
Abstract
Since therapeutic options are limited the utilization of prebiotics is suggested to prevent food allergies (FAs). Using an experimental peach allergy model we explored the effect of dietary fiber pectin, a high-methoxyl heteropolysaccharide, on the manifestation of FA. CBA/J mice were sensitized, subsequently orally boosted and provoked with peach peel extract. For dietary intervention, mice were fed a pectin containing diet before (primary-preventive) or after (secondary-preventive) sensitization. Non-treated allergic and sham-treated mice were fed a diet containing 20% cellulose. Fecal microbiota, humoral and intestinal immune cell responses were analyzed. Pectin remarkably affected the gut microbiota composition and diversity, promoting mainly the growth of Bacteroides. The frequency of mast cells, macrophages, and CD3+T cells in the lamina propria of the small intestine was reduced, whereas the frequency of B cells and CD4+T cell subpopulation was enhanced. Pectin intervention in the primary-preventive stetting significantly triggered serum IgA levels, whereas production of IgE and mMCPT-1 was reduced. Remarkably, in both settings peach allergen-specific IgG1/IgG2a ratio and specific IgE were significantly reduced to baseline. The data suggest, that dietary supplementation of pectin in both intervention approaches can diminish inflammatory responses and signs of allergic immune responses, accompanied by alteration of the gut microbiota composition.
Similar content being viewed by others

Introduction
Food allergy (FA) is a potentially life-threatening immune reaction with increasing prevalence worldwide1. FAs are typically IgE-mediated type I hypersensitivity reactions, characterized by a type 2 (T2) inflammation accompanied by the induction of T helper type 2 (TH2) cells. The activation of mast cells and basophils leads to local inflammatory and systemic reactions which may even accumulate to life-threatening anaphylactic reactions2. Peach allergy is a very frequent fruit allergy in Japan and especially in the Mediterranean area. The clinical manifestation can range from mild oral symptoms to anaphylaxis depending on the geographical distribution and the sensitization profile3. Remarkably, the reason for the geographical restriction is still unknown and immunotherapeutic strategies are not available.
Different therapeutic options such as oral immunotherapy (OIT) or sublingual immunotherapy (SLIT), which aimed to induce desensitization or peripheral tolerance, showed promising results in a limited number of studies4,5,6. So far, the only licensed product for OIT of food allergy has been approved by the US Food and Drug Administration for treatment of peanut allergy7. Thus, the recommended management of FAs is still symptomatic treatment, the avoidance of the allergenic food or a healthy diet, e.g. using prebiotics8.
Since the dysbiosis of gastrointestinal (GI) microbiota has been recently associated as a main risk factor for development of allergic airway inflammation and FA9,10,11 targeting the GI-microbiota to retain homeostasis has been discussed as preventive option for FA12,13,14. Particularly, dietary fibers, which are not digested by GI-enzymes but can be degraded by commensal bacteria in the colon, gained attention due to their ability to affect GI-microbiota composition, avoiding dysregulation, and promoting production of bacteria-derived metabolites with immune-modulating properties15,16,17. In line with this, the dietary fiber pectin has been reported to promote several additional health benefits as the improvement of physical bowel function, maintenance of blood cholesterol and glycemic response15,18,19,20. Pectin is a dietary fiber which promotes the growth of health-promoting bacteria21. Bacterial fermentation of pectin, like dietary fibers, is well known to release different metabolic products like short chain fatty acids (SCFAs), including acetate, propionate and butyrate which mediate immune-modulatory effects22,23,24. Two health claims have been accepted for pectin by the EU: 1st reduction of the blood glucose rise after meals and 2nd maintenance of normal blood cholesterol levels after consumption of at least 6–10 g pectin per meal25. Early studies on allergic asthma revealed beneficial effects of pectin supplementation in mice due to modulation of the microbiota, with increased levels of circulating SCFAs and protection against the development of allergic inflammation in the lung26,27. However, reports on the potential prophylactic and therapeutic effects of pectin in experimental food allergy models and the associated effects on the GI-immune response in a TH2-biased allergic setting are limited.
Previous own and independent studies showed that the chemical structure of pectin, which is characterized by the molecular mass, carbohydrate composition of the backbone and branches, degree of esterification and blockiness, impacts its fermentation efficacy22,28,29,30. Data provide evidence that pectin with high degree of methyl-esterification (HMP) exerts strongest effects on the immune response in naïve mice28,30. Therefore, the present study aimed to investigate the effect of dietary intervention with HMP derived from apple on the development of clinical signs, inflammation, as well as local and humoral immune responses in a peach allergy mouse model.
Results
Pectin supplementation did not prevent the development of allergy-related clinical signs
Two different intervention approaches, a primary-preventive (prophylactic) and secondary-preventive (therapeutic) treatment strategy, were compared in a mouse model of peach allergy (Fig. 1). Monitoring of body weight gain and food intake after the course of the experiments (26 days) did not reveal significant differences between the treatment groups and between treatment and control groups (Figure S1). Body core temperature was monitored up to 30 min after provocation with peach extract (PE) at day 26 (Fig. 2a). A significant temperature drop could be observed 15 min after the provocation in all PE exposed groups in comparison to the non-sensitized PBS control. However, the results did not show any difference between both, pectin treated mice and the allergic control (Fig. 2b). Noteworthy, mice with a temperature drop of more than 2 °C were sacrificed according to the animal guidelines. In addition, allergy-related clinical signs were examined in a blinded manner, including behavioral changes, consistency of the stool as well as ruffled fur (Table S1). For all PE-sensitized groups (with and without pectin treatment) mild allergy-related signs were observed after the 2nd oral boost, slightly increasing by the 3rd boost (Fig. 2c). The strongest symptoms were observed after systemic provocation with PE, demonstrating the allergy model was successfully established. However, only a slight reduction of the overall symptom score could be observed in the groups supplemented with pectin compared to the allergic control.

Experimental schedule of pectin intervention in peach allergic mice. Mice were sensitized intraperitoneally (i.p.) using peach peel extract (PE) three times (d0, d7 and d12), followed by oral (i.g.) boost of PE three times and finally mice were subjected to provocation with i.p. injection of PE. For primary-preventive pectin intervention, pectin feeding (15% HMP plus 5% cellulose) started seven days before sensitization. Secondary-preventive intervention started after sensitization of the mice. PBS and allergic controls were fed control diet (20% cellulose) during the course of the experiment. Feces samples were taken at indicated time points (*).

Allergy-related signs and symptoms after provocation. (a) Body temperature was measured before (-15 min; baseline) and up to 30 min after i.p.-provocation. (b) Body temperature of individual mice 15 min after i.p.-provocation and (c) symptom score after oral exposition and provocation. n = 3–5; *p < 0.05; **p < 0.01; ***p < 0.001, one-way ANOVA and two-way ANOVA were used in b and c, respectively.
Pectin consumption altered size of intestine and gut microbiota composition in peach allergic mice
The effect of pectin intervention on both, the intestinal tissue and gut microbiota composition was evaluated.
The length of different intestinal sections (small and large intestine and caecum) was measured and histological evaluation was performed (Figure S2a–c). Results showed that the length of the small intestine was significantly reduced in the primary-preventive pectin group (Figure S2a), whereas the length of the large intestine and caecum were significantly enlarged in both pectin intervention groups when compared to the allergic control (Figure S2b,c).
To determine, whether pectin intervention induced alterations of the gut microbiota composition, feces samples were collected at different time points (as indicated in Fig. 1) and analyzed by metagenomics 16S rRNA gene sequencing (Fig. 3). The standard cellulose diet (in both control groups and secondary-preventive group) resulted in a reduction in microbial richness as indicated by the Chao 1 index at d-2 compared to d-7. (Fig. 3a).

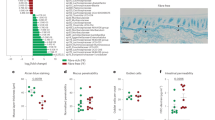
Alteration of gut microbiota composition. (a) Microbiota diversity (Chao 1) and (b) relative frequency of bacterial genera was analyzed in feces of the mice of the four treatment groups at different time points (d-7, d-2, d12, d16, d26).
Pectin intervention led to a more prominent and persistent reduction of the microbial richness, especially in the primary-preventive approach 5 days after starting pectin exposure (at d-2) and in the therapeutic approach 4 days after starting the pectin exposure (at d16). In contrast, data provide evidence that allergic sensitization (from d0 to d12) without the simultaneous exposure to pectin promoted the microbial richness in the allergic control group and the secondary-preventive group.
Above changes in the microbiome complexity correlated with a substantial shift of the bacterial composition towards Bacteroides spp., which was observed already 4–5 days after start of the pectin diet (d-2 for primary-preventive, d16 for secondary-preventive) (Fig. 3b; S3). In contrast to the increase of Bacteroides spp. in response to the pectin intervention, the relative frequency of bacteria belonging to this genus seemed to be reduced in the mice of the allergic control after allergic sensitization (d12). Moreover, the frequency of Parabacteroides spp. decreased after pectin treatment when compared to the allergic control, and the frequency of Akkermansia spp. was strongly reduced after start of pectin intervention, whereas the levels stayed constant in the allergic control group (Figure S3). Noteworthy, the pectin induced change of the composition of microbiota was consistent for all individual mice of a group (Figure S4). In contrast, the relative frequency of genus Blautia slightly increased after introduction of pectin in both treatment groups (d16 vs d12 in the secondary-preventive and d-2 vs d-7 in the primary-preventive group) (Table S4).
Pectin intervention modulated the local immune response in the intestinal tract
Considering the changes observed in the intestine size and gut microbiota after pectin intervention, the possible effects on immune cells in the lamina propria were analyzed by histology and FACS.
Histological analysis of the small intestine revealed slightly dilated lumen of the allergic control group, with no significant impact on inflammatory infiltration. Both pectin intervention groups showed dilated lumen, slightly shorter villi and minimal inflammatory infiltrate. Mild apical edema was observed in the secondary prevention group. No strong histological differences between the treatment groups could be observed. Other histopathological alterations as hypertrophic goblet cells, hypertrophic Paneth or thicker muscle layer were not detected in any sample (Figure S5).
FACS analysis of the cells from GI tissue showed a significant decrease of levels of mast cells and macrophages in both pectin intervention groups, compared to the allergic control (Fig. 4a). No clear differences were observed in the frequency of neutrophils, cDCs or eosinophils. Interestingly, when mice received pectin upon sensitization (secondary-preventive approach), pectin induced increased levels of B cells (Fig. 4b) and reduced the high frequency of T cells in the allergic setting back to normal levels of non-allergic mice. Further evaluation of T cell subpopulations revealed that pectin modulated differently the T cell subtypes. An enhanced frequency of CD4+T cells by both approaches in comparison with the allergic control was observed. In contrast, only primary-preventive pectin intervention showed a reduction on CD8+CTLs while no effect on the amount of Tregs (CD4+Fox p 3+) could be observed between the treatment groups.

Frequency of immune cells in the small intestine lamina propria. The small intestine was enzymatically treated and isolated lamina propria immune cells were analyzed via flow cytometry. Granulocytes (a): mast cells (CD45+CD117+), macrophages (CD45+CD11c+), neutrophils (CD45+Ly6G+), cDCs (CD45+CD11c+) and eosinophils (CD45+Siglec F+). Leucocytes (b): B cells (CD45+CD19+), T cells (CD45+CD3+), CTLs (CD45+CD3+CD8+), CD4 + T cells (CD45+CD3+CD4+) and Tregs (CD45+CD3+CD4+Foxp3+). n = 3–5; *p < 0.05; **p < 0.01; ***p < 0.001, one-way ANOVA was used.
Pectin intervention affects mMCPT-1 secretion
Next, the levels of mouse mast cell protease-1 (mMCPT-1), a mast cell activation marker31 were analyzed in serum (Fig. 5a) and in the small intestine homogenates (Fig. 5b). As expected, mMCPT-1 was significantly enhanced in serum and intestinal tissue in the allergic mice in comparison to the sham control mice. Although not significant, both pectin intervention approaches provided evidence for a slight reduction of mMCPT-1 level in the intestinal homogenate (Fig. 5b). Whereas the effect of pectin on serum derived mMCPT-1 in the therapeutic setting was negligible, mMCPT-1 was significantly suppressed when pectin was applied in the primary-preventive approach. Here, mMCPT-1 levels were diminished close to the baseline level of non-allergic mice (Fig. 5a).

Determination of humoral and local mMCPT-1 levels upon pectin intervention. Levels of mMCPT-1 were measured in (a) serum [mMCPT-1 µg/ml] and (b) the small intestine homogenates [mMCPT-1 µg/5 mg protein/ml] via ELISA. n = 3–5; *p < 0.05, **p < 0.01, one-way ANOVA was used.
Pectin intervention suppressed manifestation of TH2-related antibody responses
Finally, the effect on the humoral immune response in the serum of the mice was evaluated, by the determination of the values of total and allergen-specific immunoglobulins (Fig. 6 and 7).

Immunoglobulin levels in serum. Levels of (a) total IgE, (b) total IgG and (c) total IgA were analyzed in the serum of the mice after provocation via ELISA. (d) Ratio of total IgG/IgE were analyzed. n = 3–5; *p < 0.05; **p < 0.01; ***p < 0.001, one-way ANOVA was used.

Pru p 3- and PE-specific antibody response in the serum. Levels of PE-specific (a) IgE, (b) IgG1 and (c) IgG2a, as well as Pru p 3-specific (d) IgE, (e) IgG1 and (f) IgG2a were analyzed in the serum of the mice after provocation via ELISA. Ratio of (g) PE-specific and (h) Pru p 3-specific IgG1 to IgG2a values were determined for baseline and final time point, respectively. n = 3–5; *p < 0.05; **p < 0.01; ***p < 0.001, one-way ANOVA was used.
Interestingly, the primary-preventive approach significantly reduced the total IgE levels, while this effect was only moderate in the secondary-preventive approach (Fig. 6a) Pectin supplementation did not significantly alter levels of total IgG compared to the allergic control (Fig. 6b). However, when comparing the ratio of total IgG to total IgE, primary-preventive pectin supplementation significantly shifted the ratio towards IgG (Fig. 6c). Analysis of total IgA levels showed a strong decrease in the allergic control compared to the PBS treated mice, nevertheless, both pectin intervention approaches restored the total IgA levels to baseline (Fig. 6d).
Considering the modulatory effect of pectin intervention observed on the total immunoglobulin levels in serum and the modulatory effect of pectin on allergy-related immune response, allergen-specific antibody levels were determined in the serum of the mice after provocation (Fig. 7). The results indicate a strong increase of Pru p 3-specific IgE levels in the mice of the allergic control group that could be significantly reduced by pectin intervention. In comparison, only the primary-preventive intervention approach induced decreased levels of Pru p 3-specific IgG1, whereas the secondary-preventive intervention approach enhanced Pru p 3-specific IgG2a levels. Comparable effects could be observed for the PE-specific antibodies. In line with this, pectin interventions decreased the ratio of Pru p 3 and/or PE-specific IgG1/IgG2a compared to the allergic control.
Discussion
Dietary fiber can regulate intestinal microbiome, can affect the immune responses in the gut and airways, and are important for the TH1/TH2 homeostasis27. Among dietary fibers pectin has been described to provide several beneficial properties by altering the GI-microbiota composition15,18,19,20,32.
Several studies have already shown a suppressive role on allergy manifestation by increasing the production of bacteria-derived SCFAs. SCFA like butyrate, propionate and acetate have been shown to mediate a reduced risk of allergic reactions24,26,33,34,35,36,37. The underlying mechanism of how dietary fibers can modulate the allergic asthma is reviewed by Verstegen and colleagues23. However, the role of pectin intervention on the manifestation of FA and the underlying immune response remains to be elucidated. Therefore, the present study aimed to explore the modulatory effect of pectin on gut microbiota composition, and its impact on the local and systemic immune reactions in a peach allergy mouse model.
Although pectin intervention was not sufficient to prevent the drop in the core body temperature of the mice after provocation, the clinical symptoms were partially reduced in the treated mice compared to the allergic controls. Intraperitoneal allergen provocation induced a drastic temperature drop of > 2 °C, a criterion for termination of the experiment, underlining the strong allergic response in the mouse model of peach allergy used in this study. However, systemic provocation does not replicate the natural intake of a food allergens via ingestion, where the allergens encounters the immune system via the GI-mucosa. The GI-tract being supposedly the location where pectin primarily impacts the immune system. Thus, it would be interesting to monitor the effect of pectin in a murine model of peach allergy with provocation via intragastric application and/or to increase pectin dosage. However, pectin ingestion has substantial effect in the GI-microbiota and general immune responses.
Interestingly, in the allergic control group the relative frequency of Bacteroides spp. was reduced upon allergic sensitization. In both, the primary-preventive and the secondary-preventive approach pectin intervention modulated the composition of the gut microbiota in an established allergic setting. This was evidenced by a strong shift towards Bacteroides spp. as early as 4 days after starting the pectin diet, resulting in higher frequencies of Bacteroides spp. (appr. 45–60%) than the non-allergic controls (appr. 15–40%). The results are in agreement with previous own data30 and independent report21. Members of the Bacteroides genus are well-known effective pectin-degraders due to the expression of carbohydrate-active enzymes (CAZymes)38. The generated pectin-derived oligosaccharides can be further utilized by bacteria to affect microbiota composition39. In addition, Bacteroides spp. are known to produce SCFAs as acetate, propionate and butyrate40, which are reported to dampen allergic airway TH2 responses, reduce inflammatory responses in vitro and in vivo41,42, and to the protection against development of experimental allergic airway inflammation by promoting the activation of Tregs and inhibiting TH2 response without triggering a TH1 response43. In line with this, a reduction of Bacteroides spp. in the intestinal microbiota has been associated with a higher risk of FA26,44. In contrast to the literature45, the frequency of the genus Parabacteroides was reduced after pectin intervention, which probably is a consequence of the efficient utilization of the pectin by Bacteroides, promoting its predominant abundance and reducing the diversity as indicated by the Chao 1 index. A similar reduction after pectin intervention was observed for the genus Akkermansia. The implications of this difference are still under discussion. Akkermansia spp. are considered to promote beneficial health effects46, whereas other publications have reported that Akkermansia spp. are pathobionts exacerbating the development of inflammation and food allergy in fiber-deprived mice by increasing the intestinal permeability47, or promoting the development of other pathobionts48. Despite Blautia spp. are known to produce SCFAs49 and is considered to have beneficial effects promoting intestinal health, reducing inflammation, metabolic diseases and antibacterial activity the frequency of Blautia spp. was not substantially enhanced after pectin supplementation, which does not confirm previous reports50.
The present study did not aim to analyze the role of SCFAs and other bacterial-derived metabolites, rather than to investigate the pectin-induced effects on the local and systemic immune response in relation to the microbiota under allergic conditions. Actually the production of bacteria-derived 2-methylbutyric acid in response to feeding of naïve mice with the same pectin has been reported in own previous study30.
Pectin intervention strongly affects the local cellular immune response in the small intestine, as observed by the variations in the frequency of immune cells in the intestinal lamina propria. Reduced frequencies of mast cells, macrophages, T cells, and CTLs were observed, particularly in the primary-preventive pectin intervention approach. In contrast, at the same time the frequency of B cells and CD4 + T cells was enhanced. Although the effects on TH1- and TH2-promoting cytokines in gastrointestinal tissue were not investigated in the study, the reduced mMCPT-1 levels provide some evidence of a reduced anti-inflammatory response. The results support protective effects of pectin supplementation on inflammatory responses and the manifestation of allergic reactions. Mast cell numbers are increased in IgE-dependent allergies which is correlated with the release of a variety of mediators as proteases, cytokines, lipid mediators and histamine, that lead to the recruitment of leukocytes, vasodilatation and characteristic inflammatory responses51,52. Mast cells in the intestinal mucosa are very well known effector cells in IgE-mediated FAs. Studies using mast cell-deficient mice have shown that a reduction of this cell type usually translates to a reduction in the allergic response and sensitization status53. In our study, pectin intervention reduced the frequency of mast cells in the small intestine of peach allergic mice, and in addition also reduced their activation status evidenced by the mMCPT levels. This supports the reduced development of clinical signs after provocation.
Remarkably, pectin intervention was also capable to modulate the humoral immune response. Under allergic condition mice showed enhanced allergen-specific IgE- and IgG1 values52,54, whereas total IgA response was reduced. In the same context, reduced IgA levels have been associated to a higher risk of allergic disease54,55,56. Interestingly, pectin intervention was able to reverse this effect, restoring the levels of specific and total IgE and IgA to the baseline level of non-allergic mice, while no clear effect was observed in the total IgG levels. However, when the ratio IgG/IgE was evaluated, a shift towards IgG was observed. Pectin has been shown to suppress IgE production in a human myeloma cell line in vitro57, and to significantly reduce the IgE levels in mice58,59. These reports are in line with the present study further supporting the hypothesis that pectin could be beneficial to reduce allergic reaction and downregulate the inflammatory response in the intestinal mucosa. As the isotype level of allergen-specific IgG1 was diminished while IgG2a was enhanced, the ratio of specific IgG1 to IgG2a was significantly decreased, suggesting a deviation towards a TH1 immune response60.
It remains unclear which structural features of pectin mediate the observed effect. Fermentation experiments showed that LMPs are fermented in vitro more efficiently, whereas HMPs exerts higher modulatory capacity on the gut microbiota composition22,61. Since different pectin subunits are degraded at different rates, structurally different pectins might exert distinct immune modulatory effects22,62. We can therefore only speculate if additional direct effect of pectin on epithelial and immune cells might as well play a role15. It is also known that pectin can also impact on the permeability of the gut and preserve the epithelial integrity19. Rhammnogalacturan-I enriched polysaccharides have been discussed to promote this effect63. However, it remains elusive which fine structure of pectins may have a beneficial effect on allergic inflammation.
In summary, this study showed that dietary pectin intervention shifted the gut microbiota composition towards Bacteroides which was strongly associated with a modulation of the local immune response analyzed by infiltrating cells in the lamina propria. Furthermore, food allergy-related humoral TH2 responses were significantly suppressed in the mice supplemented with pectin. Thus, these results suggest a beneficial role of pectin in the manifestation of FA shifting the immune response from TH2 towards TH1, favoring immune tolerance and providing anti-inflammatory properties.
Materials and methods
Pectin diet
Apple-derived pectin Herbapekt SF 50-LV, a HMP with a degree of esterification (DE) of 57%, galacturonic acid content of 52–61 mol%, and low molecular weight (MW) of 38–47 kDa was provided by Herbstreith & Fox GmbH & Co. KG (Neuenbürg, Germany). Food pellets for dietary intervention were prepared by ssniff Spezialdiäten GmbH (Soest, Germany) and contained 20% cellulose (control diet) or 15% pectin supplemented with 5% cellulose (Table S2).
Animals
Female CBA/J mice (6–8 weeks old) were purchased from Charles River Deutschland GmbH and housed under specific pathogen-free conditions in the animal facility of the Paul-Ehrlich-Institut with free access to water and food. All animal experiments were performed in compliance with the German Animal Welfare Act and the. The authors complied with the ARRIVE guidelines. The study and protocols were reviewed by the animal welfare officer of the Paul-Ehrlich-Institut, and approved by the responsible authority, RP Darmstadt, Germany according the German animal protection law (approval number F107/2005).
Pectin intervention in peach allergic mice
For pectin dietary intervention in peach allergic mice, sensitization and provocation was performed as described recently64 (Fig. 1). Briefly, mice were sensitized intraperitoneally (i.p.) with 200 µg peach peel extract (PE; in 200 µl; n = 5) or PBS (200 µl; n = 3) at d0, d7 and d12 using alum as adjuvant (1 mg per mouse). Afterwards, animals were exposed to 500 µg PE protein or PBS by oral gavage (i.g.) for three times in a two-days interval (d19, d21, d23). Provocation was performed by i.p. injection of 100 µg PE protein (in 200 µl) or PBS at d26, and symptom scores were recorded in a blinded manner (Table S1). Mice of the control groups (PBS or allergic control) were fed control diet starting two weeks before sensitization until the end of the experiment. Composition of the control diet (20% cellulose) and the pectin diet (15% pectin and 5% cellulose) is depicted in Table S2. Mice in the primary-preventive pectin intervention group were fed control diet for one week, followed by pectin diet starting 7 days before sensitization. In the secondary-preventive pectin intervention group, mice were fed control diet until start of the pectin diet after sensitization (d12). Body temperature and symptom score were monitored up to 30 min after each oral exposition and provocation. Feces was collected at day d-7, d-2, d12, d16, and d26. Mice were sacrified by euthanasia using Co2, sera and intestinal tissue were taken and stored at − 80 °C until use.
Analysis of microbiome by metagenomic 16S rRNA gene sequencing
Bacterial DNA was extracted using the QIAamp PowerFecal Pro DNA Kit (Qiagen) and quantified using the QuantiFluor ONE dsDNA System on a Quantus™ Fluorometer (Promega). Amplification of 16S V3 and V4 region was performed by amplicon PCR using amplicon primers as described by Klindsworth et al.65, including Illumina adapter overhang sequence. Subsequently, DNA clean-up was performed and Illumina sequencing adapters were attached using the Nextera XT Index Kit and Index PCR. PCR clean-up was performed and final library was validated using the Agilent D1000 ScreenTape System (Agilent). Samples were adjusted to a final DNA concentration of 4 nM and paired-end sequencing was performed on an Illumina MiSeq benchtop sequencer (Illumina Inc., San Diego, USA) using 2 × 300 base, paired-end setup.
Processing of the sequencing data was performed by Qiime2 package66 using DADA2 (v.1.22.0)67 and Diversity (2022.8) plug-ins. Non-redundant representative sequences were taxonomically classified using the SILVA138 database68.
Intestinal tissue histology and homogenates
Length of intestinal sections were determined, and longitudinal sections of small intestinal tissue (approximately 2 cm) were taken from the jejunum (9.5 cm distal to the duodenum) of CBA/J mice. The tissue sections were fixed in 4% formalin and embedded in paraffin. Sections of 5 μm thickness were prepared using microtome (Microm HM355S, Thermo Scientific) and stained with hematoxylin and eosin (H&E) for morphologic analysis.
Intestine homogenates were prepared as previously reported69. Jejunal tissue (10 cm length) was collected, Peyer’s patches were removed and the tissue was washed with cold PBS, cut in small pieces and subsequently frozen in liquid nitrogen. The frozen tissue was minced using mortar and pistil and the obtained powder was resuspended in 300 µl of cold PBS containing 1 × protease inhibitor (Merck KGaA, Darmstadt, Germany). Samples were centrifuged at 12,000×g for 20 min and supernatant was transferred to fresh tubes. Protein concentration was determined using BCA assay (Thermo Fisher Scientific) and adjusted to 5 mg/ml.
Determination of antibody responses and mMCPT-1
For monitoring of antigen-specific antibody responses, major peach allergen Pru p 3, a non-specific lipid transfer protein (nsLTP), was purified from PE as described previously70. Natural (n) Pru p 3 (5 µg/ml) or PE (50 µg/ml) were coated on microtiter plates overnight at 4°C64. After blocking with 10% FCS in PBS for 2 h at RT, serum was added and incubated for 2 h at RT. Biotinylated anti-mouse IgE (R35-118; BD Biosciences) antibody was incubated for 1 h at RT, followed by 30 min incubation of HRP-labeled streptavidin. For detection of antigen-specific IgG1 (sIgG1) and sIgG2a, HRP-conjugated goat anti-mouse IgG1 (Thermo Fisher Scientific) or HRP-conjugated rabbit anti-mouse IgG2a (Thermo Fisher Scientific) were used. Antigen-specific antibodies were detected by addition of TMB-substrate followed by measurement of the absorbance at 450nm. Detection of total IgE, total IgG and total IgA was performed using commercial ELISA kits according to the manufacturer’s instruction (Thermo Fisher Scientific). In addition, mouse mast cell protease-1 (mMCPT-1) was quantified in serum and intestinal homogenates using commercial ELISA kits according to the manufacturer’s instruction (Thermo Fisher Scientific, Darmstadt, Germany).
Preparation of lamina propria tissue
The lamina propria dissociation was performed following an adapted protocol from Weigmann et al.71. Briefly, small intestines were harvested, fat tissue and Peyer’s patches were removed. The intestines were washed with cold PBS, opened longitudinally and cut in 1 cm pieces. The samples were further washed in 1 × Hank’s Balanced Salt Solution (HBSS) containing 5 mM dithiothreitol (DTT) at 37 °C for 20 min. Intestine pieces were subsequently passed through a 100 µm cell strainer and incubated in pre-digestion solution (1 × HBSS containing 5 mM EDTA and 10 mM HEPES) for 20 min at 37 °C using slow rotation. The samples were again passed through a 100 µm cell strainer followed by repeated incubation in pre-digestion solution. Afterwards, the intestine pieces were washed using 1 × HBSS containing 10 mM HEPES, passed through a cell strainer (100 µm) and incubated in digestion solution (0.5 mg/ml Collagenase D; 0.5 mg/ml DNase I; 1 mg/ml Dispase II in PBS) for 20 min at 37 °C using slow rotation. The samples were subsequently passed through a 40 µm cell strainer and the flow through was collected in cold FCS. The isolated cells were repeatedly washed in cold PBS, counted and used for further analysis by FACS.
FACS analysis of lamina propria cells
Single cell suspensions of lamina propria cells underwent Fc block with anti-CD16/32 (Clone 93; eBioscience, Frankfurt am Main, Germany) followed by staining with extracellular antibodies, viability dye (Thermo Fisher Scientific) and if applicable nuclear staining (true-nuclear, BioLegend). Antibodies and gating strategies are depicted in Table S3 and Figures S6 and S7, respectively. Data were acquired using FACS Symphony (BD Biosciences, Heidelberg, Germany) and analyzed via FlowJo (version 10.0.8r1; BD Biosciences).
Statistical analysis
The results are shown as combined data from two independent mouse experiments conducted under the same experimental settings. The results are represented as means ± SEM, and the data were statistically evaluated by Mann–Whitney U test or ANOVA (α = 0.05) using Graph Pad Prism version 9.5.0 https://www.graphpad.com. For statistical analysis of the sequencing data, the processed 16S V4 rRNA amplicon high throughput sequencing data was analyzed using QIIME 2 software version 2022.8.3 https://qiime2.org/.
Data availability
The datasets generated and analyzed during the current study are available in the European Nucleotide Archive repository (https://www.ebi.ac.uk/ena/browser/home), accession PRJEB81529.
Abbreviations
- FA:
-
Food allergy
- PE:
-
Peach peel extract
- i.p.:
-
Intraperitoneal
- mMCPT-1:
-
Mouse mast cell protease-1
- OIT:
-
Oral immunotherapy
- SIT:
-
Sublingual immunotherapy
- GI:
-
Gastrointestinal
- SCFA:
-
Short chain fatty acids
- HMP:
-
High-methoxyl pectin
- LMP:
-
Low-methoxyl pectin
- DE:
-
Degree of esterification
- i.g.:
-
Intragastric gavage
- HBSS:
-
Hank’s Balanced Salt Solution
- DTT:
-
Dithiothreitol
- cDCs:
-
Conventional dendritic cells
References
Feng, H. et al. Prevalence and factors for food allergy in different populations from different regions: A protocol for a systematic review and meta-analysis. PLoS ONE 16, e0261092 (2021).
Sicherer, S. H. & Sampson, H. A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 141, 41–58 (2018).
Barni, S. et al. Phenotypes and endotypes of peach allergy: What is new? Nutrients 14 (2022).
Fleischer, D. M. et al. Sublingual immunotherapy for peanut allergy: A randomized, double-blind, placebo-controlled multicenter trial. J. Allergy Clin. Immunol. 131, 119–27.e1–7 (2013).
Burks, A. W. et al. Oral immunotherapy for treatment of egg allergy in children. N. Engl. J. Med. 367, 233–243 (2012).
Vickery, B. P. et al. AR101 oral immunotherapy for peanut allergy. N. Engl. J. Med. 379, 1991–2001 (2018).
Bird, J. A. et al. Efficacy and safety of AR101 in oral immunotherapy for peanut allergy: Results of ARC001, a randomized, double-blind, placebo-controlled phase 2 clinical trial. J. Allergy Clin. Immunol. Pract. 6, 476–485.e3 (2018).
Muraro, A. et al. Managing food allergy: GA2LEN guideline 2022. World Allergy Organ. J. 15, 100687 (2022).
Rachid, R., Stephen-Victor, E. & Chatila, T. A. The microbial origins of food allergy. J. Allergy Clin. Immunol. 147, 808–813 (2021).
Bunyavanich, S. & Berin, M. C. Food allergy and the microbiome: Current understandings and future directions. J. Allergy Clin. Immunol. 144, 1468–1477 (2019).
Nance, C. L. et al. The role of the microbiome in food allergy: A review. Children (Basel, Switzerland) 7 (2020).
Kreft, L., Hoffmann, C. & Ohnmacht, C. Therapeutic potential of the intestinal microbiota for immunomodulation of food allergies. Front. Immunol. 11, 1853 (2020).
Zubeldia-Varela, E. et al. Microbiome and allergy: New insights and perspectives. J. Investig. Allergol. Clin. Immunol. 32, 327–344 (2022).
Tramper-Stranders, G. et al. Dangerous liaisons: Bacteria, antimicrobial therapies, and allergic diseases. Allergy 76, 3276–3291 (2021).
Blanco-Pérez, F. et al. The dietary fiber pectin: Health benefits and potential for the treatment of allergies by modulation of gut microbiota. Curr. Allergy Asthma Rep. 21, 43 (2021).
Jang, Y. O. et al. High-fiber diets attenuate emphysema development via modulation of gut microbiota and metabolism. Sci. Rep. 11, 7008 (2021).
Gill, S. K., Rossi, M., Bajka, B. & Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 18, 101–116 (2021).
Dang, G. et al. Pectin supplement alleviates gut injury potentially through improving gut microbiota community in piglets. Front. Microbiol. 13, 1069694 (2022).
Beukema, M., Faas, M. M. & de Vos, P. The effects of different dietary fiber pectin structures on the gastrointestinal immune barrier: Impact via gut microbiota and direct effects on immune cells. Exp. Mol. Med. 52, 1364–1376 (2020).
Wang, S. et al. Apple Polysaccharide inhibits microbial dysbiosis and chronic inflammation and modulates gut permeability in HFD-fed rats. Int. J. Biol. Macromol. 99, 282–292 (2017).
Pascale, N., Gu, F., Larsen, N., Jespersen, L. & Respondek, F. The potential of pectins to modulate the human gut microbiota evaluated by in vitro fermentation: a systematic review. Nutrients 14 (2022).
Bender, C. et al. Time-dependent fermentation of different structural units of commercial pectins with intestinal bacteria. Carbohydr. Polym. 308, 120642 (2023).
Verstegen, R. E. M. et al. Dietary fibers: Effects, underlying mechanisms and possible role in allergic asthma management. Nutrients 13 (2021).
Trompette, A. et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 15, 908–926 (2022).
EFSA Panel on Dietetic Products, Nutrition and Allergies. Scientific Opinion on the substantiation of health claims related to pectins and reduction of post-prandial glycaemic responses (ID 786), maintenance of normal blood cholesterol concentrations (ID 818) and increase in satiety leading to a reduction in ene. EFSA J 8, 1747 (2010).
Trompette, A. et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 20, 159–166 (2014).
Zhang, Z. et al. Dietary fiber intake regulates intestinal microflora and inhibits ovalbumin-induced allergic airway inflammation in a mouse model. PLoS ONE 11, e0147778 (2016).
Zwolschen, J. W., Vos, A. P., Ariëns, R. M. C. & Schols, H. A. In vitro batch fermentation of (un)saturated homogalacturonan oligosaccharides. Carbohydr. Polym. 329, 121789 (2024).
Tang, X. & de Vos, P. Structure-function effects of different pectin chemistries and its impact on the gastrointestinal immune barrier system. Crit. Rev. Food Sci. Nutr. 1–15 (2023).
Steigerwald, H. et al. Effects of pectin methyl-esterification on intestinal microbiota and its immunomodulatory properties in naive mice. Carbohydr. Polym. 334, 122007 (2024).
Miller, H. R. P. & Pemberton, A. D. Tissue-specific expression of mast cell granule serine proteinases and their role in inflammation in the lung and gut. Immunology 105, 375–390 (2002).
Larsen, N. et al. Potential of pectins to beneficially modulate the gut microbiota depends on their structural properties. Front. Microbiol. 10, 223 (2019).
Yip, W. et al. Butyrate shapes immune cell fate and function in allergic asthma. Front. Immunol. 12, 628453 (2021).
Cruz-Amaya, A. et al. Butyrate dietary supplementation promotes tolerant responses in a Pru p 3-anaphylactic mouse model. Allergy https://doi.org/10.1111/all.16033 (2024).
Di Costanzo, M., Paulis, N. de & Biasucci, G. Butyrate: A link between early life nutrition and gut microbiome in the development of food allergy. Life (Basel, Switzerland) 11 (2021).
Luu, M., Monning, H. & Visekruna, A. Exploring the molecular mechanisms underlying the protective effects of microbial SCFAs on intestinal tolerance and food allergy. Front. Immunol. 11, 1225 (2020).
Nagata, K. et al. Butyrate, valerate, and niacin ameliorate anaphylaxis by suppressing IgE-dependent mast cell activation: Roles of GPR109A, PGE2, and epigenetic regulation. J. Immunol. (Baltimore, Md. : 1950) 212, 771–784 (2024).
Luis, A. S. et al. Dietary pectic glycans are degraded by coordinated enzyme pathways in human colonic Bacteroides. Nat. Microbiol. 3, 210–219 (2018).
Tian, L. et al. Effects of pectin supplementation on the fermentation patterns of different structural carbohydrates in rats. Mol. Nutr. Food Res. 60, 2256–2266 (2016).
Parada Venegas, D. et al. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10, 277 (2019).
Qu, D. et al. Protective effects of Bacteroides fragilis against lipopolysaccharide-induced systemic inflammation and their potential functional genes. Food Funct. 13, 1015–1025 (2022).
Shon, H.-J. et al. Protective role of colitis in inflammatory arthritis via propionate-producing Bacteroides in the gut. Front. Immunol. 14, 1064900 (2023).
Pang, W. et al. Bacteroides thetaiotaomicron Ameliorates Experimental Allergic Airway Inflammation via Activation of ICOS+Tregs and Inhibition of Th2 Response. Front. Immunol. 12, 620943 (2021).
Goldberg, M. R. et al. Microbial signature in IgE-mediated food allergies. Genome Med. 12, 92 (2020).
Nie, Q. et al. Bioactive dietary fibers selectively promote gut microbiota to exert antidiabetic effects. J. Agric. Food Chem. 69, 7000–7015 (2021).
Jian, H., Liu, Y., Wang, X., Dong, X. & Zou, X. Akkermansia muciniphila as a next-generation probiotic in modulating human metabolic homeostasis and disease progression: A role mediated by gut-liver-brain axes? Int. J. Mol. Sci. 24 (2023).
Parrish, A. et al. Akkermansia muciniphila exacerbates food allergy in fibre-deprived mice. Nat. Microbiol. 8, 1863–1879 (2023).
Stoll, M. L. et al. Akkermansia muciniphila is permissive to arthritis in the K/BxN mouse model of arthritis. Genes Immun. 20, 158–166 (2019).
Liu, C. et al. Influence of glucose fermentation on CO₂ assimilation to acetate in homoacetogen Blautia coccoides GA-1. J. Ind. Microbiol. Biotechnol. 42, 1217–1224 (2015).
Liu, X. et al. Blautia-a new functional genus with potential probiotic properties?. Gut Microbes 13, 1–21 (2021).
Nakano, N. & Kitaura, J. Mucosal Mast Cells as Key Effector Cells in Food Allergies. Cells 11 (2022).
Galli, S. J. & Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 18, 693–704 (2012).
Blanco-Pérez, F. et al. Mast cells partly contribute to allergic enteritis development: Findings in two different mast cell-deficient mice. Allergy 77, 1051–1054 (2022).
Shamji, M. H. et al. The role of allergen-specific IgE, IgG and IgA in allergic disease. Allergy 76, 3627–3641 (2021).
El Ansari, Y. S. et al. Allergen-specific IgA antibodies block IgE-mediated activation of mast cells and basophils. Front. Immunol. 13, 881655 (2022).
Scheurer, S., Junker, A.-C., He, C., Schülke, S. & Toda, M. The role of IgA in the manifestation and prevention of allergic immune responses. Curr. Allergy Asthma Rep. 23, 589–600 (2023).
Iwamoto, A., Inoue, Y., Tachibana, H. & Kawahara, H. Alkali-soluble pectin suppresses IgE production in human myeloma cell line in vitro. Cytotechnology 71, 573–581 (2019).
Lim, B. O., Lee, S. H., Park, D. K. & Choue, R. W. Effect of dietary pectin on the production of immunoglobulins and cytokines by mesenteric lymph node lymphocytes in mouse colitis induced with dextran sulfate sodium. Biosci. Biotechnol. Biochem. 67, 1706–1712 (2003).
Ye, M. B. & Lim, B. O. Dietary pectin regulates the levels of inflammatory cytokines and immunoglobulins in interleukin-10 knockout mice. J. Agric. Food Chem. 58, 11281–11286 (2010).
Romagnani, S. Immunologic influences on allergy and the TH1/TH2 balance. J. Allergy Clin. Immunol. 113, 395–400 (2004).
Dongowski, G., Lorenz, A. & Proll, J. The degree of methylation influences the degradation of pectin in the intestinal tract of rats and in vitro. J. Nutr. 132, 1935–1944 (2002).
Popov, S. V. & Ovodov, Y. S. Polypotency of the immunomodulatory effect of pectins. Biochem. Biokhimiia 78, 823–835 (2013).
van den Abbeele, P. et al. A novel non-digestible, carrot-derived polysaccharide (cRG-I) selectively modulates the human gut microbiota while promoting gut barrier integrity: An integrated in vitro approach. Nutrients 12 (2020).
Steigerwald, H. et al. Peach extract induces systemic and local immune responses in an experimental food allergy model. Sci. Rep. 13, 1892 (2023).
Klindworth, A. et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41, e1 (2013).
Bolyen, E. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857 (2019).
Callahan, B. J. et al. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583 (2016).
Quast, C. et al. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596 (2013).
Blanco-Pérez, F. et al. CCR8 leads to eosinophil migration and regulates neutrophil migration in murine allergic enteritis. Sci. Rep. 9, 9608 (2019).
Toda, M. et al. Protein unfolding strongly modulates the allergenicity and immunogenicity of Pru p 3, the major peach allergen. J. Allergy Clin. Immunol. 128, 1022–30.e1–7 (2011).
Weigmann, B. et al. Isolation and subsequent analysis of murine lamina propria mononuclear cells from colonic tissue. Nat. Protoc. 2, 2307–2311 (2007).
Acknowledgements
The authors thank Prof. Hans Ulrich Endreß for providing the pectin, Tanja Leyendecker and Csaba Miskey, ATMP/Haematology, Genomics Core Facility, Paul-Ehrlich-Institut, for technical support setting up NGS. The present study was partially funded by the German Federal Ministry for Education and Research (BMBF), BMBF-ERA 01EA1901 and AC18/00031 (DIFAMEM), an IGF Project of the FEI which is supported via AiF (AiF20528) within the program for promoting the Industrial Collective Research (IGF) of the German Ministry of Economic Affairs and Energy (BMWi), and the DAAD-JSPS PPP 57710985.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
M.T., S.Sch. and S.V. conceived and designed the study. H.S., F.B.-P., M.A., M.K., A.W. and M.S. performed the experiments and data analysis; I.G.-M. and L.Q.-M. generated histological data; B.B. supported by microbiota analysis, and O.K. performed bioinformatic analysis. H.S. and F.B.-P. drafted the article, S.Sch. was involved in the data analysis and finalized the manuscript. All authors have read and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
Stefan Vieths reports personal fees from Schattauer (Allergologie Handbuch), Elsevier (Nahrungsmittelallergien und Intoleranzen), Karger (Food Allergy: Molecular Basis and Clinical Practice), and Journal of Allergy and Clinical Immunology (Associate Editor), outside the submitted work. The other authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Steigerwald, H., Albrecht, M., Blissenbach, B. et al. Dietary fiber pectin alters the gut microbiota and diminishes the inflammatory immune responses in an experimental peach allergy mouse model. Sci Rep 14, 30503 (2024). https://doi.org/10.1038/s41598-024-82210-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-82210-3
