
Abstract
Biologically synthesized nanoparticles act as growth stimulants and enhance plant tolerance to stresses. This study aimed to investigate the effect of silver nanoparticles (AgNPs) biosynthesized using an aqueous extract of Moringa oleifera as a plant stimulant on the morphological and physiological traits of Pelargonium hortorum under drought stress conditions in greenhouse experiments. First, the synthesis of AgNPs and their morphology and particle size distribution were examined using Transmission Electron Microscopy (TEM) and Scanning Electron Microscopy (SEM). Then, to evaluate the effect of nanoparticles on geranium growth, a factorial experiment was conducted in a completely randomized design, including three levels of drought stress (30, 60, and 90% field capacity) and four levels of nanoparticle foliar application (0, 10, 20, and 30%) with three replications over six months. The results showed that the synthesized nanoparticles were highly homogeneous and spherical, with an average size ranging from 8 to 12 nanometers. Foliar application at a 10% concentration significantly improved all evaluated parameters and partially mitigated the adverse effects of drought stress. The highest phenotypic coefficient of variation belonged to stem diameter (15.27%) and catalase enzyme activity (17.62%). The application of nanoparticles at a 10% concentration significantly increased leaf blade length and width, plant height, flower diameter, petal anthocyanin content, catalase enzyme activity, and chlorophyll content compared to the control. However, regarding stem diameter and flowering time, the highest increase was observed at 20% and 30% nanoparticle concentrations, respectively. Additionally, AgNPs at a 10% concentration increased phosphorus and potassium content in geranium leaves, whereas higher concentrations and drought stress reduced nitrogen and potassium levels. Therefore, foliar application of AgNPs (10%) can reduce the adverse effects of drought stress and serve as a practical, effective, and cost-efficient approach to mitigating stress and improving geranium growth and yield under water-limited conditions.
Similar content being viewed by others

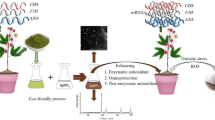

Introduction
Pelargonium hortorum is a perennial, herbaceous, evergreen flowering plant that blooms throughout spring, summer, and part of autumn. It is cultivated as both an indoor ornamental plant and in outdoor landscapes. Additionally, geranium is one of the most widely used ornamental plants and ranks second among bedding plants in international trade1.
Drought stress is one of the most significant factors reducing agricultural crop yields worldwide2and the extent of this reduction depends on the plant’s response and the stress conditions3. Therefore, mitigating the harmful effects of drought stress or enhancing plant tolerance to drought stress plays a crucial role in overcoming the challenges of agricultural production in arid and semi-arid regions4. Climate change and rising temperatures have led to increased evaporation and transpiration, intensifying water scarcity and soil salinity in arid and semi-arid regions5. The physiological and morphological responses of different plant species to drought stress vary. Drought damages the photosynthetic apparatus, reducing plant metabolism, growth, and yield. Consequently, plants adopt drought avoidance or tolerance strategies for growth and adaptation under drought conditions6.
In recent decades, there has been a growing interest in the application of nanoparticles in agriculture, genetics, and plant breeding7. Nanoparticles offer promising opportunities for mitigating the detrimental effects of biotic and abiotic stresses, contributing to the goal of sustainable agriculture on a global scale. Due to their impact on stress tolerance and crop quality, research on the application of nanoparticles is expanding8. Various reports indicate that nanoparticles enhance plant tolerance to stresses by addressing nutritional deficiencies, increasing enzymatic activity, and facilitating the colonization of roots by growth-promoting rhizobacteria under stress conditions9.
Among metallic nanoparticles, AgNPs are more widely used due to their physical and chemical properties, including catalytic efficiency, optical activity, electrical conductivity, and surface plasmon resonance10. Additionally, they play an effective role in increasing leaf surface area and dry weight of plant organs11.
Nanoparticles (NPs) can be produced through various biological, physical, and chemical methods12. However, the biological method is more cost-effective and environmentally friendly13and one of these methods is the synthesis of nanoparticles using plant extracts14. The potential use of plant materials in the green synthesis of silver nanoparticles (AgNPs) is particularly attractive due to their antioxidant-rich phytochemicals, making the process easy, cost-effective, and eco-friendly15. Several studies have been conducted on the synthesis of AgNPs using extracts from plants such as Papaver somniferum16Aloe vera17and Acacia farnesiana18.
Moringa oleifera, belonging to the Moringaceae family, grows in semi-tropical, semi-arid, and tropical regions19. Currently, it is widely cultivated in the Americas, Africa, Mexico, Malaysia, India, Sri Lanka, the Philippines, and Indonesia. It is also used in emerging countries as a source of micronutrients, a natural insecticide, and a metabolic conditioner to prevent endemic disorders20. This plant is one of the most nutrient-rich species, providing antioxidants and amino acids with anti-aging and anti-inflammatory properties. This “miracle tree” is a rich source of calcium, iron, vitamin C, and highly digestible proteins21.
Moringa leaf extract (MLE) is a potent natural growth stimulant because it contains zeatin, a natural cytokinin derivative. Moringa leaf extract is also a source of minerals, vitamins, amino acids, proteins, and phenolics, making it a potential plant growth enhancer22,23. Moringa leaf extract contains endogenous cytokinins such as zeatin, isopentyl adenine, and dihydro zeatin, which play a crucial role in enhancing stress resistance and promoting plant vigor24.
Nowadays, nanotechnology, as the most advanced technology of the present era, has penetrated all aspects of human, animal, plant, environmental, and industrial life. Through its innovations, it has influenced their current state and future, comparable to revolutionary inventions like the steam engine and the internet, with the potential to bring about fundamental transformations in other technologies and various scientific and industrial fields25.
As a result of human activities, nanoparticles have entered the environment; however, there is still no precise information about the impact of these materials on the environment and their toxic effects26. Researchers believe that AgNPs, due to their small size and high solubility, are absorbed by plants more rapidly. Therefore, applying these materials creates optimal conditions for plant growth and prevents oxidative damage induced by environmental stresses8. It appears that AgNPs can act as signaling molecules by enhancing the plant’s antioxidant defense system, reducing damage caused by reactive oxygen species, thereby increasing membrane stability and reducing ion leakage.
Additionally, studies on the foliar application of AgNPs on the morphological characteristics and antioxidant properties of artichoke under salinity stress have shown that foliar application of AgNPs improves growth traits, including plant weight, fresh and dry root weight, leaf number, and length, as well as physiological properties such as flavonoids, phenols, and antioxidants27.
Given that previous reports have demonstrated the potential of Moringa nanoparticles in enhancing crop resistance to drought stress28salinity29and heavy metal stresses23the combined strategy of drought alleviation and growth stimulation using Moringa-based AgNPs remains underexplored.
This study aims to investigate the effect of AgNPs synthesized from Moringa extract under drought stress conditions on vegetative growth and the improvement of pelargonium performance, which has not been previously studied.
Materials and methods
Preparation of aqueous extract
First, Moringa oleifera leaves were obtained from the medicinal plant collection at Hormozgan University. After washing, the leaves were dried in the shade and then ground into powder. The aqueous extract was prepared using the soaking method by adding 20 g of Moringa leaf powder to 100 mL of deionized water and heating it in a water bath for 70 min to obtain the primary extract. The resulting extract was filtered using Whatman No. 1 filter paper and a syringe filter and stored in dark-colored glass bottles at 4 °C for further studies30.
Synthesis of AgNPs
10 mL of concentrated aqueous Moringa leaf extract was mixed with 90 mL of 1 mM silver nitrate solution and stirred for 20 min using a magnetic stirrer. The mixture was then kept in the dark at 25 °C for 48 h. The color change from light brown to dark brown indicated the completion of the reaction. The synthesized nanoparticles were recovered by centrifugation at 15,000 rpm for 15 min, thoroughly washed with sterile distilled water, and dried overnight31.
Characterization of synthesized nanoparticles
Transmission electron microscopy (TEM) analysis was conducted at 70 kV accelerating voltage to determine the particle size and shape. A drop of the nanoparticle solution was placed on a copper grid and dried at room temperature. The size and morphology of the synthesized nanoparticles were examined using scanning electron microscopy (SEM).
The crystalline properties, structure, and crystallite size of the nanoparticles were analyzed using X-ray diffraction (XRD) with a Rigaku-Miniflex X-ray diffractometer (Rigaku, Tokyo, Japan), employing Cu-Kα radiation (λ = 0.15406 nm) within a 2θ range of 20° to 80°. SEM analysis was performed using a JEOL JSM5600 system (JEOL Ltd., Yuki, Japan).
For electron microscopy analysis, the reaction mixture was centrifuged three times at 12,000 rpm for 15 min. Then, a few drops of the obtained precipitate were placed on an aluminum foil, dried at room temperature, and imaged using an electron microscope.
To identify the potential biomolecules responsible for the reduction and capping of biosynthesized nanoparticles, Fourier-transform infrared (FT-IR) spectroscopy analysis was performed at room temperature using a Nicolet 6700 FTIR spectrometer.
Cultivation conditions of geranium
This experiment was conducted in the 2024–2025 agricultural year at Gorgan University, located at 36°30’ N latitude, 53°57’ E longitude, and an altitude of 155 m above sea level. Seeds of the common ornamental geranium (Pelargonium hortorum cv. Bullseye) were sown in 70-cell seedling trays. In early autumn, seedlings at the 4–6 leaf stage and 6–8 cm in height were transplanted into 12 cm diameter plastic pots. The growing medium was prepared by mixing 20% (v/v) loamy clay soil and 80% (v/v) black peat (both sieved through a 2 mm mesh). The potted plants were grown in a glass-covered greenhouse under natural light, with daytime and nighttime temperatures maintained at 25 ± 3 °C and 18 ± 3 °C, respectively. The cultivation period lasted from November 2023 to May 2024.
Application of treatments
This study examined the effects of two factors: three levels of irrigation regimes (30%, 60%, and 90% of field capacity) and four concentrations of AgNPs synthesized using aqueous Moringa leaf extract (0, 10, 20, and 30%). The experiment was conducted as a factorial arrangement based on a completely randomized design (CRD) with three replications over six months. The selected concentrations were determined based on previous reports and preliminary tests on this plant32.
Water deficit treatments were applied gravimetrically. Initially, equal amounts of gravel were placed at the bottom of each pot for drainage, and each pot was filled with an equal weight of soil (3 kg per pot). The soil in each pot was saturated with water and allowed to drain on a perforated surface for 48 h to reach field capacity. The pots were then weighed, and their soil was oven-dried at 105 °C for 48 h to determine the soil’s water-holding capacity. After calculating the soil’s field capacity moisture content, the amount of water required for each irrigation treatment was determined by weighing a sample pot in each block, calculating the water deficit, and adding the required amount of water33.
Silver nanoparticle foliar applications were carried out in three stages at 10-day intervals, using concentrations of 0, 10, 20, and 30 mL/L. Distilled water was used as the control. In each application, 100 mL of solution was sprayed. Before applying the AgNPs, ultrasonic dispersion (Hielscher UP400s, Teltow, Germany) was performed for 30 min for each concentration to ensure homogeneity and better dispersion of nanoparticles.
Trait measurements
Growth and flowering traits
Plant height was measured from the base of the stem to the last leaf using a ruler in centimeters34. To measure leaf blade length and width, all leaves of each replication were collected separately and analyzed using Image J software after photographing them, with the results expressed in square centimeters. Stem diameter was measured at the lower third of the main stem in millimeters35 using a digital caliper (Digital caliper Guanglu 0–100 mm). Flowering time was recorded as the number of days from seed sowing to the appearance of the first inflorescence36.
Petal anthocyanin content
Petal anthocyanin content was measured using the Wagner (1979) method. In this method, 0.05 g of fresh geranium petal tissue was placed in 10 mL of acidified methanol (methanol: hydrochloric acid at a 99:1 ratio) in a test tube. The mixture was transferred to a Falcon tube and kept in a dark environment at room temperature for 24 h. The extract was then centrifuged at 10,000 rpm for 15 min, and the absorbance of the supernatant was read at 550 nm using a spectrophotometer37. The total anthocyanin content was calculated using an extinction coefficient (ε) of 33,000 M−1 cm−1 with the following equation:
A represents the absorbance of the solution,
b is the cuvette path length (1 cm),
c is the concentration of the target solution (mg/g).
Catalase enzyme activity (CAT)
Catalase (CAT) enzyme activity was measured by assessing the decomposition of hydrogen peroxide using a spectrophotometer at 240 nm38. The reaction mixture contained 2600 µL of 50 mM phosphate buffer, 400 µL of hydrogen peroxide buffer, and 40 µL of enzyme extract. After mixing (by sealing the cuvette with parafilm and inverting it once), the enzymatic reaction rate was recorded as changes in absorbance over time (OD/min) at 240 nm for one minute. Enzyme activity was calculated using the Beer-Lambert law with a catalase extinction coefficient of 40 mM−1 cm−1 and expressed as µMol/gFW.min.
Measurement of nutrient elements
To measure the nutrient content, wet digestion of plant samples and extraction were performed using concentrated nitric acid and hydrogen peroxide39. For this purpose, plant samples were dried at 75 °C for 72 h, ground, and completely powdered. One gram of the obtained plant powder was weighed using a digital balance with a precision of 0.001 g and transferred into test tubes. Then, 10 mL of concentrated nitric acid (65%) was added to each sample, and they were placed in a water bath at 65 °C for 2 h. After removing the samples and cooling them to room temperature, 2.6 mL of 20% hydrogen peroxide was added to each sample. After the reaction and cooling, the samples were filtered using Whatman No. 42 filter paper and brought to a final volume of 20 mL. The phosphorus content in plant samples was then measured by the phosphovanadomolybdate complex formation method at a wavelength of 430 nm using a spectrophotometer (Varian, Cary 100). Potassium content was determined by flame emission spectrometry using a flame photometer (405G). Nitrogen content was measured by titration after distillation using the Kjeldahl apparatus40.
Chlorophyll (SPAD)
The greenness index, an indicator of chlorophyll content, was measured using a hand-held chlorophyll meter (SPAD Model 505), which provides a non-destructive method of assessing chlorophyll levels directly from the leaf surface41.
Statistical analysis
Descriptive statistics were initially performed to assess the quality and distribution of the collected data, aiming to simplify and summarize the dataset for clearer interpretation. Subsequently, inferential statistical analyses were conducted to evaluate treatment effects and interrelationships among traits. The Duncan’s multiple range test (DMRT), executed using SPSS software (version 26; IBM Corp., Armonk, NY, USA), was applied to analyze variance, compute mean squares, and assess experimental errors to identify significant differences among treatment means. DMRT at a significance level of p ≤ 0.05 was used to identify distinctions between means. Pearson correlation coefficients were calculated using SPSS (version 26; IBM Corp., Armonk, NY, USA) to evaluate the relationships between different parameter pairs. Additionally, stepwise regression analysis at p < 0.01 was conducted to determine the key traits affecting geranium flower diameter. Path analysis was performed to assess the direct and indirect effects of each trait within the model based on the traits that best explained the variations. All the aforementioned calculations (simple correlation coefficients, stepwise regression, and path analysis) were conducted using SPSS version 26, while causal diagrams were drawn using AMOS version 24.
Results
Preparation of aqueous extract and synthesis of AgNPs
After obtaining Moringa leaves from the medicinal plant collection of Hormozgan University, the aqueous extract of the plant was prepared using the soaking method. The resulting concentrated extract was then mixed with a 1 mM silver nitrate solution, and after 48 h, a visible color change from light brown to dark brown was observed, indicating initiation of nanoparticle synthesis and the completion of the reaction.
Characteristics of the synthesized nanoparticles
The size and morphology of the synthesized nanoparticles were examined using transmission electron microscopy (TEM) and scanning electron microscopy (SEM). Based on the obtained images, the nanoparticles synthesized via the green chemistry method were highly homogeneous and spherical in shape, with an average particle size ranging between 8 and 12 nm (Fig. 1).

Transmission electron microscope (TEM) image (A) and scanning electron microscope (SEM) image (B) of AgNPs synthesized by the aqueous extract of Moringa oleifera leaves.
Fourier transform infrared spectroscopy (FTIR)
The presence of distinct peaks in the FTIR spectra (Fig. 2A) within the 4000 –3500 cm−1 range indicates the stretching vibrations of the O–H stretching vibrations. Additionally, the appearance of broad and overlapping peaks in the 2800–3500 cm−1 region suggests the presence of functional groups associated with metallic nanoparticles. Furthermore, in the 500–1500 cm−1 range, the stretching peaks correspond to C = O, N-H, and O-C bond vibrations. Since the organic compounds from the plant extracts surround the nanoparticles (NPs) and contribute to their stabilization, the vibrational movements of these organic groups are observable in the FTIR spectrum (Fig. 2A).
To further assess the purity of the synthesized nanoparticles, the zeta potential analysis was analyzed, and the presence of AgNPs synthesized by Moringa was confirmed through Zeta Potential analysis (Fig. 2B).

Fourier transforms infrared (FTIR) spectrum of nanoparticles (A) and Zeta Potential spectrum of AgNPs synthesized by the aqueous extract of Moringa oleifera leaves (B).
Effect of nanoparticle foliar application on morphophysiological parameters of geranium
Descriptive statistics of evaluated traits
The descriptive statistical parameters, including mean, standard deviation, minimum, maximum, range of variation, and phenotypic coefficient of variation (PCV) for various traits, are summarized in Table 1. Among these parameters, the highest PCV percentages were observed for stem diameter (15.27%) and catalase enzyme activity (17.62%), while the lowest PCV percentages were recorded for flower diameter index (2.33%) and number of flowering branches (3.45%).
An analysis of Table 1 reveals that flowering time, chlorophyll content, and flower diameter exhibited the widest range of variation. However, these traits did not have the highest phenotypic coefficient of variation.
Effect of water stress and foliar application of synthesized nanoparticles
The results of foliar application of AgNPs synthesized using Moringa aqueous extract at four concentrations (0, 10, 20, and 30%) under drought stress conditions (30%, 60%, and 90% field capacity) over a six-month period indicated that increasing drought stress at 30% field capacity significantly reduced leaf blade length and width, plant height, stem diameter, number of flowering branches, flower diameter, and catalase enzyme activity in geranium. However, in contrast, flowering time, petal anthocyanin content, and chlorophyll content increased (Table 2).
The application of synthesized AgNPs, particularly at a 10% concentration, significantly enhanced leaf blade length and width, plant height, number of flowering branches, flower diameter, catalase enzyme activity, petal anthocyanin content, and chlorophyll content in geranium compared to the control. However, it also delayed flowering time (Table 3).
Leaf blade length and width
The mean comparison of the interaction effects of drought stress (30%, 60%, and 90% field capacity) and foliar application of synthesized AgNPs on the leaf blade length and width of geranium revealed that increased drought stress at 30% field capacity significantly reduced these traits (Table 2). However, foliar application of AgNPs at a lower concentration enhanced leaf blade length and width, whereas higher concentrations resulted in a decline in these parameters compared to the control.
The maximum leaf blade length (4.68 cm) and width (5.46 cm) were observed at a 10% nanoparticle concentration. Conversely, the lowest values were recorded at a 30% nanoparticle concentration. The interaction effects of varying concentrations of AgNPs and drought stress further demonstrated that increasing nanoparticle concentration along with higher drought stress levels resulted in a reduction in leaf blade length and width, with the highest values observed in the control plants (Fig. 3).

The effect of drought stress and AgNPs synthesized by Moringa on leaf blade length (A) and leaf blade width (B). The data are presented as mean ± standard deviation (n = 3). Data followed by the same letters are not significantly different according to Duncan’s multiple range test at P ≤ 0.05.
Plant height and stem diameter
The mean comparison of the effects of AgNPs on plant height and stem diameter of geranium showed that drought stress mildly reduced plant height and stem diameter (Table 2). However, the foliar application of nanoparticles increased plant height compared to the control but had no significant effect on stem diameter.
At a 10% nanoparticle concentration, the highest plant height (7.33 cm) was recorded, whereas the lowest height (6.56 cm) was observed at a 30% concentration (Table 3).
The interaction effect of AgNPs and drought stress showed that the highest plant height occurred at a 20% nanoparticle concentration and 90% field capacity, while the lowest plant height was recorded at a 30% nanoparticle concentration under 30% field capacity stress (Fig. 4A).
Regarding the interaction between AgNPs and drought stress on stem diameter, no significant effect was observed among different nanoparticle concentrations (Fig. 4B).

The effect of drought stress and AgNPs synthesized by Moringa on plant height (A) and stem diameter (B). The data are presented as mean ± standard deviation (n = 3). Data followed by the same letters are not significantly different according to Duncan’s multiple range test at P ≤ 0.05.
Flower diameter and number of flowering branches
The mean comparison of the effects of AgNPs on flower diameter showed that indicated drought stress led to a decrease in flower diameter in geranium at 30% field capacity (Table 2). However, the highest flower diameter (7.16 cm) was observed in the 10% nanoparticle treatment. As nanoparticle concentration increased beyond this level, the flower diameter decreased compared to the control (Table 3).
According to Fig. 5, the interaction effect of AgNPs and drought stress had a significant impact on flower diameter. The smallest flower diameter was recorded at 30% nanoparticle concentration under 30% field capacity stress. Interestingly, at 90% field capacity, flower diameter increased with higher nanoparticle concentrations, suggesting that nanoparticles had a growth-promoting effect under optimal irrigation conditions (Fig. 5A).
Regarding the number of flowering branches, different concentrations of AgNPs and drought stress did not show a significant effect. However, at 10% nanoparticle concentration under 60% field capacity, the highest number of flowering branches was recorded (Fig. 5B).

The effect of drought stress and AgNPs synthesized by Moringa on flower diameter (A) and the number of flowering branches (B). The data are presented as mean ± standard deviation (n = 3). Data followed by the same letters are not significantly different according to Duncan’s multiple range test at P ≤ 0.05.
Flowering time and petal anthocyanin content
The mean comparison of different concentrations of AgNPs and drought stress on flowering time showed that increased drought stress at 30% field capacity delayed flowering by 14 days compared to the control (Table 2). Similarly, increasing nanoparticle concentration was found to prolong the flowering time. Specifically, the foliar application of 30% nanoparticles delayed flowering by 15 days compared to the control, whereas the 20% nanoparticle concentration resulted in a relatively shorter delay in flowering (Table 3).
The interaction effect of drought stress and different nanoparticle concentrations on flowering time was significant. The results indicated that under 30% field capacity stress, flowering time increased with higher nanoparticle concentrations. However, under 90% field capacity, the longest flowering time was observed at 10% nanoparticle concentration, while the shortest flowering time was recorded at 10% nanoparticles under 60% field capacity (Fig. 6A).
Regarding petal anthocyanin content, the highest amount was observed under drought stress at 30% field capacity (Table 2). Notably, the greatest effect of the synthesized nanoparticles was observed at a 10% concentration, with a marked increase in anthocyanin content. The lowest anthocyanin content was found in the control group and at a 30% nanoparticle concentration (Table 3).
The interaction effect of AgNPs and drought stress revealed that the highest anthocyanin content was found at 30% nanoparticle concentration under 90% field capacity, showing a significant increase compared to the control (Fig. 6B). However, under drought stress at 30% field capacity, anthocyanin content decreased as nanoparticle concentration increased.

The effect of drought stress and AgNPs synthesized by Moringa on flowering time (A) and petal anthocyanin (B). The data are presented as mean ± standard deviation (n = 3). Data followed by the same letters are not significantly different according to Duncan’s multiple range test at P ≤ 0.05.
Catalase enzyme and chlorophyll content
The mean comparison of AgNPs’ effects on catalase enzyme and chlorophyll content showed that chlorophyll levels increased under drought stress at 30% field capacity, whereas catalase enzyme activity significantly declined (Table 2).
In terms of nanoparticle application, the highest levels of catalase activity and chlorophyll content were recorded at the 10% AgNPs concentration, while the lowest values were observed in the control and the 30% AgNP treatment (Table 3).
The interaction effect of AgNPs and drought stress revealed that the highest chlorophyll content was observed at 30% nanoparticle concentration under 90% field capacity, showing a significant increase compared to the control (Fig. 7B). However, under severe drought conditions (30% field capacity), chlorophyll content declined with increasing nanoparticle concentration.
The interaction effect of AgNPs and drought stress on catalase enzyme activity was not significant. Overall, as nanoparticle concentration increased under drought stress, catalase enzyme activity decreased. The lowest catalase enzyme activity was observed at 30% nanoparticle concentration under drought stress at 30% field capacity (Fig. 7A).

The effect of drought stress and AgNPs synthesized by Moringa on catalase enzyme activity (A) and chlorophyll content (B). The data are presented as mean ± standard deviation (n = 3). Data followed by the same letters are not significantly different according to Duncan’s multiple range test at P ≤ 0.05.
Leaf nutrient content (NPK)
According to the results presented in Table 4, the application of AgNPs had a significant effect on leaf nitrogen, phosphorus, and potassium levels under drought stress, with the strongest responses observed at 60% field capacity and AgNP concentrations of 10% and 20%.
The results of the mean comparison of the interaction effects of nanoparticles and drought stress on nitrogen content showed that drought conditions led to a marked decrease in nitrogen levels. However, spraying AgNPs at a 10% concentration increased nitrogen content under drought stress. The highest nitrogen content was observed in the control plant under drought stress at 60% field capacity, while the lowest nitrogen content was seen at a 30% nanoparticle concentration under drought stress at 30% field capacity.
Furthermore, AgNP foliar spraying was found to increase phosphorus content, with the maximum phosphorus and potassium levels recorded at the 10% AgNP concentration under 60% field capacity. In contrast, the lowest phosphorus concentration occurred in control plants and those treated with 10% AgNPs under 90% field capacity, while the lowest potassium content was detected in plants treated with 30% nanoparticles under severe drought stress (30% field capacity).
Correlation
The correlation matrix revealed strong interrelationships among most traits. Leaf width exhibited the highest positive correlation with catalase enzyme activity (r = 1.00), while petal anthocyanin content showed a similarly strong and significant correlation with chlorophyll content (r = 1.00) (Fig. 8). These results indicate that increases in catalase activity and chlorophyll content are associated with corresponding increases in leaf width and anthocyanin accumulation, respectively.
In contrast, flowering time demonstrated negative and significant correlations with most of the other traits, suggesting that as stress-tolerant or growth-promoting traits increase, flowering tends to be delayed.

Heat map of mutual relations of variables in correlation coefficient for growth indicators.
Principal component analysis (PCA)
Based on the component loading plot, the best traits can be selected according to the various components. In most cases, the first two or three principal components, which account for the largest proportion of variance, are used for this purpose. In this study, the first principal component explained 43.822% of the variance, the second accounted for 18.501%, and the third contributed 12.370%, cumulatively explaining 74.693% of the total variance. All traits associated with the first component had positive and significant loadings (Table 5). For the first component (catalase enzyme and leaf width), the second component (chlorophyll and petal anthocyanin), and the third component (number of flowering branches and flower diameter) showed the greatest influence.
Path analysis
To explore the underlying relationships among traits, path analysis was employed. This technique dissects the correlation between independent and dependent variables into direct and indirect effects, enabling a more accurate interpretation of causal relationships.
High correlations among traits can result in multicollinearity, which distorts the estimates and reduces the reliability of conventional path models. To address this, sequential path analysis—a method designed to minimize multicollinearity and simplify complex interrelationships was utilized in this study.
Direct effects of the examined traits on flower diameter are presented in Table 6. Stepwise regression analysis identified three models with increasing explanatory power. Among the traits, flowering time, catalase enzyme activity, and plant height exhibited the most significant direct effects on flower diameter.
The coefficient of determination (R²) for the first model was 48.2%, for the second model 64.6%, and for the third model 69.3%, indicating a progressive improvement in model fit.
In the first model, flowering time had a direct negative effect on flower diameter, with a coefficient of − 0.470.
In the second model, the direct effects were − 0.577 for flowering time and + 0.422 for catalase enzyme activity (CAT).
In the third model, the direct effects were − 0.535 for flowering time, + 0.346 for catalase enzyme activity, and + 0.237 for plant height (Table 7).
These results demonstrate that flowering time consistently exerted a strong negative influence on flower diameter across all models, while CAT activity and plant height contributed positively, enhancing flower size.
The indirect causality analysis revealed that plant height and catalase enzyme activity (CAT) were influential traits that not only affected flowering time but also exerted a positive direct impact on geranium flower diameter. As illustrated in Fig. 9, both traits showed indirect pathways to flower diameter through their effects on other variables.
The findings indicate that while flowering time influenced flower diameter only in the first model, CAT activity and plant height contributed positively to flower diameter in the second and third models, both directly and indirectly. These results underscore the critical role of CAT activity and plant height in determining flower size.
Accordingly, to enhance flower diameter, agronomic and nutritional management practices should strategically target improvements in these key traits. This approach would likely contribute to better performance and yield in geranium cultivation.

Sequential path model showing the indirect reciprocal relationships between flower diameter, flowering time index, catalase enzyme activity, and plant height.
Discussion
Drought is one of the major limiting factors restricting agricultural production, severely affecting crop yield42. Moreover, as key abiotic stresso drought impairs several physiological processes, including respiration, photosynthesis, and stomatal regulation43. Consequently, nearly all plant life processes are directly or indirectly affected by environmental stress conditions.
Among the innovative approaches to improving plant stress tolerance, the use of nanoparticles (NPs), particularly nano-fertilizers, has emerged as a promising strategy in modern agriculture. Due to their high surface area and small size, nanoparticles can easily penetrate plant cell membranes, leading to enhanced nutrient uptake efficiency, reduced soil toxicity, minimized adverse effects associated with overuse of conventional fertilizers, and a decreased need for frequent fertilizer applications44 Once absorbed, nanoparticles can modify the plant’s cellular environment by forming new pores in cell walls and membranes, thereby facilitating the uptake of water and mineral nutrients45. The increase in water leads to an increase in turgor pressure, resulting in changes in the osmotic potential of the cell and greater mineral salt absorption. At this point, enzymatic and hormonal signals increase, and the activity of growth enzymes, as well as hormones like gibberellin and auxin, rises, promoting tissue expansion and development46. Furthermore, previous studies have reported that nanoparticles may influence hormone distribution and cytoskeletal organization (e.g., microtubules) in root cells, thereby accelerating cell division and expansion in water-absorbing zones. Nanoparticles can also enter plant cells through aquaporins and ion channels, potentially affecting the expression of genes and altering physiological pathways associated with growth and stress response. Collectively, these mechanisms contribute to improved plant performance, productivity, and stress resilience47.
In recent years, biosynthesis of nanoparticles using plant extracts has attracted more attention48. The advantages of using plant extracts for the biosynthesis of AgNPs include non-toxicity, biocompatibility, cost-effectiveness, and the production of high-purity nanoparticles49. In the present study, AgNPs were synthesized using the aqueous extract of Moringa oleifera, leveraging its natural reducing and stabilizing properties to produce effective and environmentally friendly nanomaterials. Moringa Oleifera is a rich source of minerals, vitamins, proteins, and antioxidants50which can be utilized for the green synthesis of nanoparticls. Given the plant’s tropical origin and beneficial chemical compounds, this research aimed to reduce drought stress damage on some morphological and physiological traits of geranium using silver nanoparticles AgNPs synthesized by an aqueous extract of Moringa in 2024 at the Gorgan University of Agricultural sciences and Natural Resources. The results showed that the Moringa extract had a high potential to produce nanoparticles, and since the size and shape of nanoparticles play an essential role in many of their medicinal applications51the size and morphology of the AgNPs synthesized by the aqueous extract of Moringa leaves were investigated by electron microscopy. Initially, the reaction led to a color change from pale yellow to dark brown to black, resulting from the interaction between the plant extract and silver nitrate solution, similar to the findings of Ahmad et al. (2010)52 and Reddy and Gandhi (2012)53which is the first indication of AgNPs production. The color change and the appearance of strong absorption peak around 420–460 nm obtained from UV/Visible spectroscopy of the plant extract and silver nitrate solution confirmed AgNPs production, aligning closely with Roopan et al. (2013)54 results. The color change and the peak observed in the wavelength range of 400 to 500 nm in the spectroscopic spectrum were related to the surface plasmon resonance (SPR), induced by the collective oscillations of free electrons interacting with the electromagnetic field, which is specific to each type and size of nanoparticles. Transmission electron microscopy (TEM) and scanning electron microscopy (SEM) images revealed that the size and morphology of the synthesized nanoparticles in this study were highly uniform, spherical, and had an average size of 8 to 12 nm. The spherical and uniform morphology of the synthesized nanoparticles indicates the successful encapsulation of active biomolecules from the M. oleifera extract. Similarly, in the 2024 study by Fotoohiyan et al., the AgNPs synthesized using wormwood (Artemisia) extract had spherical shapes with an approximate size of 20 nm and had the highest absorption at 420 nm55.
Furthermore, in this study, different concentrations (0, 10, 20, and 30%) of AgNPs synthesized by the aqueous extract of Moringa were evaluated for their effects on morphological (leaf length and width, plant height, stem diameter, number of flowering branches, and flowering time) and physiological traits (petal anthocyanin, catalase enzyme activity, and chlorophyll content). Positive effects of AgNPs on growth indices of cucumber (performance, number of leaves, and plant height) were reported56. Additionally, AgNPs increased the growth characteristics and biochemical properties of leaf mustard (Brassica juncea), bean (Phaseolus vulgaris), and corn (Zea mays)57,58. Furthermore, nanoparticles increased the germination percentage, germination speed, seedling weight, and seedling length in savory (Satureja hortensis)8. Nanoparticles have both positive and negative effects on plant growth, and their impact depends on various factors including composition, concentration, size, and physical and chemical properties, as well as the plant species, and varies from one plant to another59.
In this study, the effect of spraying AgNPs synthesized by the aqueous extract of Moringa at four concentrations (0%, 10%, 20%, and 30%) under drought stress conditions (30%, 60%, and 90% of field capacity) over a 6-month period was investigated. The results showed that among the descriptive statistics, the phenotypic coefficient of variation (PVC) for stem diameter (15.27%) and catalase enzyme activity (17.62%) had the highest percentages of variation among the traits studied, while flower diameter and the number of flowering branches had the lowest variation with values of 2.33% and 3.45%, respectively. Furthermore, the results indicated that increasing drought stress (30% field capacity) significantly reduced the leaf blade length and width, plant height, stem diameter, number of flowering branches, flower diameter, and catalase enzyme activity in geranium. In contrast, flowering time, petal anthocyanin content, and chlorophyll content increased. In fact, with increased water stress and the reduction of turgor pressure in guard cells, stomata closed, limiting photosynthesis and thus reducing cell growth and plant organ development60. Additionally, the increased allocation of photosynthates to defense responses rather than growth was another reason for the reduction in geranium plant height under drought conditions. Similarly, drought stress caused a reduction in fenugreek61 and cumin62 heights. The increase in carotenoids and anthocyanins under stress is due to the protective role of these pigments, which help protect chlorophyll from photooxidation63.
The use of AgNPs, especially at a concentration of 10%, significantly increased the leaf blade length and width, plant height, number of flowering branches, flower diameter, catalase enzyme activity, petal anthocyanin content, and chlorophyll content in geranium plants compared to the control. However, it also delayed flowering time. The application of AgNPs enhanced the vegetative growth of the plant and increased the relative leaf humidity. Nanoparticles, due to their hydrophilic nature, gradually release water molecules, helping the plant perform well under stress conditions64. It has been reported that the use of AgNPs at a concentration of 60 mg/L significantly controlled salt stress in savory plants and increased relative content, which aligns with the present study’s results8. Additionally, AgNPs have been shown to increase growth characteristics (stem and root length, leaf area) and biochemical properties (chlorophyll, carbohydrate, protein content, and antioxidant enzymes) in canola, bean, and corn57,58.
Regarding the interaction between different concentrations of AgNPs and drought stress, the results showed an increase in leaf blade length and width. The highest plant height was observed at a 20% nanoparticle concentration and 90% field capacity, while the lowest plant height and flower diameter were found at a 30% nanoparticle concentration under 30% field capacity drought stress. Notably, under drought conditions with 90% field capacity, flower diameter increased with increasing nanoparticle concentration, suggesting that nanoparticles had a stimulatory effect on the growth of geranium plants under mild drought stress.
No significant interaction between AgNP concentrations and drought stress was observed for stem diameter. Furthermore, the diameter and number of flowers in henna⁶⁷ and sage⁶⁸ increased with AgNP treatments, which aligns with our findings. However, different nanoparticle concentrations and drought stress levels did not significantly affect the number of flowering branches in geranium. Only at a 10% nanoparticle concentration under 60% field capacity was the highest number of flowering branches observed.
The results of the interaction between nanoparticle concentrations and drought stress on flowering time showed that increasing drought stress to 30% field capacity delayed flowering time by 14 days compared to the control. Likewise, increasing nanoparticle concentration significantly delayed flowering time, with a 30% concentration delaying it by 15 days. Interestingly, a 20% nanoparticle concentration reduced flowering time. The interaction between drought stress and nanoparticle concentrations was significant; under 30% field capacity, flowering time increased with higher nanoparticle concentrations. At 90% field capacity, the longest flowering time was observed at the 10% concentration, whereas the shortest occurred at the same concentration under 60% field capacity.
Regarding the effect of synthesized nanoparticles on petal anthocyanin content under 30% field capacity, the highest anthocyanin content was recorded at the 10% nanoparticle concentration, while the lowest was seen in the control and at the 30% concentration. In the interaction between AgNPs synthesized by Moringa and drought stress, the highest petal anthocyanin content was observed at a 30% nanoparticle concentration under 90% field capacity, showing a significant increase over the control. However, under 30% field capacity, anthocyanin content decreased as nanoparticle concentration increased.
Physiological traits play a vital role in plant survival and adaptation to environmental stresses, making these indicators important for evaluation. The increase in carotenoids and anthocyanins under stress is attributed to the protective role of these pigments, which protect chlorophyll from photo-oxidation63. The measurement of photosynthetic pigment content is a key parameter that can indicate the plant’s photosynthesis rate and growth conditions.
The results of this study indicated that increasing drought stress caused damage to cell membranes. To mitigate this oxidative stress, petal anthocyanin and chlorophyll content increased in geranium plants under 30% field capacity drought stress, while catalase enzyme activity significantly decreased. The highest increase in catalase and chlorophyll content was observed with the 10% AgNPs treatment, while the lowest levels of catalase and chlorophyll were found in the control and at the 30% nanoparticle concentration.
Spraying with 10% AgNPs under drought stress enhanced antioxidant enzyme activity in geranium plants. Drought stress likely led to a decrease in chlorophyll content and photosynthesis, as well as oxidative stress, causing energy to be diverted for antioxidant activities, which resulted in reduced transport of photosynthetic products to flowers and, ultimately, a decrease in the number of flowering branches and flower diameter in plants under drought stress. Similarly, Han et al. (2022) reported that drought stress in licorice species led to decreased growth characteristics and increased antioxidant enzyme activity65.
In the study by Ehsanpour and Homaee (2015), chlorophyll content significantly increased at lower concentrations of AgNPs, while higher concentrations resulted in a decrease66. Similarly, high concentrations of nanoparticles caused a further reduction in chlorophyll pigments in Spirodela polyrhiza67A. thaliana68green algae69and black mustard70. Ma et al. (2013) showed that a reduction in chlorophyll content could result from lipid peroxidation of chloroplast membranes due to oxidative stress under high concentrations of AgNPs68. The reduction in chlorophyll pigments under AgNPs treatment may also indicate the toxic effect of AgNPs on geranium plants.
Nanoparticles affect plant metabolism by increasing the activity of nitrate reductase and glutamate dehydrogenase, thus enhancing nitrogen metabolism and boosting growth and photosynthesis. The effective concentration of elicitors varies depending on the plant species, as a concentration that stimulates one plant may have no effect on another71. In this study, since the petal anthocyanin content in geranium plants under AgNP treatment increased significantly compared to the control, this may indicate a stress response induced by AgNPs. Anthocyanins are antioxidant molecules that protect plants from damage induced by oxidative stress and free radicals72. The increase in anthocyanins under AgNP treatment is similar to results obtained in studies on Arabidopsis plants73.
Additionally, the increase in catalase activity helps reduce peroxidase effects during various stresses and plays a crucial role in plant resistance to environmental stress conditions. This enzyme is involved in the breakdown of cellular peroxides under environmental stress74. Furthermore, the increase in chlorophyll and petal anthocyanin content under the 10% AgNP treatment suggests a protective role of AgNPs as an ethylene production inhibitor.
In the study by Najafi et al. (2013), the effect of nanoparticles on growth characteristics and secondary metabolite production in Phausalus vulgaries was observed75. Treatment with silver nanoparticles increased plant growth and some secondary metabolites, such as flavonoids, at lower concentrations, which enhanced antioxidant properties. This may be due to the effect of AgNPs on enzymes responsible for producing these compounds. Silver nanoparticles can modify plant growth and secondary metabolite production.
In any case, the application of nanoparticles not only improves vegetative growth and plant performance under drought stress compared to control plants but also delays flowering time. However, at higher concentrations, the results are the opposite, showing an inhibitory effect. Therefore, according to the above reports, the appropriate concentration of nanoparticles to influence plant growth indices varies between species. The positive or negative effects of nanoparticles on plants depend on the type of nanoparticles, their concentration, method, and duration of application, and the plant species76.
In this study, the effect of synthesized AgNPs on the levels of nitrogen, phosphorus, and potassium elements in geranium leaves was also investigated. The most significant effect was observed at a soil capacity of 60% with 10% and 20% nanoparticle concentrations. Under stress conditions, nitrogen content significantly decreased, and spraying with 10% AgNPs increased nitrogen, potassium, and phosphorus under drought stress conditions. Additionally, the lowest phosphorus content was observed in the control plant and at a 10% nanoparticle concentration under 90% field capacity, while the lowest potassium content was found at a 30% nanoparticle concentration under 30% field capacity in drought stress conditions.
In medicinal plants, nitrogen plays an essential role in the development and division of new cells containing essential oils and in the biosynthesis of these oils and active compounds. Since essential oils are composed of terpenoid compounds, which are built from isoprenoid units and require NADPH and ATP for their formation, the presence of elements such as nitrogen is critical for their production. Phosphorus, as a component of RNA and DNA, also acts as a molecular bridge in phospholipid membranes. Potassium is a highly consumed cation in plants and plays a significant role in cell expansion, leaf mobility, metabolic adjustments, germination, osmotic regulation, and stomatal movements under stress.
Improving crops for drought tolerance has always faced challenges77highlighting the need for innovative methods to address this issue. Nanoparticles, at specific concentrations, may have a significant toxic effect on plants due to the production of reactive oxygen and nitrogen species78. Therefore, more studies at the cellular and molecular levels are needed to determine the negative and positive effects of nanoparticles under stress conditions78. Given that nanoparticles derived from Moringa plant extract have significant biological potential, further studies and investigation of side effects and optimization of conditions could lead to their use in the food, pharmaceutical, and agricultural industries.
The summary of the effect of foliar application of silver nanoparticles (AgNPs) synthesized by Moringa oleifera on improving the yield and quality of Pelargonium hortorum under drought stress is in Fig. 10. The analysis shows that drought stress alone significantly reduces leaf blade length, and leaf blade width, plant height, stem diameter, and flower diameter, along with a decline in nutrient (N and K) levels. However, chlorophyll content (SPAD index) and catalase (CAT) enzyme activity increase, indicating an intrinsic stress response.
In contrast, plants treated with AgNPs under drought stress demonstrate enhanced physiological and biochemical traits, including larger flower diameter, increased petal anthocyanin content, higher chlorophyll levels, and greater CAT enzyme activity. Additionally, nutrient retention (N and K) improves, suggesting better stress tolerance. These results indicate that AgNPs help mitigate drought-induced damage by boosting antioxidant activity, maintaining nutrient balance, and enhancing stress adaptation mechanisms. Thus, AgNPs hold the potential as a viable approach to improving plant drought resilience.

The summary of the effect of foliar application of AgNPs synthesized by Moringa oleifera on improving the yield and quality of Pelargonium hortorum under drought stress.
Conclusion
The findings of this study demonstrated that geranium plants are sensitive to drought stress, and both quantitative and qualitative plant traits are disrupted under such conditions. However, foliar spraying with AgNPs at a concentration of 10% significantly increased the leaf length and width, plant height, number of flowering branches, flower diameter, catalase enzyme activity, anthocyanin content in petals, chlorophyll content, and the levels of nitrogen, potassium, and phosphorus in geranium leaves, while also delaying the flowering time. These results suggest that AgNPs play a pivotal role in activating the plant’s antioxidant defense system, helping to alleviate the harmful effects of drought stress. By stabilizing critical compounds like anthocyanins and chlorophyll, and improving growth parameters, AgNP treatment mitigates drought-induced damage. This approach also helps in postponing flowering, which could be beneficial under stress conditions.
In conclusion, the application of silver nanoparticles presents a promising strategy for enhancing the resilience of geranium plants to drought stress. However, further investigations are necessary to explore the underlying mechanisms of plant responses to nanoparticles and to refine their use for optimal results in diverse agricultural settings.
Future directions and research limitations
This study showed that foliar application of silver nanoparticles (AgNPs) synthesized from Moringa oleifera enhances drought tolerance and improves the growth of Pelargonium hortorum. Nevertheless, additional research is essential to refine application methods and evaluate the long-term effects of AgNPs. Large-scale field trials should validate AgNPs’ effectiveness and explore their interactions with soil microbiomes. Furthermore, it is vital to assess the environmental risks, economic feasibility, and regulatory challenges is also crucial. The study’s limitations include controlled greenhouse conditions and a single growing season. Addressing these gaps will be crucial in establishing AgNPs as a sustainable and effective strategy for enhancing crop resilience to drought stress.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding authors, Ali Salehi Sardoei and Sediqeh Afsharipour, upon reasonable request via email at alisalehisardoei1987@gmail.com and s.afshari996@gmail.com.
References
Bayat, H., Karimi, M. & Babaei, S. First report of Pelargonium flower break virus (PFBV) in Iran and determining some of its molecular properties (2021).
Bhusal, N., Han, S. G. & Yoon, T. M. Impact of drought stress on photosynthetic response, leaf water potential, and stem Sap flow in two cultivars of bi-leader Apple trees (Malus× domestica Borkh). Sci. Hortic. 246, 535–543 (2019).
Arbona, V. et al. Physiological, metabolic, and molecular responses of plants to abiotic stress. Stress Signal. Plants: Genomics Proteom. Perspective Volume. 2, 1–35 (2017).
Amirkhiz, K. F., Dehaghi, M. A., Sanavy, S. A. M. M. & Rezazadeh, A. Evaluation of changes in fatty acid profile, grain, and oil yield of Carthamus tinctorius L. in response to foliar application of polyamine compounds under deficit irrigation conditions. Ind. Crops Prod. 161, 113231 (2021).
Mahalingam, R. Consideration of combined stress: A crucial paradigm for improving multiple stress tolerance in plants. Combined stresses in plants: Physiological, molecular, and biochemical aspects 1–25 (2015).
Naghizadeh, M. et al. Exogenous application of melatonin mitigates the adverse effects of drought stress on morpho-physiological traits and secondary metabolites in Moldavian balm (Dracocephalum moldavica). Physiol. Mol. Biology Plants. 25, 881–894 (2019).
Solgi, M. & Taghizadeh, M. The effects of silver nitrate, thymol, green silver nanoparticles and Chitosan on vase life of carnation cut flowers cv. white liberty. Plant. Productions. 40, 1–12 (2017).
Nejatzadeh, F. Effect of silver nanoparticles on salt tolerance of Satureja hortensis l. during in vitro and in vivo germination tests. Heliyon 7 (2021).
Iqbal, S., Waheed, Z. & Naseem, A. Nanotechnology and abiotic stresses. Nanoagronomy 37–52 (2020).
Oldenburg, S. J. Silver nanoparticles: properties and applications. Sigma-Aldrich Co. Nd (2014).
Ngo, Q. B. et al. Effects of nanocrystalline powders (Fe, Co and Cu) on the germination, growth, crop yield and product quality of soybean (Vietnamese species DT-51). Adv. Nat. Sci. NanoSci. NanoTechnol. 5, 015016 (2014).
Mohanpuria, P., Rana, N. K. & Yadav, S. K. Biosynthesis of nanoparticles: technological concepts and future applications. J. Nanopart. Res. 10, 507–517 (2008).
Mittal, A. K., Chisti, Y. & Banerjee, U. C. Synthesis of metallic nanoparticles using plant extracts. Biotechnol. Adv. 31, 346–356 (2013).
Das, R. K. et al. Biological synthesis of metallic nanoparticles: plants, animals and microbial aspects. Nanatechnol. Environ. Eng. 2, 1–21 (2017).
Soliman, M. et al. Rapid green synthesis of silver nanoparticles from blue gum augment growth and performance of maize, fenugreek, and onion by modulating plants cellular antioxidant machinery and genes expression. Acta Physiol. Plant. 42, 1–16 (2020).
Vijayaraghavan, K., Nalini, S. P. K., Prakash, N. U. & Madhankumar, D. One step green synthesis of silver nano/microparticles using extracts of Trachyspermum ammi and Papaver somniferum. Colloids Surf. B Biointerfaces. 94, 114–117 (2012).
Tippayawat, P., Phromviyo, N., Boueroy, P. & Chompoosor, A. Green synthesis of silver nanoparticles in aloe vera plant extract prepared by a hydrothermal method and their synergistic antibacterial activity. PeerJ 4 e2589 (2016).
Mohammed, A. E., Al-Qahtani, A., Al-Mutairi, A., Al-Shamri, B. & Aabed, K. Antibacterial and cytotoxic potential of biosynthesized silver nanoparticles by some plant extracts. Nanomaterials 8, 382 (2018).
Fahey, J. W. Moringa oleifera: a review of the medical evidence for its nutritional, therapeutic, and prophylactic properties. Part 1. Trees Life J. 1, 1–15 (2005).
Foidl, N., Makkar, H. P. S. & Becker, K. The potential of Moringa oleifera for agricultural and industrial uses (2001).
Wasif Nouman, W. N. et al. Potential of Moringa oleifera L. as livestock fodder crop: A review (2014).
Rady, M. M., Varma, B. & Howladar, S. M. Common bean (Phaseolus vulgaris L.) seedlings overcome NaCl stress as a result of presoaking in Moringa oleifera leaf extract. Sci. Hortic. 162, 63–70 (2013).
Howladar, S. M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 100, 69–75 (2014).
Price, M. L. The moringa tree. ECHO Tech. Note. 17391, 1–19 (2007).
Azim, Z. et al. A review summarizing uptake, translocation and accumulation of nanoparticles within the plants: current status and future prospectus. J. Plant. Biochem. Biotechnol. 32, 211–224 (2023).
Largia, M. J. V., Shilpha, J., Satish, L., Swamy, M. K. & Ramesh, M. Elicitation: an efficient strategy for enriched production of plant secondary metabolites. Phytochemical Genomics: Plant. Metabolomics Med. Plant. Genomics 477–497 (2023).
Shahraki, H., Mahdi Nezhad, N. & Fakheri, B. A. The effect of synthesis nanosilver by plant extract on morphological and antioxidant properties of artichoke (Cynara scolymus L.) under salinity stress. Plant. Productions. 44, 103–114 (2021).
Perumalsamy, H. et al. A comprehensive review on Moringa oleifera nanoparticles: importance of polyphenols in nanoparticle synthesis, nanoparticle efficacy and their applications. J. Nanobiotechnol. 22, 71 (2024).
Rady, M. M. & Mohamed, G. F. Modulation of salt stress effects on the growth, physio-chemical attributes and yields of Phaseolus vulgaris L. plants by the combined application of Salicylic acid and Moringa oleifera leaf extract. Sci. Hortic. 193, 105–113 (2015).
Nikbakht, M. & Pour Ali, P. Biological production and antibacterial effect of synthesized ag with aqua extract and methanol Anab. Med. Sci. J. Islam Azad Uni. 12, 112–118 (2015).
Kaviani, N. & Osfoori, M. Biological Preparation of silver nanoparticles using artemisia sieberi. Modares J. Biotechnol. 9, 23–27 (2018).
Abd El-Mageed, T. A., Semida, W. M. & Rady, M. M. Moringa leaf extract as biostimulant improves water use efficiency, physio-biochemical attributes of squash plants under deficit irrigation. Agric. Water Manag. 193, 46–54 (2017).
Khorasaninejad, S., Alizadeh Ahmadabadi, A. & Hemmati, K. The effect of humic acid on leaf morphophysiological and phytochemical properties of Echinacea purpurea L. under water deficit stress. Sci. Hortic. 239, 314–323 (2018).
Soroori, S., Danaee, E., Hemmati, K. & Ladan Moghadam, A. Effect of spermidine foliar application on some morphophysiological traits and secondary metabolites of marigold (Calendula officinalis L.) under drought stress. J. Plant. Environ. Physiol. 17, 108–125 (2022).
Rahbarian, P. & Salehi, S. A. L. I. Effect waste of palm trees and sand and perlite mixed with some growth indices Ficus Benjamina (2014).
Sardoei, A. S., Roien, A., Sadeghi, T., Shahadadi, F. & Mokhtari, T. S. Effect of vermicompost on the growth and flowering of African marigold (Tagetes erecta). Am. Eurasian J. Agric. Environ. Sci. 14, 631–635 (2014).
Wagner, G. J. Content and vacuole/extravacuole distribution of neutral sugars, free amino acids, and anthocyanin in protoplasts. Plant. Physiol. 64, 88–93 (1979).
Claiborne, A. Handbook of Methods for Oxygen Radical Research 283–284 (CRC Press, 1985).
Abdel-Shafy, H. I., Hegemann, W. & Teiner, A. Accumulation of metals by vascular plants. Environ. Manage. Health. 5, 21–24 (1994).
Yazdanpanah, A. & Motallebifard, R. The effects of chicken manure and potassium fertilizer on yield and nitrogen, phosphorus, potassium, zinc and copper uptake of potato. Appl. Soil. Res. 4, 60–71 (2017).
Afsharipour, S., Mirzaalian Dastjerdi, A. & Seyedi, A. Optimizing Cucumis sativus seedling vigor: the role of pistachio wood vinegar and date palm compost in nutrient mobilization. BMC Plant. Biol. 24, 407 (2024).
Li, W. et al. Impacts of drought stress on the morphology, physiology, and sugar content of Lanzhou Lily (Lilium Davidii var. unicolor). Acta Physiol. Plant. 42, 1–11 (2020).
Yang, X. et al. Response mechanism of plants to drought stress. Horticulturae 7, 50 (2021).
Peivandi, M., Jamkani, Z. & Mirza, M. Effect of iron nano-chelate with iron chelate on growth and antioxidant enzymes activity of Satureja hortensis. J. New. Cell. Mol. Biotechnol. 2, 25–32 (2011).
Tiwari, D. K. et al. Interfacing carbon nanotubes (CNT) with plants: enhancement of growth, water and ionic nutrient uptake in maize (Zea mays) and implications for nanoagriculture. Appl. Nanosci. 4, 577–591 (2014).
Taiz, L., Zeiger, E., Møller, I. M. & Murphy, A. Plant Physiology and Development (2015).
Aslani, F. et al. Effects of engineered nanomaterials on plants growth: an overview. Sci. World J. 2014, 641759 (2014).
Choi, O. et al. The inhibitory effects of silver nanoparticles, silver ions, and silver chloride colloids on microbial growth. Water Res. 42, 3066–3074 (2008).
Salehi, M. & Tamaskani, F. Pretreatment effect of nanosilver on germination and seedling growth of wheat under salt stress. in Proceeding of First Iranian Congress in Seed Sciences and Technology. Gorgan, Iran (2008).
Adedapo, A. A., Falayi, O. O. & Oyagbemi, A. A. Evaluation of the analgesic, anti-inflammatory, anti-oxidant, phytochemical and toxicological properties of the methanolic leaf extract of commercially processed Moringa oleifera in some laboratory animals. J. Basic. Clin. Physiol. Pharmacol. 26, 491–499 (2015).
Brunner, T. J. et al. In vitro cytotoxicity of oxide nanoparticles: comparison to asbestos, silica, and the effect of particle solubility. Environ. Sci. Technol. 40, 4374–4381 (2006).
Ahmad, N. et al. Rapid synthesis of silver nanoparticles using dried medicinal plant of Basil. Colloids Surf. B 81, 81–86 (2010).
Reddy, G. R. & Gandhi, N. N. Environmental friendly biosynthesis, characterization and antibacterial activity of silver nanoparticles by using Senna Saimea plant leaf aqueous extract. Int. J. Iins Pharm. Life Sci. 2, 186–193 (2012).
Roopan, S. M. et al. Low-cost and eco-friendly phyto-synthesis of silver nanoparticles using Cocos nucifera Coir extract and its larvicidal activity. Ind. Crops Prod. 43, 631–635 (2013).
Fotoohiyan, Z., Karimdadi, M., Salehi Sardoei, A., Bovand, F. & Ghorbanpour, M. Green synthesis of silver nanoparticles using Artemisia persica extract and evaluation of their activity on onion Gray rot fungus. Biomass Convers. Biorefin. 1–13 (2024).
Shams, G., Ranjbar, M. & Amiri, A. Effect of silver nanoparticles on concentration of silver heavy element and growth indexes in cucumber (Cucumis sativus L. negeen). J. Nanopart. Res. 15, 1–12 (2013).
Salama, H. M. H. Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea mays L). Int. Res. J. Biotechnol. 3, 190–197 (2012).
Sharma, K., Sharma, R., Shit, S. & Gupta, S. Nanotechnological application on diagnosis of a plant disease. in International conference on advances in biological and medical sciences, Singapore 15–16 (2012).
Van Nguyen, D. et al. Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant. Growth Regul. 41, 364–375 (2022).
Sourour, A., Afef, O., Mounir, R. & Mongi, B. Y. A review: morphological, physiological, biochemical and molecular plant responses to water deficit stress. Int. J. Eng. Sci. 6, 1805–2319 (2017).
Baghbani-Arani, A., Modarres-Sanavy, S. A. M., Mashhadi-Akbar-Boojar, M. & Mokhtassi-Bidgoli, A. Towards improving the agronomic performance, chlorophyll fluorescence parameters and pigments in Fenugreek using zeolite and vermicompost under deficit water stress. Ind. Crops Prod. 109, 346–357 (2017).
Shekofteh, H. & Dehghani Fatehabad, R. Effect of water stress and potassium on yield and yield components of Cumin (Cuminum cyminum L.) (2016).
Inze, D. & Van Montagu, M. Oxidative stress in plants. Curr. Opin. Biotechnol. 6, 153–158 (1995).
Sabertanha, B., Fakheri, B., Mahdinezhad, N. & Alizade, Z. Effects of silver nanoparticles elicitor and drought stress on the expression of beta-carotene hydroxylase (bch) gene on the yield of saffron carotenoid (Crocus sativus L). Crop Biotechnol. 6, 1–13 (2017).
Han, Y. et al. Important changes in germination, seedling tolerance, and active components content due to drought stress on three licorice (Glycyrrhiza) species. Ind. Crops Prod. 175, 114240 (2022).
Bagherzadeh Homaee, M. & Ehsanpour, A. A. Physiological and biochemical responses of potato (Solanum tuberosum) to silver nanoparticles and silver nitrate treatments under in vitro conditions. Indian J. Plant. Physiol. 20, 353–359 (2015).
Jiang, H., Li, M., Chang, F., Li, W. & Yin, L. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 31, 1880–1886 (2012).
Ma, C. et al. Physiological and molecular response of Arabidopsis thaliana (L.) to nanoparticle cerium and indium oxide exposure. ACS Sustain. Chem. Eng. 1, 768–778 (2013).
Oukarroum, A., Barhoumi, L., Pirastru, L. & Dewez, D. Silver nanoparticle toxicity effect on growth and cellular viability of the aquatic plant Lemna gibba. Environ. Toxicol. Chem. 32, 902–907 (2013).
Amooaghaie, R., Tabatabaei, F. & Ahadi, A. Alterations in HO-1 expression, Heme Oxygenase activity and endogenous NO homeostasis modulate antioxidant responses of Brassica nigra against nano silver toxicity. J. Plant. Physiol. 228, 75–84 (2018).
Khosroushahi, A. Y. et al. Improved taxol production by combination of inducing factors in suspension cell culture of Taxus baccata. Cell. Biol. Int. 30, 262–269 (2006).
Guo, N. et al. Anthocyanin biosynthetic genes in Brassica rapa. BMC Genom. 15, 1–11 (2014).
Qian, H. et al. Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana. J. Environ. Sci. 25, 1947–1956 (2013).
Kafi, M. Effects of chilling stress on physiological and biochemical traits of three hybrid of corn (Zea mays L.) in seedling stage. J. Plant. Res. (Iranian J. Biology). 29, 540–552 (2016).
Najafi, S., Heidari, R. & Jamei, R. Influence of silver nanoparticles and magnetic field on phytochemical, antioxidant activity compounds and physiological factors of Phaseolus vulgaris. Tech. J. Eng. Appl. Sci. 3, 2812–2816 (2013).
Ma, C. et al. Reduced silver nanoparticle phytotoxicity in Crambe abyssinica with enhanced glutathione production by overexpressing bacterial γ-glutamylcysteine synthase. Environ. Sci. Technol. 49, 10117–10126 (2015).
Karimi Afshar, A., Baghizadeh, A. & Mohammadi-Nejad, G. Screening of Iranian Cumin (Cuminum cyminum L.) ecotypes under normal moisture and drought conditions using tolerance indices. J. Ethno-Pharmaceutical Prod. 2, 17–22 (2021).
Banan, A. et al. Salinity modulates biochemical and histopathological changes caused by silver nanoparticles in juvenile Persian sturgeon (Acipenser persicus). Environ. Sci. Pollut. Res. 27, 10658–10671 (2020).
Author information
Authors and Affiliations
Contributions
“In this collaborative research, M.B. led the research team, proposed the idea, and provided the plants. P.Z. was responsible for conducting the greenhouse experiments, while A.S.S focused on data analysis and interpretation. Z.F. contributed significantly to writing and editing the manuscript, ensuring its scientific accuracy and consistency. A.Gh. collected key data and drafted the initial version of the manuscript. A.D. supplied the necessary materials and equipment. S.A. conducted laboratory work and contributed to writing and editing. All authors reviewed the final manuscript and approved its publication, confirming their agreement with the study’s content and findings.”
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Babarabie, M., Zolfaghary, P., Sardoei, A.S. et al. The effect of foliar application of silver nanoparticles synthesized by Moringa oleifera on improving the yield and quality of Pelargonium hortorum under drought stress. Sci Rep 15, 23239 (2025). https://doi.org/10.1038/s41598-025-05457-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-05457-4
