
Abstract
This study investigated the influence of various nanoparticles (NPs), including carbon-based NPs, Al2O3-NPs, SiO2-NPs, and TiO2-NPs, on the growth and physiology of okra plants cultivated in soil impacted by rocky desertification. The morphological characteristics, photosynthetic parameters, antioxidant capacity, nutrient uptake of the okra plants, and available nutrient content in the soil were evaluated. The results demonstrated a significant improvement in the growth, photosynthetic properties, chlorophyll content, and activities of antioxidant enzymes (catalase, peroxidase, and superoxide dismutase) in the okra plants treated with NPs compared to the control. Additionally, the NP-treated plants exhibited enhanced total nitrogen content, and hydrolysis nitrogen content in soil was also significantly increased, indicating improved nitrogen uptake. Notably, TiO2-NPs demonstrated the most pronounced effects on plant growth and photosynthetic characteristics, while TiO2-NPs and SiO2-NPs showed higher antioxidant enhancement compared to other NPs. These findings suggest the potential of nanoparticles as a promising agricultural solution to address challenges in regions like Guizhou Province, China.
Similar content being viewed by others


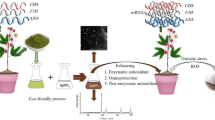
Introduction
In southwestern China, karst rocky desertification is a significant ecological issue that hinders the economic and social development of the region. This process involves severe soil erosion, extensive exposure of basement rocks, a drastic decrease in soil productivity, and the formation of a desert-like landscape. The control of rocky desertification has been identified as a priority in national projects for economic and social development1. Numerous studies have endeavored to elucidate the causes and environmental ramifications of soil rocky desertification, providing comprehensive insights into this phenomenon.Wang et al.2., found that different combinations of basement carbonate rocks contribute to the development of rocky desertification. In the Shibanqiao catchment of Guanling County, Guizhou Province, they conducted a systematic study on the impact of land use and land cover changes on soil erosion over the past 40 years. The research utilized environmental information extracted from lake sediment to analyze the relationship between soil erosion and rocky desertification. Xiuqin et al.3, reported that accelerated soil erosion occurred during the implementation of ecological engineering between 1990 and 2002. They suggested that future ecological constructions and engineering projects should prioritize soil erosion prevention and avoid unnecessary soil disturbance. Wang et al.4, proposed an erosion-creep-collapse mechanism to explain underground soil loss in the karst rocky desertification of Chenqi village, Puding county, Guizhou province. By controlling soil collapse, soil loss can be effectively prevented. Afforestation of degraded land is a crucial strategy for preventing soil erosion and promoting ecosystem recovery in rocky desertification areas. Cheng et al.5, studied biomass increment and carbon accumulation in different aged Z. bungeanum stands, as well as the distribution of carbon stock among various biomass components and soil depths. Furthermore, Cheng et al.6, analyzed variations in biomass and nitrogen storage, as well as nitrogen distribution in biomass components and soil depths. These studies highlight the importance of understanding and managing biomass and nutrient dynamics in combating rocky desertification. Additionally, Zhao & Hou7, discussed five human-related factors that contribute to soil loss in rural karst mountainous areas of Guizhou Province. These factors further exacerbate the problem of rocky desertification. The control of karst rocky desertification is vital for the sustainable development of southwestern China, particularly Guizhou Province, a less developed region. Through thorough research and understanding of the causes and mechanisms of soil erosion and desertification, effective strategies can be implemented to prevent and mitigate this ecological problem.
Owing to their unique properties, nanoparticles (NPs) have garnered significant attention in recent research for applications in agriculture and ecological control. These materials exhibit exceptional characteristics that make them promising candidates for addressing various challenges in these fields. Biochar nanoparticles have been studied as a soil amendment for a sandy loam soil, which may play a role in addressing soil degradation and erosion8. Desert Control, a company specializing in desertification control, has proposed the use of nano-clay to turn unproductive desert land into fertile soil9. This innovative approach suggested that nanomaterials, such as nanoclay, may offer a promising solution for soil improvement in arid and semiarid regions. Cecchin et al.10, provided a comprehensive review of the application of nanomaterials in soil remediation, discussing their toxicity to soil microorganisms and the potential for integration with other remediation technologies, including bioremediation. They proposed the concept of nanobioremediation (NBR), which aims to combine these two technologies for more effective, efficient, and sustainable remediation outcomes. Hussain et al.11, present a comprehensive review of bioremediation techniques for environmental pollutants, highlighting the integration of nanoparticles. Their analysis illuminated the potential of nanotechnology-based bioremediation as a sustainable and cost-effective solution for mitigating environmental contamination. NPs possess potential for enhancing soil quality and facilitating plant growth in regions impacted by desertification12. However, the effects of different NPs on plant growth and physiology remain complex and variable. As reported by Kong et al.13, the impact of NPs on root and shoot development varies significantly depending on the type of nanoparticles, incubation conditions, and plant species. Our previous studies14 further support this variability, demonstrating that Al2O3-NPs (ANPs), SiO2-NPs (SNPs) exert positive effects on growth and photosynthetic properties in okra plants under water deficit conditions. However, TiO2-NPs (TNPs) were found to have detrimental effects, significantly impairing both growth and photosynthetic performance in okra plants.
Okra, scientifically known as Abelmoschus esculentus, is a vital economic crop recognized for its nutritional benefits and economic significance. It is abundant in essential vitamins and minerals, particularly dietary fiber, vitamin C, and folate, thereby making it an excellent component of a healthy diet. The Food and Agriculture Organization (FAO) reported that global okra production reached approximately 5.2 million tons in 2020, with India and Nigeria being the primary producers15. Beyond its nutritional advantages, okra significantly impacts the livelihoods of farmers, particularly in arid regions where it enhances income and food security amid challenging environmental conditions. Du to its adaptability to harsh, rocky terrains, this warm-season crop demonstrates remarkable drought resistance, thriving with minimal water, an essential characteristic for areas with scarce moisture. Additionally, its rapid growth cycle and resilience to high temperatures enable efficient biomass production, yielding substantial economic and nutritional value. Thus, this study investigated the influence of NPs on plant growth in soils degraded by rocky desertification, offering new strategies to enhance crop production in the Karst rocky desertification region of Guizhou Province, China.
Materials and methods
Materials
Soil samples were collected from a low rocky desertification area in Zhanjie Town, Qingzhen County, Guizhou Province, China (106°39′58.05″E, 26°56′21.13″N). The chemical properties of the soil surrounding the rocky desertification area are presented in Table 1. Okra seeds, used as the cultivated plant, were obtained from Guiyang University. The drought-tolerant okra cultivar “Xianzhi” was identified based on its agronomic traits and physiological and biochemical responses to water deficiency. This cultivar demonstrates significant resilience under conditions of limited water availability, making it a promising option for cultivation in arid environments.This study employed four types of NPs, namely carbon-based NPs (CNPs), Al2O3-NPs (ANPs), SiO2-NPs (SNPs) and TiO2-NPs (TNPs), which were purchased from Beijing Dk nano S&T Ltd, China. Notably, all NPs exhibited a nearly spherical morphology with an average particle size ranging from 20 to 40 nm.
Methods
Experimental design
A pot experiment was conducted in a greenhouse at Guiyang University under controlled temperature (25 ± 2 °C) and humidity (70 ± 5%), from April to July in 2024. Okra was planted in plastic pots (28.0 cm depth × 26.0 cm diameter) filled with 2 kg of soil with moderate rocky desertification. Prior to sowing, different types of NPs were added to the soil at the following concentrations: CNPs at 60 mg/kg, ANPs at 60 mg/kg, SNPs at 15 mg/kg, and TNPs at 60 mg/kg on the basis of our previous study16. Additionally, 15 g of compound fertilizer (N: P:K = 12:18:1) was applied per treatment before sowing. The direct seeding method was used, with 5–8 seeds planted per pot. Okra was also planted without NPs as a control. The experiment consisted of five treatments, each replicated five times, resulting in a total of 25 pots arranged in a completely randomized design.
Determination
Plant growth characteristics were measured after 48 days. Photosynthetic parameters, including photosynthetic rate (A), transpiration rate (E), intercellular CO2 concentration (Ci), stomatal conductance (Gs), vapor pressure deficit (VPD), leaf temperature (T), relative humidity of leaf chamber (Rh) were determined using a portable photosynthesis system (GFS-3000, EU). Chlorophyll content was measured with a SPAD-502plus analyzer. Catalase (CAT), superoxide dismutase (SOD), peroxidase (POD) activity, and malondialdehyde (MDA) content were determined using kits from Suzhou Grith Biotechnology Co., LTD (China). According to the method of Chun et al..,, total nitrogen (N), total phosphorus (P), and total potassium (K) in plants were analyzed using a continuous flow analyzer (Auto Analyzer 3, Germany) and flame spectrometry (Sherwood M425, UK), respectively. Soil available nutrient content (N, P, K) was measured according to Wang et al.17, the alkali diffusion method for nitrogen, colorimetry for phosphorus, and atomic absorption spectroscopy (AAS) for potassium.
Data analysis
All data were presented as mean ± standard deviation of triplicate samples and analyzed using one-way analysis of variance (ANOVA) followed by Duncan’s test in Origin 8.0. The significance of treatment effects was considered at a P value < 0.05. The subordinate function value method18, was employed to comprehensively evaluate the drought resistance of okra plants.
Results
The effect of various NPs on growth of okra plant in soil affected by rocky desertification
The morphological characteristics of okra grown in soil affected by rocky desertification were determined, as shown in Table 2. The okra growth, as indicated by various parameters including blade number (BN), number of green leaves (GL), maximum blade width per plant (MBW), maximum blade length per plant (MBL), length of main root (LMR), plant height (PH), stem diameter (SD), fresh weight (FW), dry weight (DW), root-shoot ratio (RS), was dramatically significantly enhanced by the application of NPs (CNPs, SNPs, TNPs, and ANPs), compared to the control group. However, no significant differences were observed among the various NP treatments.
The effect of various NPs on photosynthetic property of okra plant in soil affected by rocky desertification
As illustrated in Table 3, with the exception of Gs, all other photosynthetic parameters, including net photosynthetic rate (A), intercellular CO2 concentration (Ci), transpiration rate (E), leaf temperature (T), relative humidity (Rh), and vapor pressure deficit (VPD), exhibited significant increases in NPs-treated okra plants compared to the control, with the notable exceptions of A and T in okra plants treated with ANPs. Notably, the okra plants exposed to SNPs exhibited the highest values for A, Ci, E, Gs, and VPD, surpassing the other NP treatments. Furthermore, as presented in Fig. 1, the leaf chlorophyll content, including total chlorophyll (Chl t), chlorophyll a (Chl a), and chlorophyll b (Chl b), were significantly enhanced in the NPs-treated okra plants compared to the control, while the Chl a/b ratio was decreased.

The leaf chlorophyll contents of okra plant treated with various NPs. A: total chlorophyll (Chl t), chlorophyll a (Chl a), and chlorophyll b (Chl b), B: Chl a/b ratio.
The effect of various NPs on antioxidative activity and nutrient uptake of okra plant in soil affected by rocky desertification
The antioxidative activity of okra plants is presented in Fig. 2. The activities of CAT, POD, and SOD in okra plants exposed to NPs were significantly improved compared to the control group. Specifically, the POD activity of okra plants treated with CNPs was significantly higher than those treated with other NPs. No significant differences in MDA content were identified among all treatments and the control. According to Fig. 3, total N of okra plant were both observably increased by NPs compared to the control, while total P and K of that were observably decreased. Similarly, no significant differences were observed among the various NPs.
The effect of various NPs on available nutrition content of soil affected by rocky desertification region
As shown in Fig. 4, the analysis of soil nutrient availability revealed that the levels of hydrolysis N were generally higher in the soils exposed to NPs compared to the control group. However, the available K content in the NP-treated soils was significantly lower than the control, except for ANPs treatment. No substantial differences were found in the available P content across the various NP treatments and the control.

The antioxidant ability of okra plant treated with various NPs.A:Catalase (CAT), B:Superoxide dismutase (SOD), C:Peroxidase (POD), D: Malondialdehyde (MDA).

The nutrient content of okra plant treated with various NPs. A:Total nitrogen (N), B:Total phosphorus (P), C: Total potassium (K).

The available nutrient content of soil A: available N, B: available P, C: available K.
Subordinate function analysis
The application of NPs have been found to enhance the resistance abilities of okra plants grown in soil affected by rocky desertification. Analysis of the morphological properties (Table 4) and photosynthetic physiological indices (Table 5) indicated that the TNPs had the most significant promoting effect compared to other NPs. Furthermore, the biochemical indices (Table 6), such as antioxidant activity and N/P/K nutrient content, also showed that both TNPs and SNPs had the best promoting effect on the resistance abilities of okra plants in this challenging soil environment.
Discussion
This studies have indicated that the application of NPs, including CNPs, SNPs, TNPs, and ATPs can significantly improve the growth of okra plant in soil affected by rocky desertification. Similar studies have been reported in Dracocephalum moldavica19 and broad bean20. NPs improve soil physicochemical properties, increasing porosity, water-holding capacity, and nutrient availability, which facilitates root development and nutrient uptake, as reflected in elevated hydrolysis N (soil) and total N (plant) levels. However, the specific mechanisms vary among NP types and warrant further investigation to clarify interactions between NPs, soil, and plant responses under these conditions21.
In this work, NPs has been shown to enhance the photosynthetic capacity and chlorophyll content of okra plants under conditions of rocky desertification. Previous studies, such as that by Venkatachalam et al.22, on cotton plants (Gossypium hirsutum L.), have reported similar findings. The improved photosynthetic efficiency associated with NPs application can be attributed to the enhanced activity of the Rubisco enzyme, which plays a crucial role in carbon dioxide absorption and photosynthesis23. The unique properties of NPs, including their high surface area-to-volume ratio and increased reactivity, may contribute to improved nutrient availability and uptake by the plants, ultimately leading to enhanced photosynthetic performance and chlorophyll synthesis24. Therefore, the observed increase in total N contents in okra plants may have a positive impact on photosynthesis. Photosynthesis is influenced by the specific leaf nitrogen content, which is partly related to the partitioning of nitrogen in photosynthetic enzymes, pigment content, and the size, number, and composition of chloroplasts25. Previous studies have reported a linear increase in photosynthesis with the increase in leaf N content in sugarcane26. Additionally, N stimulates leaf growth through the synthesis of proteins involved in cell growth, cell division, and cell wall and cytoskeleton synthesis, thereby increasing the photosynthetic area27. Furthermore, the studies of Ye et al.28, have revealed that N plays a substantial role in plant responses and tolerance to abiotic stresses, including its involvement in physiological and biochemical mechanisms related to photosynthesis, osmoprotection, stomatal regulation, water-nutrient absorption, nutrient translocation, and enzyme activation. Under abiotic stress conditions, the observed decrease in the Chl a/b ratio induced by nanomaterial treatments can be attributed to stress-induced alterations in chlorophyll metabolism. Specifically, chlorophyll a is more susceptible to degradation under stress due to increased oxidative stress and the activity of chlorophyll-degrading enzymes, such as chlorophyllase. This selective degradation leads to a reduction in the Chl a/b ratio. Furthermore, under stress, plants may downregulate the synthesis of chlorophyll a to reduce the risk of photodamage to the photosynthetic apparatus, while maintaining or even increasing chlorophyll b levels to adjust light-harvesting efficiency. This adjustment can help plants optimize photosynthesis under suboptimal conditions.
The application of NPs has shown promising results in improving total N content of okra plants and hydrolysis N content in soil. However, the observed decrease in total P and K content suggested that the effects of NPs on plant nutrient absorption are not uniform across different nutrients. The underlying mechanism behind this phenomenon can be attributed to the unique properties and interactions of NPs with the soil and plant systems. NPs, due to their high surface-to-volume ratio and enhanced reactivity, can potentially alter the availability, mobility, and uptake efficiency of various nutrients in the soil29. In the case of nitrogen, NPs may have facilitated their increased solubility, bioavailability, and subsequent absorption by the okra plants, leading to the observed higher concentrations. On the other hand, the decrease in phosphorus and potassium content could be related to the complex interactions between NPs, soil properties, and phosphorus dynamics30. NPs may have influenced the adsorption, precipitation, or immobilization of phosphorus and and potassium in the soil, thereby reducing its accessibility and uptake by the plants. This was also supported by a significant reduction in soil available K content. Additionally, NPs may have altered the soil’s pH, redox conditions, or the activities of soil microorganisms, which can indirectly affect the availability and utilization of phosphorus by the plants (Sembada & Lenggoro31.
This study demonstrated that applying NPs to soil degraded by rocky desertification enhanced antioxidant enzyme activity of okra plants, thereby protecting plant from stress-induced damage. This protective effect is likely attributed to the detoxification of reactive oxygen species (ROS), which accumulate in karst rocky desertification areas32. The increased activity of antioxidant enzymes, such as SOD, CAT, and APX, suggested that NPs application promoted efficient ROS scavenging. These findings align with previous research indicating that NPs can improve cell membrane integrity, leading to enhanced antioxidant enzyme activity and stress tolerance33,34. The enhanced antioxidant enzyme activities display a critical role in improving plant resistance to the adverse stresses associated with rocky desertification35. The significantly enhanced activity of okra POD treated with CNPs compared to other NPs can be attributed to the unique physicochemical properties of CNPs. CNPs exhibit high surface areas, biocompatibility, and the ability to modulate enzyme conformation through non-covalent interactions. These features may stabilize the enzyme’s active site and enhance its catalytic efficiency. Furthermore, CNPs’ ability to act as electron transfer mediators could facilitate the redox reactions catalyzed by POD, potentially increasing its activity. In contrast, other NPs, such as metal-oxide nanoparticles(ANPs), may induce structural perturbations or aggregation, leading to reduced enzymatic activity. Studies have demonstrated that the interaction between enzymes and CNPs can lead to improved stability and catalytic performance under various conditions36. However, NPs did not alter the MDA activity in plants, suggesting that environmental stressors such as drought and nutrient deficiency in rocky desertification soil imposed greater stress on okra. Consequently, the promotional effects of NPs were limited, which may also be dose-dependent.
The existing literature suggested that NPs primarily enhance plant growth, physiological parameters, productivity, and nutritional value by modulating various intrinsic signaling pathways. These pathways include chlorophyll biosynthesis, carbohydrate metabolism, antioxidant machinery, and hormone synthesis, ultimately leading to improved plant growth and yield37. NPs exhibit antioxidant properties, scavenging ROS and reducing oxidative stress induced by abiotic stress. They also enhance the activity of antioxidant enzymes, contributing to a robust antioxidant defense system. Furthermore, NPs aid in osmotic adjustment, maintain cell membrane integrity, and modulate plant hormone signaling pathways, thereby promoting stress tolerance in plants38. While the specific mechanisms may vary depending on nanoparticle characteristics, plant species, stress type, and dosage, further research is necessary to fully elucidate the precise molecular pathways involved in the stress mitigation properties of NPs. Nonetheless, the existing evidence indicated that NPs have the potential to enhance plant growth, productivity, and resilience in the face of abiotic stress39.
It is worth noting that different nanoparticles recorded lack of significant differences in improving plant growth. One plausible explanation is that the fundamental mechanisms by which nanoparticles influence plant growth are not entirely dependent on their specific material composition. Instead, they may share common pathways, such as the ability to penetrate plant cells, alter membrane permeability, or modulate ion transport40. Additionally, the size and surface charge of nanoparticles, which are critical determinants of their bioavailability and interaction with plant cells, may play a moresignificant role than their material composition in driving growth responses. Another factor could be the experimental conditions, such as the concentration of nanoparticles applied, the duration of exposure, and the plant species used, which may obscure material-specific effects. Furthermore, it is possible that the responses of plants to nanoparticles are governed by complex cellular and molecular processes, including hormonal regulation and stress responses, which may override the subtle differences in nanoparticle properties41. While this phenomenon underscores the potential universality of nanoparticle-induced plant growth promotion, it also raises questions about the long-term implications and the mechanisms that drive these effects42. Nanoparticles possess the ability to enhance nutrient transport, improve soil quality, and increase crop yields. However, their persistence and accumulation in the environment pose ecological risks. Notably, the risk assessment of their impact on soil microbial populations emerges as a critical priority for future research.
Conclusion
The findings of our study indicate that the use of nanoparticles can greatly improve the growth of okra in soil affected by rocky desertification. Nanoparticles can enhance photosynthetic efficiency, nitrogen absorption, and antioxidant enzyme activity in plants, thereby resulting in improved growth outcomes and enhanced stress tolerance. This research highlights the potential of nanomaterials as an effective solution for agricultural production in rocky desertification areas of Guizhou Province, China.
Data availability
All data analysed during this study are included in this published article and supplementary information file.
References
Liu, Y. Household livelihood choices under the different eco-environment in the karst area: A case study of Anshun City, Southwest of China. Environ. Res. 197, 111171. https://doi.org/10.1016/j.envres.2021.111171 (2021).
Wang, S. et al. How types of carbonate rock assemblages constrain the distribution of karst Rocky desertified land in Guizhou Province, PR China: phenomena and mechanisms. Land. Degrad. Dev. https://doi.org/10.1002/ldr.591 (2004).
Xiuqin, W., Yun-long, C. & Tao, Z. Effects of land use/land cover changes on Rocky Desertification-A case study of a small karst catchment in Southwestern China. Energy Procedia. 5, 1–5. https://doi.org/10.1016/J.EGYPRO.2011.03.001 (2011).
Wang, J. et al. Erosion-creep-collapse mechanism of underground soil loss for the karst Rocky desertification in Chenqi village, puding County, Guizhou, China. Environ. Earth Sci. 72, 2751–2764. https://doi.org/10.1007/s12665-014-3182-0 (2014).
Cheng, J. et al. Biomass accumulation and carbon sequestration in an age-sequence of Zanthoxylum bungeanum plantations under the grain for green program in karst regions, Guizhou Province. Agric. For. Meteorol. 203, 88–95. https://doi.org/10.1016/J.AGRFORMET.2015.01.004 (2015).
Cheng, J. et al. Changes in above- and below-ground nitrogen stocks and allocations following the conversion of farmland to forest in Rocky desertification regions. Agric. Ecosyst. Environ. 232, 9–16. https://doi.org/10.1016/J.AGEE.2016.07.019 (2016).
Zhao, L. & Hou, R. Human causes of soil loss in rural karst environments: a case study of Guizhou. China Sci. Rep. 9 (1), 3225. https://doi.org/10.1038/s41598-018-35808-3 (2019).
Chen, X., Duan, M., Zhou, B. & Cui, L. Effects of Biochar nanoparticles as a soil amendment on the structure and hydraulic characteristics of a sandy loam soil. Soil Use Manag. 38, 836–849. https://doi.org/10.1111/sum.12740 (2021).
Zhang, Z. & Huisingh, D. Combating desertification in China: monitoring, control, management and revegetation. J. Clean. Prod. 182, 765–775. https://doi.org/10.1016/j.jclepro.2018.01.233 (2018).
Cecchin, I., Reddy, K. R., Thomé, A., Tessaro, E. & Schnaid, F. Nanobioremediation: integration of nanoparticles and bioremediation for sustainable remediation of chlorinated organic contaminants in soils. Int. Biodeterior. Biodegrad. 119, 419–428. https://doi.org/10.1016/J.IBIOD.2016.09.027 (2017).
Hussain, A. et al. In-situ, Ex-situ, and nano-remediation strategies to treat polluted soil, water, and air - A review. Chemosphere 289, 133252. https://doi.org/10.1016/j.chemosphere.2021.133252 (2022).
Fatima, F., Hashim, A. & Anees, S. Efficacy of nanoparticles as nanofertilizer production: a review. Environ. Sci. Pollut Res. 28, 1292–1303. https://doi.org/10.1007/s11356-020-11218-9 (2021).
Kong, I. C., Ko, K. S. & Koh, D. C. Comparisons of the effect of different metal oxide nanoparticles on the root and shoot growth under shaking and Non-Shaking incubation, different plants, and binary mixture conditions. Nanomaterials (Basel Switzerland). 11 (7), 1653. https://doi.org/10.3390/nano11071653 (2021).
Wang, J. et al. Effects of nano oxide particles on the growth and photosynthetic characteristics of Okra plant under water deficiency. Folia Horticulturae. 36 (3), 1–13. https://doi.org/10.2478/fhort-2024-0029 (2024).
FAO. Introduction To Seed Systems (FAO Plant Production and Protection Division, 2020).
Wang, J., Xu, J., Yang, M. & Miao, M. Influence of SiO2, Al2O3 and TiO2 nanoparticles on okra seed germination under PEG-6000 simulated water deficit stress. Journal of Seed Science. 45 e202345029. (2023). https://doi.org/10.1590/2317-1545v45274866
Wang, J. et al. Azolla (Azolla pinnata) mitigated water stress and enhanced fruit yield of Mango plants (Mangifera indica L.) in Arid-degraded soil. J. Soil. Sci. Plant. Nutr. 23, 1173–1184. https://doi.org/10.1007/s42729-022-01112-3 (2023b).
Chen, Z. Q. & Song, R. Two-sided eigenvalue estimates for subordinate processes in domains. J. Funct. Anal. 226 (1), 90–113. https://doi.org/10.1016/j.jfa.2005.05.004 (2005).
Gohari, G. et al. Titanium dioxide nanoparticles (TiO2 NPs) promote growth and ameliorate salinity stress effects on essential oil profile and biochemical attributes of Dracocephalum moldavica. Sci. Rep. 10, 912. https://doi.org/10.1038/s41598-020-57794-1 (2020).
Abdel Latef, A. A., Srivastava, A. K., El-sadek, M. S., Kordrostami, M. & Tran, L. P. Titanium dioxide nanoparticles improve growth and enhance tolerance of broad bean plants under saline soil conditions. Land. Degrad. Dev. 29, 1065–1073. https://doi.org/10.1002/ldr.2780 (2018).
Rizwan, M., Iqbal, M. A., Imtiaz, H., Hamid, A. & Sheikh, U. A. A. Cactus Pear: a weed of dry-lands for supplementing food security under changing climate. Planta Daninha. https://doi.org/10.1590/s0100-83582020380100040 (2018).
Venkatachalam, P. et al. Enhanced Plant Growth Promoting Role of Phycomolecules Coated Zinc Oxide Nanoparticles with P Supplementation in Cotton (Gossypium Hirsutum L.). Plant Physiology and Biochemistry110118–127 (PPB, 2017). https://doi.org/10.1016/j.plaphy.2016.09.004
Yamori, W., Masumoto, C., Fukayama, H. & Makino, A. Rubisco activase is a key regulator of non-steady-state photosynthesis at any leaf temperature and, to a lesser extent, of steady-state photosynthesis at high temperature. Plant. Journal: Cell. Mol. Biology. 71 (6), 871–880. https://doi.org/10.1111/j.1365-313X.2012.05041.x (2012).
Eker, F. et al. A comprehensive review of nanoparticles: from classification to application and toxicity. Molecules (Basel Switzerland). 29 (15), 3482. https://doi.org/10.3390/molecules29153482 (2024).
Bassi, D., Menossi, M. & Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 8, 2327. https://doi.org/10.1038/s41598-018-20653-1 (2018).
Meinzer, F. C. & Zhu, J. Nitrogen stress reduces the efficiency of the C4CO2 concentrating system, and therefore quantum yield, in Saccharum (sugarcane) species. J. Exp. Bot. 49, 1227–1234. https://doi.org/10.1093/JXB/49.324.1227 (1998).
Mu, X. & Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant. Physiol. Biochemistry: PPB. 158, 76–82. https://doi.org/10.1016/j.plaphy.2020.11.019 (2021).
Ye, J. Y., Tian, W. H. & Jin, C. W. Nitrogen in plants: from nutrition to the modulation of abiotic stress adaptation. Stress Biology. 2, 4. https://doi.org/10.1007/s44154-021-00030-1 (2022).
Thabet, S. G. & Alqudah, A. M. Unraveling the role of nanoparticles in improving plant resilience under environmental stress condition. Plant. Soil. 503, 313–330. https://doi.org/10.1007/s11104-024-06581-2 (2024).
Wang, X., Xie, H., Wang, P. & Yin, H. Nanoparticles in plants: uptake, transport and physiological activity in leaf and root. Mater. (Basel Switzerland). 16 (8), 3097. https://doi.org/10.3390/ma16083097 (2023c).
Sembada, A. A. & Lenggoro, I. W. Transport of nanoparticles into plants and their detection methods. Nanomaterials (Basel Switzerland). 14 (2), 131. https://doi.org/10.3390/nano14020131 (2024).
Esfandiari, E. & Gohari, G. Response of ROS-Scavenging systems to salinity stress in two different wheat (Triticum aestivum L.) cultivars. Notulae Botanicae Horti Agrobotanici Cluj-napoca. 45, 287–291. https://doi.org/10.15835/nbha45110682 (2017).
Fazelian, N., Movafeghi, A., Yousefzadi, M., Rahimzadeh, M. & Zarei, M. Impact of silver nanoparticles on the growth, fatty acid profile, and antioxidative response of Nannochloropsis oculata. Acta Physiol. Plant. 42. https://doi.org/10.1007/s11738-020-03101-4 (2020).
Zia-Ur-Rehman, M. et al. Nanoparticles assisted regulation of oxidative stress and antioxidant enzyme system in plants under salt stress: A review. Chemosphere 314, 137649. https://doi.org/10.1016/j.chemosphere.2022.137649 (2023).
Laxa, M., Liebthal, M., Telman, W., Chibani, K. & Dietz, K. J. The role of the plant antioxidant system in drought tolerance. Antioxid. (Basel Switzerland). 8 (4). https://doi.org/10.3390/antiox8040094 (2019).
Wang, W. et al. Inorganic carbon utilization: A target of silver nanoparticle toxicity on a submerged macrophyte. Environ. Pollution (Barking Essex: 1987). 318, 120906. https://doi.org/10.1016/j.envpol.2022.120906 (2023).
Verma, N., Kaushal, P., Gahalot, D., Sidhu, A. K. & Kaur, K. Mechanistic aspect of zinc oxide nanoparticles in alleviating abiotic stress in Plants- A sustainable agriculture approach. BioNanoScience 13, 1645–1661. https://doi.org/10.1007/s12668-023-01192-0 (2023).
Zafar, S. et al. Impact of Zn nanoparticles synthesized via green and chemical approach on Okra (Abelmoschus esculentus L.) growth under salt stress. Sustainability 13 (7), 3694. https://doi.org/10.3390/su13073694 (2021).
Taha, R. S., Seleiman, M. F., Shami, A., Alhammad, B. A. & Mahdi, A. H. Integrated application of selenium and silicon enhances growth and anatomical structure, antioxidant defense system and yield of wheat grown in salt-stressed soil. Plants 10 (6), 1040. https://doi.org/10.3390/plants (2021).
Yue, J., Feliciano, T. J., Li, W., Lee, A. & Odom, T. W. Gold nanoparticle size and shape effects on cellular uptake and intracellular distribution of SiRNA nanoconstructs. Bioconjug. Chem. 28 (6), 1791–1800. https://doi.org/10.1021/acs.bioconjchem.7b00252 (2017).
Tumburu, L., Andersen, C. P., Rygiewicz, P. T. & Reichman, J. R. Molecular and physiological responses to titanium dioxide and cerium oxide nanoparticles in Arabidopsis. Environ. Toxicol. Chem. 36 (1), 71–82. https://doi.org/10.1002/etc.3500 (2017).
Khan, S., Zahoor, M., Sher Khan, R., Ikram, M. & Islam, N. U. The impact of silver nanoparticles on the growth of plants: the agriculture applications. Heliyon 9 (6), e16928. https://doi.org/10.1016/j.heliyon.2023.e16928 (2023).
Acknowledgements
DeepSeek R170b artificial intelligence tool was employed to refine and polish the English text of this manuscript.
Funding
This study was supported by the program for Natural Science Research in Guizhou Education Department (QJJ-[2023]-024), The Sixth Batch of Guizhou Province High-level Innovative Talent Training Program (GCC[2022]009), Innovation and Entrepreneurship Training plan for college students in Guizhou Province in 2024 (S2024109760212,2024109760072,2024109760064), and The program of Excellent Innovation Talents in Guizhou Province (GCC[2023]071).
Author information
Authors and Affiliations
Contributions
Jiyue Wang, Xiuhuan Meng, Shuai Wei and Xiaojian Tian, Methodology; Validation, formal analysis, Investigation; Nian Chen, Xiaojian Tian, and Shuai Wei; Data curation, Jiyue Wang, Xiuhuan Meng and Shuai Wei, Writing-original draft preparation; Denghong Shi, Chenggang Liang, Funding acquisition, Project administration. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, J., Tian, X., Wei, S. et al. Effect of nanoparticles on the growth of okra cultivated in soil affected by rocky desertification. Sci Rep 15, 18930 (2025). https://doi.org/10.1038/s41598-025-03838-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-03838-3
