
Abstract
The Sichuan Basin has yielded abundant sauropod dinosaurs from the Middle-Late Jurassic, and Mamenchisauridae had predominated the dinosaur faunae during the Late Jurassic in the Sichuan Basin. Here, we describe a new sauropod dinosaur, Tongnanlong zhimingi gen. et sp. nov. from the Upper Jurassic Suining Formation in the Sichuan Basin, southwestern China. The holotype includes three dorsal and six caudal vertebrae, scapula, coracoid, and some hindlimb bones. It is diagnosed by the complex structures of the dorsal and anterior caudal vertebrae compared to other mamenchisaurids. Phylogenetic analysis shows that it is more closely related to Mamenchisaurus than to Omeisaurus. The huge-sized scapula and coracoid also indicate that the specimen belongs to an extremely gigantic individual with a body length approaching about 25 ~ 26 m. The new specimen enriches the diversity of Mamenchisauridae and provides additional information for understanding the evolution and diversity of eusauropod dinosaurs.
Similar content being viewed by others
Introduction
Sauropods were gigantic, quadrupedal herbivores and the largest terrestrial dinosaurs ever existed. They appeared in the Late Triassic, achieved a global distribution by the Middle Jurassic, and finally died out at the end of the Late Crertaceous1. More than 150 genera have been identified, including over 20 genera from the Jurassic period in China1,2,3. The southwestern China is a key region that yields Middle-Late Jurassic sauropod dinosaurs, particularly the Sichuan Basin. The sauropod fauna from the Middle-Late Jurassic Sichuan Basin has previously been considered an endemic fauna which differs from the contemporaneous sauropod faunae from the Pangaean terrestrial faunae. This distribution has often been interpreted by the “East Asian Isolation” hypothesis, which occurred during the Jurassic to Early Cretaceous1,4,5. However, the “East Asian Isolation” hypothesis is challenged by more phylogenetic analyses and studies of neosauropod dinosaurs from China6,7 and mamenchisaurid dinosaurs discovered from Africa4 which support the idea that they were distributed globally during the Middle Jurassic.
The sauropod fauna from the Middle Jurassic shows a different pattern compared to that from the Late Jurassic in the Sichuan Basin. Comparing to the abundant and diverse Middle Jurassic sauropods like Shunosaurus, Omeisaurus, Datousaurus and Dashanpusaurus, the Late Jurassic dinosaur fauna became simplified and was predominated by Mamenchisaurus2,8. However, the sauropods diversity and disparity in the Late Jurassic have been gradually revised based on newly discovered sauropods fossils in recent years. Xing et al. (2015) reported a new sauropod, Qijianglong guokr, from the Late Jurassic Suining Formation, which is the first non-Mamenchisaurus sauropod dinosaur of this formation except the only reported sauropod Mamenchisaurus anyuensis9,10.
Here we report a new sauropod dinosaur, Tongnanlong zhimingi, gen. et sp. nov., from the Upper Jurassic Suining Formation in the Sichuan Basin. Anatomical and phylogenetic analysis demonstrate that Tongnanlong is a mamenchisaurid eusauropod. This discovery increases the taxonomic diversity of mamenchisaurid and strengthens the idea that Mamenchisauridae was a non-endemic clade that radiated widely in Asia and Africa during the Middle-Late Jurassic.
Geological background
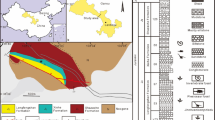
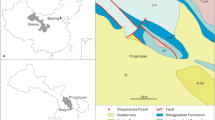
The Sichuan Basin is an important Mesozoic continental basin located in southwestern China11,12,13. The terrestrial strata of the basin are fully exposed and widely distributed, from the bottom to top including the Upper Triassic Xujiahe Formation, the Lower Jurassic Zhenzhuchong and Ziliujing formations, the Middle Jurassic Xintiangou and the Lower Shaximiao formations, and the Upper Jurassic Upper Shaximiao, Suining and Penglaizhen formations, and the Lower Cretaceous Jiaguan Formation. Among them, the Jurassic deposits of the Sichuan Basin is famous for yielding abundant dinosaur fossils and tracks8,14. Notably, the era of Suining Formation, along with the Upper Shaximiao Formation, belongs to the late Late Jurassic is controversial. Most of the sedimentology studies assigned the Suining Formation to the Upper Jurassic15,16,17but some chemical analyses of zircon U-Pb dating indicated that it belongs to the Lower Cretaceous13,18. Dinosaur and sporopollen assemblies support the Suining Formation to the Late Jurassic2,8,19. Here, in this study, we use the division scheme that suggested the Suining Formation belongs to the Late Jurassic based on the features of its dinosaur fauna8.
In the Sichuan Basin, almost 80% of the dinosaur and non-dinosaur vertebrate fossils were unearthed from the Middle-early Upper Jurassic strata of the Lower Shaximiao and Upper Shaximiao formations2,8. Dong et al. (2015) considered that the dinosaurs from the Lower Shaximiao and Upper Shaximiao formations shows different fauna assemblages and assigned them to the Shunosaurus Fauna and Mamenchisaurus Fauna, respectively2. Comparatively, the Lower Jurassic Zhenzhuchong Formation only yields several sauropod tracks but no bones, while the Ziliujing Formation includes several basal sauropodmorphs and theropod tracks14. The Upper Jurassic Suining Formation has yielded rare dinosaur bones and tracks, such as Qijianglong guokr from the Suining Formation and M. anyuensis materials from the top of the Suining Formation and the bottom of the Penglaizhen Formation9,10,14. It is noteworthy that the layer of Qijianglong guokr is lower than the specimen reported in this paper based on our field work.
In 1998, the fossil specimen reported in this paper was excavated from a building site in the Tongnan District of Chongqing area together with many well-preserved, articulated post-cranial skeletal bones (Fig. 1). Our field work shows that the fossiliferous site belongs to the lower portion of the Upper Jurassic Suining Formation, overlying by the Quaternary deposits. The Suining Formation consists of purplish red mudstone and sandstone. Abundant invertebrate fossils are known from this formation, especially ostracods and stoneworts, along with some freshwater bivalves, conchostracans. A few vertebrates are also known from this formation, such as fish Ceratodus szechuanensis, turtle Plesiochelys tatsuensis, and dinosaur Mamenchisaurus anyuensis9,20,21,22,23,24.
The new specimen described in this study shows a nearly in-situ burial state and thus likely experienced a very short-distant transportation before burial. Its layered deposits are characterized by purplish red sandstone with a wavy texture, indicating a lakeshore to shallow lake environment and relatively arid climate. Here, we provide a systematic description of this new specimen and discuss its implications for sauropod palaeogeography.

Maps of the fossil locality in Tongnan District, Sichuan Basin, China. The star represents the fossil site. (A) Sketch map of China showing the geographical locations of Sichuan Basin. The base map of China is modified after the Standard Map Service of the National Administration of Surveying, Mapping and Geoinformation of China (http://bzdt.ch.mnr.gov.cn/) (No. GS (2016)1585); (B) Locality of Tongnan District (marked by a irregular black area); (C) Fossil site and its geological background (marked by a red rectangle in B) modified from the 1:200 000 standard map of Suining (Map No.: H-48-(16) published in 1981).
Results
Systematic paleontology
Saurischia25.
Sauropodomorpha26.
Sauropoda27.
Eusauropoda28.
Mamenchisauridae29.
Tongnanlong zhimingi gen. et sp. nov.
Etymology. The generic name Tongnan, Chinese Pinyin, refers to the Tongnan District, where the holotype was found; the word ‘long’ in Chinese Pinyin for dragon. The specific name is in honor of the world-famous dinosaur scientist Mr. Dong Zhiming for his great contributions to the dinosaur research in the Sichuan Basin.
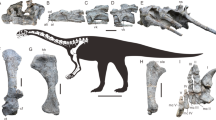
Holotype. An incomplete individual that includes three dorsal and six caudal vertebrae, left shoulder girdle (scapula and coracoid) and hindlimb elements (partial tibia, fibula, three metatarsals, and two unguals). The specimen is housed in Tongnan Museum, with the specimen number of TNM 0254.
Type locality and horizon. The building site of Dafo Street (with GPS N30°10’44”, E105°48’17”), Tongnan District; Upper Jurassic, Suining Formation.
Diagnosis. A huge-bodied mamenchisaurid dinosaur with the following unique combination of characters: dorsal centra opisthocoelous, camerate pneumatic structures of dorsal centra, well-deveolped spinopostzygapophysial lamina of dorsal vertebrae, anterior caudal centra procoelous, neural spine of anterior caudal vertebrae oriented posterodorsally, small lamina within the pleurocoels on dorsal centra, neural spine of the dorsal vertebrae and postozygapophyses forming laminae and fossae, fossa developed between the postzygapopaphysis and hyposphene on anterior caudal vertebra, S-shaped suture of scapula and coracoid, obtuse angle of the scapular and coracoid articular surfaces of glenoid, coracoid fossa much close to the scapular margin than to the glenoid margin, scapular acromion weakly developed and bearing no hook-like process.
Description
The specimen was excavated from a quarry, including three primarily articulated dorsal vertebrae, six caudal vertebrae, one anterior caudal neural spine, one nearly complete anterior chevron, three isolated metatarsals and two claws, complete left scapular and coracoid, one complete right fibula, proximal end of a tibia and some isolated dorsal rib fragments (Figs. 2, 3, 4 and 5, Tables S1, S2).

Fossil remains of Tongnanlong zhimingi (TNM 0254). (A) Skeletal outlines showing recovered elements in blue color. The skeletal reconstruction is the proportional scaling of Mamenchisaurus youngi with copyright Scott Hartman (2022) (https://www.skeletaldrawing.com/sauropods-and-kin); (B1-B3) Dorsal vertebrae in lateral view; (B4) dorsal neural spine in posterior view; (C1-C5) Caudal vertebrae in lateral view; (C6) caudal neural spine in lateral view; (C7) chevron in lateral-posterior view; (D) Scapula and coracoid in lateral view; (E) Fibula in anterior view; (F1-F3) Metatarsals in dorsal view; (G1, G2) Claws in lateral view.
Dorsal vertebrae. Three dorsal vertebrae were naturally connected when being excavated but later on isolated due to improper storage. The dorsal vertebrae are medially opisthocoelous. The anterior articular surface of the dorsal centrum is convex and elliptical in shape whilst the posterior articular surface is concave and pot-like in shape. The margin of the convex anterior articular surface and its dorsal centrum is clear and forms a ridge-like line. The height of each dorsal centrum is greater than its width as in other mamenchisaurids. The broken surface of the dorsal centrum shows typical grid structure and camerate of internal tissue texture (Fig. 3B3, C3), which is similar to those of the middle-posterior dorsal centra in Mamenchisaurus youngi30. The ventral surface of the dorsal centrum bears no keel, and is transversely smooth and concave, which is similar to most sauropods like Omeisaurus tianfuensis, Shunosaurus lii, Mamenchisaurus youngi, and Analong chuanjieensis9,30,31,32. The pleurocoel of the dorsal centrum is oval and located much anterodorsally. The pleurocoel of the dorsal centrum bears strong ventral margin, but its dorsal margin flushes into a lateral surface, which is similar to those of the middle dorsal centra in Mamenchisaurus youngi (ZDM 0083), but different from those clearly whole pleurocoel margin in Omeisaurus tianfuensis (ZDM 5002)9,30. Moreover, the pleurocoel develops a further deep excavation which is divided by the vertically orientated internal laminae. But in other eusauropod dinosaurs like Mamenchisaurus youngi (ZDM 0083), Shunosaurus lii (ZDM 5008) and Omeisaurus tianfuensis (ZDM 5002), the laminae in the pleurocoel of the dorsal centra are absent8,30. The neural arches of the dorsal vertebrae were artificially destroyed after excavation. Their neural spines are relatively positioned backward, and each extends nearly to the posterior end of its centrum. The dorsal neural spine is oriented vertically and slightly posterodorsally. The postzygapophyseal surface is relatively small and its lamina is less developed (Fig. 3D). In lateral view, the spinopostzygapophyseal lamina of the neural spine is thin and well developed. The prezygapophyses are lean and form an acute angle of about 60°, but the postzygapophyses are nearly horizontal. The head of the dorsal neural spine is coarse and weakly bifurcated. In posterior view, the coarse spinal ridge extends vertically along the midline and becomes bifurcated toward the base of the neural spine and further forms a clearly vertically oriented rhombic spinopostzygapophysial fossa (i.e. cen. sp-poz. f) (Fig. 3D1). Moreover, the base of the neural spine and the postogapophyses also form four lateral spinopostzygapophysial fossae which are divided by small laminae (i.e. med. sp-poz. f and lat. sp-poz. f) (Fig. 3D1). These fossae are symmetrically distributed on bilateral sides of the neural spinal ridge, and the medial ones (i.e. f1) are much bigger and deeper than the lateral ones (i.e. f2) (Fig. 3D1). The central spinopostzygapophysial fossa and medial spinopostzygapophysial fossae are divided by small, thin vertical laminae (i.e. med. sp-poz. l). The lateral spinopostzygapophysial fossae become shallow till missing when extending upward to the less lower half of the neural spine. The spinopostzygapophysial fossae and laminae structures are unique that never been seen in other eusauropods and mamenchisaurids such as in Mamenchisaurus and Omeisaurus8,30. In lateral view, the neural canal is enclosed by pedicel that is partly located in its centrum rather than the whole neural arch. In posterior view, the neural canal forms a deep fossa with a vertically oval-shaped outline (Fig. 3C5). The height ratio of the neural canal to its centrum is only about 14% (5 cm and 33 cm, respectively). The neural arch is partly preserved and positioned relatively high, and the transverse process is above the horizontal level of the neural canal, which indicates that the dorsal vertebrae belong to the middle part of the dorsal series.

Dorsal vertebrae of Tongnanlong zhimingi. (A-C) Dorsal centra, (1–6) in left lateral, right lateral, dorsal, anterior, posterior and ventral views; (D1-D3) Dorsal neural spine in posterior, lateral and dorsal views; (E1,E2) Dorsal neural spine in lateral-posterior and lateral views; (F) Dorsal neural spine in anterior-lateral views; Scale bar = 10 cm. cen.sp-poz.f, central spinopostzygapophysial fossa; pl, pleuroceol; lat.sp-poz.f (f1), lateral spinopostzygapophysial fosssa; poz, postzygapophysis; di, diapophysis; med.sp-poz.f, medial spinopostzygapophysial fossa; med.sp-poz.l (f1), medial spinopostzygapophysial lamina; n.c, neural canal; sp, neural spine; spoz.l, suprapostzygapophysial lamina.
Caudal vertebrae. Six caudal vertebrae are preserved with three of them belonging to the anterior portion and the other three belonging to the middle-posterior portion. The three anterior caudal centra are procoelous. The posterior articular process of the anterior caudal centrum is medially convex and forms a marked condyle. The condyle length is about one third of its centrum (with length of 6 cm and 19 cm, respectively) (Fig. 4B). The margin of the posterior articular process and its centrum is clear and forms an oval ridge-like outline. The anterior caudal centra are taller than wide. The lateral surfaces of the caudal centra are smooth and shrunk inwardly, and bear no lateral fossa. The ventral surface of the caudal centrum bears a shallow longitudinal hollow along its midline. The caudal rib is low and situated wholly on the centrum, much closer to the posterior articular process than to the anterior one (Fig. 4A1, B1, C1). The hyposphenal ridge of the neural arch is present and slender (Fig. 4G2). The postzygapophysis extends less to the posterior articular surface of its centrum. The anterior neural spine is posterodorsally oriented and blade-like in shape, with a width/length ratio of about 20% (1.4 cm and 6.5 cm, respectively) (Fig. 4G). The anterodorsal margin of the neural spine is situated anteriorly and does not extend to the postzygapophyses. In anterior view, the spinoprezygapophyseal lamina is clearly developed and positioned near to the base of the neural spine. In posterior view, the spinopostzygapophyseal lamina is weakly developed, which makes the posterior surface of the neural spine vertically concave with a central ridge dividing the concavity into two parts (i.e. sp.r in Fig. 4G3). The postzygapophyses are much more vertical and their faces form a small acute angle of about 30°. In posterior view, a dorsoventrally elongated fossa is present between the postzygapophyses and hyposphene (i.e. poz-hyp. f in Fig. 4G1, G2), which is a unique character that differs from the absence of fossa in the anterior caudal vertebrae of other eusauropods like Mamenchisaurus and Omeisaurus1,8,30.
An isolated caudal vertebra is partly preserved but clearly different from the three anterior caudals (Fig. 4D), which belongs to the middle-posterior part of the caudal series. Its centrum is weakly amphicoelous. Both the anterior and posterior articular surfaces are ellipse-shaped. The ventral surface bears a clear concavity on the midline. Its prezygapophysis is weakly developed, extending anteriorly no more than the articular surface. The neural spine is low and extends posteriorly more than the posterior articular surface. Another two naturally articulated posterior caudal centra are preserved (Fig. 4E). Their length is clearly greater than their height and width (16 cm, 14 cm and 10 cm, respectively), which makes the centra laterally compressed in shape. Both centra are weakly amphicoelous. Both the anterior and posterior articular surfaces are oval in outline. The ventral surface of each centrum bears a clear anteroposteriorly elongated vascular groove. The neural canal is situated wholly above its centrum.
Chevron. The proximal end of the preserved chevron is bridged dorsally by a bone bar (Fig. 4F). The proximal end also bears a deep haemal canal, and its shaft below the canal curves slightly backward, which indicates that it belongs to an anterior chevron.

Caudal vertebrae and chevron of Tongnanlong zhimingi. (A-E) Caudal vertebrae, (1–6) in left lateral, right lateral, dorsal, ventral, anterior and posterior views; (F1-F4) Chevron in anterior-lateral, posterior-lateral, lateral and top views; (G1-G3) Caudal neural spine in left lateral, posterior, right lateral and dorsal views. (A-G) with scale bar = 10 cm. hyp, hyposphene; n.c, neural canal; poz, postzygapophysis; prz, prestzygapophysis; sp, neural spine; sp.r, ridge of neural spine; poz-hyp.f, postzygapopaphysis-hyposphene fossa.
Ribs. The dorsal ribs are broken. One nearly complete dorsal rib is more than 150 cm long. In proximal view, the capitulum and tuberculum are both robust, forming an acute angle. The plate-like process is developed anteriorly to the tuberculum, which indicates that it is an anterior dorsal rib.
Scapula. The left scapula and coracoid are completely preserved (Fig. 5A). The scapula is relatively large, and its blade is long and straight. The ratio of the maximal dorsoventral height to the total anteroposterior length of the scapula approaches about 60% (109 cm and 182 cm, respectively), this ratio only reaches 50% in that of Mamenchisaurus youngi (ZDM 0126)30. The proximal end of the scapula is extremely extended whilst the distal end is only weakly extended dorsoventrally. The highest point of the dorsal margin of the blade is clearly lower than the dorsal margin of the proximal end. The cross section of the scapular blade at the base is D-shaped, with the lateral surface strongly convex dorsoventrally and medial surface flat, which is similar to that in Analong chuanjieensis31Dashanpusaurus dongi6Bellusaurus sui33 but different from the rectangular shape in Shunosaurus lii (ZDM 5007)6. The scapular proximal plate develops a strong acromial ridge, extending dorsally from the margin of the proximal plate and ventrally to the margin of the glenoid. The area of the anterior acromial ridge forms a distinct concavity, which is similar to that in Mamenchisaurus youngi (ZDM 0126)30. The acromion is well-developed on the dorsal end of the proximal plate and forms a rounded outline other than a posteriorly directed hook-like process, which is similar to that in Mamenchisaurus jingyanensis34. The suture of scapula and coracoid is S-shaped other than nearly straight as those in other eusauropods like Mamenchisaurus youngi (ZDM 0126) and Omeisaurus tianfuensis (ZDM 5002). Also, the suture and the scapular blade midline forms a broad obtuse angle (about 110°) other than nearly perpendicular as in that of Mamenchisaurus youngi (ZDM 0126). The gleniod is thick mediolaterally. Its scapular and coracoid articular surfaces form a broad obtuse angle (about 110°), differing from the right angle (nearly 90°) in that of Mamenchisaurus youngi (ZDM 0126)30 and neosauropods like Ruyangsaurus35.
Coracoid. The coracoid is relatively quadrilateral in shape with a dorsoventral height greater than anteroposterior length (about 68 cm and 46 cm, respectively). In lateral view, the dorsal margin of the coracoid lies below the level of the scapular proximal plate and separates the latter by a broad V-shaped notch. The glenoid articular surface of the coracoid is weakly concave. The lateral surface of the coracoid develops a convex notch that attaches the biceps. The coracoid fossa is big and subcircular (with a diameter of about 9 cm), which is located much closer to the scapular margin of the coracoid than to the glenoid margin of the coracoid (with distances of 7 cm and 18 cm, respectively).
Fibula and tibia. The right fibula is completely preserved (Fig. 5B). The fibula is slender with a shaft length of about 103 cm. The fibular proximal end is extremely extended, and its proximal margin is relatively straight (Fig. 5B2). The distal end is more extended anteroposteriorly than mediolaterally. Both the proximal and distal articular surfaces of the fibular are coarse. The cross-section of the middle shaft of fibula is D-shaped. The left fibula only preserves the proximal and distal ends (Fig. 5C, D). The anterior surface of the distal end of the fibular shaft bears a clear longitude ridge that separates the anterior surface into two parts.
Only the proximal end of the left tibia is preserved (Fig. 5E). In posterior view, the tibial lateral surface is thinner and greater than its medial surface, which further form a clear vertical crest as those in other eusauropods1.
Pes. Three metatarsals are completely preserved (Fig. 5F, G, H). The metatarsals are smooth laterally with the middle shafts shrunk inwardly. The metatarsal V is well preserved and dorsoventrally compressed (Fig. 5H). Its proximal end is extremely extended mediolaterally while the distal end weakly extended dorsoventrally. The distal articular surface is fan-shaped. The maximal transverse width of the distal end of the metatarsal V is only half of its proximal end (8 cm and 16 cm, respectively). The ratio of the shaft length to the proximal transverse width of metatarsal V is about 1.4 (23 cm and 16 cm, respectively), which is less than 1.5 as in other eusauropods1. The dorsal surface of the shaft of metatarsal V bears a ridge along the midline. Another two metatarsals are relatively slender and only slightly longer than the metatarsal V (Fig. 5F, G). Their proximal ends are mediolaterally expanded with clear articular surfaces developed on the medial and lateral sides, which indicate they belong to middle metatarsals and possibly metatarsal III or IV.
Pedal ungual. Two unguals were discovered (Fig. 5I, G). They are almost identical in shape but different in size. The ventral margins of unguals are only slightly curved but dorsal margins strongly curved. These unguals are strongly compressed mediolaterally and their tips are not that sharp. The lateral surface of each ungual bears a shallow groove. Their proximal articular surfaces are slightly concave. The ventral margin of each ungual develops a tuberosity along its distal half portion. In sauropod dinosaurs, the manus only bears one ungual in digit I and hoof-like bones in digits II-V; the pes bears claws I-V, and these unguals are similar in shape with the first pedal ungual being the biggest and unguals II-V being smaller in size, for example, the pedal ungual I is twice as large as the pedal ungual II in Omeisaurus tianfuensis9,36. Thus, we infer that the two unguals belong to the pes.

Shoulder girdle and hindlimb bones of Tongnanlong zhimingi. (A) Left scapula and coracoid in external view; (B1-B3) right fibula in anterior, proximal and distal views; (C1,C2) proximal end of left fibula in anterior and posterior views; (D1-D3) distal end of the left fibula in anterior, posterior and distal views; (E1-E2) proximal end of the left tibia in anterior, posterior and proximal views; (F1-F6) Metatarsal III in dorsal, ventral, dorsal-lateral, ventral-lateral, proximal and distal views; (G1-G6) Metatarsal IV in dorsal, ventral, lateral, proximal and distal views; (H1-H5) Metatarsal V in dorsal, ventral, lateral, proximal and distal views; (I1-J3) Claws in internal, external and proximal views. (A) with scale bar = 100 cm; (B-J) with scale bar = 10 cm. Abbreviations: ac, acromion; acr, acromial ridge; co, coracoid; cof, coracoid fossa; gl, glenoid; sc, scapular blade.
Comparative discussion
The phylogenetic analysis placed Tongnanlong as a derived eusauropod dinosaur stably located in the clade of Mamenchisauridae. Tongnanlong has a closer relationship to Jingiella, Mamenchisaurus and Chuanjiesaurus than to Omeisaurus. Phylogenetic result also supports Mamenchisauridae as a stable group in Eusauropoda (Fig. 6, Fig. S1).

The strict consensus of the 35 most parsimonious trees showing the systematic position of Tongnanlong zhimingi among sauropod dinosaurs (with consistency index = 0.335 and retention index = 0.685).
Tongnanlong shares some characters with Mamenchisaurus hochuanensis29Mamenchisaurus youngi30Mamenchisaurus anyuensis9and Omeisaurus tianfuensis36 which supports the assignment to Mamenchisauridae: the ratio of scapular total length/blade minimal width of more than 6, the dorsal margin of the scapular acromion within a rounded outline, the opisthocoelous dorsal vertebrae and procoelous anterior caudal centra, pneumatic structures and bifurcated neural spine of dorsal vertebrae, dorsal centra anteroposteriorly longer than dorsoventrally tall, anterior caudal central height greater than its width, lateral pleurocoels on dorsal centra.
Tongnanlong differs from other mamenchisaurids for bearing the following unique combination of characters, which support the erection of a new genus and species other than assigning it to known mamenchisaurids: complex spinopostzygapophaysial laminae and fossae of the dorsal vertebrae (also in Xinjiangtitan shanshanensis), small vertical laminae dividing the lateral pleurocoels on dorsal centra, the suture of scapula and coracoid curved into S-shaped (also in Tienshanosaurus chitainensis), a fossa well-develooped between the postzygapopaphyses and hyposphene of the anterior caudal neural spine. Moreover, Tongnanlong has the following differences compared to other eusauropods, particularly the mamenchisaurid taxa from the adjacent regions in southwestern China, especially the Sichuan Basin and Yunnan Province.
Tongnanlong differs from Mamenchisaurus anyuensis9,23 in various aspects. Due to the brief description of M. anyuensis in published literature, the following comparisons are based on firsthand, personal observation of its holotype (AL 001). The distal end of scapular blade is only slightly extended and its exterior surface is concave in Tongnanlong but relatively extended with a flat lateral surface in M. anyuensis. The hook-like process of the scapular acromion is absent in Tongnanlong but developed in M. anyuensis. The acromial ridge is curved and extends dorsally above the glenoid in Tongnanlong but it is straight and extends posteriorly to the glenoid in M. anyuensis. The scapular proximal plate is much extended dorsoventrally in Tongnanlong than that in M.anyuensis. The coracoid outline is relatively quadrolateral in Tongnanlong but regularly subrounded in M.anyuensis. The exterior surface of coracoid is slightly concave in Tongnanlong while convex in M. anyuensis. The sizes of dorsal vertebrae, scapula and coracoid of Tongnanlong are all bigger than those of M. anyuensis, which indicates that Tongnanlong is a larger mamenchisaurid individual than M. anyuensis (Fig. S2).
Tongnanlong differs from Mamenchisaurus youngi (ZDM 0083)30 in various aspects, such as less extended distal end of scapular blade than that in M. youngi, rounded outline and absent hook-like process of the scapular acromion whereas straight outline with weakly hook-like process in M. youngi; coracoid fossa less close to the scapular suture than that in M. youngi, and irregularly quadrilateral of coracoid but regularly oval in M. youngi.
Tongnanlong differs from Mamenchisaurus hochuanensis29 by having lateral pleurocoels on dorsal centra that are much developed than that in M. hochuanensis. The caudal neural spine is blade-like in Tongnanlong whilst rod-like in M. hochuanensis. The metatarsal V of Tongnanlong is much slenderer with a ratio of the proximal width to its shaft length about 50% (16 cm and 24 cm, respectively), but the ratio reaches to 83% (15 cm and 18 cm, respectively) in M. hochuanensis. Also, the scapular distal end in Tongnanlong is less extended than that in M. hochuanensis (ZDM 0126)37.
Tongnanlong differs from the Mamenchisaurus jingyanensis34 in that it bears well developed lateral pleurocoels on dorsal centra, but pleurocoels are absent in M. jingyanensis. The articular surface of middle caudal centrum of Tongnanlong is oval and laterally compressed but circular in M. jingyanensis. The scapular proximal plate is much extended dorsally in Tongnanlong than that in M. jingyanensis. The scapular blade and proximal plate form a nearly right angle on the dorsal margin in Tongnanlong, but it is a broad obtuse angle in M. jingyanensis. The suture of scapula and coracoid is curved and S-shaped, but it is almost straight in M. jingyanensis.
Tongnanlong differs from the Omeisaurus tianfuensis (ZDM 5002)36 in that the head of the dorsal neural spine is weakly bifurcated of Tongnanlong while not bifurcated in O. tianfuensis. The lateral pleurocoels of dorsal centra only had clear ventral margin in Tongnanlong whilst a wholly sub-rounded margin in O. tianfuensis. The pleurocoel of dorsal centrum in Tongnanlong is shallower than that in O. tianfuensis which is extremely developed and almost pokes through the dorsal centrum.
Tongnanlong differs from the Zigongosaurus fuxiensis (ZDM 00251; CQMNH CV 025011)8,38 in that the height of scapular proximal plate is smaller than the length of scapular blade in Tongnanlong, whereas the proximal plate height is greater than its blade in Z. fuxiensis. The shape of the coracoid is irregular in Tongnanlong but regularly oval in Z. fuxiensis.
Tongnanlong differs from the Shunosaurus lii (IVPP V 9065)8,32,39 in that the dorsal centra are opisthocoelous but weakly amphicoelous in S. lii. The dorsal pleurocoels strongly developed in Tongnanlong whilst weakly developed in S. lii (ZDM 5008). The scapular proximal plate is less extended in S. lii (ZDM 5003) than that in Tongnanlong.
Tongnanlong differs from Analong chuanjieensis31 in that the neural canals are nearly circular-shaped but considerably slot-shaped in A. chuanjieensis. The centrum is taller than wide in Tongnanlong while conversely in A. chuanjieensis. The neural spine of anterior caudal vertebrae is plate-like in Tongnanlong whilst rod-like in A. chuanjieensis.
Tongnanlong differs from Qijianglong guokr10 in some aspects even though some similarities are present in their morphology, for example the strongly opisthocoelous dorsal centra, and they were both yielded from the Suining Formation. The articular surfaces of dorsal and caudal centra are anteroposteriorly shorter than dorsoventrally tall in Tongnanlong while conversely in Q. guokr. In dorsal view, the dorsal neural spine forms a clear midline groove in Q. guokr but absent in Tongnanlong. The dorsal neural spine is thinner anteroposteriorly in Q. guokr than those in Tongnanlong. Also, the pleurocoels on the dorasl centrum is smaller and bears no vertical laminae in Q. guokr compared to those in Tongnanlong. The sizes dorsal centra are bigger in Tongnanlong than those in Q. guokr, which indicates a much huger individual than the latter. However, it is difficult to make further detailed comparison due to the fragmentary nature of the bones or the lack of overlapping bones, such as the partly preserved anterior caudal centra in Tongnanlong and the nearly complete mid-distal caudal verterbrae in Q. guokr.
Tongnanlong differs from Yuanmousaurus jiangyiensis40 in that the dorsal and caudal centra are clearly taller than wide whilst nearly equal in Y. jiangyiensis. The scapular proximal plate is much extended in Tongnanlong than that in Y. jiangyiensis, which shows a ratio of proximal plate width/blade length of scapular about 0.91 in Tongnanlong but only 0.69 in Y. jiangyiensis. Also, the sizes of scapula and coracoid are greater in Tongnanlong than that in Y. jiangyiensis, which indicates that Tongnanlong is a much larger individual than the 17 m long Y. jiangyiensis.
Tongnanlong differs from Chuanjiesaurus ananensis41 in that the caudal centra are taller than wide while almost equal in C. ananensis. The coracoid fossa is positioned much further away from the glenoid margin of coracoid in C. ananensis than that in Tongnanlong. The scapula-coracoid suture is clear in Tongnanlong but almost fused in C. ananensis. The size of scapula and coracoid are greater in Tongnanlong than that in C. ananensis, which indicates that Tongnanlong is much bigger than C. ananensis.
Tongnanlong differs from Jingiella dongxingensis42 in that the anterior caudal centra are typically procoelous but generally amphicoelous in the latter. Also, the sizes of anterior caudal centra in Tongnanlong are much larger than those in J. dongxingensis. The dorsal neural spines are much higher than wide in Tongnanlong but transversely compressed in J. dongxingensis though this might be caused by their different positions. Complex fominae and laminae above the postzygapophyses of the dorsal vertebrae are well-developed in Tongnanlong but absent in J. dongxingensis. The ventral mid-keel on the anterior caudal centra is present in J. dongxingensis but absent in Tongnanlong.
Moreover, Tongnanlong displays differences from the Middle-Late Jurassic mamenchisaurids that lived in northwestern China, especially the mamenchisaurids from Xinjiang Province, such as Tienshanosaurus chitaiensis, Xinjiangtitan shanshanensis, Klamelisaurus gobiensis, Hudiesaurus sinojapanorum and Mamenchisaurus sinocanadorum.
Tongnanlong differs from Tienshanosaurus chitaiensis43 in that the sizes of dorsal and anterior caudal vertebrae, scapula and coracoid are bigger than those of the latter, which indicates a much larger individual of the former. Due to the limited description of T. chitaiensis in published literature, it is difficult to make further vertebrate comparisons between the two taxa. Even though both display a S-shaped suture of scapula and coracoid, it is much curved dorsally in Tongnanlong but ventrally in T. chitaiensis. The acromial ridge is obtuse, curved and extends dorsally above the glenoid in Tongnanlong but nearly straight and only extends further posteriorly to than the glenoid in T. chitaiensis. Ventrally, the ratio of the lower border width to glenoid surface length in the coracoid is approximately 0.6 in Tongnanlong but nearly 1.2 in T. chitaiensis.
Tongnanlong differs from Xinjiangtitan shanshanensis in many aspects though they both came from the Late Jurassic and have many anatomical similarities44,45. Although both of them bear complex spinopostzygapophysial fossae and laminae structures, the spinopostzygapophysial fossae are much deeper and their number reaches five (two lateral and two medial fosssae separately, and one central fossa) in Tongnanlong but shallow and three fossae (two lateral fossae within a vertical ridge seeing in dorsal 6 and one central fossa seeing in dorsal 3, 6, 9, 11) in X. shanshanensis. The spinopostzygapophysial laminae is much narrower and blade-like in shape in Tongnanlong but broad, ridge-like in shape (seeing dorsal 3, 6, 9, 11) in X. shanshanensis. Also, the dorsal neural spine is much higher dorsally and thinner anteroposteriorly in Tongnanlong than those in X. shanshanensis. Although both pleurocoels on the dorsal centra are deep, there are small vertical laminae dividing the pleurocoels in Tongnanlong but absent in X. shanshanensis.
Tongnanlong differs from the Klamelisaurus gobiensis46 in that the dorsal neural spine is higher than the latter. The shape of the anterior caudal neural spine is blade-like in Tongnanlong but claviform in K. gobiensis. In dorsolateral view, the acromial ridge is located closer to the posterior margin of the proximal portion of the scapula in K. gobiensis than that in Tongnanlong. Regarding the partially reconstructed dorsal junction between the scapula and coracoid, their suture is S-shaped in Tongnanlong but straight in K. gobiensis. The coracoid is irregularly quadrilateral in outline in Tongnanlong but slightly trapezoidal in K. gobiensis. The lateral surface of the coracoid is concave anteroposteriorly in Tongnanlong but gently convex in K. gobiensis. The sizes of the dorsal centra, coracoid, fibula and proximal tibia are much larger in Tongnanlong than those in K. gobiensis, which indicates a much larger individual of the former.
Given the difficulties in identifying the precise position of the isolated dorsal vertebra of Hudiesaurus sinojapanorum47comparisons could only show that the ventral surfaces of the dorsal centra bear no midline ridges in Tongnanlong, while the midline ridge is well-developed in H. sinojapanorum, and the outline of the posterior articular surface of the dorsal centrum is much rounded in H. sinojapanorum but oval in Tongnanlong. Moreover, it is challenging to make comparisons between Tongnanlong and Mamenchisaurus sinocanadorum due to their non-overlapping skeletal parts, as only the lower jaw and cervical series were preserved in the latter48.
Diversity of Mamenchisauridae in the Sichuan Basin
The Jurassic strata in the Sichuan Basin has yielded abundant sauropod dinosaurs since the first dinosaur discovery in 1915, and is now well-known and represented by Jurassic sauropod dinosaurs like Shunosaurus, Omeisaurus and Mamenchisaurus1,8. The Jurassic dinosaurs from the Sichuan Basin are divided into three dinosaur faunae: (1) the Early Jurassic Lufengosaurus Fauna with basal sauropodmorphs predominately represented by Lufengsaurus, (2) the Middle Jurassic Shunosaurus Fauna with sauropods predominately represented by Shunosaurus and Omeisaurus, and (3) the Late Jurassic Mamenchisaurus Fauna with sauropods predominately represented by Mamenchisaurus2. Since the family Mamenchisauridae was established based on the fossils from the Upper Shaximiao Formation in the Sichuan Basin29the validity and evolutionary relationships of this family have experienced several controversies1,]. It was originally considered as a junior synonym of the family Euhelopodidae49then it was abandoned based on the following phylogenetic analyses50,51. The clade including Omeisaurus and Mamenchisaurus from China was assigned to a new family Omeisauridae instead of Mamenchisauridae52. Some studies treated the Mamenchisauridae as a clade of Asian eusauropod taxa from the Middle Jurassic to the earliest Cretaceous8,30,53. Mamenchisauridae is now mainly supported to be a valid clade that includes the Middle-Late Jurassic eusauropods Omeisaurus, Mamenchisaurus and closely related forms6,42,52.
Present studies supports that the clade Mamemchisauridae consists of more than fifteen genera in China (i.e. Tongnanlong, Mamenchisaurus, Omeisaurus, Zigongsaurus, Abrosaurus, Qijianglong, Chuanjiesaurus, Yuanmousaurus, Eomamenchisaurus, Jingiella, Tienshanosaurus, Klamelisaurus, Hudiesaurus, Xinjiangtitan, Huangshanlong and Anhuilong) from the Middle-Late Jurassic strata, which are commonly widespread geographically in southwestern China (Sichuan, Yunnan, Guangxi provinces)8,40,42,54), northwestern China (Gansu and Xinjiang provinces)29,55 and central China (Anhui Province)56. Among these mamenchisaurid taxa, the genera Mamenchisaurus and Omeisaurus have higher species diversities than other taxa. The reported Mamenchisaurus includes six species: M. constructus57 M. hochuanensis, M. youngi30 M. anyuensis9 M. jingyanensis34 M. sinocanadorum58 and the genus Omeisaurus includes eight species: O. junghsiensis59 O. changshouensis60 O. tianfuensis, O. fuxiensis, O. luoquanensis36 O. maoianus61 O. jiaoi62 and O. puxiani63. Although the validity of some species of Omeisaurus and Mamenchisaurus were debated, they still indicate an extremely high mamenchisaurid diversity in China. Moreover, the Late Jurassic stratum in Tanzania has yielded the sauropod dinosaur Wamweraccaudia keranjei, which belongs to Mamenchisauridae4. It suggests that Mamenchisauridae had a wide geographical distribution not only limited to East Asia but also Africa during the Late Jurassic. Mamenchisaurids were also reported from the earliest Cretaceous (controversial era,?Late Jurassic) Phu Kradung Formation in northeastern Thailand which is not far away from the Sichuan Basin in southwestern China42 supporting that the genus Mamenchisaurus existed from the Middle Jurassic till the earliest Cretaceous. It is noteworthy that the sauropod dinosaurs in the Sichuan Basin had attained a high diversity, and the Sichuan Basin was an ideal habitat for sauropod dinosaurs compared to other contemporaneous mainlands in North America, Europe and Africa during the Middle-Late Jurassic period.
Body size of Tongnanlong zhimingi
Tongnanlong preserves complete scapular, coracoid and fibula bones but only partial limbs and vertebrate series, which makes the estimation of body size difficult. However, the length comparisons of scapula, coracoid and fibula with other eusauropods could provide some indications. Length comparisons of scapula and fibula show that Tongnanlong is observably greater than those in Mamenchisaurus and Omeisaurus which had a body length of about 20 m. Based on the ratio of fibula to body length of Mamenchisaurus youngi (ZDM 0083), Mamenchisaurus hochuanensis (ZDM 0126) and Omeisaurus tianfuensis (ZDM 5002), which had nearly complete individuals preserved, the estimated body length of Tongnanlong probably reaches about 23 ~ 26 m, and based on the ratio of scapula length to body length, the estimated body length Tongnanlong could be up to 24.5 ~ 28 m. It is noteworthy that the body length of Tongnanlong could reach 33 m based on the scapular data of Mamenchisaurus jingyanensis, but this data is uncertain for the inaccurate body length of Mamenchisaurus jingyanensis. Mamenchisaurus jingyanensis is known from an incomplete individual without complete vertebrate series and limbs, and its body length of 20 m was estimated by considering it to be similar to Mamenchisaurus hochuanensis (ZDM 0126)34. Herein, we cautiously estimated that Tongnanlong probably achieved a body length of about 25 ~ 26 m, representing a huge-sized mamenchisaurid dinosaur and might be the biggest individual by owning the biggest scapula so far. Some studies suggested that huge-bodied reptiles could reduce death risks during predation and thus gained competitive advantages within the dinosaur fauna42,64,65which might be a driving factor that facilitated the mamenchisaurids to gradually dominate the dinosaur fauna in Late Jurassic.
Conclusion
Mamenchisauridae was distributed globally in the Late Jurassic rather than an endemic fauna which was previously considered limited to East Asia. Tongnanlong zhimingi gen. et sp. nov. from the Upper Jurassic Suining Formation of Chongqing area in the Sichuan Basin is assigned to Mamenchisauridae based on its more complex structures of dorsal and anterior caudal vertebrae compared to those of other mamenchisaurid dinosaurs. The large size of scapula and fibula also implies that it is a gigantic mamenchisaurid individual and might be the biggest one so far. The dorsal vertebrae of Tongnanlong provides additional information to help us understanding the skeletal changes of sauropods in Eastern Asia. The newly reported Tongnanlong enriches the diversity of eusauropods and provides new information on the understanding of the sauropod diversity and evolutionary trend from the Middle Jurassic to the Late Jurassic when their bodies became larger.
Methods
Anatomy
Anatomical descriptions and comparisons in this study followed the Romerian nomenclature and orientation instead of standardized terms (e.g. using anterior/posterior instead of cranial/caudal). Measurements of the specimen were taken using a caliper and photographed with a Nikon D7000 camera.
Phylogenetic analysis
We conducted a phylogenetic analysis to determine the systematic position of Tongnanlong using the data matrix of Ren et al. (2023, 2024)6,42. With the addition of Tongnanlong in the matrix, 87 taxa and 400 characters were analyzed (see in the Supplementary). We first analyzed the data matrix under the “New Technology” search options using sectorial search, ratchet, tree drift, and tree fuse options with default parameters (10000 maximum trees and 1000 random seeds). Then the “Traditional Search” was applied under the tree bisection reconnection branch (TBR) swapping algorithm (10000 random seeds, 10000 replicates, 100 trees saved per replication and 10000 maximum trees, consistency index = 0.335 and retention index = 0.685). Phylogenetic analysis yields 35 most parsimonious trees with 1407 steps. The strict consensus of 35 most parsimonious trees excludes the taxon Wamweraccaudia keranjei as it was actively relocated in different clades of the 35 most parsimonious trees. Phylogenetic analysis was performed using TNT version 1.6 under maximum parsimony66 and character mapping was carried out using Mesquite version 3.8167.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank. The LSIDs of this publication is EF21F164-0723-4F9A-AA6E-A7531CFC35F7 for this publication, 941C1474-8944-4197-B508-E66041A9418B for the genus Tongnanlong, and 25ED5383-9D83-4633-93F2-9F9727B14BDA for the species Tongnanlong zhimingi.
Data availability
Data is provided online freely within the supplementary information files and available on request. Correspondence and requests for supplementary materials should be addressed to X.W.
Abbreviations
- TNM:
-
Tongnan Museum
- ZDM:
-
Zigong Dinosaur Museum
- CQMNH:
-
Chongqing Museum of Natural History
- IVPP:
-
Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences
References
Upchurch, P., Barrett, P. M. & Dodson, P. Sauropoda in The Dinosauria (Second edition) (eds. Weishampel, D. B., Dodson, P., Osmolska, H.) 259–322. (University of California Press, 2004).
Dong, Z. M., You, H. L. & Peng, G. Z. Palaeovertebrata Sinica, Volume II. Amphibians, Reptilians, and Avians, Fascicle 5 (Serial No. 9): Ornithischian Dinosaurs (Science Press, 2015).
Xu, X., Clark, J. M., Eberth, D. A. & Currie, P. J. The Shishugou Fauna of the Middle-Late Jurassic transition period in the Junggar Basin of western China. Acta Geol. Sin (Eng Ed). 96, 1115–1135. https://doi.org/10.1111/1755-6724.14996 (2022).
Mannion, P. D., Upchurch, P., Schwarz, D. A. & Wings, O. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zool. J. Linn. Soc. 9, 784–909. https://doi.org/10.1093/zoolinnean/zly068 (2019).
Zhou, Z. H., Barrett, P. M. & Hilton, J. An exceptionally preserved Lower Cretaceous ecosystem. Nature 421, 807–814. https://doi.org/10.1038/nature01420 (2003).
Ren, X. X. et al. Re-examination of Dashanpusaurus Dongi (Sauropoda: Macronaria) supports an early Middle Jurassic global distribution of neosauropod dinosaurs. Palaeogeogr Palaeoclimatol Palaeoecol. 610, 1–11. https://doi.org/10.2139/ssrn.4150550 (2023).
Xu, X. et al. A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs. Nat. Commun. 9, 1–9. https://doi.org/10.1038/s41467-018-05128-1 (2018).
Peng, G. Z., Ye, Y., Gao, Y. H., Shu, C. K. & Jiang, S. Jurassic Dinosaur Faunas in Zigong (Sichuan People’s Publishing House, 2005).
He, X. L., Yang, S. H., Cai, K. J., Li, K. & Liu, Z. W. A new species of sauropod, Mamenchisaurus anyuensis sp. nov. In Proceeding of 30th International Geology Congress, 83–86 (China Economy Press, 1996).
Xing, L. D. et al. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. J. Vert Paleontol. 35 e889701 https://doi.org/10.1080/02724634.2014.889701 (2015).
Luo, L., Qi, J., Zhang, M. Z., Wang, K. & Han, Y. Z. Detrital zircon U–Pb ages of Late Triassic-Late Jurassic deposits in the western and northern Sichuan Basin margin: constraints on the foreland basin provenance and tectonic implications. Inter J. Earth Sci. : Geol. Rundschau. 103, 1553–1568. https://doi.org/10.1007/s00531-014-1032-7 (2014).
Shao, T. B., Cheng, N. F. & Song, M. S. Provenance and tectonic-paleogeographic evolution: constraints from detrital zircon U–Pb ages of Late Triassic–Early Jurassic deposits in the northern Sichuan Basin, central China. J. Asian Earth Sci. 127, 12–31. https://doi.org10.1016/j.jseaes.05.027 (2016). (2016).
Liu, G. et al. Detrital zircon U–Pb dating of Suining Formation sandstone from the Daba mountains, northeastern Sichuan and its stratigraphic implications. Palaeoworld 26, 380–395. https://doi.org/10.1016/j.palwor.2017.03.002 (2017).
Ye, Y., Peng, G. Z., Jiang, S., Hao, B. Q. & Xing, L. D. Dinosaur Tracks in Sichuan Basin (Geological Publishing House, 2022).
Huang, D. Y. Jurassic integrative stratigraphy and timescale of China. Sci. China Earth Sci. 62, 223–255. https://doi.org/10.1360/N072017-00443 (2019).
Huang, D. Y., Fang, Y. N., Li, J. G. & Li, X. Lithostratigraphic subdivision and correlation of the Jurassic in China. J. Stratigr. 45, 364–374. https://doi.org/10.19839/j.cnki.dcxzz (2021).
Zeng, Q., Liu, H. Q., Yuan, Z. P. & Yang, J. H. The occurrence characteristics, burial environment and protection status and suggestions of Anyue dinosaur fossils. Sich Nonf Met. 2, 9–13 (2020).
Wang, J., Norell, M. A., Pei, R., Ye, Y. & Chang, S. C. Surprisingly young age for the mamenchisaurid sauropods in south China. Cret Res. 104, 104176. https://doi.org/10.1016/j.cretres.2019.07.006 (2019).
Wang, Q. W., Kan, Z. Z., Liu, X. H., Liang, B. & Zhu, B. The Mesozoic sporopollen assemblage in the Sichuan Basin and its significance to paleovegetation and paleoclimate. Acat Geol. Sichuan. 28 (2), 89–95 (2008).
Chen, X. L. et al. Flood-overlake sedimentary characteristics of the Suining Formation (Lower Jurassic) in western Sichuan depression. Acta Sedimentol. Sin. 32, 912–920 (2014).
Gao, H. C., Zheng, R. C., Ke, G. M. & Wen, H. G. Characteristics of sedimentary facies of the Suining Formation of Upper Jurassic in western Sichuan Basin. J. Palaeogeol. 8, 467–476 (2006).
Kan, Z. Z., Liang, B., Wang, Q. W. & Zhu, B. Trace fossils in the Suining and Penglaizhen formations as buried places of dinosaur fossils, in Anyue, Sichuan and their environmental significance. Acata Geol. Sichuan. 25, 68–71 (2005).
Li, Z. G. Quantitative analysis of Mamenchisaurus anyuensis. Master thesis, Chengdu University of Technology (2013).
Xia, Z. S., Yuan, C. M. & Li, R. N. Mesozoic Stratigraphy and Palaeology of Sichuan Basin (Sichuan People’s Publishing House, 1982).
Seeley, H. G. On the classification of the fossil animals commonly named Dinosauria. Proc. R. Soc. London 43, 165–171 (1887).
von Huene, F. Die fossile reptil-ordnung Saurischia: ihreentwicklung und geschichte. Gebrüder Borntraeger. Monogr. Geol. Paläontol Ser. 1 (4), 1–361 (1932).
Marsh, O. C. Principal characters of American Jurassic dinosaurs (Part 1). Amer J. Sci. Ser. 3. 16, 411–416 (1878).
Upchurch, P. The evolutionary history of sauropod dinosaurs. Philos. Trans. R Soc. Lond. Ser. B. 349, 365–390. https://doi.org/10.1098/rstb.1995.0125 (1995).
Young, C. C. & Zhao, X. J. Mamenchisaurus hochuanensis. (ed. Institute of Vertebrate Paleontology and Paleoanthropology Monograph, Series I). (Science Press, 1972).
Ouyang, H. & Ye, Y. The First Mamenchisaurian Skeleton with Complete Skull: Mamenchisaurus youngi. (Sichuan Publishing House of Science and Technology, 2002).
Ren, X. X., Sekiya, T., Wang, T., Yang, Z. W. & You, H. L. A revision of the referred specimen of Chuanjiesaurus anaensis Fang et al., 2000: a new early branching mamenchisaurid sauropod from the Middle Jurassic of China. Hist. Biol. 33, 1872–1887. https://doi.org/10.1080/08912963.2020.1747450 (2021).
Zhang, Y. H. The Middle Jurassic Dinosaur Fauna from Dashanpu of Zigong, Sichuan (Vol. 3), Sauropoda: Shunosaurus lii (Sichuan Pubublishing House of Science and Technology, 1988).
Moore, A. J., Mo, J. Y., Clark, J. M. & Xu, X. Cranial anatomy of Bellusaurus Sui (Dinosauria: Eusauropoda) from the Middle-Late Jurassic Shishugou Formation of northwest China and a review of sauropod cranial ontogeny. PeerJ 6, e4881. https://doi.org/10.7717/peerj.4881 (2019).
Zhang, Y. H., Li, K. & Zeng, Q. H. A new species of sauropod from the Late Jurassic of the Sichuan Basin (Mamenchisaurus jingyanensis sp. nov). J. Chengdu Univ. Tech. 25, 61–68 (1998).
Lü, J. C. et al. Osteology of the Giant Sauropod Dinosaur Ruyangosaurus giganteus Lü et al., 2009 (Geological Publishing House, 2014).
He, X. L., Li, C. & Cai, K. J. The Middle Jurassic Dinosaur Fauna from Dashanpu, Zigong, Sichuan (Vol. 4), Sauropod Dinosaurs (2), Omeisaurus tianfuensis (Sichuan Publishing House of Science and Technology, 1988).
Ye, Y., Ouyang, H. & Fu, Q. M. New material of Mamenchisaurus hochuanensis from Zigong. Vert PalAs. 1, 266–271 (2001).
Hou, L. H., Zhou, S. W. & Cao, Y. S. New sauropod dinosaurs from Sichuan. Vert PalAs. 14, 160–165 (1976).
Dong, Z. M., Zhou, S. W. & Zhang, Y. Z. The Jurassic Dinosaurs of the Sichuan Basin, Chinese Palaeontologica Sinica Series C (Science Press, 1983).
Lü, J. C. et al. New eusauropod dinosaur from Yuanmou of Yunnan Province, China. Acta Geol. Sin (Eng Ed). 80, 1–10 (2006).
Sekiya, T. Re-examination of Chuanjiesaurus anaensis (Dinosauria: Sauropoda) from the Middle Jurassic Chuanjie Formation, Lufeng County, Yunnan Province, southwest China. Mem. Fukui Prefect Dinosaur Mus. 10, 1–54 (2011).
Ren, X. X., Wang, X. R., Ji, Y. N., Guo, Z. & Ji, Q. The first mamenchisaurid from the Upper Jurassic Dongxing Formation of Guangxi, southernmost China. Hist. Biol. https://doi.org/10.1080/08912963.2024.2309287 (2024).
Young, C. C. A new dinosaurian from Sinkiang. Pala. Sin. New ser. C, 2. Whole Ser. No. 105, 1–29 (1937).
Zhang, X. Q., Li, N., Xie, Y., Li, D. Q. & You, H. L. Redescription of the dorsal vertebrae of the mamenchisaurid sauropod Xinjiangtitan Shanshanesis Wu et al. 2013. Hist. Biol. 36, 49–75 (2022).
Wu, W. H., Zhou, C. F., Oliver, W., Sekiya, T. & Dong, Z. M. A new gigantic sauropod dinosaur from the Middle Jurassic of Shanshan, Xinjiang. Glob Geol. 32, 437–446. https://doi.org/10.3969/j.issn.1004-5589.2013.03.002 (2013).
Moore, A. J. et al. Osteology of Klamelisaurus gobiensis (Dinosauria, Eusauropoda) and the evolutionary history of Middle–Late Jurassic Chinese sauropods. J. System Paleontol. 18, 1299–1393. https://doi.org/10.1080/14772019.2020.1759706 (2020).
Dong, Z. M. A gigantic sauropod (Hudiesaurus sinojapanorum gen. et sp. nov.) from the Turpan Basin, China. In Dong, Z. M. (edit.) Sino–Japanese Silk Road Dinosaur Expedition. 102–110 (China Ocean Press, 1997).
Moore, A. J. et al. Re-assessment of the Late Jurassic eusauropod Mamenchisaurus sinocanadorum Russell and zheng, 1993, and the evolution of exceptionally long necks in mamenchisaurids. J. Sys Palaeont. 21 (1), 2171818. https://doi.org/10.1080/14772019.2023.2171818 (2023).
Romer, A. S. Osteology of the Reptiles (University of Chicago Press, 1956).
Canudo, J. I., Ruiz-Omeñaca, J. I., Barco, J. L. & Royo-Torres, R. Saurópodos asiáticos en el Barremiense inferior (Cretácico Inferior) de España? Ameghiniana 39, 443–452 (2002).
Wilson, J. A. & Upchurch, P. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. J. Syst. Palaeontol. 7, 199–239. https://doi.org/10.1017/S1477201908002691 (2009).
Wilson, J. A. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zool. J. Linn. Soc. 136, 217–276. https://doi.org/10.1046/j.1096-3642.2002.00029.x (2002).
Suteethorn, S., Le Loeuff, J., Buffetaut, E., Suteethorn, V. & Wongko, K. First evidence of a mamenchisaurid dinosaur from the Upper Jurassic–Lower Cretaceous Phu Kradung Formation of Thailand. Act. Palaeon. Pol. 58, 459–469 (2009). https://doi.org/10.4202/app. 0155. (2013).
Lü, J. C., Li, T. G., Zhong, S. M., Ji, Q. & Li, S. X. A new mamenchisaurid dinosaur from the Middle Jurassic of Yuanmou, Yunnan province, China. Acta Geol. Sin (Eng Ed). 82, 17–26 (2008).
Xu, X., You, H. L. & Mo, J. Y. Palaeovertebrata Sinica, Volume II. Amphibians, Reptilians, and Avians, Fascicle 6 (Serial No. 10): Saurischian Dinosaurs. (Science Press, 2022).
Ren, X. X., Huang, J. D. & You, H. L. The second mamenchisaurid dinosaur from the Middle Jurassic of eastern China. Hist. Biol. (32), 602–610. https://doi.org/10.1080/08912963.2018.1515935 (2020).
Young, C. C. On a new sauropod from Yiping, Szechuan, China. Acta Palaeontol. Sin. 3, 491–505 (1954).
Russell, D. A. & Zheng, Z. A large mamenchisaurid from the Junggar Basin, Xinjiang, People’s Republic of China. Can. J. Earth Sci. 30, 2082–2095. https://doi.org/10.1139/e93-180 (1993).
Young, C. C. On a new sauropoda, with notes on other fragmentary reptiles from Szechuan. Bull. Geol. Soc. China. 19, 279–316 (1939).
Young, C. C. New sauropods from China. Vert PalAs. 2, 1–28 (1958).
Tang, F., Jin, X. S., Kang, X. M. & Zhang, G. J. A Complete Sauropod Dinosaur from Jingyan, Sichuan: Omeisaurus maoianus (China Ocean Press, 2001).
Jiang, S., Li, F., Peng, G. Z. & Ye, Y. A new species of Omeisaurus from the Middle Jurassic of Zigong, Sichuan. Veterbr PalAs. 49, 185–194. https://doi.org/10.1631/jzus.B1000197 (2011).
Tan, C. et al. A new species of Omeisaurus (Dinosauria: Sauropoda) from the Middle Jurassic of Yunyang, Chongqing, China. Hist. Biol. 33, 1817–1829. https://doi.org/10.1080/08912963.2020.1743286 (2021).
Kanayama, Y. & Iwasa, Y. Why did sauropod dinosaurs grow so big? A possible answer from the life history theory. J. Theor. Biol. 508, 110485. https://doi.org/10.1016/j.jtbi.2020.110485 (2021).
Bates, K. T. et al. Temporal and phylogenetic evolution of the sauropod dinosaur body plan. R Soc. Open. Sci. 3, 150636. https://doi.org/10.1098/rsos.150636 (2016).
Golobof, P. A. & Morales, M. E. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39, 144–153. https://doi.org/10.1111/cla.12524 (2023).
Mesquite. A modular system for evolutionary analysis version 3.81 (2023). http://www.mesquiteproject.org
Acknowledgements
We thank Dr. Xinxin Ren for his thorough reviews that largely improved the manuscript. We also appreciate the Dr. Xuanyu Zhou, whose constructive comments have greatly improved an earlier version of this manuscript. We also give thanks to Guangming Ren, Weihua Pang, Xiuping Wang and Xiao Mei who give much conveniences for preparing the specimen.
Author information
Authors and Affiliations
Contributions
X.W described the specimen and wrote the main manuscript. Y.T, J.D and L.L prepared Figs. 1, 2, 3, 4 and 5 and tables. X.W, Y.L and Y. L measured the specimen. G.W, D.L, G.P and S.Z prepared figure 6 and supplementary information. C.L refined the MS, checked and polished the English expressions, and gave constructive suggestions on dealing with the reviewers’ opinions. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wei, X., Tan, Y., Jiang, S. et al. A new mamenchisaurid from the Upper Jurassic Suining Formation of the Sichuan Basin in China and its implication on sauropod gigantism. Sci Rep 15, 24808 (2025). https://doi.org/10.1038/s41598-025-09796-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-09796-0
