly change rhizosphere soil environmental factors and rhizosphere soil microbial structure, promote rhizosphere soil microbial diversity, and effectively mitigate the challenges associated with continuous cropping, provides favorable ways and Utilizing scientific methods to manage planting in order to increase crop yield.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
National Bureau of Statistics. Announcement of the National Bureau of Statistics on cotton production in 2022[EB/OL].(2022-12-26) (2022-12-30),http://www.stats.gov.cn/xxgk/sjfb/zxfb2020/202212/t20221227_1891259.html (2022).
Xi, H. et al. Effects of cotton-maize rotation on soil microbiome structure. Mol. Plant. Pathol. 22 (6), 673–682 (2021).
Chen, H., Yang, L. & Zhang, F. H. Effects of Long-term cotton continuous cropping on Soil Physical and Chemical Properties and Nematode Community in Xinjiang. J. Chin. J. Appl. Ecol. 32 (12), 4263–4271 (2021).
Chen, X. L. & Yao, C. H. Effects of continuous cropping of cotton on main physical and chemical properties of soil. J. China Cotton. 41 (06), 23–24 (2014).
Zhu, S. et al. Rotary and subsoiling tillage rotations influence soil carbon and nitrogen sequestration and crop yield. J. Plant. Soil. Environ. 68 (02), 89–97 (2022).
Li, L. et al. GWAS and bulked segregant analysis reveal the loci controlling growth habit-related traits in cultivated peanut (Arachis hypogaea L). BMC Genom 23,403 (2022).
Zhang, L. W. & Wang, L. W. Development status, problems and policy suggestions of Peanut Industry in China. J. China Oils Fats. 45 (11), 116–122 (2020).
Wan, S. B. & Zhang, J. L. Xinjiang Peanut Industry Development Strategy and countermeasures. J. J. Peanut Sci. 48 (02), 66–68. https://doi.org/10.14001/j.issn.1002-4093.2019.02.011 (2019).
Shi, B. I. et al. Research on the current status, trends, and Development Strategies of Xinjiang Peanut production. J. Xinjiang Agricultural Sci. 54 (03), 574–584 (2017).
Venter, Z. S., Jacobs, K. & Hawkins, H. J. The impact of crop rotation on soil microbial diversity: a meta-analysis. J. Pedobiologia. 59, 215–223. https://doi.org/10.1016/j.pedobi.2016.04.001 (2016).
Chen, Y. et al. Evolutions and managements of soil microbial community structure drove by continuous cropping. J. Front. Microbiol. 13, 839494. https://doi.org/10.3389/fmicb.2022.839494 (2022).
Zhang, B. H. Study on the effects of fertilization on soil microbial diversity of soybean and maize rotation and reclamation.D. Shanxi Agricultural University,2021.https://doi.org/10.27285/d.cnki.gsxnu.2021.000001
Meriles, J. M. et al. Glyphosate and previous crop residue effect on deleterious and beneficial soil-borne fungi from a peanut-corn-soybean rotations. J. Phytopathol. 154, 309–316 (2006).
Zou, X. X. et al. Rotational strip intercropping of maize and peanuts has multiple benefifits for agricultural production in the northern agropastoral ecotone region of China. J. Eur. J. Agron. 129,126304 (2021).
Katsvairo, T., Cox, W. J. & Es, H. V. Tillage and Rotation effects on Soil Physical Characteristics.J. Agron. J. 94 (2), 299–304. https://doi.org/10.2134/agronj2002.0299 (2002).
Sharma, N. K. et al. Increasing farmer’s income and reducing soil erosion using intercropping in rainfed maize-wheat rotation of Himalaya. India J. Agric. Ecosyst. Environ. 247, 43–53. https://doi.org/10.1016/j.agee.2017.06.026 (2017).
Guochun, L. et al. Legacy effects of wheat season organic fertilizer addition on microbial co-occurrence networks, soil function, and yield of the subsequent maize season in a wheat-maize rotation system.J. J. Environ. Manage. 347, 119160–119160 (2023).
Qin, S. et al. Breaking continuous potato cropping with legumes improves soil microbial communities,enzyme activities and tuber yield. PloS One. 12 (5), e0175934. https://doi.org/10.1371/journal.pone.0175934 (2017).
Berzsenyi, Z., Győrffy, B. & Lap, D. Effect of crop rotation and fertilisation on maize and wheat yields and yield stability in a long-term experiment.J. Eur. J. Agron. 13. 0301 (00), 00076–00079. https://doi.org/10.1016/S1161- 2-3, 225–244. (2000).
Bennett, A. J., Bending, G. D., Chandler, D., Hilton, S. & Mills, P. Meeting the demand for crop production: the challenge of yield decline in crops grown in short rotations. J. Biol. Reviews Camb. Philosophical Soc. 87 (1), 52–71. https://doi.org/10.1111/j.1469-185X.2011.00184.x (2012).
Xuan, D. T. et al. Different crop rotation systems as drivers of change in soil bacterial community structure and yield of rice, Oryza sativa. J. Biology Fertility Soils. 48, 217–225. https://doi.org/10.1007/s00374-011-0618-5 (2012).
Soman, C., Li, D., Wander, M. M. & Kent, A. D. Long-term fertilizer and crop-rota-tion treatments differentially affect soil bacterial community structure. J. Plant. Soil. 413, 145–159. https://doi.org/10.1007/s11104-016-3083-y (2017).
McDaniel, M. D., Grandy, A. S., Tiemann, L. K. & Weintraub, M. N. Crop rotation complexity regulates the decomposition of high and low quality resi-dues. J. Soil Biol. Biochemistry. 78, 243–254. https://doi.org/10.1016/j.soilbio.2014.07.027. (2014).
Tiemann, L. K., Grandy, A. S., Atkinson, E. E., Marin, S. E. & McDaniel, M. D. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. J. Ecol. Lett. 18.8, 761–771. https://doi.org/10.1111/ele.12453. (2015).
Kost, C. et al. Metabolic exchanges are ubiquitous in natural microbial communities. J. Nat. Microbiol. 8.12, 1–9 (2023).
Sharma, S. K. et al. Microbial community structure and diversity as indicators for evaluating soil quality. J. Biodiversity Biofuels Agroforestry Conserv. Agricult. Sustain. Agric. Rev.. 5, 317–358 (2011).
Yu, L. et al. Molecular characterization of root-associated fungal communities in relation to health status of Pisum sativum using barcoded pyrosequencing. J. Plant. Soil. 357 (01), 395–405 (2012).
Kracmarova, M. et al. Soil microbial communities following 20 years of fertilization and crop rotation practices in the Czech Republic. J. Environ. Microbiome. 17 (1), 13. https://doi.org/10.1186/s40793-022-00406-4 (2022).
Tian, K. et al. Effects of long-term fertilization and residue management on soil organic carbon changes in paddy soils of China: a meta-analysis. J. Agric. Ecosyst. Environ. 204, 40–50 (2015).
Uroz, S. et al. Pyrosequencing reveals a contrasted bacterial diversity betweenoak rhizosphere and surrounding soil. J. Enviromental Microbiol. Rep. 2(2), 281–288 (2010).
Watts, C. W. et al. Effects of mechanical energy inputs on soil respiration at theaggregate and field scales. J. Soil. Till Res. 53 (34), 231243 (2000).
Deng, Y. Q. Effects of crop Rotation on soil Microbial Community Structure, Enzyme Activity and Nutrients of Tomato Continuous cropping.D (Haikou:Hainan University, 2018).
Uzoh, I. M. et al. Legume-maize rotation effect on maize productivity and soil fertility parameters under selected agronomic practices in a sandy loam soil. J. Sci. Rep. 9 (1), 1–9 (2019).
Zhang, L. L. et al. Soil oxidoreductase activity and kinetic characteristics of different cropping systems. J. Ecol. Environ. Sci. 18 (01), 343–347 (2009).
Ding, J. N. et al. Effects of maize-soybean rotation on soil enzyme and Root Microbial Community Diversity. J. Heilongjiang Agricultural Sci. 04, 11–16 (2021).
Jin, H. Y. et al. Response of Soil Fungal communities in Diversified rotations of Wheat and different crops. J. Huan Jing Ke Xue. 43 (6), 3338–3347. https://doi.org/10.13227/j.hjkx.202109069 (2022).
Yu, Z. H. et al. Effect of different fertilization measures on soil enzyme activity characteristics of farmland black soil profile. J. Soil. Crops. 7 (03), 276–283 (2018).
Li, C. et al. Effect of monoculture soybean on soil microbial community in the Northeast China. J. Plant. Soil. 330, 423–433 (2010).
Zhu, L. et al. Analysis of Bacterial Community diversity in soybean continuous cropping soil based on high-throughput sequencing. J. Soybean Sci. 36 (3), 419–424 (2017).
Tang, H. et al. Prokaryotic diversity in continuous cropping and rotational cropping soybean soil. J. FEMS Microbiol. Lett. 298 (2), 267–273 (2009).
Xi, H. et al. Effects of cotton-maize rotation on soil microbiome structure. J. Mol. Plant. Pathol. 22, 673–682 (2021).
Perez, B. C. et al. The effect of crop sequences on soil microbial,chemical and physical indicators and its relationship with soybean sudden death syndrome (complex of Fusarium species). J. Span. J. Agricultural Res. 12 (1), 252–264 (2014).
Pérez, B. C. et al. Soybean fungal soil-borne diseases: a parameter for measuring the effect of agricultural intensification on soil health. J. Can. J. Microbiol. 60 (2), 73–84 (2014).
Bi, B. Y. & Han, F. P. GC-MS analysis of Alfalfa Root exudates at different planting years on the Loess Plateau. J. Acta Grassland Sinica. 26 (03), 611–617 (2018).
Liu, D. D., Li, M. & Liu, R. J. Research Progress on Plant Root Periphery Growth-promoting Bacteria in China. J. Chin. J. Ecol. 35 (03), 815–824 (2016).
Simpson, R. T. et al. Preferential accumulation of microbial carbon in aggregate structures of no-tillage soils. J. Soil. Sci. Soc. Am. J. 68 (4), 1249 (2004).
Zhang, W. et al. Comprehensive Assessment of 16S rRNA gene amplicon sequencing for Microbiome profiling across multiple habitats. Microbiol. Spectr. 11 (3), e0056323 (2023).
Ranjan, R., Rani, A., Metwally, A., McGee, H. S. & Perkins, D. L. Analysis of the microbiome: advantages of whole genome shotgun versus 16S amplicon sequencing. Biochem. Biophys. Res. Commun. 469 (4), 967–977 (2016).
Caporaso, J. G. et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. J. ISME J. 6, 1621–1624 (2012).
Oros, S. M. & Smalla, K. Semi-nested PCR approach to amplify large 18S rRNA gene fragments for PCR-DGGE analysis of soil fungal communities. Laboratory Protocols in Fungal Biology: Current Methods in Fungal Biology. 289 – 98 (2013).
Bachy, C. et al. Accuracy of protist diversity assessments: morphology compared with cloning and direct pyrosequencing of 18S rRNA genes and ITS regions using the conspicuous tintinnid ciliates as a case study. J. ISME J. 7, 244–255 (2013).
Bengtsson, P. J. et al. Improved software detection and extraction of ITS1 and ITS 2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. J. Methods Ecol. Evol. 4, 914–919 (2013).
Caporaso, J. G. et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proceedings of the national academy of sciences. 108(supplement_1), 4516–4522 (2011).
Bao, S. D. Soil agrochemical analysis 3rd edition.M. China Agriculture Press, (2000).
Wright, P., Falloon, R. & Hedderley, D. A long-term vegetable crop rotation study to determine effects on soil microbial communities and soilborne diseases of potato and onion. New. Zeal J. Crop Hortic. 45, 1–26. https://doi.org/10.1080/01140671.2016.1229345 (2016).
Puniya, R., Pandey, P. C., Bisht, P. S., Singh, D. K. & Singh, A. P. Effect of long-term nutrient management practices on soil micronutrient concentrations and uptake under a rice-wheat cropping system. J. Agric. Sci. 157 (3), 226–234. https://doi.org/10.1017/S0021859619000509 (2019).
Xiong, C. & Lu, Y. Microbiomes in agroecosystem: diversity, function and assembly mechanisms. J. Environ. Microbiol. Rep. 14 (6), 833. https://doi.org/10.1111/1758-2229.13126 (2022).
Gupta, A. et al. Linking soil microbial diversity to modern agriculture practices: a review.J. Environ. Res. Public. Health. 19 (5), 3141. https://doi.org/10.3390/ijerph19053141 (2022).
Markalanda, S. H., McFadden, C. J., Cassidy, S. T. & Wood, C. W. The soil microbiome increases plant survival and modifies interactions with root endosymbionts in the field. Ecol. Evol. 12 (1), e8283 (2022).
Chen, L. H. et al. Application of Trichoderma Harzianum SQR-T037 bio-organic fertiliser significantly controls Fusarium wilt and affects the microbial communities of continuously cropped soil of cucumber. J. Sci. Food Agric. 92 (12), 2465–2470 (2012).
Chen, W. et al. Mechanisms by which organic fertilizer and effective microbes mitigate peanut continuous cropping yield constraints in a red soil of south China. J. Appl. Soil. Ecol. 128, 23–34 (2018).
Ling, N., Zhang, W., Tan, S., Huang, Q. & Shen, Q. Effect of the nursery application of bioorganic fertilizer on spatial distribution of Fusarium oxysporum f. sp. niveum and its antagonistic bacterium in the rhizosphere of watermelon. J. Appl. Soil. Ecol. 59, 13–19 (2012).
Larkin, R. P. et al. Effects of different potato cropping system approaches and water management on soilborne diseases and soil microbial communities. J. Phytopathol. 101 (1), 58–67. https://doi.org/10.1094/PHYTO-04-10-0100 (2011).
Larkin, R. P., Honeycutt, C. W., Griffn, T. S., Olanya, O. M. & He, Z. Q. Potato growth and yield characteristics under different cropping system management strategies in northeastern U.S. J. Agron. 11 (1), 165. https://doi.org/10.3390/agronomy11010165 (2021).
Pan, L. J. et al. Screening and evaluation of high-yield peanut varieties suitable for peanut-cotton Rotation System. J. J. Peanut. 52 (04), 60–68. https://doi.org/10.14001/j.issn.1002-4093.2023.04.008 (2023).
Long, X. E. et al. Phosphate levels influence the utilisation of rice rhizodeposition carbon and the phosphate-solubilising microbial community in a paddy soil. J. Soil. Biology Biochem. 118, 103–114 (2018).
Dong, Q. Q. et al. Effects of maize and peanut strip intercropping on plant nitrogen uptake and soil microbial community. J. Chin. J. Oil Crop Sci. 44 (06), 1296–1306. https://doi.org/10.19802/j.issn.1007-9084.2021283 (2022).
Wang, X. R. et al. Root morphology and physiological mechanism of differences in potassium uptake efficiency of different cotton varieties. J. Cotton Sci. 28 (02), 152–159 (2016).
Liu, H. et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. J. New. Phytologist. 229 (5), 2873–2885 (2021).
Song, Y. et al. FERONIA restricts Pseudomonas in the rhizosphere microbiome via regulation of reactive oxygen species. J. Nat. Plants. 7, 644–654 (2021).
Li, X. G. et al. Acidification suppresses the natural capacity of soil microbiome to fight pathogenic fusarium infections. J. Nat. Commun. 14, 5090 (2023).
Deng, J. J., Yin, Y., Zhu, W. X. & Zhou, Y. B. Variations in soil bacteral community diversity and structures among different revegetation types in the Baishilazi nature reserve. J. Front. Microbiol. 9,2874 (2018).
Yu, J., Liu, F., Tripathi, B. M. & Steinberger, Y. Changes in the composition of soil bacterial and fungal communities after revegetation with Caraganamicrophylla in a desertified semiarid grassland.J. J. Arid Environ. 182, 1042–1062 (2020).
Xu, G. H. et al. Influence of Fallow and Crop Rotation on Microbial Carbon in Farmland Soil. J. Chin. J. Soil. Sci. 49 (04), 897–901. https://doi.org/10.19336/j.cnki.trtb.2018.04.20 (2018).
Wang, X. Y. et al. Response of potato yield, soil chemical and microbial properties to different rotation sequences of green manure-potato cropping in north China. J. Soil. Till Res.. 217, 1052–1073. https://doi.org/10.1016/j.still.2021.105273 (2022).
Sudini, H., Liles, M. R., Arias, C. R., Bowen, K. L. & Huettel, R. N. Huettel exploring soil bacterial communities in different peanut-cropping sequences using multiple molecular approaches. J. Phytopathol. 101 (7), 819–827 (2011).
Dong, Y. F. et al. Effects of different cultivation patterns on soil microbial community and enzyme activity in continuous cropped pepper field. J. Acta Agric. Zhejiangensis. 31 (9), 1485–1492. https://doi.org/10.13930/j.cnki.cjea.140755 (2019).
Herrmann, M. et al. Predominance of cand. Patescibacteria in groundwater is caused by their preferential mobilization from soils and flourishing under oligotrophic conditions. J. Front. Microbiol. 10, 1407. https://doi.org/10.3389/fmicb.2019.01407 (2019).
Janvier, C. et al. Soil health through soil disease suppression: which strategy from descriptors to indicators? J. Soil. Biol. Biochem. 39 (1), 1–23 (2007).
Luo, Q., Hiessl, S. & Steinbüchel A.Functional diversity of Nocardia in metabolism. J. Environ. Microbiol. 16 (1), 29–48 (2014).
Shekhar, N., Bhattacharya, D., Kumar, D. & Gupta, R. K. Biocontrol of wood-rotting fungi with Streptomyces violaceusniger XL-2. J. Can. J. Microbiol. 52 (9), 805–808 (2006).
Rafael, V., Elke, L. F. & de, N. C. Rhizospheric Streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda. J. BioControl. 54, 807–816 (2009).
Paz, A. et al. Biotransformation of phenolic compounds by Bacillus aryabhattai. J. Bioprocess. Biosyst Eng. 42, 1671–1679 (2019).
Peng, L. et al. Mobilization of recalcitrant phosphorous and enhancement of pepper P uptake and yield by a new biocontrol and bioremediation bacterium Burkholderia cepacia CQ18. J. Appl. Microbiol. 130 (6), 1935–1948 (2021).
Zeng, Y. H. et al. Gemmatimonas Groenlandica sp. nov. is an aerobic anoxygenic phototroph in the phylum gemmatimonadetes. J. Front. Microbiol. 11 https://doi.org/10.3389/fmicb.2020.606612 (2021).
Mäkipää, R. et al. Interactions between soil-and dead wood-inhabiting fungal communities during the decay of Norway spruce logs. J. ISME. 11 (9), 1964–1974 (2017).
Xiong, W. et al. Bio-fertilizer application induces soil suppressiveness against Fusarium wilt disease by reshaping the soil microbiome. J. Soil. Biol. Biochem. 114, 238–247 (2017).
Xu, L., Ravnskov, S., Larsen, J., Nilsson, R. H. & Nicolaisen, M. Soil fungal community structure along a soil health gradient in pea fields examined using deep amplicon sequencing. J. Soil. Biol. Biochem. 46, 26–32 (2012).
Shen, Z. et al. Effect of biofertilizer for suppressing Fusarium wilt disease of banana as well as enhancing microbial and chemical properties of soil under greenhouse trial. J. Appl. Soil. Ecol. 93, 111–119 (2015).
Zhao, J. et al. The rhizosphere microbial community response to a bio-organic fertilizer: finding the mechanisms behind the suppression of watermelon Fusarium wilt disease. J. Acta Physiol. Plant. 40, 1–14 (2018).
Folman, B. W., Summerbell, L. B., Boddy, R. C. & de,, Living in a fungal world: impact of fungi on soil bacterial niche development. J. FEMS Microbiol. Rev. 29 (4), 795–811. https://doi.org/10.1016/j.femsre.2004.11.005 (2005).
Ma, B. L. et al. The carbon footprint of maize production as affected by nitrogen fertilizer and maize-legume rotations. J. Nutr. Cycl. Agroecosyst. 94 (1), 15–31. https://doi.org/10.1007/s10705-012-9522-0 (2012).
Plaza, B. D. et al. Innovative cropping systems to reduce N inputs and maintain wheat yields by inserting grain legumes and cover crops in southwestern France. J. Eur. J. Agron. 82(1),331 – 41 , 2017). https://doi.org/10.1016/j.eja.2016.05.010
Roesch, M. G. E., Arbuckle, J. G. & Tyndall, J. C. Barriers to implementing climate resilient agricultural strategies: the case of crop diversification in the US Corn Belt. J. Global Environ. Change. 48 (1). https://doi.org/10.1016/j.gloenvcha.2017.12.002,206 – 15. (2018).
Tao, C. Y. et al. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. J. Microbiome. 8 (01), 137. https://doi.org/10.1186/s40168-020-00892-z (2020).
Six, J. et al. Bacterial and fungal contributions to carbon sequestration in agroeco systems. J. Soil. Sci. Soc. Am. J. 70, 555–569 (2006).
Gao, S. C. et al. Effect of fertilization on bacterial community in black soil of Northeast China under continuous cropping of soybean. J. China Agricultural Sci. 50 (07), 1271–1281 (2017).
Li, L. R. et al. Effect of crop planting patterns on Soil microorganisms and Crop pests in Farmland. J. Chin. Agricultural Sci. Bull. 37 (29), 99–106 (2021).
Wang, G. Z., Jia, J. Y. & Zhang, J. L.Plant soil feedback theory and its applications and prospects in natural and agricultural. J. Ecosyst. Acta Ecol. Sinica. 41 (23), 9130–9143 (2021).
Funding
This work was supported by (1) the National Key Research and Development Program of China (2022YFD1000105); (2) China Agriculture Research System of MOF and MARA (CARS-13); (3) Xinjiang Uygur Autonomous Region’s “Tianshan Talent” Training Program - High-level Talents for Development of Systemrn Agricultural and Animal Husbandry Industry (2023SNGGGCC018).
Author information
Authors and Affiliations
Contributions
Conceptualization, C.F.Y., L.Q. and H.X.F; formal analysis: L.Q. and C.F.Y.; investigation: C.F.Y. and L.Q.; methodology, C.F.Y. and H.X.F.; writing of (original draft): C.F.Y.; writing (review, and editing): H.X.F. and S.S.T. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cui, F., Li, Q., Shang, S. et al. Effects of cotton peanut rotation on crop yield soil nutrients and microbial diversity. Sci Rep 14, 28072 (2024). https://doi.org/10.1038/s41598-024-75309-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-75309-0
