
Abstract
Drought events are expected to become increasingly frequent, with potentially severe aftermaths on agriculture through direct and indirect effects on crops. It is thus necessary to understand how management practices can mitigate the impacts of droughts on yields, harmful organisms and ecosystem service providers in different soil contexts. Soil disturbance reduction is often suggested as one such practice.
In this study, we investigated the effects of drought (50% reduction in natural precipitation), tillage regime (conservation vs. conventional tillage), and the pivotal soil nutrient phosphorous on crop yield, as well as on the control of weeds, pests, and pathogens. We set our manipulative experiment in 18 arable fields in Northern Italy, and drought conditions were simulated with rainout shelters.
Drought had a negative effect on yields and increased the biomass and species richness of weeds. Conservation tillage had lower crop disease incidence but higher weed biomass than conventional tillage. Drought and conventional tillage both reduced the number of synergies between the different ecosystem services indicators. Soil phosphorus increased weed biomass, but decreased disease incidence in soybean. Arthropod pests and predators were not significantly affected by any of the tested variables.
Against the predictions, the effects of conservation tillage on drought mitigation and ecosystem services were mixed, indicating that complex combinations of multiple interventions will be required to reduce the negative effects of drought, weeds and pests under a changing climate.
Similar content being viewed by others


Introduction
Climate change is one of the greatest environmental challenges of our time with far-reaching consequences on many human activities, including agriculture1. Meteorological drought conditions, defined as precipitation deficits, are expected to become increasingly common2, which might negatively impact agriculture affecting crop yields3. Meteorological drought, in fact, is linked with agricultural drought, defined as a deficit in soil moisture and, therefore, in soil water availability to plants4. Increasing temperatures—while generally accelerating crop development and growth5—also lead to higher evapotranspiration and eventually water stress in plants, severely limiting crop productivity6. Climate-change-induced drought can also indirectly impact crop yields by generally favouring organisms noxious to the crop itself; these may include weeds, animal pests and pathogens. Given the wildly different biology and general ecology of these 3 groups, sustainable agroecological strategies to control them – either through natural enemies (biological control as an ecosystem service) or other means – need to focus on different aspects of the agroecosystem7. The situation is complicated by the fact that the same management action can differentially affect different ecosystem services, leading to trade-offs between them8. Consequently, there is a great interest in understanding how to minimize such trade-offs and exploit the generally rarer ecosystem service synergies—commonly defined, respectively, as negative and positive associations between two or more services in response to a given variable9,10.
Among harmful organisms, weeds deserve particular attention as they represent one of the most important biotic factors limiting crop yields worldwide11. Many weed species with traits related to efficient water exploitation are expected to expand their ranges and increase in abundance under drier conditions12. One of the key advantages of weeds over crops is their diversity, both at the genetic and community level, which allows them to exploit a variety of niches and persist through environmental changes and stresses, including attempts at weed control13. In contrast with weed biomass, however, weed diversity can also have positive effects on the functioning of agroecosystems, including support to beneficial arthropods and other wildlife14. Additionally, weeds do not only compete against the crop but also against each other, with some weed species potentially limiting dominant species and mitigating their negative effects15. Seeds are a key life stage through which drought could exert its effect on weeds. Seed germinability is in fact known to respond dramatically to drought-induced changes in the physical and biological features of soil16,17. Additionally, soil microorganisms can degrade buried seeds18 and therefore perform biological weed control19, an attractive alternative to herbicides and their deleterious environmental effects20. The effects of climate change on soil microbiomes and the associated ecosystem services are understudied21, but there is evidence that arid conditions can negatively impact soil microbe abundance and diversity22, potentially eroding their ability to degrade seeds – although physiological and evolutionary adaptation mechanisms might maintain soil microbial functions in certain conditions23.
Other important yield limiting factors for crops worldwide are invertebrate pests and pathogens, both of which are projected to be affected by climate change as well24. Just like weeds, many pest species could shift their ranges and generally benefit from climate change, increasing in numbers25 – although in some cases they might also suffer from negative impacts of more extreme climatic conditions and events26. Additionally, plants suffering from water stress in drought conditions might have reduced chemical defenses or increased nutrient concentration, becoming a higher-quality resource for herbivores as per the plant stress hypothesis27,28. Natural enemies such as predators or parasitoids are often seen as beneficial organisms, because they can act as an environmentally friendly tool for the control of pests29. Climate change effects on these biocontrol agents are expected to be complex, with the repercussions on pest control being equally difficult to predict30. As for plant pathogens, climate change could not only alter their biology and range, but also impact the physiology of host plants through drought and other stresses, potentially making plants more vulnerable to infection31.
Soil features and management play a pivotal role in influencing crop productivity and sustainability. Conservation tillage regimes that reduce soil disturbance improve soil physical features32 and favor a series of beneficial organisms living in the soil such as natural enemies of pests and weeds33,34 and microorganisms linked with soil fertility35, which in turn can influence the related ecosystem services34,36,37. Conservation tillage has also been proposed as a way to reduce carbon emissions and mitigate climate change38. Even more crucially in the context of this paper, conservation tillage has been proposed as a way to reduce the deleterious effects of drought on crops, either directly by reducing soil moisture loss39 or indirectly by favoring beneficial organisms40. Conservation tillage, however, also presents significant drawbacks, including influencing weed communities41 often resulting in a higher incidence of weeds in the fields42. A careful choice of soil management strategies might thus prove pivotal in offsetting some of the most harmful effects that climate change has on agriculture either through negative impacts on beneficial organisms or positive impacts on weeds, pests and pathogens. Another factor that pertains to soil and climate change is nutrient availability. The ability of plants to exploit soil nutrients can be significantly hampered by drought, as it has been shown for the important limiting macronutrient phosphorous43. It is well known that phosphorus has a pivotal role in influencing crop yield44 and modulating multiple ecosystem services45,46,47. It is also considered one of the best indicators of soil fertility 48. The ability of plants to uptake soil phosphorous has in fact been shown to be particularly sensitive to negative effects caused by drought when compared to other limiting macronutrients such as nitrogen49,50. It follows that the soil phosphorous concentration necessary for crops to reach a given yield may vary depending on the severity of drought conditions.
The aim of this study was to investigate the combined effects of climate-change-induced drought (simulated through rainout shelters), tillage regime and soil phosphorous availability (detected as a natural gradient in the experimental fields) on the yields of soybean (Glycine max (L.)) and wheat (Triticum aestivum L.), and on important parameters linked with the natural control of weeds, pests and pathogens; the latter included weed biomass production, weed diversity, weed seed decay, herbivore arthropod abundance, predatory arthropod abundance and soybean leaf disease incidence. The study took place in 9 field pairs (conventional vs. conservation tillage) in the Friuli plain of Northern Italy. We hypothesized that conservation tillage and natural phosphorous availability would mitigate the deleterious effects of drought on ecosystem services and yield, and that tillage and drought would also influence the number and identity of trade-offs and synergies among ecosystem services. Specifically, we expected conservation tillage and normal rainfall to increase synergies and reduce trade-offs, as stressful factors such as conventional tillage and excessive fertilization have been found to cause an opposite effect34. Given the high influence that weeds have on crops11, it was reasonable to assume that weed control and crop yield would be frequently correlated, while weed control and arthropod predator abundance was considered a likely trade-off due to the positive effects that weeds can have on natural enemy populations14.
Methods
Study area and experimental design
Experiments were carried out between 2018 and 2019 across 9 field pairs in the lowland area of the Friuli Venezia Giulia Region (NE Italy), within an agricultural landscape (c. 615 km2) characterized by temperate climate (mean annual precipitation of c. 1300 mm; mean annual temperature of 13 °C). Each pair included one conventional tillage field and one conservation tillage field (Fig. 1). In our case, all techniques characterized by non-inversion of soil for at least 8 to a maximum of 18 years were considered as “conservation tillage”. The distance between fields in a pair was less than 300 m except for one in which it was 900 m (Supplementary Table S1). The minimum distance between different pairs was 450 m, the maximum distance was 39 km; the specific coordinates of each experimental field can be found in Supplementary Table S1. In all fields, soybean was sown in May 2018 and harvested in October 2018, while wheat was sown in November 2018 and harvested in June 2019.

Experimental design with a picture showing a drought treatment plot under a rainout shelter. Each of the 9 field pairs included a conservation tillage field and a conventional tillage field; in each field, we selected a 20 × 60 m area where no agrochemicals were applied; in each of these areas, we selected a drought treatment plot and a control rainfed plot; in each plot, we had a normally sown half (for yield, disease, predators and pests) and an unsown half (for weeds); in the unsown half, se selected two subplots for the study of weeds, and we performed the seed burial experiment.
Soil characterization
Topsoil cores of 40 cm depth were collected in each field at the end of June 2018 and analyzed for content of carbon and phosphorous and for texture (i.e. silt, clay and sand content) (Supplementary Table S2). Soil phosphorus was quantified using the Olsen method51. A gradient in natural soil phosphorous availability detected across the selected experimental fields allowed to test the interactive effect of tillage regime and drought across varying soil fertility conditions. Phosphorous was not significantly linked with tillage regime in our data; additionally, phosphorous was only measured in normal rainfed conditions, as phosphorous soil levels are mainly dependent on artificial fertilization on the short term, while it is phosphorous availability to plants that is influenced by drought49,50.
Drought treatment
In each field, we defined a 20 × 60 m cultivated strip at the field margin in which agrochemicals (including fertilizers) were not used. Roughly at the center of this strip, we selected a 4 × 4 m drought treatment plot and an equally sized rainfed control plot (Fig. 1). The drought and the rainfed plot were placed at around 1 m from each other, to ensure homogeneity of conditions. Half of the area of each drought and rainfed plot was normally sown with the crop, while the other half was left unsown. The sown half represented a realistic situation for measuring crop yield production under the different treatments, while the unsown half was used to gather more data on weed diversity and growth. The latter is a less realistic scenario as weeds experienced increased growth, being free of competition with crops, but the method allowed us to gather more detailed data on weed community responses to drought, tillage and phosphorous that would be valuable for understanding baseline weed ecology.
The drought treatment was implemented using 4 × 4 m removable rainout shelters52. The roofs of the shelters were covered with 18 polycarbonate gutters (11 cm wide, 400 cm long and 3 cm deep) evenly distributed in order to exclude about 50% of the precipitations, with the space between one gutter and the next being roughly 11 cm wide. The shelters were 170 cm high on one side and 150 cm high on the opposite side, water was collected in a common gutter at the back of the shelter and then released on field at about 5 m from the plot through a plastic tube. Shelters were placed in the field one week after soybean sowing and temporarily removed in between soybean harvesting and wheat sowing. It is important to note that the exclusion of 50% of precipitations does not mean that shelters are able to simulate a 50% drought, as underground and air humidity can percolate from surrounding areas, which is inevitable under realistic field conditions; in spite of these known limitations, rainout shelters remain a standard and widespread method for the study of drought effects on vegetation, due to their versatility and ease of use53. We had a total of 36 experimental plots, 9 for each tillage/drought combination.
To better quantify the effect of the rainout shelters on soil moisture, from May to October 2018 (the soybean period) we monitored three field couples using ECH2O EC-5 soil humidity probes and ICT Em5b data loggers (Supplementary Table S1). In each drought and rainfed plot, two probes were buried at 20 cm of depth and data loggers registered humidity levels every 30 min from the placement to the removal of the shelters. Mean volumetric water contents per month are reported in Supplementary Table S3.
Crop yield
At soybean and wheat harvest phase, we harvested 1 m2 of each crop in each experimental plot using a plastic quadrat as a reference. We then dried the seeds (70 °C for 72 h) and measured their dry weight, which was used as a proxy for yield. In one of the rainfed plots of conservation tillage, wheat could not be harvested due to storm damage.
Herbivores, predators and pathogen survey
Two visual surveys were carried out on soybean in the first half of July and in the first half of September of 2018. During each survey, 20 soybean plants per plot were randomly selected, and all leaves of each plant were carefully checked. We recorded the number of potentially damaging herbivore arthropods and potentially beneficial predatory arthropods in each plot. Additionally, during these surveys we recorded the incidence of pathogens, measured as the percentage of surveyed plants that showed signs of infection. Herbivores, predators and pathogens were not identified to the species level, although herbivores and predators were categorized into their main families/orders. These surveys could not be carried out on wheat due to entomological operator unavailability.
Buried seed decay
We selected 3 weed species, representing different seed shapes and sizes, to test the seed decay capabilities of soil physical features and soil microbes under the different treatments. The species were Abutilon theophrasti Medicus, Amaranthus retroflexus L. and Digitaria sanguinalis (L.) Scop. Mature seeds of these species were hand-collected at the experimental farm of the University of Padova “Lucio Toniolo” (45°20′53.99″N, 11°57′5.46″E). The seeds were air-dried, mechanically cleaned, and stored at 4 °C until use. For each species, metal wire mesh bags (mesh size: 1 mm2) were prepared, each with 50 seeds sealed inside. This prevented invertebrates from preying on the seeds and confounding the effects of seed decay; the bags were flexible enough to allow the contact between soil and seeds54. In each drought and rainfed plot, we buried 4 seed bags per species. Due to organizational limitations, we could perform this experiment in only 8 of the 9 field pairs, resulting in a total of 128 buried seed bags per plant species. Bags were buried side by side at the depth of 15 cm, in a 60 × 25 cm hole, roughly at the center of the unsown half of the plot. Seeds remained underground from December 2018 to June 2019 (6 months). In one of the field pairs, data was lost due to external interference.
The recovered seeds initially underwent a crush test55, during which crushed seeds were classified as degraded. The remaining seeds were then tested for germinability by putting them in a growth chamber, i.e. seeds were placed in Petri dishes with filter paper bedding and 2 mL of distilled water, at 15/25 °C with a 12/12 h light/dark photoperiod54. Petri dishes were sealed with Parafilm, and more water was added when needed. Every 2 days, seeds were checked and the ones with a visible root were considered germinated and removed. When no new germinations were observed for a period of 10 days, seeds were moved in cold storage at 4 °C for 3 weeks and then moved back in the growth chamber and the process repeated, to interrupt a possible state of quiescence. After another 10 days since the last observed germination, the remaining seeds were cut in half, individually stored into Eppendorf vials with 1% tetrazolium and left at 30 °C in the dark for 24 h. At the end of the process, seeds were considered still vital if their interior was colored red. The proportion of the number of degraded and non-vital seeds over the total number of seeds was used as the measure of seed decay.
Weed diversity and biomass
Weed diversity was evaluated in two 1 × 1 m subplots placed in the unsown half of each drought and rainfed plot, at the distance of at least 70 cm from the border of the drought/rainfed plot. Due to botanical operator availability, surveys were carried out in February and April of 2019, after shelters had been in place exerting their action through all the soybean period, but before the sowing of wheat. Plant species were identified during the field survey, resulting in a measure of weed species richness in each subplot. At the time of wheat harvest in June 2019, all weed biomass was gathered from the subplots, dried for 48 h at 65 °C and then weighed. In one of the rainfed plots of conservation tillage, weed biomass could not be gathered due to external interference.
Data analysis: effects of drought, tillage and soil phosphorous
We used linear mixed effects models (LMMs) to evaluate the effect of drought treatment, tillage regime and soil phosphorus content on the yield of soybean and wheat as well as on weed seed decay, weed biomass and weed diversity, respectively. We started by testing the 3-way interaction between drought, tillage and phosphorous (which henceforth may be collectively indicated as “explanatory variables”) and removing non-significant interactions to avoid overfitting and to correctly interpret the main effects. For predator abundance, herbivore abundance and disease incidence, we ran a Poisson, negative binomial and binomial generalized linear mixed effects model (GLMM) respectively. The choice of LMMs rather than GLMMs for seed decay and weed species richness, and model choice in general, was informed by the validation plots. Weed seed decay was square-root transformed in order to meet model assumptions. Models were calculated using package lme456 and validation plots were generated with package DHARMa57 in R v3.6.258.
In the models for soybean and wheat yield, we used field pair ID and field ID as nested random factors. For most of the other models we used pair ID, field ID and plot ID as nested random factors to account for multiple measures in each plot – i.e. predators, herbivores and pathogens were surveyed twice per plot, weed biomass was collected in two subplots in each plot and weed seed decay was tested with multiple seed bags (4 for each of the 3 species) per plot. For weed species richness, we also used subplot ID as a nested random factor along with the previously mentioned ones, as each subplot was surveyed twice, while for the seed decay model we included seed species as the first nested random factor along with the others.
Data analysis: ecosystem services trade-offs and synergies
To evaluate trade-offs and synergies between ecosystem service indicators, we needed to have the same number of measurements for each indicator; we therefore calculated, for each plot, the mean values of the indicators that were measured multiple times per plot, so that they could be correlated with the indicators that were measured only once per plot. Additionally, some of our indicators (herbivore abundance, disease incidence, weed biomass, weed species richness) were actually indicators of ecosystem disservices, i.e. they were positively related to the success of noxious organisms. In order to calculate synergies and trade-offs, all indicators needed to be positively related with ecosystem services rather than with disservices. For disservices indicators, therefore, measures were transformed so that higher values corresponded to higher ecosystem services34. For instance, herbivore abundance measures were transformed into an indicator of herbivore control according to the formula herbivore control = (Xmax – X)/Xmax, where X is the value of each measure of herbivore abundance and Xmax is the highest value of X in the dataset, resulting in an index ranging from 0 (minimum herbivore control in the studied context) to an ideal 1 (total herbivore control in the studied context). The same principle was applied to disease control, weed biomass control and weed diversity control. We calculated the index also for the latter, considering weed species richness as a measure of weed success against crops, for simplicity and coherence with the other indicators—although, as mentioned in the introduction, weed diversity can also have positive effects on agroecosystems15,59.
Pairwise correlation coefficients are a simple and yet effective and widespread method to test for synergies and trade-offs between ecosystem services, including agricultural services34,60. We thus calculated Pearson correlations between the ecosystem service indicators for the conservation and conventional tillage datasets, and for the drought treatment and rainfed treatment datasets, separately. We did not consider the correlations between wheat yield and between organisms that were specifically sampled only on soybean (pests, predators, pathogens), nor the correlations of such organisms with weeds, as weeds were sampled after soybean harvest. We did consider the correlation between soybean yield and weed control (including seed decay) as crops can both influence soil microbial communities with long lasting consequences on crops that succeed them61 and, by outcompeting weeds, they can limit weed contribution to seed bank and have legacy effects on the weed control in following seasons62. Analyses were performed using package corrplot63 in R v3.6.2
Finally, to check the effect of tillage and drought stress on the overall provision of all the considered services, we used a Nonmetric MultiDimensional Scaling (NMDS) plot64 and carried out an analysis of similarities (ANOSIM)65, in both cases using Bray–Curtis distance66 and using the vegan67 package in R v3.6.2. All ecosystem service data were normalized into a 0 to 1 scale (were 1 represented the highest possible value of a given service in the dataset) for this analysis.
Results
Drought treatment, tillage regime and soil phosphorus had varying effects on the measured ecosystem service indicators – though for most indicators, interactions between explanatory variables were not significant and were thus removed from the models.
Crop yield
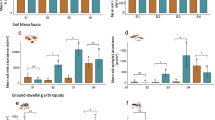
For soybean, crop yield was significantly lower in the drought treatment than in the rainfed control, with a 21% reduction (Table 1, Fig. 2a). Wheat yield, like soybean, was significantly lower in the drought treatment than in the rainfed control, with a 13% reduction (Table 1, Fig. 2b). Neither tillage regime nor soil phosphorus had significant effects on either crop yield.

Significant (P < 0.05) effects of drought treatment on soybean yield (a) and wheat yield (b) according to LMMs. Solid lines indicate model estimates, dots indicate partial residuals. Plots were generated using package visreg v2.7.0 in R. Explanatory variables and interactions that had non-significant effects are not shown.
Herbivores, predators and pathogens in soybean
Arthropod taxa surveyed in the study are reported in Supplementary Table S4. Tillage, drought treatment and phosphorous had no effect on herbivore or predator abundance (Table 1). Disease incidence, on the other hand, was higher in conventional than in conservation tillage, and this difference was more accentuated in the rainfed control (69%) than under drought conditions (53%) (Table 1, Fig. 3a). Disease incidence also steeply decreased with soil phosphorus content (Fig. 3b).

Significant (P < 0.05) effects of the interaction of drought treatment and tillage regime (a), and of phosphorous content (b) on soybean disease incidence according to the binomial GLMM. Solid lines indicate model estimates, partial residuals are indicated by dots for (a) and a rug for (b). Plots were generated using package visreg v2.7.0 in R. Explanatory variables and interactions that had non-significant effects are not shown.
Seed decay
Seed decay slightly decreased with increasing soil phosphorus content (Table 1, Fig. 4a), while tillage regime and drought treatment had no effect.

Significant (P < 0.05) effects of explanatory variables on weed control parameters, including interred weed seed decay (a), weed biomass (b–d) and weed biodiversity (e) according to LMMs. Seed decay incidence was square-root transformed to meet model assumptions. Solid lines indicate model estimates, dots indicate partial residuals. Plots were generated using package visreg v2.7.0 in R. Explanatory variables and interactions that had non-significant effects are not shown.
Weed biomass and diversity
The detailed list of weed species recorded in the study is reported in Supplementary Table S5. In the case of weed biomass, all three explanatory variables had significant effects with no interactions (Table 1). Conservation tillage, drought conditions and high phosphorous content all increased weed biomass separately (Fig. 4b–d), with increases caused by conventional tillage and drought being 26% and 27%, respectively. As for weed species richness, it increased (17%) with the drought treatment (Fig. 4e) with no significant effects of the other variables.
Ecosystem services trade-offs and synergies
The NMDS plot and the ANOSIM did not show any significant difference between tillage and drought treatments in terms of global ecosystem services provision, although tillage had a near-significant effect (0.05 < P < 0.10) (Supplementary Fig. S1). When considering pairwise correlations between services (Fig. 5), the drought scenario and conventional tillage presented a lower number of significant or near-significant trade-offs and synergies if compared to the alternative (normal rainfall and conservation tillage respectively).

Significant (P < 0.05) and near-significant (0.05 < P < 0.10) trade-offs (in red) and synergies (in green) between ecosystem service indicators in the two tillage regimes (upper row) and in the two rainfall scenarios (lower row). We report the value of the Pearson correlation coefficients between services. Grey squares represent relationships that were not tested due to variables being measured in different seasons.
The most frequent synergies were between soybean and wheat crop yield, and between the yield of these crops and weed control (either biomass or diversity), while the most frequent trade-offs were between soybean yield and indicators of pest control (either herbivore control or predator abundance).
Discussion
Our study showed that drought, tillage regime and soil phosphorus content can influence crop yield as well as weed, pest and pathogen control. In particular, both conservation and conventional tillage presented strengths and weaknesses, while simulated climate change drought mostly had undesirable effects. As expected, drought treatment reduced the yields of both studied crops, which can at least partially be a direct effect of the increase drought stress on the plant68. Indirect effects mediated by the increase in harmful organism populations are likely to depend mostly on weeds. Weeds were indeed confirmed as one of the most critical factors to consider in modern day ecosystems as well as in future climate change scenarios11,12. Weed biomass production was especially boosted by drought, conservation tillage and soil phosphorous.
Before further discussing the results, it is worth mentioning a potential limitation of our study, which is the fact that we only measured each variable for a single year (either 2018 or 2019). One consequence of this fact is that our study is only fit to reveal relatively short-term effects of drought, while multiple-year studies may be needed to reveal long-term effects69,70,71. Additionally, crop yield, weed and insect populations, as well as ecosystem functions can fluctuate—sometimes dramatically—over time, in response to weather fluctuations, climate shifts and other environmental variables. Studies aiming at fully capturing this variability are usually repeated for 5 or more years72,73,74, a time frame which was outside the scope or the budget of our project. The space-for-time approach, replacing temporal replication with spatial replication, is popular for studies focusing on long-time trends such as the effects of climate change75; similarly, the relatively high number of sampled sites spread across different areas of the study region ensures that we captured some degree of environmental variability in spite of the single-year samplings, improving the generalizability of our results. Nonetheless, our conclusions remain valid for the climate and environmental conditions found in the study areas in the sampling years, and further generalizability would require additional, longer-term experiments.
Drought treatment did not affect herbivore abundance, in contrast with studies showing a positive effect25,76. Effects on herbivores may require a longer period of drought to become apparent69,70. While some studies had reported negative effects on natural enemies77, others showed that climate change conditions, including drought, can occasionally increase natural enemy populations or biological control78. In our case, however, while predatory arthropods were more abundant in drought conditions, the difference was not significant, making it unlikely that they played a role in the lack of increase in pest arthropod numbers. Additionally, neither pests nor predators were influenced by tillage regime or soil phosphorus.
Disease incidence was influenced by tillage regime, with conventional tillage fields being more affected by crop pathogens than conservation tillage. This stands in contrast with a sizeable portion of the available literature, according to which conservation tillage practices makes crops more susceptible to disease79,80, although some sources also report lower disease incidence in conservation tillage81, which could be linked with a higher abundance and richness of non-pathogenic microorganisms competing with pathogens82,83. These discrepancies might be caused by differences in the physiological state of the crops, which may alter their susceptibility to infection31. For instance, conservation tillage is known to improve soil water conservation and reduce drought stress84, which is particularly important in arid conditions. This might mean that, in a future climate change scenario with increasing droughts, the recorded higher susceptibility of conventional tillage crops to disease might be furtherly exacerbated. It should however be noted that the drought treatment reduced disease incidence in conventional tillage (although it remained higher than in conservation tillage). This might be explained by a lower residual humidity on leaves in drought conditions, as leaf humidity favors leaf pathogens31. Considering also our results on drought, this factor might be more important than a possible increased susceptibility caused by drought, even potentially reducing the pathogen problems of conventional tillage agriculture. The clear trend of disease incidence steeply decreasing with soil phosphorus content is in line with most of the literature on the subject, which reports that phosphorus often improves plant health either through direct effects on pathogen multiplication or effects on plant metabolism and defenses85.
Weed biomass and richness were influenced by multiple factors. As expected, weed biomass was in general higher in conservation than in conventional tillage12,42 and, alongside weed species richness to a lesser degree, was positively influenced by drought conditions favouring hardy, drought-resistant weed species over the crops. While there was no significant interaction between these two explanatory variables, it is likely that, in future climate change scenarios, conservation tillage fields will be at a higher risk of suffering from problems related to weed biomass increases if compared with conventional fields, requiring efficient weed control strategies, such as thick mulching and robotic weeding, to remain viable without herbicide overuse. It is however important to note that usually cover crops are included in the conservation tillage regime also as a weed control strategy86, and this element was not incorporated in our manipulative experiments.
Soils with high phosphorus content generally increased weed biomass production. Phosphorous is indeed a nutrient that is more efficiently exploited by weed species rather than crops87. However, another factor may be at play, as phosphorus was also linked with lower buried seed decay. While the design of our study is not fit to provide a definite mechanistic explanation of this phenomenon, it is known that phosphorous can influence soil microbial communities, and while this influence is generally regarded as beneficial, in some conditions it might shift community composition in a way that alters certain ecosystem functions88. This might mean that weed control through seed decay would be increasingly inefficient in soils with high phosphorous content. Overall, conventionally tilled fields in low-phosphorus soils might be at a lower risk of weed-related problems in increasingly dry conditions.
Altogether, drought at the levels tested in our experiment did not significantly alter the overall provision of the suite of ecosystem services that we studied, while tillage only had a near-significant effect. When considering specific pairwise correlations between services, however, it appears that stressful environmental factors—both drought conditions and conventional tillage—reduce the number of significant or near-significant ecosystem service indicator trade-offs and synergies, similarly to what was reported by Tamburini et al. (2016)34 for conventional tillage and artificial fertilization. In climate change drought conditions, due to the observed loss of positive associations between ecosystem services, it could thus become more difficult to improve multiple services with a single management action, and each service might require a specifically tailored action. In that scenario, such management actions would nonetheless have a lower chance of negatively impacting non-target services, due to reduced negative associations among them.
Synergies between soybean yield and wheat yield indicate that these different crop types tend to be positively influenced by a similar set of variables. Alongside that, synergies between crop yield and weed biomass/diversity control were relatively common. This is in line with our initial expectations and, when considering the interaction between weeds and wheat, it further confirms the role of weeds as the main biotic limiting factor for crops. When considering the interaction between soybean and weeds, the relatively common yield/weed control synergies also confirm the importance of successful crop competition against weeds in limiting weed incidence in the following seasons. On the other hand, the most common trade-offs were between soybean yield and pest control, in the form of either herbivore suppression or predator abundance, which seems to indicate that increasing crop success might result in more severe pest infestations. Additionally, soybean success in conventional tillage was linked simultaneously with an increase in the control of weed diversity and a decrease in the number of arthropod predators; as we hypothesized, since weed diversity can support biological control agents by offering favorable microhabitats and alternative prey14, success in weed control might result in negative consequences for pest control in some instances.
Conclusions
Our results highlight some important consequences of drought on crop yield and multiple parameters related to weed, pest and pathogen control, as well as the potential effects of soil management regime through tillage and soil phosphorus content, paving the way for future research lines. Experiments focused on the potentially interactive effects of multiple environmental factors on yield are pivotal, given the importance of the topic for agricultural productivity in a rapidly transforming world, and our still limited knowledge of the topic89. Contrary to expectations, the effects of conservation tillage on ecosystem services and drought mitigation were mixed, suggesting a challenging path ahead for climate change adaptation. Weeds were confirmed to be the most likely biotic limiting factors for crops in climate change scenarios. Sustainable weed management strategies will become increasingly important, especially for conservation tillage fields, which are more susceptible to weeds. The use of fast-growing cover crops able to outcompete weeds90, the protection of seed predator communities36 and the correct management of soil for enhancing seed-degrading microorganisms18 might all represent promising ways of mitigating the problem without an increase in herbicide use, all while continuing to reap some of the benefits associated with conservation tillage (including improved disease control). Soil phosphorous seems to be an important variable which might require a fine tuning to balance between its desirable (disease control) and undesirable effects (weed increase, possibly also through seed decay reduction). There is indication, however, that this type of trade-offs (and synergies) between ecosystem services will be less frequent under climate change conditions, especially in conventionally tilled fields, implying that we will require complex combinations of interventions to reduce the negative effects of drought, weeds and pests.
Data availability
The datasets generated and analyzed during the described study are available from the corresponding author on reasonable request.
References
Wreford, A., Moran, D. & Adger, N. Climate change and agriculture: Impacts, adoption, and mitigation. Climate Change and Environmental Concerns: Breakthroughs in Research and Practice (OECD, 2010). https://doi.org/10.4018/978-1-5225-5487-5.ch001.
Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 3, 52–58 (2013).
Marini, L. et al. Crop rotations sustain cereal yields under a changing climate. Environ. Res. Lett. 15, 124011 (2020).
Behrang Manesh, M. et al. Linkage of agricultural drought with meteorological drought in different climates of Iran. Theor .Appl. Climatol. 138, 1025–1033 (2019).
Richter, G. M. & Semenov, M. A. Modelling impacts of climate change on wheat yields in England and Wales: Assessing drought risks. Agric. Syst. 84, 77–97 (2005).
Raderschall, C. A., Vico, G., Lundin, O., Taylor, A. R. & Bommarco, R. Water stress and insect herbivory interactively reduce crop yield while the insect pollination benefit is conserved. Glob. Chang. Biol. 27, 71–83 (2021).
Deguine, J.-P. et al. Agroecological crop protection for sustainable agriculture. Adv. Agron. 178, 1–59 (2023).
Raudsepp-Hearne, C., Peterson, G. D. & Bennett, E. M. Ecosystem service bundles for analyzing tradeoffs in diverse landscapes. Proc. Natl. Acad. Sci. USA 107, 5242–5247 (2010).
Howe, C., Suich, H., Vira, B. & Mace, G. M. Creating win-wins from trade-offs? Ecosystem services for human well-being: A meta-analysis of ecosystem service trade-offs and synergies in the real world. Glob. Environ. Chang. 28, 263–275 (2014).
Vallet, A. et al. Relationships between ecosystem services: Comparing methods for assessing tradeoffs and synergies. Ecol. Econ. 150, 96–106 (2018).
Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 144, 31–43 (2006).
Peters, K., Breitsameter, L. & Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 34, 707–721 (2014).
Allan, E. et al. More diverse plant communities have higher functioning over time due to turnover in complementary dominant species. Proc. Natl. Acad. Sci. USA 108, 17034–17039 (2011).
Barbercheck, M. E. & Wallace, J. Weed-insect interactions in annual cropping systems. Ann. Entomol. Soc. Am. 114, 276–291 (2021).
Adeux, G. et al. Mitigating crop yield losses through weed diversity. Nat. Sustain. 2, 1018–1026 (2019).
Hoveland, C. S. & Buchanan, G. A. Weed seed germination under simulated drought. Weed Sci. 21, 322–324 (1973).
Karimmojeni, H., Bazrafshan, A. H., Majidi, M. M., Torabian, S. & Rashidi, B. Effect of maternal nitrogen and drought stress on seed dormancy and germinability of Amaranthus retroflexus. Plant Species Biol. 29, 1–8 (2014).
Chee-Sanford, J. C., Williams, M. M., Davis, A. S. & Sims, G. K. Do microorganisms influence seed-bank dynamics?. Weed Sci. 54, 575–587 (2006).
Kennedy, A. C. & Kremer, R. J. Microorganisms in weed control strategies. J. Prod. Agric. 9, 480–485 (1996).
Annett, R., Habibi, H. R. & Hontela, A. Impact of glyphosate and glyphosate-based herbicides on the freshwater environment. J. Appl. Toxicol. 34, 458–479 (2014).
Jansson, J. K. & Hofmockel, K. S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 18, 35–46 (2020).
Maestre, F. T. et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. 112, 15684–15689 (2015).
Allison, S. D. Microbial drought resistance may destabilize soil carbon. Trends Microbiol. 31, 780–787. https://doi.org/10.1016/j.tim.2023.03.002 (2023).
Gregory, P. J., Johnson, S. N., Newton, A. C. & Ingram, J. S. I. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 60, 2827–2838 (2009).
Skendžić, S., Zovko, M., Živković, I. P., Lešić, V. & Lemić, D. The impact of climate change on agricultural insect pests. Insects 12, 440 (2021).
Jactel, H., Koricheva, J. & Castagneyrol, B. Responses of forest insect pests to climate change: not so simple. Curr. Opin. Insect. Sci. 35, 103–108 (2019).
Gutbrodt, B., Dorn, S., Unsicker, S. B. & Mody, K. Species-specific responses of herbivores to within-plant and environmentally mediated between-plant variability in plant chemistry. Chemoecology 22, 101–111 (2012).
White, T. C. 1% A hypothesis to explain outbreaks of looper caterpillars, with special reference to populations of Selidosema suavis in a plantation of Pinus radiata in New Zealand. Oecologia 16, 279–301 (1974).
DeBach, P. & Rosen, D. Biological Control by Natural Enemies (Cambridge University Press, Cambridge, 1991).
Thomson, L. J., Macfadyen, S. & Hoffmann, A. A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 52, 296–306 (2010).
Elad, Y. & Pertot, I. Climate change impacts on plant pathogens and plant diseases. J. Crop. Improv. 28, 99–139 (2014).
Chabert, A. & Sarthou, J. P. Conservation agriculture as a promising trade-off between conventional and organic agriculture in bundling ecosystem services. Agric. Ecosyst. Environ. 292, 106815 (2020).
Jacobsen, S. K., Sigsgaard, L., Johansen, A. B., Thorup-Kristensen, K. & Jensen, P. M. The impact of reduced tillage and distance to field margin on predator functional diversity. J. Insect. Conserv. https://doi.org/10.1007/s10841-022-00370-x (2022).
Tamburini, G., De Simone, S., Sigura, M., Boscutti, F. & Marini, L. Soil management shapes ecosystem service provision and trade-offs in agricultural landscapes. Proc. Biol. Sci. 283, 20161369 (2016).
Liu, C. et al. Soil bacterial diversity and potential functions are regulated by long-term conservation tillage and straw mulching. Microorganisms 8, 836 (2020).
Lami, F., Boscutti, F., Masin, R., Sigura, M. & Marini, L. Seed predation intensity and stability in agro-ecosystems: Role of predator diversity and soil disturbance. Agric. Ecosyst. Environ. 288, 106720 (2020).
Höflich, G. et al. Influence of long-term conservation tillage on soil and rhizosphere microorganisms. Biol. Fertil. Soils 29, 81–86 (1999).
Almagro, M., Garcia-Franco, N. & Martínez-Mena, M. The potential of reducing tillage frequency and incorporating plant residues as a strategy for climate change mitigation in semiarid Mediterranean agroecosystems. Agric. Ecosyst. Environ. 246, 210–220 (2017).
Holland, J. M. The environmental consequences of adopting conservation tillage in Europe: reviewing the evidence. Agric. Ecosyst. Environ. 103, 1–25 (2004).
Madejón, P. et al. Could conservation tillage increase the resistance to drought in Mediterranean faba bean crops?. Agric. Ecosyst. Environ. 349, 139295 (2023).
Boscutti, F. et al. Conservation tillage affects species composition but not species diversity: A comparative study in Northern Italy. Environ. Manag. 55, 443–452 (2015).
Peigné, J., Ball, B. C., Roger-Estrade, J. & David, C. Is conservation tillage suitable for organic farming?. A review. Soil Use Manag 23, 129–144 (2007).
Zhang, H. et al. Drought promotes soil phosphorus transformation and reduces phosphorus bioavailability in a temperate forest. Sci. Total Environ. 732, 139295 (2020).
Syers, J. K., Johnston, A. E. & Curtin, D. Efficiency of soil and fertilizer phosphorus use. Food Agric. Org. United Nations 9, 5–50 (2008).
Macdonald, G. K. et al. Guiding phosphorus stewardship for multiple ecosystem services. Ecosyst. Health Sustain. 2, 01251 (2016).
Macintosh, K. A. et al. Transforming soil phosphorus fertility management strategies to support the delivery of multiple ecosystem services from agricultural systems. Sci. Total Environ. 649, 90–98 (2019).
Jarvie, H. P. et al. The pivotal role of phosphorus in a resilient water-energy-food security nexus. J. Environ. Qual. 44, 1049–1062 (2015).
Gosling, P. & Shepherd, M. Long-term changes in soil fertility in organic arable farming systems in England, with particular reference to phosphorus and potassium. Agric. Ecosyst. Environ. 105, 425–432 (2005).
Mariotte, P. et al. Plant uptake of nitrogen and phosphorus among grassland species affected by drought along a soil available phosphorus gradient. Plant Soil 448, 121–132 (2020).
He, M. & Dijkstra, F. A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 204, 924–931 (2014).
Sims, J. T. Soil test phosphorus: Olsen P. in Methods of phosphorus analysis for soils, sediments, residuals, and waters 20–21 (2000).
Kundel, D. et al. Design and manual to construct rainout-shelters for climate change experiments in agroecosystems. Front. Environ. Sci. 6, 1–9 (2018).
Hoover, D. L., Wilcox, K. R. & Young, K. E. Experimental droughts with rainout shelters: a methodological review. Ecosphere 9, e02088 (2018).
Nikolić, N. et al. Weed seed decay in no-till field and planted riparian buffer zone. Plants 9, 293 (2020).
Sawma, J. T. & Mohler, C. L. Evaluating seed viability by an unimbibed seed crush test in comparison with the tetrazolium test. Weed Technol. 16, 781–786 (2002).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. CRAN: Contributed Packages Preprint at https://doi.org/10.32614/CRAN.package.DHARMa (2020).
R Core Team. R: A language and environment for statistical computing. Preprint at https://www.R-project.org/ (2019).
Schumacher, M., Dieterich, M. & Gerhards, R. Effects of weed biodiversity on the ecosystem service of weed seed predation along a farming intensity gradient. Glob. Ecol. Conserv. 24, e01316 (2020).
Mouchet, M. A. et al. An interdisciplinary methodological guide for quantifying associations between ecosystem services. Glob. Environ. Chang. 28, 298–308 (2014).
Benitez, M. S., Ewing, P. M., Osborne, S. L. & Lehman, R. M. Rhizosphere microbial communities explain positive effects of diverse crop rotations on maize and soybean performance. Soil Biol. Biochem. 159, 108309 (2021).
Melander, B., Rasmussen, I. A. & Olesen, J. E. Legacy effects of leguminous green manure crops on the weed seed bank in organic crop rotations. Agric. Ecosyst. Environ. 302, 107078 (2020).
Wei, T. & Simko, V. corrplot: Visualization of a Correlation Matrix. CRAN: Contributed Packages Preprint at https://doi.org/10.32614/CRAN.package.corrplot (2017).
Minchin, P. R. An evaluation of the relative robustness of techniques for ecological ordination. in Theory and models in vegetation science: Proceedings of Symposium, Uppsala, July 8–13, 1985 (eds. Prentice, I. C. & van der Maarel, E.) 89–107 (Springer Netherlands, Dordrecht, 1987). https://doi.org/10.1007/978-94-009-4061-1.
Clarke, K. R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143 (1993).
Bray, J. R. & Curtis, J. T. An Ordination of the Upland Forest Communities of Southern Wisconsin. vol. 27 (1957).
Oksanen, J. et al. vegan: Community Ecology Package. CRAN: Contributed Packages Preprint at https://doi.org/10.32614/CRAN.package.vegan (2025).
Fathi, A. & Tari, D. B. Effect of drought stress and its mechanism in plants. Int. J. Life Sci. 10, 1–6 (2016).
Hassan, K., Carrillo, Y. & Nielsen, U. N. Prolonged drought causes negative plant-soil feedbacks in grassland species under field conditions. Soil Biol. Biochem. 172, 108772 (2022).
Hassan, K., Carrillo, Y., Islam, T. & Nielsen, U. N. Prolonged drought legacies influence the performance of foliar herbivores on legumes through shifts in plant–soil biotic interactions. Entomol. Exp. Appl. 173, 60–70 (2025).
Power, S. A. et al. DRI-Grass: A new experimental platform for addressing grassland ecosystem responses to future precipitation scenarios in South-East Australia. Front. Plant. Sci. 7, 1373 (2016).
Bopp, M. C., Kazakou, E., Metay, A. & Fried, G. Relative importance of region, seasonality and weed management practice effects on the functional structure of weed communities in French vineyards. Agric. Ecosyst. Environ. 330, 107892 (2022).
Jungers, J. M. et al. Diversifying bioenergy crops increases yield and yield stability by reducing weed abundance. Sci. Adv. 7, 8531–8560 (2021).
Paredes, D., Rosenheim, J. A. & Karp, D. S. The causes and consequences of pest population variability in agricultural landscapes. Ecol. Appl. 32, e2607 (2022).
Lovell, R. S. L., Collins, S., Martin, S. H., Pigot, A. L. & Phillimore, A. B. Space-for-time substitutions in climate change ecology and evolution. Biol. Rev. 98, 2243–2270 (2023).
Cannon, R. J. C. The implications of predicted climate change for insect pests in the UK, with emphasis on non-indigenous species. Glob. Chang. Biol. 4, 785–796 (1998).
Morecroft, M. D., Bealey, C. E., Howells, O., Rennie, S. & Woiwod, I. P. Effects of drought on contrasting insect and plant species in the UK in the mid-1990s. Glob. Ecol. Biogeogr. 11, 7–22 (2002).
Staley, J. T. et al. Drought stress differentially affects leaf-mining species. Ecol Entomol 31, 460–469 (2006).
Guo, X. W., Fernando, W. G. D. & Entz, M. Effects of crop rotation and tillage on blackleg disease of canola. Can. J. Plant Path. 27, 53–57 (2005).
Bockus, W. W. & Shroyer, J. P. The impact of reduced tillage on soilborne plant pathogens. Annu. Rev. Phytopathol. 36, 485–500 (1998).
Hu, X. et al. Conventional and conservation tillage practices affect soil microbial co-occurrence patterns and are associated with crop yields. Agric. Ecosyst. Environ. 319, 107534 (2021).
Muneret, L. et al. Evidence that organic farming promotes pest control. Nat. Sustain 1, 361–368 (2018).
van Bruggen, A. H. C. & Finckh, M. R. Plant diseases and management approaches in organic farming systems. Annu. Rev. Phytopathol. 54, 25–54 (2016).
Sher, A. et al. Conservation tillage improves productivity of sunflower (Helianthus annuus L.) under reduced irrigation on sandy loam soil. PLoS ONE 16, 1–9 (2021).
Walters, D. R. & Bingham, I. J. Influence of nutrition on disease development caused by fungal pathogens: Implications for plant disease control. Annal. Appl. Biol. 151, 307–324 (2007).
Teasdale, J. R. et al. Cover crops and weed management. in Non chemical weed management: Principles, concepts and technology 239 (CABI, 2007).
Little, N. G., Ditommaso, A., Westbrook, A. S., Ketterings, Q. M. & Mohler, C. L. Effects of fertility amendments on weed growth and weed-crop competition: A review. Weed Sci. 69, 132–146 (2021).
Wu, W. et al. Meta-analysis of the impacts of phosphorus addition on soil microbes. Agric. Ecosyst. Environ. 340, 108180 (2022).
Rezaei, E. E. et al. Climate change impacts on crop yields. Nat. Rev. Earth Environ. 4, 831–846 (2023).
Brust, J., Claupein, W. & Gerhards, R. Growth and weed suppression ability of common and new cover crops in Germany. Crop. Prot. 63, 1–8 (2014).
Acknowledgements
Source vector art should be attributed to Dave Angelini (herbivore icon, https://creativecommons.org/licenses/by/3.0/), Melissa Broussard (predator icon, https://creativecommons.org/licenses/by/3.0/), Freepik.com (wheat and soybean icons) and Flaticon.com (disease control and weed seed decay icons, designed by users Smashicon and imaginationlol respectively).
Funding
This research was funded by the project BIRD171833/17 "Predazione e mortalità naturale dei semi come strategia di controllo delle infestanti in agricoltura conservativa of the University of Padova, and by the project "Efficacia dell’agricoltura conservativa per la mitigazione dei cambiamenti climatici in paesaggi agricoli" (CUP: F66C18000040002) of ERSA - Agenzia Regionale per lo Sviluppo Rurale. Open access fees were funded by the National Biodiversity Future Center (NBFC), Project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU (Project code CN_00000033). NBFC was not involved on study design, writing or any decision about the paper.
Author information
Authors and Affiliations
Contributions
F. L.: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. F. B.: Funding acquisition, Investigation, Methodology, Resources, Supervision, Writing – review & editing. S. B.: Funding acquisition, Resources, Writing – review & editing. M. F.: Funding acquisition, Resources, Writing – review & editing. R. M.: Investigation, Methodology, Writing – review & editing. N. N.: Investigation, Methodology, Writing – review & editing. M. S.: Investigation, Resources, Writing – review & editing. L. M.: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lami, F., Boscutti, F., Barbieri, S. et al. Drought conditions, tillage regime and soil phosphorous modulate the incidence of weeds, pests and pathogens in arable crops. Sci Rep 15, 24383 (2025). https://doi.org/10.1038/s41598-025-04042-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-04042-z
