
Abstract
Scientific knowledge of vertebrate scavenging communities has increased over the last decades. However, there is a lack of information on the Neotropics. We aimed to describe the vertebrate scavenger community of the tropical dry forest of western Ecuador and to identify the factors influencing carcass consumption patterns by this guild. Between November 2022 and May 2023, we placed 64 experimental carcasses of different sizes in eight camera-trapping stations (32 carcasses for each category). At each station, we simultaneously placed two carcasses at least 100 m apart: one small-sized (chicken) and one medium-sized (goats or pigs) carcass. We fitted Generalized Linear Mixed Models to assess the effects of carcass size and season (rainy and dry) on the scavenger species richness and abundance, considering all scavenger species and by species group. We also fitted Linear Mixed Models to assess differences in carcass detection time, total consumption time and rate, using the carcass size, vulture abundance, and percentage of canopy cover as predictors. The scavenger community was dominated by obligate scavengers (i.e., vultures), and the species detected were the Black Vulture (Coragyps atratus), the Turkey Vulture (Cathartes aura), and the King Vulture (Sarcoramphus papa). Facultative mammalian scavengers such as Ocelot (Leopardus pardalis), Tayra (Eira barbara), Crab-eating Raccoon (Procyon cancrivorus) and domestic dog (Canis lupus familiaris) were recorded in a smaller number of carcasses. No facultative avian scavenger species were detected. Carcass size influenced all the variables analyzed, except the scavenger species richness. While the season influenced the vulture abundance, especially of S. papa. Facultative scavenger richness and abundance did not show significant differences. Carcass total consumption time was not explained by any of the variables included in the models. Both carcass detection time and consumption rates were influenced by carcass size, particularly in the case of consumption rates, where larger carcasses exhibited higher consumption rates than smaller ones. The dominance of vulture species in the scavenger community suggests that their ecological role is indispensable for the dry forest of western Ecuador. The loss of obligate scavengers could have important implications for ecosystem services and human well-being.
Similar content being viewed by others

Introduction
Vertebrate scavengers play an essential role in ecosystems by consuming dead animals, which accelerates the process of organic matter decomposition and nutrient recycling1,2,3. Two functional groups use carrion as a food resource. Obligate scavengers (i.e., vultures) are species that feed almost exclusively on carrion, while facultative scavengers (e.g., mammalian carnivores, avian species like raptors and corvids) often, but not always, feed on carrion3,4,5. Vultures help to eliminate potential foci of disease transmission due to their gregarious habits and ability to consume large amounts of biomass in relatively short periods3,6. Despite this, anthropogenic threats such as poisoning with pesticides or rodenticides, lead contamination, human-wildlife conflict, and illegal hunting make them one of the most threatened groups of birds worldwide4,7,8,9,10.
Carcass size is one of the main factors influencing their detection and consumption patterns11,12. Species richness and abundance in vertebrate scavenger communities often vary depending on carcass size and biomass availability11,12,13. Likewise, carcass consumption patterns (e.g. detection time, carcass consumption time and rate) varies due to climate-related factors, such as temperature or precipitation14,15. Also, it has been demonstrated that the type of landscape where the carcasses appear is a factor influencing the time taken to detect and consume them16. However, there is a notable geographical bias in the amount of information about the influence of climatic factors and landscape on scavenger communities, as available studies have mainly been conducted in septentrional countries12,17,18. Although there are no distinct seasons per se in the Neotropics, some regions exhibit cyclical rainfall patterns that mark two seasons19; therefore, it is foreseeable that these dynamics may influence the scavenger communities. In globally threatened ecosystems such as tropical dry forests, which are highly dependent on their rainfall patterns, it is essential to assess the influence of carcass size, rainy and dry seasons on the composition and dynamics of vertebrate scavenger communities11,20,21.
Interactions among scavenging species can facilitate food detection and reduce carcass consumption time22,23. For example, some vulture species play an essential role in maximizing the foraging success of other vulture species (i.e., heterospecifics) by providing visual cues to food location and opening up carcasses, enabling access to food22,23,24. To prevent agonistic encounters, some scavengers avoid using the resource in the presence of species that may exhibit aggressive behaviors, such as domestic dogs (Canis lupus familiaris)25,26. The lack of information on carrion use, foraging behavior, and the richness and abundance of vertebrate scavengers in tropical dry forests makes it difficult to design and implement effective conservation strategies (e.g. multi-species action plans) for species with declining populations, such as some New World vultures8,21,27,28,29,30. Given the vulnerability of the tropical dry forest, which has historically lost 75% of its original extent21,31,32scientific research on vertebrate scavenger communities in this critically threatened ecosystem is a priority.
A comprehensive understanding of climate-related and landscape factors that influence vertebrate scavenger communities and their dynamics, is essential to developing conservation measures and maintaining the ecosystem services2,6,16. By consuming carcasses and maintaining healthy ecosystems, vulture species are essential for biodiversity conservation and achieving sustainable development goals33,34. We aimed to describe the scavenging guild exploiting carrion and to evaluate the influence of carcass size and season on the vertebrate scavenger community (species richness and abundances) and carcass consumption patterns (detection time, total consumption time, consumption rate) in the tropical dry forest of western Ecuador. We predicted that (1) larger carcasses (medium-sized carcasses in the experimental system), as a resource with greater available biomass, would have a higher number of scavenger species and a higher abundance, that (2) scavenger abundance would differ between the rainy and dry seasons, that (3) carcass consumption time and detection time would be lower on medium-sized carcasses and during the dry season due to lower canopy cover, and that (4) carcass consumption rate would be higher on medium-sized carcasses11,35.
Methods
Study area
The study was conducted in three private nature reserves located in the dry forest of western Ecuador. The Lalo Loor Reserve and the Jama-Coaque Reserve are located in Manabí Province, while the Bosque Protector Cerro Blanco is situated in Guayas Province (Fig. 1). The Lalo Loor Reserve has 180 ha and is managed by the Ceiba Foundation36. The Jama-Coaque Reserve has 840 ha belonging to the Third Millennium Alliance37and the Bosque Protector Cerro Blanco has an area of 6,078 ha and is managed by the Pro-Bosque Foundation38.
The Manabí and Guayas reserves are distanced from each other by approximately 200 km but share the same climatic and environmental characteristics. The altitude of the reserves ranges from 80 to 500 m above sea level. The study area has a rainy season, between January and May, characterized by rainfall ranging from 500 to 1000 mm, and a dry season from June to December, with precipitation below 500 mm. The average annual temperature is 26° C, with a 1 to 2° C decrease during the dry season39,40.
The landscape at the three sites consists of remnants of dry semi-deciduous forest, urbanized areas, and agricultural land. The vegetation is xerophytic and deciduous, dominated by shrub species. The tree species reach a canopy height of 10 to 25 m. During the dry season, between 25 and 75% of the tree and shrub species lose their leaves, and the herbaceous vegetation disappears. The representative plant species of the area are Ceiba trischistandra, Cavanillesia platanifolia, and Cecropia angustifolia39,40.
The three reserves are part of the Tumbes-Chocó-Magdalena hotspot. A total of 543 vertebrate species have been recorded in the area surrounding the reserves, including three species of obligate scavengers: the Black Vulture (Coragyps atratus), the Turkey Vulture (Catharthes aura), and the King Vulture (Sarcoramphus papa). There is no information or estimate of the vulture population size in the study area. However, there are informal records of breeding pairs and chicks of both the Turkey and the King Vultures. The most common large herbivores are the Collared Peccary (Dicotyles tajacu) and deer of the Mazama genus. Native carnivores include the Sechura Fox (Lycalopex sechurae), the Tayra (Eira barbara), and the Ocelot (Leopardus pardalis)39,41. In the study area, natural carcasses consist mainly of medium-sized wild herbivores. However, it is possible to find larger carcasses with less frequency of domesticated introduced species such as cattle (Bos taurus), horses (Equus caballus), or donkeys (Equus asinus) that occasionally die inside or near the reserves.

Location of the three study sites in the dry forest of western Ecuador and the camera-trapping stations within each study area. The map was created using QGIS 3.28 software (URL: https://www.qgis.org/).
Data collection
Between November 2022 and May 2023, we systematically monitored 64 experimental carcasses at eight camera-trapping stations, which were placed in the Cerro Blanco Protected Forest (4 stations), the Jama-Coaque Reserve (3 stations), and the Lalo Loor Reserve (1 station) (Fig. 1). The camera-trapping stations were separated from each other by at least 1 km. At each camera-trapping station, we simultaneously placed two carcasses of different sizes at least 100 m apart: a small-sized carcass weighing between 1.5 and 4 kg of domestic chickens (Gallus gallus), and a medium-sized carcass weighing between 15 and 40 kg of domestic pigs (Sus scrofa var. dom) or domestic goats (Capra hircus).
We placed a total of four sets of carcasses. At each camera-trapping station, we placed carcasses on two occasions in the dry season (November and December) and on another two occasions in the rainy season (March and April). We did not place carcasses in January and February to avoid the transition period between the rainy and dry seasons. Carcasses placed at the same camera-trap station were approximately 30 days apart from the previous one. These were monitored from the moment they were placed until their biomass was consumed entirely, leaving only skin, bones, fur, or feathers42.
We used one camera trap for each carcass placed in the field. The camera traps were placed 50 cm above the ground and 6–7 m from the carcasses. We set the camera traps to hybrid mode (capturing two photos and a 20-second video) with a 30-second delay and a “Normal” sensor level. The time elapsed between each of the two photos and the video was of one second by camera default. The use of videos made it possible to clearly verify the number of individuals and their stages in case the photos were not clear enough. We checked the memory cards and batteries every 15 days after placing the carcasses.
At each camera-trapping station, we recorded the time and date of carcass placement, the weight of the carcass (in kg), and the time and date of retrieval from the camera trap. We estimated canopy cover at the time of carcass placement by taking a photo pointing skyward and determining the percentage of the photo that was covered by the tree canopy. Then, we obtained the following data on the scavenger community from the analyzed photos and videos: total scavenger species richness, vulture species richness, facultative scavenger species richness, abundance of each species, vulture abundance, and mammalian scavenger abundance. Abundance was calculated as the highest number of individuals of the same species recorded in the same photo or photo-sequence. It was estimated by recognizing individuals by their plumage or fur patterns, age, and sex11especially for S. papa, C. aura, L. pardalis, and E. barbara. The abundance of vultures and carnivorous mammals was estimated as the sum of the number of individuals of each species for each group in the same carcass.
We estimated three variables related to carcass consumption. (1) Detection time was the time in hours it took the scavenger species to locate the carcass. (2) Total consumption time, which was the time in hours when the carcass was consumed entirely since it was placed in the field. Meanwhile, (3) the consumption rate was estimated based on the relationship between carcass weight (kg) and total consumption time (h)11,12,16.
Data analysis
We analyzed 62 carcasses because one camera trap was stolen, and another had a damaged SD card. We constructed Generalized Linear Mixed Models (GLMMs) to assess differences in scavenger species richness and abundance, using the total scavenger species richness, vulture species richness, total carnivore mammal species richness, species abundance, vulture abundance, and mammal abundance as response variables. Carcass size and season were used as predictor variables. Camera-trapping stations were treated as a random factor to control for the lack of independence of carcasses placed simultaneously at the same camera-trapping station. Negative binomial family models were fitted to address overdispersion43. The R package “lme4” fit the mixed models44.
The variables of carcass detection time, total consumption time, and consumption rate were normalized by the natural logarithm12 and selected as response variables to fit Linear Mixed Models (LMMs). The predictor variables were carcass size, vulture abundance, and percentage of canopy cover. Variance inflation factor (VIF) analysis was used to measure the multicollinearity of model variables, and variables with values greater than five were excluded45. No interactions between any of the model terms were included.
We performed the model selection based on the Akaike information criterion (AICc). Only models with a ΔAICc < 2 were considered valid46. If we obtained more than one candidate model, we performed model averaging using the package “MuMIn”47. We used the R package “MuMIn”47 to calculated the R2 marginal (R2m) to evaluate the proportion of variance explained by the fixed effects alone, and R2 conditional (R2c) to see the proportion of variance explained by the fixed and the random effects. All tests were performed using R software version 4.4.148. We considered the results to be statistically significant when p < 0.05.
Results
Scavenger community
We detected seven species of vertebrate scavengers, including three obligate scavengers: the Black Vulture (Coragyps atratus), the Turkey Vulture (Cathartes aura) and the King Vulture (Sarcoramphus papa), and four species of mammalian facultative scavengers: the Ocelot (Leopardus pardalis), the Tayra (Eira barbara), the Crab-eating Raccoon (Procyon cancrivorus) and the domestic dog (C. l. familiaris) (Fig. 2). No facultative avian scavenger species were detected.
The species that most frequently visited the carcasses were C. aura (87.1%) and C. atratus (85.5%), followed by S. papa (48.4%), L. pardalis (25.8%), and E. barbara (11.3%). While the species that visited the carcasses least frequently were C. l. familiaris (3.2%) and P. cancrivorus (1.6%). We detected that C. aura and C. atratus visited 100% of medium-sized carcasses and between 69% and 80% of small carcasses in both the dry and rainy seasons, and S. papa visited mainly medium-sized carcasses in the dry season (86.7%) and the rainy season (62.5%). In contrast, C. l. familiaris and P. cancrivorus were only recorded in small carcasses during the dry season, and E. barbara was not recorded in medium-sized carcasses during the rainy season (Fig. 3).
Vultures were the most abundant species, with groups of 15.66 ± 11.15 (mean ± SD) individuals. Coragyps atratus was the most abundant species, recorded with a mean abundance of 10.48 ± 9.75 and a maximum of 49 individuals. It was followed by C. aura and S. papa with mean abundances of 3.35 ± 2.14 and 1.82 ± 2.85 individuals, respectively. Cathartes aura was recorded in groups of up to nine individuals, while S. papa was recorded in groups with a maximum of 11 individuals. Facultative mammalian scavengers were less abundant, with a mean of less than one individual per carcass. L. pardalis had the highest mean in this group, with 0.34 ± 0.62 individuals (Supplementary Table 1).

Native vertebrate scavenger species recorded in the dry forest of western Ecuador. (A) Black Vulture (Coragyps atratus), (B) Turkey Vulture (Cathartes aura), (C) King Vulture (Sarcoramphus papa), (D) Ocelot (Leopardus pardalis), (E) Tayra (Eira barbara), (F) Crab-eating Raccoon (Procyon cancrivorus).

Percentage of small-sized and medium-sized carcasses visited by seven species of vertebrate scavengers during the rainy and dry season in the dry forest of western Ecuador.
Carcass consumption patterns
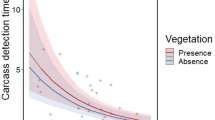
The most frequent detector species was C. aura, which detected 54.84% and 58.06% of small and medium-sized carcasses, respectively. Leopardus pardalis detected 22.58% of small-sized carcasses, while C. atratus detected 22.58% of medium-sized carcasses. Mean (± SD) carcass detection time was 20.16 ± 18.99 h. Small-sized carcasses were detected in a mean time of 25.07 ± 23.82 h, and medium-sized carcasses were detected in a mean time of 15.25 ± 10.78 h (Supplementary Table 1).
Mean (± SD) carcass total consumption time was 47.64 ± 31.73 h. In medium-sized carcasses, the mean time of total consumption was 40.87 ± 21.37 h; in small-sized carcasses, the mean time of total consumption increased to 54.40 ± 38.67 h. The consumption period of small-sized carcasses varied between 3.73 and 149.13 h (Supplementary Table 1).
Mean (± SD) carcass consumption rate was 0.47 ± 0.75 kg/h. The mean consumption rate was higher in medium-sized carcasses (0.86 ± 0.91 kg/h) than in small-sized carcasses (0.08 ± 0.11 kg/h).
Influence of carcass size and season on scavenging dynamics
Species richness and abundance of facultative scavengers did not show significant differences (p-value > 0.05) influenced by the carcass size or season (Supplementary Table 2). The model-averaged indicated that carcass size influenced all the variables analyzed, except the scavenger species richness. While the season influenced the vulture abundance and the abundance of S. papa. The camera-trapping station (random effect) highly increased the variance explained for the variables: vulture abundance, the abundance of C. atratus and S. papa (Table 1). Medium-sized carcasses had higher scavenger species richness, higher vulture species richness and abundance than small-sized carcasses. During the dry season, both small and medium-sized carcasses recorded higher scavenger species richness and higher vulture abundance compared to the rainy season (Fig. 4).

Boxplot of the differences in the variables of the scavenger community studied in the tropical dry forest of Ecuador according to carcass size and season (rainy vs. dry).
Carcass total consumption time was not explained by any of the variables included in the LMMs. In contrast, Carcass detection time was influenced by carcass size, with a low R2 marginal value, explaining low variability in the model. Also, carcass size had a significant influence on the consumption rate. The camera-trapping station (random effect) increased the variance explained for the carcass detection time and consumption rate (Table 2). Larger carcasses showed a higher consumption rate compared to smaller ones.
Discussion
We found that the vertebrate scavenger assemblage in the dry forest of western Ecuador was dominated by the obligate scavengers (i.e., vultures), with three vulture species recorded scavenging on experimental carcasses: C. atratus, C. aura, and S. papa. Facultative scavengers were recorded to a lesser extent, only involving mammalian carnivore species: L. pardalis, E. barbara, P. cancrivorus, and C. l. familiaris (Fig. 3). Our results indicate that the carcass size influenced all the variables analyzed, except the scavenger species richness. The season influenced the abundance of vultures recorded on carcasses, especially that of S. papa (Fig. 4). Furthermore, the total consumption time of carcasses was not explained by any of the variables in the model. Still, the carcass size influenced the detection time and consumption rate.
Our findings suggest that in the Neotropical dry forest, vultures may be the dominant species on carcasses (Fig. 3). Similarly, in the Brazilian Cerrado dry forest, the scavenger community was dominated by vultures, such as S. papa, C. aura, and C. atratus11. This probably occurs because the mammalian facultative scavengers in the Neotropics tend to be medium-sized species (e.g., L. pardalis, E. barbara). In contrast, in African savannas and European temperate forests, the facultative scavengers tend to be larger mammalian carnivores such as lions (Panthera leo), spotted hyenas (Crocuta crocuta), and bears (Ursus arctos), which may be more dominant and monopolize carcasses35,49,50. The greater dominance of vultures on carrion may also be due to their efficiency in locating carcasses (via visual and olfactory abilities) compared to mammals (only via olfactory abilities)22,23,51.
Contrary to our predictions, the total scavenger species richness in the tropical dry forest was not significantly influenced by carcass size or season. This is opposite to studies conducted in other tropical ecosystems where medium and large carcasses recorded more scavenging species than the smaller ones11,35. This pattern is repeated in various ecosystems worldwide, where larger carcasses recorded scavenger communities composed of more species2,14,16. However, vulture abundance was influenced by carcass size. The larger carcasses have more biomass for consumption, allowing more animals to feed on them2,35.
The absence of facultative avian scavenger species is striking despite the presence of raptor species that could potentially feed on them, such as the Red-throated Caracara (Ibycter americanus) and the Crested Caracara (Caracara plancus)52. This could be because vultures dominate carcasses and exploit them faster than other birds35,53. Likely, the high abundance of vultures in the carcasses, which are larger than other raptors, scares away facultative avian scavengers that prefer to use other resources available in the forest.
The influence of the camera-trapping station on the abundance of S. papa, may be related to the type of landscape in which the carcass appears19,53. S. papa seems to have a more specialized foraging behavior, restricted to the carcasses available within or near natural landscapes5. This reliance on the dry forest could make them more dependent on medium-sized carcasses and more vulnerable to reductions in natural food sources. On the other hand, C. atratus and C. aura have a flexible and opportunistic diet, frequently feeding on fish scraps along beaches, roadkill animals, and garbage dumps in cities54,55,56. These urban and suburban areas provide a stable food source for their diet5, and their visits to the forest in our study area may be more related to searching for additional resources. This may explain the higher percentage of visits to small-sized carcasses compared to S. papa (Fig. 3).
The rainy and dry seasons affected the vulture abundance, as well as that for C. atratus and S. papa (Fig. 4). This could be due to variations in canopy cover since the dry forest is an ecosystem characterized by semi-deciduous vegetation21,40. The abundance of the two vulture species was higher in the dry season. The lower canopy covers likely facilitated the detection of carcasses by visual cues in species that rely primarily on their sense of sight, such as C. atratus and S. papa51.
The presence of the detector species C. aura and L. pardalis, which rely mainly on their sense of smell to locate food5,22would explain why the detection time was not influenced by the season or carcass size. Likewise, changes in canopy cover would not be significant as they do not directly affect the ability to detect carcasses. Species with a developed sense of smell play an important role in ecological communities, as their presence alerts other scavenging species to the existence of available food, favoring the use of this resource22,51.
In the three protected areas studied, natural carcasses consist mainly of the remains of ungulate herbivores’ corpses, such as D. tajacu and Mazama sp39,41. As well as introduced domestic species such as C. hircus, S. scrofa var. dom., and cattle (Bos taurus) that occasionally die inside or near forested areas. The availability of carcasses in natural conditions, especially the larger ones, would depend on the presence of predators and their successful hunting events35,57. It is of great concern that no large felids (i.e., puma Puma concolor, jaguar Panthera onca) have been recorded in the last decade in the three protected areas58,which would probably reduce the possibility of finding carcasses from predation events. Specific seasonal factors could also influence the abundance and availability of food resources. However, data related to climatic factors, such as temperature and rainfall, were not directly measured, which could provide a better explanation of their influence on the assemblage and consumption patterns of vertebrate scavengers.
Implications for conservation
The high abundance of vultures could explain the low species richness of mammalian carnivores found and the absence of avian facultative scavengers in the dry forest of western Ecuador, as obligate scavengers are more efficient at detecting and consuming carrion16,22,23. We detected mixed groups of vultures up to 51 individuals, which could have restricted the use of carcasses by mammals57,59. This suggests that in the critically endangered tropical dry forest20obligate scavengers are probably fulfilling an essential role in removing organic matter and potential sources of infection for humans, which could hardly be covered by facultative scavengers alone. The loss of vultures could have important implications for ecosystem services and human well-being2,6.
Our findings provide evidence of the significance of carcass size for vulture abundance and consumption rates, as well as the importance of rainy and dry seasons for the total abundance of vultures and each vulture species. Meanwhile, the lack of significant explanatory variables for the total consumption time indicates the need for further studies on scavenger communities and their consumption patterns in tropical dry forests. It is necessary to prioritize research efforts on understudied species with globally declining populations, such as S. papa, which is listed as Near Threatened in Ecuador8,11,60,61.
We also highlight the record of domestic dogs on carcasses in the Ecuadorian western dry forest. These alien species compete for food with native scavengers26,62 and are considered a major threat to vultures in Ecuador, either through direct attacks on individuals62,63 by causing fake news and conflicts with rural inhabitants, which leads in turn to unintentional subsequent poisoning of obligate scavengers7,33,64,65. Expanding the study area to obtain a more representative sample size and achieve a comprehensive understanding of interspecific interactions in scavenger communities, particularly those involving vulture species, is essential22,23,24. It is also necessary to estimate the populations of vulture species in order to evaluate their changes over time2,8,66as well as to understand human-vulture interactions67,68. This will facilitate the design and implementation of multi-species conservation plans suited to the particular social-ecological contexts of critically endangered neotropical ecosystems.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Blázquez, M., Sánchez-Zapata, J. A., Botella, F., Carrete, M. & Eguía, S. Spatio-temporal segregation of facultative avian scavengers at ungulate carcasses. Acta Oecol. 35, 645–650. https://doi.org/10.1016/j.actao.2009.06.002 (2009).
Sebastián-González, E. et al. Scavenging in the anthropocene: human impact drives vertebrate scavenger species richness at a global scale. Glob Chang. Biol. 25, 3005–3017. https://doi.org/10.1111/gcb.14708 (2019).
Newsome, T. M. et al. Monitoring the dead as an ecosystem indicator. Ecol. Evol. 11, 5844–5856. https://doi.org/10.1002/ece3.7542 (2021).
Ogada, D., Torchin, M., Kinnaird, M. & Ezenwa, V. Effects of Vulture declines on facultative scavengers and potential implications for mammalian disease transmission. Conserv. Biol. 26, 453–460. https://doi.org/10.1111/j.1523-1739.2012.01827.x (2012).
Bildstein, K. L. Vultures of the World: Essential Ecology and Conservation (Cornell University Press, 2022).
DeVault, T. L. et al. Ecosystem services provided by avian scavengers. In Why Birds Matter: Avian Ecological Function and Ecosystem Services (eds Şekercioğlu, C. H., Wenny, D., Whelan et al.) 235-270 (University of Chicago Press, 2016).
Restrepo-Cardona, J. S. et al. Anthropogenic threats to the vulnerable Andean Condor in Northern South America. PLoS One. 17, e0278331. https://doi.org/10.1371/journal.pone.0278331 (2022).
Santangeli, A., Buechley, E. R., Mammola, S. & Lambertucci, S. A. Priorities for research and action to prevent a new world Vulture crisis. Biol. Conserv. 270, 109563. https://doi.org/10.1016/j.biocon.2022.109563 (2022).
Oliva-Vidal, P. et al. Second-generation anticoagulant rodenticides in the blood of obligate and facultative European avian scavengers. Environ. Pollut. 315, 120385. https://doi.org/10.1016/j.envpol.2022.120385 (2022).
Oliva-Vidal, P. et al. Griffon vultures, livestock and farmers: unraveling a complex socio-economic ecological conflict from a conservation perspective. Biol. Conserv. 272, 109664. https://doi.org/10.1016/j.biocon.2022.109664 (2022).
Naves-Alegre, L. et al. Uncovering the vertebrate scavenger guild composition and functioning in the Cerrado biodiversity hotspot. Biotropica 53, 1582–1593. https://doi.org/10.1111/BTP.13006 (2021).
Orihuela-Torres, A., Pérez-García, J. M., Sánchez-Zapata, J. A., Botella, F. & Sebastián-González, E. Scavenger guild and consumption patterns of an invasive alien fish species in a mediterranean wetland. Ecol. Evol. 12, e9133. https://doi.org/10.1002/ece3.9133 (2022).
van Overveld, T. et al. Integrating Vulture social behavior into conservation practice. Ornithol. Appl. 122, 1–20. https://doi.org/10.1093/condor/duaa035 (2020).
Walker, M. et al. Factors influencing scavenger guilds and scavenging efficiency in Southwestern Montana. Sci. Rep. 11, 4254. https://doi.org/10.1038/s41598-021-83426-3 (2021).
Vandersteen, J. et al. Carcass use by mesoscavengers drives seasonal shifts in Australian alpine scavenging dynamics. Wildl. Res. 50, 1031–1045. https://doi.org/10.1071/WR22100 (2023).
Oliva-Vidal, P., Sebastián-González, E. & Margalida, A. Scavenging in changing environments: Woody encroachment shapes rural scavenger assemblages in Europe. Oikos 12 (e09310). https://doi.org/10.1111/oik.09310 (2022).
Turner, K., Abernethy, E., Conner, M., Rhodes, O. & Beasley, J. Abiotic and biotic factors modulate carrion fate and vertebrate scavenging communities. Ecology 92, 2413–2424. https://doi.org/10.1002/ecy.1930 (2017).
Allen, M., Elbroch, M. & Wittmer, H. Scavenging by fishers in relation to season and other scavengers. Ecol. Res. 36, 1–7. https://doi.org/10.1111/1440-1703.12198 (2021).
Tonkin, J., Bogan, M., Bonada, N., Rios-Touma, B. & Lytle, D. Seasonality and predictability shape Temporal species diversity. Ecology 98, 1201–1216. https://doi.org/10.1002/ecy.1761 (2017).
Etter, A. et al. Lista roja de ecosistemas de Colombia. (2017). https://doi.org/10.13140/RG.2.2.10861.08165
Souza, C. et al. Seasonally dry tropical forest Temporal patterns are marked by floristic stability and structural changes. CERNE 27 (e-102355). https://doi.org/10.1590/01047760202127012355 (2021).
Naves-Alegre, L., Morales-Reyes, Z., Sánchez-Zapata, J. A. & Sebastián-González, E. Scavenger assemblages are structured by complex competition and facilitation processes among vultures. J. Zool. 318, 260–271. https://doi.org/10.1111/jzo.13016 (2022).
Beirne, C. et al. Scouts vs. usurpers: alternative foraging strategies facilitate coexistence between Neotropical cathartid vultures. Ibis 166, 1368–1383. https://doi.org/10.1111/ibi.13327 (2024).
Oliva-Vidal, P., Villalba, D., Colomer, M. A. & Margalida, A. Heterospecific visual cues and trophic facilitation processes used by a solitary bone-eating Vulture. Ecosphere 15, e4941. https://doi.org/10.1002/ecs2.4941 (2024).
Cunningham, C. et al. Top carnivore decline has cascading effects on scavengers and carrion persistence. Proc. Biol. Sci. 285, 20181582. https://doi.org/10.1098/rspb.2018.1582 (2018).
Monar-Barragán, P. et al. Impacts of free-ranging dogs on a community of vertebrate scavengers in a high Andean ecosystem. Trop. Conserv. Sci. 16, 1–10. https://doi.org/10.1177/19400829231218409 (2023).
Ferrer-Paris, J. R. et al. An ecosystem risk assessment of temperate and tropical forests of the Americas with an outlook on future conservation strategies. Conserv. Lett. 12, e12623. https://doi.org/10.1111/conl.12623 (2018).
Méndez, D., Olea, P. P., Sarasola, J. H. & Vargas, F. H. Large Andean Condor aggregations at carcasses exacerbate the threat of poisoning. J. Raptor Res. 55, 444–446. https://doi.org/10.3356/JRR-20-26 (2021).
Rivas, C. A., Guerrero-Casado, J. & Navarro-Cerillo, R. M. Deforestation and fragmentation trends of seasonal dry tropical forest in ecuador: impact on conservation. Ecosyst. 8, 46. https://doi.org/10.1186/s40663-021-00329-5 (2021).
Restrepo-Cardona, J. S. & Hull, V. A call to mitigate threats and fill existing knowledge gaps to facilitate the conservation of the Andean Condor in Colombia. Ornito Colom. 23, 44–47. https://doi.org/10.59517/oc.e556 (2023).
Linares-Palomino, R., Oliveira-Filho, A. T. & Pennington, R. T. Neotropical seasonally dry forests: diversity, endemism, and biogeography of Woody plants. In Seasonally Dry Tropical Forests. (eds Dirzo, R., Young, H. S., Mooney, H. A. & Ceballos, G.) 3-21 (Island, 2011). https://doi.org/10.5822/978-1-61091-021-7_1
Cuesta, F. et al. Priority areas for biodiversity conservation in Mainland Ecuador. Neotrop. Biodivers. 3, 93–106. https://doi.org/10.1080/23766808.2017.1295705 (2017).
Moleón, M. et al. Humans and Scavengers: The Evolution of Interactions and Ecosystem Services. BioScience 64, 394–403. (2014). https://doi.org/10.1093/biosci/biu034
Santangeli, A. et al. The global contribution of vultures towards ecosystem services and sustainability: An experts’ perspective. iScience 27, 109925. (2024). https://doi.org/10.1016/j.isci.2024.109925
Moleón, M., Sánchez-Zapata, J. A. & Sebastián-González, E. Owen-Smith, N. Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 124, 1391–1403. https://doi.org/10.1111/OIK.02222 (2015).
Fundación Ceiba. Bosque Seco Lalo Loor. (2023). https://ceiba.org/conservation/dry-forests/lalo-loor/
Third Millennium Alliance. Protecting & Restoring the Most Endangered Rainforest on Earth. (2021). https://www.tma.earth/
Fundación Pro-Bosque. Bosque Protector Cerro Blanco. https://bosquecerroblanco.org/ (2023).
Albuja, L. et al. Fauna De Vertebrados Del Ecuador (Escuela Politécnica Nacional, 2012).
Ministerio del Ambiente del Ecuador. Sistema De Clasificación De Los Ecosistemas De Ecuador Continental (Ministerio del Ambiente del Ecuador, 2013).
Vallejo, A. F. & Boada, C. Dicotyles Tajacu. In Mamíferos Del Ecuador Version 2018.0 (eds Brito, J., Camacho, M. A., Romero, V. & Vallejo, A. F.) (Pontificia Universidad Católica del Ecuador, (2024).https://bioweb.bio/faunaweb/mammaliaweb/FichaEspecie/Dicotyles%20tajacu
Orihuela-Torres, A. et al. Unravelling the vertebrate scavenger assemblage in the gobi desert, Mongolia. J. Arid Environ. 190, 104509. https://doi.org/10.1016/J.JARIDENV.2021.104509 (2021).
Lindén, A. & Mäntyniemi, S. Using the negative binomial distribution to model overdispersion in ecological count data. Ecology 92, 1414–1421. https://doi.org/10.1890/10-1831.1 (2011).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear Mixed-Effects models using lme4. J. Stat. Softw. 67, 1–48. https://doi.org/10.18637/jss.v067.i01 (2015).
James, G., Witten, D., Hastie, T. & Tibshirani, R. An Introduction To Statistical Learning: with Applications in R (Springer, 2017).
Burnham, K. P. & Anderson, D. R. Model Selection and Multimodel Inference. A Practical information-theoretic Approach (Springer, 2002).
Bartoń, K. Package ‘MuMIn’: Multi-model inference. R Package Ver. 1.48.11. (2025). https://doi.org/10.32614/CRAN.package.MuMIn
R Core Development Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2021).
Butler, J. R. A. & du Toit, J. T. Diet of free-ranging domestic dogs (Canis familiaris) in rural zimbabwe: implications for wild scavengers on the periphery of wildlife reserves. Anim. Conserv. 5, 29–37. https://doi.org/10.1017/S136794300200104X (2002).
Olea, P. P., Iglesias, N. & Mateo-Tomás, P. Temporal resource partitioning mediates vertebrate coexistence at carcasses: the role of competitive and facilitative interactions. Basic. Appl. Ecol. 60, 63–75. https://doi.org/10.1016/j.baae.2022.01.008 (2022).
Naves-Alegre, L., Morales-Reyes, Z., Sánchez-Zapata, J. A., Sebastián-González, E. & Ovaskainen, O. Scavenging in the realm of senses: Smell and vision drive recruitment at carcasses in Neotropical ecosystems. Proc. Biol. Sci. 289, 20220843. (2022). https://doi.org/10.1098/rspb.2022.0843
Morales, J. D. Descubriendo El Bosque Seco Tropical Con Las Aves De Cerro Blanco (The Wild GYE Initiative, 2022).
Wenting, E. et al. Influence of tree cover on carcass detection and consumption by facultative vertebrate scavengers. Ecol. Evol. 14, e10935. https://doi.org/10.1002/ece3.10935 (2024).
De Araujo, G., Peres, C., Baccaro, F. & Guerta, R. Urban waste disposal explains the distribution of black vultures (Coragyps atratus) in an Amazonian metropolis: management implications for birdstrikes and urban planning. PeerJ 6, e5491. https://doi.org/10.7717/peerj.5491 (2018).
Keppeler, R., Ticiani, D., Teston, G., Onghero, O. & Favretto, M. Registro de comportamento de Pesca Por Urubu (Coragyps atratus) no Brasil. Biota. Amazon. 10, 62–63 (2020).
Hill, J. et al. Diets of black vultures and Turkey vultures in coastal South carolina, USA with a review of species’ dietary information. SENA 21, 11–27. https://doi.org/10.1656/058.021.0102 (2022).
Mateo-Tomás, P., Olea, P., Moleón, M., Selva, N. & Sánchez-Zapata, J. A. Both rare and common species support ecosystem services in scavenger communities. Glob Ecol. Biogeogr. 26, 1459–1470. https://doi.org/10.1111/geb.12673 (2017).
Salas, J. A., Navas, I. B., Merchán, M. B., Medranda-Benavides, J. & Hurtado, C. M. Riqueza, abundancia relativa y patrones de actividad de Mamíferos Medianos y grandes En El Bosque protector Cerro Blanco (Guayas, Ecuador). Mamm. Aequat. 4, 9–23. https://doi.org/10.59763/mam.aeq.v4i.53 (2022).
Morales-Reyes, Z. et al. Scavenging efficiency and red Fox abundance in mediterranean mountains with and without vultures. Acta Oecol. 79, 81–88. https://doi.org/10.1016/j.actao.2016.12.012 (2017).
Freile, J. F. et al. Lista Roja De Las aves Del Ecuador (Ministerio Del Ambiente, Aves Y Conservación, Comité Ecuatoriano De Registros Ornitológicos, Fundación Charles Darwin, Universidad Del Azuay (Red Aves Ecuador y Universidad San Francisco de Quito, 2019).
BirdLife International. Species factsheet: Sarcoramphus papa. (2025). https://datazone.birdlife.org/species/factsheet/king-vulture-sarcoramphus-papa
Restrepo-Cardona, J. S. et al. New records of domestic dog negative impacts on native vertebrates in the Ecuadorian Andes. Neotrop. Biol. Conserv. 20, 93–101. https://doi.org/10.3897/neotropical.20.e148221 (2025).
Restrepo-Cardona, J. S., Kohn, S., Narváez, F., Pineida, R. & Vargas, F. H. Life history of the Andean Condor in Ecuador. Trop. Conserv. Sci. 17, 1–8. https://doi.org/10.1177/19400829241238005 (2024).
Lambertucci, S. et al. Presumed killers? Vultures, stakeholders, misperceptions, and fake news. Conserv. Sci. Pract. 3, e415. https://doi.org/10.1111/csp2.415 (2021).
Margalida, A., Campión, D. & Donázar, J. A. Scavenger turned predator: European vultures’ altered behaviour. Nature 480, 457. https://doi.org/10.1038/480457b (2011).
Sáenz-Jiménez, F. et al. Estimating the Andean Condor Vultur gryphus population at the Northern edge of its geographical range: a citizen science-based approach. Bird. Conserv. Int. 35, 1–9. https://doi.org/10.1017/S0959270925000061 (2025).
Restrepo-Cardona, J. S., Saenz-Jimenez, F. & Lieberman, A. A. Traditional knowledge and perceptions towards the Andean Condor (Vultur gryphus linnaeus, 1758) in the central Andes of Colombia. Ethnoscientia 4, 1–7. https://doi.org/10.22276/ethnoscientia.v4i1.211 (2019).
Ballejo, F., Plaza, P. I. & Lambertucci, S. A. The conflict between scavenging birds and farmers: field observations do not support people’s perceptions. Biol. Conserv. 248, 108627. https://doi.org/10.1016/j.biocon.2020.108627 (2020).
Acknowledgements
We thank The Peregrine Fund, Fundación Cóndor Andino Ecuador, and the Third Millennium Alliance for their logistical and financial support for the development of the research. To The Wild GYE Initiative for their support in the field. To Moisés Tenorio for his assistance with the fieldwork in Manabí. To the Cerro Blanco Protective Forest and the Lalo Loor Reserve for providing access to the study areas. To Esther Sebastián-González for her help in the statistical models and model selection. We also thank the Associate Editor and two anonymous reviewers for their valuable comments, which significantly improved the content of this paper.
Author information
Authors and Affiliations
Contributions
P.M.B.: Conceptualization, methodology, validation, data analysis and statistical analysis, writing-original draft, writing-review and editing. F.H.V.: Conceptualization, methodology, writing-review, supervision. R.P.: Data collection, methodology, writing-review. A.S.: Data collection, methodology. E.V.: Methodology, validation, statistical analysis, writing-review and editing, supervision. F.N.: Conceptualization, methodology, validation. S.K.: Conceptualization, methodology, validation. J.D.M.: Data collection, methodology. J.S.R.C.: Conceptualization, methodology, validation, writing-review and editing, supervision.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Monar-Barragán, P., Vargas, H., Pineida, R. et al. Vulture dominance in a scavenger assemblage in the Neotropical dry forest. Sci Rep 15, 24019 (2025). https://doi.org/10.1038/s41598-025-08406-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-08406-3
