
Abstract
The circadian clock is a cell-autonomous process that regulates daily internal rhythms by interacting with environmental signals. Reports across species show that infection can alter the expression of circadian genes; however, in teleosts, these effects are influenced by light exposure. Currently, no reports analyze the direct effects of bacterial exposure on the zebrafish clock. Using zebrafish Z3 cells, we demonstrate that exposure to heat-killed Streptococcus pneumoniae (HK-Spn) augments the expression of core repressive factors in a light- and time-dependent manner. In constant darkness, HK-Spn highly upregulated cry1a, per3, and per1b expression. In the presence of light, HK-Spn exposure rapidly and strongly upregulated per2 and cry1a, and this was proportionally increased with light intensity. The combinatorial effect of light and HK-Spn on per2 and cry1a was not duplicated with H2O2, a known byproduct of light exposure. However, the ROS inhibitor N-acetyl cysteine was sufficient to block HK-Spn augmentation of per2, cry1a, and per3. These findings demonstrate that exposure to an inactive bacteria influences the expression of zebrafish clock genes under different light conditions.
Similar content being viewed by others
Introduction
Circadian rhythms are 24-hour oscillations that control different aspects of our physiology, including sleep patterns, metabolism, body temperature, hormone secretion, and immune response1,2,3. In vertebrates, these oscillations are regulated by a transcriptional/ translational feedback loop known as the molecular clock4. The activating arm of the molecular clock consists of the transcription factors Clock and Bmal, which dimerize and recognize promoter elements called E-boxes, leading to the circadian expression of hundreds of genes. Clock:Bmal also drives the production of the core repressive factors, Per and Cry, which inhibit Clock:Bmal activity, leading to a negative feedback loop that ultimately terminates Per and Cry expression4. This cell intrinsic cycle takes about 24 h to complete and is what drives daily rhythms in gene expression.
The molecular clock constantly aligns with environmental cues, such as light, to maintain synchronization with the day-night cycle5,6,7. Within vertebrates, light regulation of the clock has been extensively studied in mice, where photic input is only effective in vivo as it must be sensed through photosensitive retinal ganglion cells (pRGCs) in the eye. pRGCs signals directly activate a brain region called the suprachiasmatic nucleus (SCN), which serves as the master clock in the animal4. Peripheral murine cells, either in vivo or ex vivo, are unresponsive to light and instead rely on physiological cues such as body temperature, nutrients, or hormones to synchronize their timing8. In contrast, in zebrafish (Danio rerio), every cell directly detects and responds to light, whether in vivo or in vitro, making it a valuable tool for studying the vertebrate molecular clock in a whole organism or cell culture9,10,11. Zebrafish contain the same components of the molecular clock of other vertebrates (Clock, Bmal, Per, and Cry proteins); the difference between these systems stems from the fact that, in zebrafish, light drives transcriptional induction of per2 and cry1a due to the presence of D-boxes in their promoters11,12,13,14,15,16,17,18. This light-responsive feature allows the autonomous resetting of peripheral clocks, a characteristic that is also shared by lower organisms such as Drosophila19,20. Though mammals experience a delay between the neural detection of light and the resetting of peripheral clocks8, light remains one of the most important environmental cues that drive circadian timing across all domains of life.
Circadian rhythms are well known to control diverse aspects of our physiology, including activity, sleep, cognition, and metabolism3. Circadian rhythms also drive time-of-day-dependent differences in a variety of immune pathways and have been linked to the severity of infection in humans and diverse animal models ranging from mice to Drosophila2,21,22,23,24,25,26,27,28. In humans, clock disruption due to behavioral changes such as shift work is associated with decreased health and increased risk of respiratory infection29,30. Moreover, vaccine efficacy can differ depending on the time of administration31,32,33. Incredibly, exposure to blue light has been reported to improve appendicitis prognosis in patients due to its ability to activate REV-ERBα, a master component of the circadian stabilization loop34. The first animal model demonstrating time-of-day dependent susceptibility to infection was published in 1969, finding that mice were more resistant to Streptococcus pneumoniae (Spn) during their active phase35. No cellular explanation for this differential bacterial control was described for over 50 years until a 2020 report demonstrated that circadian regulation of phagocytosis of Spn by murine macrophages is dependent on the master clock protein BMAL136. Curiously, a 2012 study had already reported that in Drosophila, the master clock repressor Timeless (tim) differentially influences phagocytosis during day and night, regulating resistance against Spn28. These findings suggest a strategy that may be conserved across species despite the partial divergence of the core clock proteins, whereby the circadian phase dictates how cells respond to pathogens.
Dissecting these cellular mechanisms is complicated because substantial evidence demonstrates that active infection can directly alter the expression of the circadian clock. Infection with Helicobacter pylori has been shown to upregulate the expression of Bmal1 in both murine and human cells37. In contrast, there are many more examples of infection downregulating clock genes. Human cells infected with the hepatitis C virus have decreased PER2 and CRY2, and mice infected with the eukaryotic parasites Trypanosoma brucei and Plasmodium chabaudi have decreased Bmal1, Per1, and Dbp38,39. In zebrafish, RNA-seq analysis revealed that infection with the microsporidium Pseudoloma neurophilia downregulated the expression of multiple genes related to circadian rhythms in the fish brain, including per1b and nr1d140. Not surprisingly, several reports in teleosts demonstrate that light exposure is a critical variable that impacts how clock genes respond during infection, innate immune activity, and ultimately, the survival of zebrafish during bacterial infection41,42,43,44. Finally, another limitation is that these studies do not address whether changes in circadian gene expression result from the pathogen directly (e.g. detection of microbial ligands, secretion of virulence factors), inflammation, or host tissue damage. Several reports have shown that, at least in murine models, exposure to pathogen-associated molecular patterns (PAMPs) alone can alter circadian gene expression, most commonly resulting in decreased amplitude of expression45,46,47,48,49,50,51. However, no studies have investigated whether microbial ligands affect circadian gene expression in zebrafish.
Considering this, this study aimed to characterize the impact of exposure to an inactivated Gram-positive bacterium, Streptococcus pneumoniae, on the expression of two light-responsive clock genes from the repressive arm: per2 and cry1a, and two non-light-responsive genes, per1b and per3, in zebrafish cells.
Results
Basal and light-induced expression of circadian genes on Z3 cells
In the last two decades, numerous studies have demonstrated the importance of light regulating the zebrafish clock10,11,12,13,14,15,16,17,18,52,53,54,55,56,57,58,59,60. Light activates the transcription of the clock repressors per2 and cry1a, directly resetting the phase of the clock11,14,15,16,17,18. Because different light conditions and circadian phases of exposure have been shown to have varying induction levels on per2 and cry1a expression15,58,59, we first sought to characterize these genes by a qRT-PCR time course in our cells. We choose per3 and per1b as control genes not induced by light11,60. We first entrained Z3 cells, a zebrafish embryonic fibroblast-like cell line11, for at least three days in 12 h light and 12 h dark (12:12 LD, low-intensity white light ~ 4.12 × 1018 photons/s/m2). To assess the self-sustained circadian rhythm, cells were then maintained in continual darkness (DDDD) for 48 h, during which time we analyzed the transcripts expression of per2, cry1a, per3, and per1b every 4 h (Fig. 1a). Consistent with previous literature11,53, per2 and cry1a show minimal oscillations in constant darkness. To determine circadian rhythmicity, we analyzed the expression of these genes in DDDD, using the JTK algorithm on BioDare2 software. This is a non-parametric test that is widely used in circadian rhythm studies to detect oscillating molecules. While per2 and cry1a appear to oscillate in the dark, neither of these genes passed the JTK rhythmicity test. In contrast, per1b and per3 showed robust and significant oscillations in the dark, confirmed by a positive JTK rhythmicity test.

Effect of light exposure in the expression of per2, cry1a, per3, and per1b on Z3 cells. (a) Z3 cells were entrained for three LD cycles of 12:12 h. After the third day of entrainment, cells were exposed to constant darkness (DDDD) and mRNA expression of the genes per2, cry1a, per3, and per1b were analyzed every 4 h for 48 h. The eJTK test detected circadian rhythmicity on per3 and per1b, with a p-value of 0.01. No rhythmicity was detected in per2 and cry1a in DDDD. (b) mRNA expression of the genes per2, cry1a, per3 and per1b after a 12-hour exposure of low-intensity light (~ 4.12 × 1018 photons/s/m2) with subsequent darkness (LDDD). Samples were taken every 4 h for 48 h. Transcript levels were measured against time zero. Yellow bar: light exposure. Grey bar: constant darkness. Means ± SD are shown. Three replicates per group, one experiment.
To validate the effect of light on the induction of per2 and cry1a, we repeated these studies with an additional 12-hour light exposure during the first 12 h of analysis (Fig. 1b). As predicted, light drove immediate increases in per2 and cry1a (0–12 h), but not to per1b and per3, which do not respond to light. After light exposure, all four genes maintained oscillations in the following 36 h of darkness. These data demonstrate that the light-regulated expression patterns of these genes, previously shown in other studies11, are maintained in our experimental conditions.
Heat-killed Streptococcus pneumoniae augment gene expression of the zebrafish clock’s repressive arm
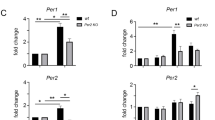
Streptococcus pneumoniae (Spn) is a Gram-positive pathogen that has been shown to elicit a circadian-dependent host response when infecting different organisms, including mice and Drosophila28,35,36. However, no studies have shown whether the sole presence of this bacteria directly affects the clock in zebrafish. Therefore, we wondered if exposure to an inactive form of Spn affects the expression of zebrafish clock genes. To test this, we incubated Z3 cells with heat-killed Streptococcus pneumoniae (HK-Spn) and exposed them to the same light conditions that had been tested previously. In constant darkness, HK-Spn substantially augmented the amplitude of cry1a, per3, and per1b oscillations within the first 24 h of exposure (Fig. 2a). In contrast, a small but statistically significant augmentation of per2 did occur in response to HK-Spn at 12 and 16 h. Importantly, the gene expression pattern of these genes in the presence of HK-Spn does not show any changes in phase but rather augments the natural expression of these genes.

HK-Spn exposure augments the expression of per2, cry1a, per3, and per1b on zebrafish cells. (a) After entrainment, Z3 cells were exposed to PBS or heat-killed Streptococcus pneumoniae (HK-Spn) and maintained in constant darkness (DDDD). RNA samples were collected every 4 h throughout 48 h, and the relative transcript expression of per2, cry1a, per3, and per1b was measured. (b) Z3 cells were exposed to PBS or heat-killed Streptococcus pneumoniae (HK-Spn) under 12-hour low-intensity light (~ 4.12 × 1018 photons/s/m2) followed by constant darkness (LDDD). RNA samples were collected every 4 h throughout 48 h, and the relative transcript expression of per2, cry1a, per3, and per1b was measured. All time points were compared against time zero samples. Yellow bar: light period. Grey bar: dark period. Means ± SD are shown. Asterisks (*) indicate statistically significant differences *p < 0.05,** p < 0.01,***p < 0.001, ****p < 0.0001, Two-way ANOVA with Šídák’s multiple comparisons test. Three replicates per group, one experiment.
Because several reports in teleost have shown that light plays a differential role in how the clock interacts with microbes, we decided to analyze the exposure to HK-Spn in the context of light. As in Fig. 1, immediate increases in per2 and cry1a occurred in response to light, but surprisingly, exposure to HK-Spn further amplified the light-induced expression of per2 and cry1a (Fig. 2b). Moreover, the amplitude of cry1a expression was also augmented during the subsequent dark period (20–36 h), while per2 was not affected. In contrast, there was no substantial increase in amplitude in per3 and per1b expression in this condition, but small significant differences were seen at 4 (per3) and 32 h (per3 and per1b). Together, these results demonstrate that the sole presence of bacteria, in the absence of an infection, is sufficient to affect the expression of light-inducible (per2, cry1a) and non-inducible (per3, per1b) clock genes. Moreover, the expression patterns of HK-Spn augmentation follow the natural expression of these genes, although the effect seems to depend on light conditions present during bacteria exposure.
HK-Spn acutely augments per2, cry1a, and per3 in different light conditions
Given that per1b, per2, per3, and cry1a are part of a negative feedback loop that can affect their own expression, we decided to focus on the acute induction of genes within the first 4 h of exposure. At this time point, the changes observed are most likely to stem from the direct effects of HK-Spn exposure. Because the intensity of light affects the expression of per2 and cry1a14,18, and since the most prominent impact of HK-Spn exposure was seen in per2 light-driven expression (Fig. 2b), we next sought to determine whether HK-Spn would have the same effect at different light intensities. At 4 h, HK-Spn is insufficient to induce per2 or cry1a expression in the dark (Fig. 3). As in Fig. 2b, low-intensity light (~ 4.12 × 1018 photons/s/m2) induces per2 and cry1a expression (~ 3-fold and ~ 2.5-fold on average, respectively), which is further augmented in the presence of HK-Spn (~ 7.7-fold and ~ 4.7-fold on average, respectively). Notably, per2 and cry1a induction is strongly increased with high-intensity light (~ 3.96 × 1019 photons/s/m2; ~10.4-fold and ~ 4.4-fold, respectively), which is further magnified in the presence of HK-Spn (per2 ~ 21.7-fold and cry1a ~ 9.3-fold). These results show that immediately following HK-Spn exposure, per2 and cry1a are proportionally augmented by increasing light levels, but darkness impairs this acute activation.

Light intensity proportionally augments HK-Spn effect on per2 and cry1a but not in per3 and per1b. Z3 cells were entrained and exposed to HK-Spn under two light conditions, low-intensity light (Low-Int light,~4.12 × 1018 photons/s/m2) and high-intensity light (High-Int light, ~ 3.96 × 1019 photons/s/m2) for 4 h. Transcript expression of per2, cry1a, per3, and per1b was compared against time zero samples. Means ± SEM are shown. Asterisks (*) indicate statistically significant differences ** p < 0.01,****p < 0.0001, Two-way ANOVA with Tukey’s multiple comparisons test. Six experiments were conducted for dark and low-intensity light conditions, and three were conducted for high-intensity light conditions. Three replicates per group.
In the case of per3, neither low-intensity light, high-intensity light, nor HK-Spn alone was sufficient to induce expression (Fig. 3). However, when HK-Spn was added in the presence of light at either intensity, there was a significant effect (~ 1.3-fold increase). In contrast, per1b shows diminished expression during light exposure, consistent with reports that show inhibition by light60. Moreover, HK-Spn did not augment per1b expression at this time point, as seen in Fig. 2. These results suggest that light is needed for HK-Spn acute effects on per2, cry1a and per3. Hence, we propose that a common transcriptional activating factor generated by light allows HK-Spn-induced acute augmentation.
Exogenous H2O2 does not replicate light and HK-Spn combinatorial effects
Previous reports have described that light drives the expression of per2 and cry1a through the activation of D-box regions in their promoters16,17,18. Light induction of per2 and cry1a has also been related to the production of reactive oxygen species (ROS) by different light wavelengths, an effect mediated by activation of MAP kinases and D-boxes52,53. Moreover, H2O2 is sufficient to induce the expression of per2 and cry1a, which has also been suggested to occur through the activation of D-boxes52,53. In line with this, we wondered whether H2O2 is sufficient to mimic the effect of light and HK-Spn augmentation. To test this, we exposed Z3 cells to 300 µM H2O2 alone or in combination with HK-Spn in dark conditions and monitored gene expression after 4 h.
As expected, H2O2 induced robust expression of per2 and cry1a (Fig. 4). However, contrary to light, there was no combinatorial effect with HK-Spn. Moreover, H2O2 was not sufficient to induce per3 or per1b, alone or in combination with HK-Spn. This is consistent with previous reports indicating H2O2 activates D-boxes, regions absent in per1b and per352. These results indicate that H2O2 does not recapitulate the combinatorial effect of light and HK-Spn on per2, cry1a, and per3.

Hydrogen peroxide does not replicate light and HK-Spn effects. Relative expression of per2, cry1a, per3, and per1b in Z3 cells incubated with 300 µM hydrogen peroxide (H2O2) and HK-Spn for 4 h in the dark. Means ± SEM are shown. Asterisks (*) indicate statistically significant differences ****p < 0.0001, Two-way ANOVA with Tukey’s multiple comparisons test. Three experiments with three replicates per group.
Role of ROS in HK-Spn mediated effect on per2, cry1a, and per3 expression
Even though H2O2 was not sufficient to induce a combinatorial effect with HK-Spn, we wondered if the light-generated ROS may activate alternative pathways from exogenous H2O2 and thus contribute to the augmentation effect of HK-Spn. To test this, we treated Z3 cells with the antioxidant N-acetylcysteine (NAC) and evaluated gene expression at 4 h in light and dark conditions. NAC was not sufficient to inhibit light-mediated induction of per2 and cry1a (Fig. 5a), but instead, it significantly decreased the effect of HK-Spn augmentation of per2, cry1a, and per3. These results lead us to hypothesize that ROS generated in response to HK-Spn may be responsible for the augmentation of per2, cry1a, and per3. Unexpectedly, NAC induced per1b expression even in the absence of HK-Spn, suggesting that per1b expression may be sensitive to redox balance in a distinct manner.

Role of ROS in HK-Spn and light induction of clock genes. (a) Relative expression of per2, cry1a, per3, and per1b of Z3 cells exposed to HK-Spn and incubated in dark and low-intensity light conditions for 4 h. A 2-hour pre-treatment of 6 mM of N-acetyl cysteine (NAC) was used to suppress ROS production. Means ± SEM are shown. Asterisks (*) indicate statistically significant differences *p < 0.05, ***p < 0.001, ****p < 0.0001, Two-way ANOVA with Tukey’s multiple comparisons. Five to eight experiments, three replicates per group. (b) Dichlorodihydrofluorescein (DCF) fluorescence was measured after 2 h of PBS or HK-Spn exposure in dark and low-intensity light conditions. A 2-hour pre-treatment of 6 mM of N-acetyl cysteine (NAC) was used to suppress ROS production. Fold induction of relative fluorescent units was calculated against the dark PBS-treated cells. Means ± SEM are shown. Asterisks (*) indicate statistically significant differences ***p < 0.001,****p < 0.0001, Two-way ANOVA with Tukey’s multiple comparisons test. Four experiments, three replicates per group.
Because the innate immune detection of bacterial components often results in ROS production, we hypothesized that Z3 cells might generate ROS in response to HK-Spn exposure, contributing to the augmentation of per and cry genes. To test this, we analyzed the generation of ROS by the DCFH-DA (DCF) assay. DCFH-DA is a cell-permeable reagent that gets trapped in cells upon cleavage by intracellular esterases and fluoresces upon oxidation by intracellular ROS61,62. A pilot study to identify the best timepoint for analysis was conducted to analyze ROS production at 0.5, 1, 2, and 4 h post stimulation with HK-Spn. All timepoints showed similar patterns (i.e., fold differences between groups were similar at all timepoints) though, as reported previously in the literature, we also observed general increases in DCF baseline signal that correlates with incubation time. As we reasoned that changes in ROS would have to precede the activation of gene expression, we selected the 2-hour timepoint to analyze with sufficient replicates. Thus, after 2 h of stimulation with HK-Spn and LD exposure, cells were incubated with DCF, washed, and the fluorescent signal was measured at 490 nm excitation and 530 nm emission wavelengths. Fold induction was calculated against the basal levels of ROS detected in the dark (Fig. 5b). NAC, which inhibits ROS production, was used as a negative control. Exposure to light for 2 h did not induce a significantly higher DCF signal when compared to dark. Similarly, HK-Spn did not show detectable changes in the DCF-ROS signal in either dark or light conditions, but NAC treatment decreased ROS levels in all conditions. These results suggest that light and HK-Spn do not generate higher ROS production detectable by DCF in our conditions, suggesting NAC’s effects may be independent of ROS.
Discussion
Several studies have demonstrated changes in circadian gene expression during infection with bacteria, viruses, and parasites37,38,39,40,42,43,44. However, one question that has remained unanswered is whether this is due to direct effects of bacteria (or lack of bacteria) or indirect effects such as inflammation and damage from an infection. In this study, we demonstrate for the first time that the presence of inactive bacteria, specifically the Gram-positive Streptococcus pneumoniae, can induce changes in the expression of zebrafish genes from the repressor arm of the circadian clock suggesting that vertebrate hosts perceive and react to the presence of microbes. This finding is intriguing as our current models of circadian clocks do not provide any mechanistic reasons as to how or why the detection of bacteria would affect the core timekeeping machinery. Interestingly, these gene-specific effects are affected by the light conditions and the temporal expression of these genes. Here we show that heat-killed Streptococcus pneumoniae potently augmented the expression of per2 and cry1a in a manner synergistic with light and independently augments cry1a, per3, and per1b in constant darkness. This effect is surprising as several studies have demonstrated that exposure to bacterial PAMPs decreases the expression of clock genes38,39,40,45,46,47,48,49,50,51,63,64. In line with this, inflammatory compounds also decrease the amplitude of per3 oscillations in larval zebrafish65. It is clear that inflammatory and microbe recognition pathways can affect the circadian clock but these effects may vary depending on the time of exposure, the type of microorganism, and environmental conditions39,50,51,65,66.
Our results demonstrate different effects on clock gene expression depending on light exposure, an effect that is consistent with previous studies. In rainbow trout (Oncorhynchus mykiss), a skin infection with the ectoparasite Argulus foliaceus altered the expression and rhythmicity of clock genes in a manner dependent on light conditions42. Moreover, fish infected and maintained in constant light conditions had more parasite load after seven days of infection than those exposed to regular LD cycles. In a zebrafish model infected with tilapia lake virus, light availability significantly enhanced the antiviral immune response and changed the gene expression of clock genes (e.g., upregulation of arntl2 in the kidney in LD conditions)43. In a recent study, infection of zebrafish larvae with the gram-negative Klebsiella pneumoniae showed a general trending of upregulated clock gene expression in the light but downregulation in the dark44. These studies demonstrate that changes in clock expression are specific to both the pathogen and the light context during exposure, having consequences that can influence the disease outcome.
To investigate the combinatorial effects seen between light and HK-Spn exposure, we also examined the contribution of ROS in this pathway. Several reports have demonstrated a bidirectional interaction between ROS and clock genes in Drosophila, zebrafish, and mammalian systems4852,53,67,68,69,70,71,72,73. Moreover, light-generated ROS have been related to the light-driven expression of per2 and cry1a in zebrafish52,53. However, our results showed that in contrast to light, there is no synergistic effect between H2O2 and HK-Spn. This, together with the fact that low-intensity white light did not produce detectably higher amounts of ROS and light induction of per2 and cry1a was not reduced by a ROS inhibitor, indicates that in our conditions, light-generated ROS do not play a role in per2 and cry1a induction.
An intriguing fact is that the addition of the antioxidant NAC prevented HK-Spn from inducing per2, cry1a, and per3, suggesting a ROS-activated pathway in HK-Spn augmented response. Nonetheless, we were unable to detect a higher ROS signal in the presence of HK-Spn. A possible explanation could be that HK-Spn exposure generates a type of ROS not detectable by DCF as this assay presents a limitation in detecting different ROS species. DCF is oxidized by various radicalized ROS, including peroxyl, alkoxyl, hydroxyl, carbonate, nitrogen dioxide, and peroxynitrite, but can only detect cellular peroxides, such as H2O2, if they are decomposed to radicals61. Alternatively, the effects of NAC may be a consequence of other characteristics of this compound, such as a reducing agent, inhibitor of proteasome inhibitors, and suppressor of the transcription factor NF-κB, any of which may affect gene expression74,75,76. In PAC-2 cells, NAC was shown to block per2 and cry1a expression induced by blue light52; however, here we observed that in Z3 cells, NAC has no effect on light-induced cry1a and can even augment expression of per2. These differences could possibly be attributed to cell-specific effects and/or the use of different light wavelengths. Interestingly, the addition of NAC alone was sufficient to drive per1b expression in the light, an effect that has not been previously reported. It is uncertain if this effect results from NAC alone or a combinatorial effect of NAC and light, but it supports the idea that complex interactions exist between redox regulation and the circadian clock that remains unexplored.
The current understanding of the transcriptional regulation of per and cry genes in zebrafish is through promoter elements called E-boxes and D-boxes, driven by clock-related transcription factors such as Clock:Bmal and Tef, respectively12,13. Our initial thought about the regulation of HK-Spn effects was the activation of these regions by modulation of core components of the clock; however, we cannot discount the possibility that non-circadian transcription factors contribute to these effects. In mammals, transcription factors activated by microbial exposure, such as HIF-1⍺ and NF-κB, have been shown to directly interact with E-boxes or modify CLOCK:BMAL interaction with these regions, respectively77,78,79,80,81,82. Future studies analyzing the contribution of circadian and non-circadian transcription factors activated by HK-Spn may answer these questions. Additionally, other elements such as promoter architecture, such as the number of spacing of binding sites, or interactions between factors, as well as post-transcriptional modifications, could be at play in this regulation83,84,85. Other questions to be addressed in future studies will investigate the nature of the heat-resistant ligand from Spn, the effect of the exposure to other types of bacteria, and the identification of the receptors that contribute to this outcome. Regardless, the findings presented in this work provide new insights into the regulatory pathways of zebrafish circadian genes and present, for the first time, the impacts of an inactive bacteria on the molecular clock of the zebrafish. Understanding the interactions between different types of bacteria and the circadian clock can provide valuable insights into the intricate relationship between these systems.
Methods
Z3 cell culture
Z3 cells, a fibroblast-like embryonic cell line from zebrafish (Danio rerio)11, were maintained at room temperature in Z3 Media: Leibovitz L-15 medium (Gibco, 11415064) supplemented with 15% HyClone Fetal Bovine Serum (Cyvita, 16777-238), 2µM L-Glutamine (Sigma, G3126) and 1% Penicillin/Streptomycin (Sigma, P4333). Cells were seeded at 1 × 106, 5 × 105, or 5 × 104 cells/well into 6-, 12- or 96-well plates, respectively, grown until confluent and entrained in 12:12 LD cycle for three days using low-intensity white light (LED, 400–700 nm, ~ 4.12 × 1018 photons/s/m2). After entrainment, cells remained in the dark unless light was noted in the figure legend.
Light sources
Cells in light conditions were exposed to low-intensity (~ 500 lux, ~ 4.12 × 1018 photons/s/m2) or high-intensity (5000 lux, ~ 3.96 × 1019 photons/s/m2) white light (LED, 400–700 nm) for 4–12 h, as noted in the figure legend. Cells in dark conditions were kept at room temperature in constant darkness and covered in aluminum foil. Treatments of all groups and harvest of the dark condition groups were performed under indirect dim red light (LED, 600–750 nm, ~ 0–2 lux, ~ 1.67 × 1016 photons/s/m2), which does not induce per2.
Heat-killed bacteria
Heat-killed Streptococcus pneumoniae (HK-Spn) was prepared by growing wild-type D39 in Todd-Hewitt Broth (Thermo Scientific, CM0189B) with 2% Yeast Extract (Sigma, Y1625) at 37 °C with 5% CO2 until they reached an OD600 of 0.4. Colony-forming units were enumerated by serial dilution and plating on Columbia Blood Agar (Thermo Scientific, OXCM331B) with 5% Sheep’s Blood (Fisher Scientific, 50863755), and the remaining bacteria were then pelleted, resuspended in PBS, and heated to 100 °C for 1 h. HK-Spn was stored in aliquots at -20 °C until use. The effective dose of HK-Spn was determined empirically based on augmentation of per2 and cry1a after 4 h in light conditions. For all experiments shown here, the actual dose used was between 3.7 × 108 - 2.25 × 109 CFU/mL.
Bacterial exposure and pharmacological treatments
PBS, HK-Spn, and 300 µM H2O2 (Sigma, H1009) were applied after 12 h of the last entrainment cycle (time zero). N-Acetyl-L-cysteine (NAC) (Sigma, A9165) was added 2 h before time zero to a final concentration of 6 mM. All treatments were added directly to the cell media under dim red light. No media changes were made after entrainment.
RNA extraction and qRT-PCR
Cells were harvested in DNA/RNA Shield (Zymo, R110050) by scraping, flash-frozen in liquid nitrogen, and stored at -80 °C until extraction. The following products were used for qRT-PCR analysis: Quick-RNA Miniprep Kit (Zymo, R1055), iScript cDNA Synthesis (Bio-Rad, 1708891), PowerUp SYBR Green (Applied Biosystems, A25777), CFX Connect Real-Time thermocycler (Bio-Rad). Primers: actb1 (NM_131031) Fwd: ATCTTCACTCCCCTTGTTCAC; Rev: TCATCTCCAGCAAAACCGG, per1b (NM_212439) Fwd: TGCGCGTAATGGAGAGTATATG; Rev: CTTCGTTCAGTGGAGAGGTTC, per2 (NM_182857) Fwd: ACGAGGACAAGCCAGAGGAACG; Rev: GCACTGGCTGGTGATGGAGA, per3 (NM_131584) Fwd: CAAGTACAAGCAAACAGCGAG; Rev: ACTACCACAAAAGAGTCCGTG, cry1a (NM_001077297) Fwd: GGAGTGTGAACGCAGGAAG; Rev: AAACCCCTTAAGACTGGCAG. ΔΔCT was used to calculate relative mRNA expression86. Values were normalized to actb1 and compared to time zero samples.
Rhythmicity test
An eJTK rhythmicity test was performed on 48 h gene expression of per1b, per2, per3, and cry1a in constant darkness, using the online software BioDare2 (https://biodare2.ed.ac.uk/)87. Data were not detrended and were analyzed with the following settings: BD2 eJTK test method, and eJTK Classic analysis presets. p-value of < 0.01 was considered statistically significant.
Measurement of intracellular ROS
Cells were stimulated at time zero under dim red light, then maintained at room temperature in the dark or exposed to low-intensity light for 2 h. The following steps were all performed under dim red light: After treatments, cells were washed once with Hanks Balanced Salt Solution (HBSS) (Gibco, 14025092) and incubated for 45 min in 10 µM 2′,7′-Dichlorodihydrofluorescein diacetate (DCFH-DA) (Sigma, D6883). Cells were washed once with HBSS to remove extracellular DCFH-DA and placed in plain HBSS to read fluorescence at 490 nm excitation and 530 nm emission wavelengths using a Spectramax iD3 Multi-Mode Microplate Reader (Molecular Devices). Fold change was calculated by dividing the value of each sample by the average of the PBS dark control.
Statistical analysis
All experiments were performed thrice with three replicates unless denoted in the figure legend. Two-way ANOVA analyzed differences between two groups at different time points with Šídák’s multiple comparisons test. Two-way ANOVA with Tukey’s multiple comparisons was used to compare more than two groups and conditions.
Data availability
Data is available from the corresponding author upon reasonable request.
References
Marcheva, B. et al. In Circadian Clocks (eds Kramer, A. & Merrow, M.), Vol. 217 127–155 (Springer, 2013).
Scheiermann, C., Kunisaki, Y. & Frenette, P. S. Circadian control of the immune system. Nat. Rev. Immunol. 13, 190–198 (2013).
Patke, A., Young, M. W. & Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell. Biol. 21, 67–84 (2020).
Partch, C. L., Green, C. B. & Takahashi, J. S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 24, 90–99 (2014).
Boivin, D. B., Duffy, J. F., Kronauer, R. E. & Czeisler, C. A. Dose-response relationships for resetting of human circadian clock by light. Nature 379, 540–542 (1996).
Hannay, K. M., Forger, D. B. & Booth, V. Seasonality and light phase-resetting in the mammalian circadian rhythm. Sci. Rep. 10, 19506 (2020).
Jiménez, A., Lu, Y., Jambhekar, A. & Lahav, G. Principles, mechanisms and functions of entrainment in biological oscillators. Interface Focus 12, 20210088 (2022).
Mohawk, J. A., Green, C. B. & Takahashi, J. S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 35, 445–462 (2012).
Whitmore, D., Foulkes, N. S. & Strähle, U. Sassone-Corsi, P. Zebrafish clock rhythmic expression reveals independent peripheral circadian oscillators. Nat. Neurosci. 1, 701–707 (1998).
Whitmore, D., Foulkes, N. S. & Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature 404, 87–91 (2000).
Pando, M. P., Pinchak, A. B., Cermakian, N. & Sassone-Corsi, P. A cell-based system that recapitulates the dynamic light-dependent regulation of the vertebrate clock. Proc. Natl. Acad. Sci. U.S.A. 98, 10178–10183 (2001).
Vatine, G., Vallone, D., Gothilf, Y. & Foulkes, N. S. It’s time to swim! Zebrafish and the circadian clock. FEBS Lett. 585, 1485–1494 (2011).
Sacksteder, R. E., Kimmey, J. M. & Immunity Infection, and the zebrafish clock. Infect. Immun. 90, e00588–e00521 (2022).
Tamai, T. K., Young, L. C. & Whitmore, D. Light signaling to the zebrafish circadian clock by cryptochrome 1a. Proc. Natl. Acad. Sci. U.S.A. 104, 14712–14717 (2007).
Cermakian, N. et al. Light induction of a vertebrate clock gene involves signaling through blue-light receptors and MAP kinases. Curr. Biol. 12, 844–848 (2002).
Vatine, G. et al. Light directs zebrafish period2 expression via conserved D and E boxes. PLoS Biol. 7, e1000223 (2009).
Mracek, P. et al. Regulation of per and cry genes reveals a central role for the D-box enhancer in light-dependent gene expression. PLOS ONE 7, e51278 (2012).
Weger, B. D. et al. The light responsive transcriptome of the zebrafish: function and regulation. PLoS ONE 6, e17080 (2011).
Agrawal, P. et al. Drosophila CRY entrains clocks in body tissues to light and maintains Passive membrane properties in a non-clock body tissue Independent of Light. Curr. Biol. 27, 2431–2441e3 (2017).
Ozturk, N. Light-dependent reactions of animal circadian photoreceptor cryptochrome. FEBS J. 289, 6622–6639 (2022).
Abele, S. H., Meadows, K. E., Medeiros, D. & Silver, A. C. Time is on the immune system’s side, yes it is. Yale J. Biol. Med. 92, 225–231 (2019).
Ince, L. M. et al. Influence of circadian clocks on adaptive immunity and vaccination responses. Nat. Commun. 14, 476 (2023).
Edgar, R. S. et al. Cell autonomous regulation of herpes and influenza virus infection by the circadian clock. Proc. Natl. Acad. Sci. U.S.A. 113, 10085–10090 (2016).
Lundy, S. R. et al. Effect of time of day of infection on chlamydia infectivity and pathogenesis. Sci. Rep. 9, 11405 (2019).
Borrmann, H., McKeating, J. A. & Zhuang, X. The circadian clock and viral infections. J. Biol. Rhythms 36, 9–22 (2021).
Mul Fedele, M. L., Senna, C. A., Aiello, I., Golombek, D. A. & Paladino, N. Circadian rhythms in bacterial sepsis pathology: What we know and what we should know. Front. Cell. Infect. Microbiol. 11, 773181 (2021).
Bellet, M. M. et al. Circadian clock regulates the host response to Salmonella. Proc. Natl. Acad. Sci. U.S.A. 110, 9897–9902 (2013).
Stone, E. F. et al. The circadian clock protein Timeless regulates phagocytosis of Bacteria in Drosophila. PLoS Pathog. 8, e1002445 (2012).
Gu, F. et al. Total and cause-specific mortality of U.S. nurses working rotating night shifts. Am. J. Prev. Med. 48, 241–252 (2015).
Loef, B. et al. Shift work and respiratory infections in health-care workers. Am. J. Epidemiol. 188, 509–517 (2019).
Long, J. E. et al. Morning vaccination enhances antibody response over afternoon vaccination: A cluster-randomised trial. Vaccine 34, 2679–2685 (2016).
Wang, W. et al. Time of day of vaccination affects SARS-CoV-2 antibody responses in an observational study of health care workers. J. Biol. Rhythms. 37, 124–129 (2022).
Hazan, G. et al. Biological rhythms in COVID-19 vaccine effectiveness in an observational cohort study of 1.5 million patients. J. Clin. Invest. 133, e167339 (2023).
Lewis, A. J. et al. Blue light enhances bacterial clearance and reduces organ injury during sepsis*. Crit. Care Med. 46, e779–e787 (2018).
Feigin, R. D., Joaquin, V. H. S., Haymond, M. W. & Wyatt, R. G. Daily periodicity of susceptibility of mice to pneumococcal infection. Nature 224, 379–380 (1969).
Kitchen, G. B. et al. The clock gene Bmal1 inhibits macrophage motility, phagocytosis, and impairs defense against pneumonia. Proc. Natl. Acad. Sci. U.S.A. 117, 1543–1551 (2020).
Li, T. et al. H. pylori infection induced BMAL1 expression and rhythm disorder aggravate gastric inflammation. EBioMedicine 39, 301–314 (2019).
Benegiamo, G. et al. Mutual antagonism between circadian protein period 2 and Hepatitis C virus replication in hepatocytes. PLoS ONE 8, e60527 (2013).
Rijo-Ferreira, F. et al. Sleeping sickness is a circadian disorder. Nat. Commun. 9, 62 (2018).
Midttun, H. L. E., Vindas, M. A., Whatmore, P. J., Øverli, Ø. & Johansen, I. B. Effects of Pseudoloma neurophilia infection on the brain transcriptome in zebrafish (Danio rerio). J. Fish Dis. 43, 863–875 (2020).
Du, L. Y. et al. The innate immune cell response to bacterial infection in larval zebrafish is light-regulated. Sci. Rep. 7, 12657 (2017).
Ellison, A. R., Wilcockson, D. & Cable, J. Circadian dynamics of the teleost skin immune-microbiome interface. Microbiome 9, 222 (2021).
Mazur, M. et al. Effects of light and circadian clock on the antiviral immune response in zebrafish. Fish Shellfish Immunol. 140, 108979 (2023).
Ding, H. et al. Klebsiella pneumoniae alters zebrafish circadian rhythm via inflammatory pathways and is dependent on light cues. Heliyon 10, e30829 (2024).
Okada, K. et al. Injection of LPS causes transient suppression of biological clock genes in rats. J. Surg. Res. 145, 5–12 (2008).
Yamamura, Y., Yano, I., Kudo, T. & Shibata, S. Time-dependent inhibitory effect of lipopolysaccharide injection on PER1 and PER2 gene expression in the mouse heart and liver. Chronobiol. Int. 27, 213–232 (2010).
Curtis, A. M. et al. Circadian control of innate immunity in macrophages by miR-155 targeting Bmal1. Proc. Natl. Acad. Sci. U.S.A. 112, 7231–7236 (2015).
Wang, Y. et al. Endotoxin disrupts circadian rhythms in macrophages via reactive oxygen species. PLoS ONE 11, e0155075 (2016).
Shimizu, T., Watanabe, K., Anayama, N. & Miyazaki, K. Effect of lipopolysaccharide on circadian clock genes Per2 and Bmal1 in mouse ovary. J. Physiol. Sci. 67, 623–628 (2017).
Chen, S., Fuller, K. K., Dunlap, J. C. & Loros, J. J. A Pro- and anti-inflammatory axis modulates the macrophage circadian clock. Front. Immunol. 11, 867 (2020).
Silver, A. C. Pathogen-associated molecular patterns alter molecular clock gene expression in mouse splenocytes. PLoS ONE 12, e0189949 (2017).
Pagano, C. et al. Evolution shapes the responsiveness of the D-box enhancer element to light and reactive oxygen species in vertebrates. Sci. Rep. 8, 13180 (2018).
Hirayama, J., Cho, S. & Sassone-Corsi, P. Circadian control by the reduction/oxidation pathway: Catalase represses light-dependent clock gene expression in the zebrafish. PNAS 104, 15747–15752 (2007).
Hirayama, J., Cardone, L., Doi, M. & Sassone-Corsi, P. Common pathways in circadian and cell cycle clocks: Light-dependent activation of Fos/AP-1 in zebrafish controls CRY-1a and WEE-1. Proc. Natl. Acad. Sci. U.S.A. 102, 10194–10199 (2005).
Ben-Moshe, Z. et al. The light-induced transcriptome of the zebrafish pineal gland reveals complex regulation of the circadian clockwork by light. Nucleic Acids Res. 42, 3750–3767 (2014).
Moore, H. A. & Whitmore, D. Circadian rhythmicity and light sensitivity of the zebrafish brain. PLoS ONE 9, e86176 (2014).
Ramos, B. C. R., Moraes, M. N. C. M., Poletini, M. O., Lima, L. H. R. G. & Castrucci, A. M. L. From blue light to clock genes in zebrafish ZEM-2S cells. PLoS ONE 9, e106252 (2014).
Mracek, P. et al. ERK signaling regulates light-induced gene expression via D-box enhancers in a differential, wavelength-dependent manner. PLoS ONE 8, e67858 (2013).
Steindal, I. A. F. & Whitmore, D. Zebrafish circadian clock entrainment and the importance of broad spectral light sensitivity. Front. Physiol. 11, 1002 (2020).
Vallone, D., Gondi, S. B., Whitmore, D. & Foulkes, N. S. E-box function in a period gene repressed by light. PNAS 101, 4106–4111 (2004).
Eruslanov, E. & Kusmartsev, S. In Advanced Protocols in Oxidative Stress II (ed. Armstrong, D.), Vol. 594 57–72 (Humana Press, 2010).
Gardiner, B. et al. In Measuring Oxidants and Oxidative Stress in Biological Systems (eds. Berliner, L. J. & Parinandi, N. L.), Vol. 34 39–60 (Springer International Publishing, 2020).
Rijo-Ferreira, F. & Takahashi, J. S. Circadian rhythms in infectious diseases and symbiosis. Semin. Cell Dev. Biol. 126, 37–44 (2022).
Huang, H. et al. Immunological and inflammatory effects of infectious diseases in circadian rhythm disruption and future therapeutic directions. Mol. Biol. Rep. 50, 3739–3753 (2023).
Mosser, E. A. et al. Identification of pathways that regulate circadian rhythms using a larval zebrafish small molecule screen. Sci. Rep. 9, (2019).
Spišská, V., Kubištová, A., Novotný, J. & Bendová, Z. Impact of prenatal LPS and early-life constant light exposure on circadian gene expression profiles in various rat tissues. Neuroscience. 551, 17–30 (2024).
Krishnan, N., Davis, A. J. & Giebultowicz, J. M. Circadian regulation of response to oxidative stress in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 374, 299–303 (2008).
Lai, A. G. et al. CIRCADIAN CLOCK-ASSOCIATED 1 regulates ROS homeostasis and oxidative stress responses. Proc. Natl. Acad. Sci. U.S.A. 109, 17129–17134 (2012).
Tamaru, T. et al. ROS stress resets circadian clocks to coordinate pro-survival signals. PLoS ONE 8, e82006 (2013).
Schippers, J. H., Lai, A. G. & Mueller-Roeber, B. Dijkwel, P. P. Could ROS signals drive tissue-specific clocks? Transcription 4, 206–208 (2013).
Tahara, Y. et al. In vitro and in vivo phase changes of the mouse circadian clock by oxidative stress. J. Circadian Rhythm. 14, 4 (2016).
Arthaut, L. D. et al. Blue-light induced accumulation of reactive oxygen species is a consequence of the Drosophila cryptochrome photocycle. PLoS ONE 12, e0171836 (2017).
Chen, A. et al. Circadian clock1a coordinates neutrophil recruitment via nfe212a/duox-reactive oxygen species pathway in zebrafish. Cell. Rep. 42, 113179 (2023).
Zafarullah, M., Li, W. Q., Sylvester, J. & Ahmad, M. Molecular mechanisms of N-acetylcysteine actions. Cell. Mol. Life Sci. (CMLS) 60, 6–20 (2003).
Halasi, M. et al. ROS inhibitor N -acetyl- L -cysteine antagonizes the activity of proteasome inhibitors. Biochem. J. 454, 201–208 (2013).
Tenório, M. C. D. S., Graciliano, N. G., Moura, F. A., Oliveira, A. C. M. D. & Goulart, M. O. F. N-acetylcysteine (NAC): Impacts on human health. Antioxidants 10, 967 (2021).
Cavadini, G. et al. TNF-α suppresses the expression of clock genes by interfering with e-box-mediated transcription. Proc. Natl. Acad. Sci. U.S.A. 104, 12843–12848 (2007).
Brocato, J., Chervona, Y. & Costa, M. Molecular responses to hypoxia-inducible factor 1 α and beyond. Mol. Pharmacol. 85, 651–657 (2014).
Wu, Y. et al. Reciprocal regulation between the circadian clock and hypoxia signaling at the genome level in mammals. Cell Metabol. 25, 73–85 (2017).
Shen, Y. et al. NF-κB modifies the mammalian circadian clock through interaction with the core clock protein BMAL1. PLoS Genet. 17, e1009933 (2021).
Jantsch, J. et al. Toll-like receptor activation and hypoxia use distinct signaling pathways to stabilize hypoxia-inducible factor 1α (HIF1A) and result in differential HIF1A-dependent gene expression. J. Leukoc. Biol. 90, 551–562 (2011).
Rahman, M. M. & McFadden, G. Modulation of NF-κB signalling by microbial pathogens. Nat. Rev. Microbiol. 9, 291–306 (2011).
Gordân, R. et al. Genomic regions flanking E-box binding sites influence DNA binding specificity of bHLH transcription factors through DNA shape. Cell. Rep. 3, 1093–1104 (2013).
Nakahata, Y. et al. A direct repeat of e-box-like elements is required for cell-autonomous circadian rhythm of clock genes. BMC Mol. Biol. 9, 1 (2008).
Cox, K. H. & Takahashi, J. S. Circadian clock genes and the transcriptional architecture of the clock mechanism. J. Mol. Endocrinol. 63, R93–R102 (2019).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 25, 402–408 (2001).
Zieliński, T., Hay, J. & Millar, A. J. In Plant Circadian Networks (eds Staiger, D., Davis, S. & Davis, A. M.), Vol. 2398, 15–32 (Springer US, 2022).
Acknowledgements
This work was supported by the National Institutes of Health / NIGMS (R35GM147509). J.M.K. is a Pew Scholar in the Biomedical Sciences, supported by The Pew Charitable Trusts. We thank Prof. Carrie Partch for the kind donation of Z3 cells.
Author information
Authors and Affiliations
Contributions
CMF and RES performed the experiments. CMF, RES and JMK designed the experiments. CMF and RES wrote the paper with additional edits from AGD and JMK. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Morales Fénero, C., Sacksteder, R.E., Diamos, A.G. et al. Heat-inactivated Streptococcus pneumoniae augments circadian clock gene expression in zebrafish cells. Sci Rep 14, 27805 (2024). https://doi.org/10.1038/s41598-024-78888-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-78888-0
