
Abstract
Many survivors of childhood cancer experience significant side effects leading to inferior quality of life like chemotherapy-induced cognitive impairment (CICI). A growing body of literature implies that the gut-brain axis modulates neurological and inflammatory physiology. Cancer therapy may alter the gut microbiome in ways that modify the risk of CICI. We utilized a commonly used pediatric cancer chemotherapy drug, doxorubicin (DOXO), in our juvenile rat model, including both male and female rats. At 5 weeks of age, Long Evans rats were treated with weekly intraperitoneal DOXO (2 mg/kg/dose for 4 weeks). Compared to PBS-treated rats, DOXO treatment caused impairment of visual memory among female rats and spatial memory among male rats, when tested 3–4 weeks after DOXO exposure. In both sexes, DOXO-induced cognitive deficits do not persist among the surviving animals. Microbiome analyses were conducted in fecal samples collected 48 hours following the final DOXO injection. Compared to PBS-treated rats, female DOXO-treated rats had significant differences in beta diversity. Significant changes in the abundances of specific species, genera, and phyla were observed. These experimental results set the stage for further investigations of how chemotherapy-induced changes in gut microbiome might contribute to CICI, and point toward therapeutic interventions.
Similar content being viewed by others
Introduction
Anthracyclines (including daunomycin and doxorubicin [DOXO]) are used in more than 60% of curative regimens for children, adolescents, and young adults diagnosed with cancer1. Although effective as a cancer drug, DOXO induces neurotoxicity despite its inability to cross the blood-brain barrier (BBB)2. Chemotherapy-induced cognitive impairment (CICI) significantly impacts the quality of life for pediatric cancer survivors3,4and includes measurable deficits in executive functioning, working memory, information processing speed, calculations, multitasking, fine motor skills, and concentration5,6. In the United States alone, there are approximately 500,000 childhood cancer survivors, and from this subset, at least 35% will exhibit both short and long-term neurocognitive impairments directly related to the treatment they received7. Not only does CICI impact the quality of life for children surviving cancer, but also that of the parents and families of the children and emphasizing the need for psychosocial aftercare8. A significant underlying factor that plays a role in poorer neurocognitive outcomes is the age of treatment, where young age is more susceptible9. Further investigations of the impact of DOXO on cognition are necessary to better understand the pathophysiology and to suggest interventions to mitigate or prevent CICI.
Multiple in vivo studies indicate that DOXO induces cognitive dysfunction, along with increased neurodegeneration, decreased neurogenesis, and increased pro-inflammatory markers10,11,12,13,14. However, many of these preclinical models for DOXO-induced cognitive impairment do not accurately reflect pediatric dosing used clinically, use animals of only one sex, and do not test the persistence of observed deficits. Despite being extensively studied for the past several decades, there is no clearly defined mechanism for DOXO-induced cognitive impairment, making it challenging to mitigate these neurotoxic effects.
Recent literature suggests the gut-brain axis plays a critical role in modulating chemotherapy-induced neurotoxic effects15,16,17. Paclitaxel treatment induced cognitive impairment in mice, reduced microglial immunoreactivity, and increased pro-inflammatory cytokines/chemokines in the brain; additionally, paclitaxel treatment impacted the bacterial taxa, which may modulate the neuropathology18. Similarly, methotrexate-induced neurotoxicity has been associated with gut microbiota changes in a rat model, implying a critical role of the gut-brain axis in the pathogenesis of MTX-induced neurotoxicity19. Although these preclinical studies do not explore DOXO, they offer insights into how chemotherapy can broadly modulate the gut-brain axis.
Clinical studies also exemplify the role of the gut-brain axis in patients treated with chemotherapy. Specifically, breast cancer patients treated with chemotherapies, including DOXO, had altered beta diversity measures after chemotherapy treatment and increased TNF-α levels20. Further, the cognition of breast cancer patients was monitored throughout the chemotherapy treatment, and researchers made a few associations between gut microbiome changes and cognitive decline20. While many studies report the relevance of microbiome changes related to DOXO-induced cardiovascular toxicity, there are limited in vivo models and even fewer pediatric models investigating the microbiome implications in DOXO-induced neurotoxicity.
Here, we investigated whether DOXO induces cognitive dysfunction in a juvenile rat model, including both males and females, and whether these cognitive deficits will persist into adulthood. We further explored the impact of DOXO treatment on the gut microbiome and identified alterations that possibly contribute to cognitive dysfunction.
Methods
Animal model and reagents
Long Evans rats (16 males and 16 females) were purchased from Charles River Laboratories (Wilmington, Massachusetts) at 4 weeks of age and were allowed to acclimate to the vivarium environment and handling for 1-1.5 weeks prior to any experiments. The rats were housed in same-sex groups of two or three rats per cage with a 12/12 hours light/dark cycle and ad libitum access to food (LabDiet 5053) and water. At 5 weeks of age, the rats were injected intraperitoneally with DOXO (2 mg/kg/dose once weekly for 4 weeks) (Doxorubicin HCl 2 mg/mL, Fresenius Kabi, LLC. – 63323010161 purchased from Henry Schein, Catalog No. 1164027) or equal volume of saline (control) (1X PBS (Phosphate Buffered Saline pH 7.4, Gibco, Catalog No. 10010049)). This regimen is approximately equivalent to 25 mg/m2 per course21,which is typical dosing for some pediatric cancer regimens2. All injected solutions were sterilized using 0.22 µM syringe filters (VWR, Catalog No. 76479-016). The weights of the rats were monitored throughout the treatment period and during the post-treatment period. All experiments were approved by the Animal Institute and Use Committee of Rutgers University and were conducted following the “Guide for the Care and Use of Laboratory Animals.” The ARRIVE guidelines were followed for all experiments described here. All methods were conducted in accordance with the relevant guidelines and regulations.
Microbiome collection and analysis
Fecal samples were collected 48-hours after the final DOXO injection by placing each rat into a sterile cage to collect a pea-sized fecal sample and it was placed directly into the barcoded tubes with DNA stabilization buffer provided by Transnetyx (Cordova, TN, USA). The fecal samples stored in the stabilization buffer were shipped to Transnetyx to conduct the DNA extraction, library preparation, and sequencing. DNA extraction was fully automated using the Qiagen DNeasy 96 PowerSoil Pro QIAcube HT extraction kit and protocol for the inhibitor-free extraction of high molecular weight genomic DNA. Quality control was completed, followed by conversion of the genomic DNA into sequencing libraries. Unique dual indexed (UDI) adapters were utilized. The KAPA HyperPlus library preparation was used for the library preparation. Libraries were sequenced using shotgun sequencing with a depth of 2 million 2 × 150 bp read pairs using Illumina NextSeq 2000. All sequencing data was uploaded by Transnetyx onto One Codex software, and it was analyzed against a database with microbial reference genomes. First, a K-mer based classification was applied, where every sequence was compared against the database by exact alignment (k = 31). Next, artifact automatic filtering was performed based on the relative frequency of unique k-mers in the sample and any artifacts from sequencing were filtered out. Last, the species level abundance estimation is based on the sequencing depth and coverage across every reference genome. All samples had more than 1,000,000 reads (range: 1,206,306-2,822,548 reads). Analysis was conducted using built-in Jupyter notebooks provided on One CodeX portal (https://www.onecodex.com/tags/notebooks/). For the analysis, we utilized abundance estimates that were generated by One CodeX. Subsequently, we performed a multiplicative replacement, followed by the Analysis of Compositions of Microbiomes (ANCOM) test22 on Jupyter notebooks.
Alpha diversity analysis was performed to determine the species richness and evenness, using both Shannon Index and Observed Taxa. Beta diversity analysis was performed to determine the differences between samples and statistical significance was calculated using the permutational multivariate analysis of variance (PERMANOVA). The results were visualized using Principal Coordinates Analysis (PCoA) with the Bray-Curtis matrix. The Benjamini-Hochberg false discovery rate (FDR) was utilized to adjust for the beta diversity. To identify the differentially abundant taxa between the groups, we utilized the ANCOM test to identify the species (or other taxonomic ranks) that were significantly different between the groups of interest (PBS vs. DOXO, PBS Female vs. DOXO Female, PBS Male vs. DOXO Male). Subsequently, for the taxa identified as significantly different by the ANCOM test, we assessed the normality within each group by the Shapiro-Wilk test. The statistical tests used for each panel are specified in the legend. GraphPad Prism 10.4 (GraphPad Software, San Diego, CA) was utilized to perform the Shapiro-Wilk normality test, followed by either an unpaired t-test (normally distributed data) or Mann Whitney test (non-normally distributed data). Any ANCOM results that calculated a W = 0 was not considered significant even if the Unpaired t-test or Mann Whitney test showed significance. Results are considered statistically significant when the p-value ≤ 0.05.
We used the One Codex functional analysis pipeline to identify metabolic pathways in each sample. Subsequently, an ANCOM test was performed with a Holm-Bonferroni multiple comparisons correction, to identify differences between groups in pathway abundances unstratified by taxa. Males and females were analyzed both separately and combined by sex or by treatment.
Behavioral testing
Behavioral testing was conducted at 2 timepoints to assess acute effects (10–12 weeks of age; 2–4 weeks after the last DOXO injection) and long-term effects (28–30 weeks of age; 5 months after the last DOXO injection) of DOXO treatment at a juvenile age. The open field (OF) test was conducted to assess the locomotion for the rats after DOXO and PBS injections and to confirm that DOXO does not induce locomotor deficits. This test was conducted in an arena (69 × 69 × 60 cm) with visual cues for 6 min, and the total track length, center track length, center time, and center entries were recorded.
To assess the cognitive functioning of the rat model, we utilized the object placement (OP) and object recognition (OR), as previously published by our laboratory23,24 with slight modifications, and conducted these tests in the same apparatus used for the OF test. All the objects used for these tests have been validated prior to testing. Our first detection method for cognitive dysfunction consisted of the novel OR test, which tests the visual and recognition memory25 and is a hippocampus-independent memory test26. In the OR test, the testing interval for the rat was 120 min, and the test was performed as previously published23,24. Our second detection method was the OP test, which tests the spatial memory and is a hippocampus-dependent memory test26,27. In the OP test, the testing interval for the rat was 20 min, and the test was performed as previously published23,24. The time the rat spent with each object was recorded. Exploration for the OP and OR tests was defined as the rat having physical contact with the object, such as sniffing, whisking, or touching the object. The preference score is the ratio of the time spent exploring the novel (or relocated) object to the total time exploring both objects, and that is expressed as a percentage23. As previously published, for both OP and OR, cognitive functioning was assessed using a preference score threshold of 55%, where percentages above this threshold indicate intact memory23.
Statistical analysis was performed to compare the different groups through GraphPad Prism 10.4 (GraphPad Software, San Diego, CA) (https://www.graphpad.com). The Shapiro-Wilk test was used to test for normality within each group. A one sample t test (parametric; normally distributed data) or Wilcoxon signed-rank test (nonparametric; non-normally distributed data) with hypothetical mean of 50 was used to determine whether the PBS-treated control rats displayed a preference score higher than chance, which indicates intact memory. The contingency was performed using the Fisher’s exact test two-sided and the number of rats, however the graph is a representation of the percentage of rats with the significance displayed from the number of rats.
Ethics approval
All animal studies were performed in compliance with the Animal Institute and Use Committee of Rutgers University and were conducted following the “Guide for the Care and Use of Laboratory Animals.”
Results
DOXO induces cognitive dysfunction in juvenile rats
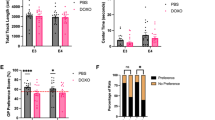
Male and female juvenile rats were treated with a clinically relevant DOXO regimen. DOXO-treated male rats displayed significant weight loss starting at 14 weeks of age, compared to the PBS-treated male rats (Fig. 1a). Behavioral testing was conducted to evaluate the impact of DOXO on cognitive function. In the open field (OF) test, there was no significant effect of DOXO on total track length (Fig. 1b) or center time (Fig. 1c), indicating no locomotor deficits or anxiety-like behavior. Object placement (OP) test results showed that the PBS-treated female and male rats received a mean OP preference score significantly higher than chance, suggesting intact cognition (Fig. 1d; One sample t test: PBS female, t = 2.232, df = 7, p = 0.0608; PBS male, t = 4.179, df = 6, p = 0.0058). DOXO-treated male rats exhibited spatial memory deficits (Fig. 1e; PBS males compared to DOXO males: Fisher’s exact test, p = 0.0291). Lastly, we conducted the object recognition (OR) test of visual memory. PBS-treated female and male rats demonstrated intact visual memory, with mean OR preference scores significantly higher than chance, while DOXO-treated rats did not (Fig. 1f; Wilcoxon signed rank test: PBS female, sum of signed ranks (W) = 32, p = 0.0234; PBS male, sum of signed ranks (W) = 32, p = 0.0234). DOXO-treated female rats were more likely to exhibit visual memory deficits (Fig. 1g; PBS females compared to DOXO females: Fisher’s exact test, p = 0.0070). Our results suggest that DOXO treatment induces visual memory deficits in female rats and spatial memory deficits in male rats.

DOXO induces cognitive dysfunction in juvenile Long Evans rats. (a) Growth curve shows the weights of rats in grams (g) during and after DOXO treatment. The grey shaded region represents the period of DOXO treatment, starting at 5 weeks of age, the rats were treated with 2 mg/kg/week for 4 weeks. The groups are separated by sex and treatment. Two weeks after the last DOXO dose, open field test was conducted on the rats to assess for locomotion: (b) bar graph shows the total track length in centimeters (cm) for each rat and (c) bar graph shows the time each rat spent in the center. (d) Bar graph represents the object placement (OP) preference score percentage for each group. (e) Stacked bar graph represents the percentage of the total number of rats for that group that either performed above the threshold and displayed a preference or that performed below and displayed no preference. (f) Bar graph represents the object recognition (OR) preference score percentage for each group. For both panels d and f, the red dotted line represents the 55% threshold. (g) Stacked bar graph represents the percentage of the total number of rats for that group that either performed above the threshold and received a preference or that performed below and received no preference. For all panels, the groups are separated by sex and treatment (n=7-8 per group) with biological replicates per group shown as a circle on the graph. Data represent mean ± SEM, biological replicates per group are displayed as a circle on the graph. Statistical analysis was performed using: 2way ANOVA with Tukey’s multiple comparisons test (a), one-way ANOVA with Tukey’s multiple comparisons test (b, c), One sample t test with hypothetical mean at 50 (d), Contingency Fisher’s exact test utilized on number of animals per group and the significance displayed here is based off the number of rats per group that received a preference or no preference for each sex was conducted individually (e, g), Wilcoxon signed-rank test with hypothetical mean at 50 (f). *p < 0.05, **p < 0.01, n.s.: not significant.

DOXO-induced cognitive dysfunction in juvenile rats does not persist 5 months after the last dose. (a) Survival curve shows the percentage of rats that survived during and after DOXO treatment over 5 to 28 weeks of age. The grey shaded region represents the period of DOXO treatment. The groups are separated by sex and treatment. Five months after the last DOXO dose, open field test was conducted on the rats to assess for locomotion: (b) bar graph shows the total track length in centimeters (cm) for each rat and (c) bar graph shows the time each rat spent in the center. (d) Bar graph represents the object placement (OP) preference score percentage for each group. (e) Stacked bar graph represents the percentage of the total number of rats for that group that either performed above the threshold and displayed a preference or that performed below and displayed no preference. (f) Bar graph represents the object recognition (OR) preference score percentage for each group. For both panels d and f, the red dotted line represents the 55% threshold. (g) Stacked bar graph represents the percentage of the total number of rats for that group that either performed above the threshold and received a preference or that performed below and received no preference. For all panels, the groups are separated by sex and treatment (n=4-8 per group) with biological replicates per group displayed as a circle on the graph. Data represent mean ± SEM. Statistical analysis was performed using: One sample t test with hypothetical mean at 50 (d, f). *p < 0.05, **p < 0.01.
DOXO-induced visual and Spatial memory deficits do not persist into adulthood
Few to no in vivo studies investigate the long-term impact of DOXO-induced cognitive deficits in a pediatric model. Patients may experience symptoms from months to years after chemotherapy treatment; therefore, establishing a model that recapitulates long-term deficits to better understand the neuro-pathophysiology of CICI is essential. We previously demonstrated that the chemotherapeutic drug, methotrexate, induced cognitive deficits in juvenile rats that persisted well into adulthood, more than 12 months after drug exposure24. To assess the long-term effects of DOXO treatment, we monitored survival and assessed cognitive function. Interestingly, our results showed that 50% of the DOXO-treated male rats died within 3 months post-treatment (Fig. 2a). These results highlight that DOXO treatment increases toxicity for male rats, indicating a sex-specific response. At five months after DOXO or PBS treatment, there were no significant differences between the groups in total track length and center time (Fig. 2b and c, respectively). Additionally, there were no significant differences in the OP (Fig. 2d and e) and OR (Fig. 2f and g) testing, indicating that cognitive deficits do not persist 14 weeks after the final DOXO dose. Interestingly, at 29–30 weeks of age, the DOXO-treated male rats failed to attain a preference score significantly higher than chance for both spatial memory (Fig. 2 d; One sample t test: PBS female, t = 4.356, df = 7, p = 0.0033; DOXO female, t = 2.510, df = 7, p = 0.0404; PBS male, t = 4.685, df = 6, p = 0.0034) and visual memory (Fig. 2f; One sample t test: PBS female, t = 4.010, df = 7, p = 0.0051; DOXO female, t = 2.735, df = 7, p = 0.0291; PBS male, t = 2.790, df = 7, p = 0.0269) tests. Our results show that DOXO-induced cognitive impairment does not persist into adulthood like some chemotherapies.
DOXO treatment induces changes in the gut Microbiome
Given the increasing literature suggesting an important role of the gut-brain axis for cognition28we next explored how DOXO treatment impacts the gut microbiome 48 h following the last DOXO dose. There were no significant differences between DOXO-treated and PBS-treated rats in alpha diversity metrics (Shannon Index and Observed Taxa; Supplementary Fig. 1a and 1b, respectively). Next, we used the beta diversity to show the differences between samples and visualized our results using a Principal Coordinates Analysis (PCoA) plot. We observed partial clustering of DOXO-treated rats from PBS-treated rats in the PCoA plot; however, no significant differences were found in the beta diversity (Supplementary Fig. 1c). Next, we identified the specific taxa across the taxonomic ranks that are significantly impacted by DOXO treatment, and we found differences only at the genus level when the data for the males and females were combined (Supplementary Fig. 1 d). Specifically, we found a significant decrease in the mean relative abundance of Muribaculum (Supplementary Fig. 1e, ANCOM: W = 15) and Paramuribaculum (Supplementary Fig. 1f, ANCOM: W = 2). Further, we found a significant increase in the mean relative abundance of the following: Parabacteroides (Supplementary Fig. 1 g, ANCOM: W = 3), Dubosiella (Supplementary Fig. 1 h, ANCOM: W = 2), and Lachnotalea (Supplementary Fig. 1i, ANCOM: W = 1). Our results indicate that the gut microbiome is impacted by DOXO treatment.
Given that we have previously highlighted sex differences in the response to DOXO, we separated the data by sex. We observed no significant differences in the Shannon Index (Fig. 3a) and Observed Taxa (Fig. 3b), indicating no differences in the alpha diversity. We found that our pediatric DOXO regimen induced significant changes in the beta diversity in DOXO-treated female rats only, indicating that the bacterial species are altered after DOXO treatment (Fig. 3c; PERMANOVA: p = 0.008, FDR = 0.016). In the PCoA plot, PC1 explains 47.6% of the variance between all samples, whereas PC2 explains a further 14.8% of the variance (Fig. 3c). Our results indicate that there are significant microbiome differences induced by DOXO treatment in the female rats. Next, we delved deeper into identifying the specific species that are impacted by DOXO treatment (Fig. 3d). In female rats, the mean relative abundance of species Duncaniella MGBC103508 was increased after DOXO treatment (Fig. 3e, ANCOM: W = 378). In male rats, the mean relative abundance of species Porphyromonadaceae bacterium UBA7183 was significantly increased after DOXO treatment (Fig. 3f, ANCOM: W = 96). Further, we analyzed the differential abundance across all the taxonomic ranks and found significant differences at the order, class, and phylum ranks for the DOXO-treated female rats compared to the PBS-treated female rats. To capture the broader taxonomic level trends, we focused on the phylum level changes induced by DOXO treatment in the female rats (Fig. 3g). We found a significant increase in the mean relative abundance of Actinobacteria (Fig. 3h, ANCOM: W = 3), and a decrease in the mean relative abundance of Bacteroidetes (Fig. 3i, ANCOM: W = 1) in the DOXO-treated female rats compared to the PBS-treated male rats. No changes in these phyla were found in the male rats. To explore the functional implications of these taxonomic shifts in the microbial communities induced by DOXO treatment in female rats, we performed pathway analysis of the predicted metabolic pathways. Interestingly, we identified a few pathways associated with DOXO treatment, including biotin biosynthesis I (ANCOM: W = 111), fatty acid elongation (ANCOM: W = 124), and C4 photosynthetic carbon assimilation cycle (ANCOM: W = 132) (Fig. 3j). Altogether, our results suggest that female rats were more likely to exhibit changes in the gut microbiome after DOXO treatment. While the relationship between the microbiome composition and cognitive function was not directly established in this study, the parallel between the microbiome alterations and visual memory impairment in female rats warrants further exploration of the possibility of a sex-specific gut-brain axis involvement.

DOXO treatment significantly impacts the gut microbiome. Fecal samples were collected 48-hours post the last DOXO injection and samples were subjected to shotgun whole genome sequencing and bioinformatics analysis. Box plots represent measures of alpha diversity: (a) Shannon Index and (b) Observed Taxa. (c) The Principal Coordinates Analysis (PCoA) plot using the Bray-Curtis Dissimilarity metric and 95% Confidence Ellipses represents the beta diversity. (d) The community bar plot represents the mean relative abundance for the different species listed on the right. (e) The box plot for the female rats treated with or without DOXO for the mean relative abundance for species Duncaniella MGBC103508. (f) The box plot for the male rats treated with or without DOXO for the mean relative abundance for species Porphyromonadaceae bacterium UBA7183. (g) The community bar plot represents the mean relative abundance for the different phyla. (h) Box plot for the female rats treated with or without DOXO for the mean relative abundance for phylum Actinobacteria. (i) Box plot for the female rats treated with or without DOXO for the mean relative abundance for phylum Bacteroidetes. (j) Heat map displaying the significant pathways dysregulated in DOXO-treated female rats, where the scale bar range represents the mean abundance for that group. Statistical analysis was performed using: Mann Whitney test (e, f), Unpaired t test (h, i). In all the panels, the groups are separated by treatment (n=4-5 per group). Each biological replicate is displayed as a circle on the graph. *p < 0.05, **p < 0.01.
Discussion
Although the neurotoxic effects of DOXO have been well-documented in animal studies, these studies were often limited to one sex, did not fully mimic a pediatric CICI model, and do not provide a clear understanding of the pathophysiology leading to DOXO-induced cognitive impairment. To address this, in this study, we utilized both male and female rats and demonstrated DOXO-induced changes in the gut microbiome and DOXO-induced cognitive impairment. We showed that DOXO treatment at a juvenile age induces cognitive deficits into young adulthood. We observed that DOXO-treated males are more likely to exhibit spatial memory deficits, and DOXO-treated females are more likely to exhibit visual memory deficits. However, these memory deficits did not persist into adulthood, as we observed no significant deficits 5 months following the last DOXO injection. We found that DOXO-treated female rats have distinct microbial shifts in the gut microbiome.
DOXO does not cross the BBB2, yet it induces neurotoxicity and symptoms of CICI in some patients, possibly through alterations in the gut-brain axis. Here, we reported that DOXO treatment altered the gut microbiome and decreased the mean relative abundance of Bacteroidetes and increased Actinomycetota in female rats. Whereas others have reported in male rats that DOXO decreases the abundance of Bacteroidetes and Verrucomicrobia and increases Proteobacteria and Epsilonbacteraeota29. However, another commonly used pediatric chemotherapy, methotrexate, induced changes in the gut microbiome and significantly decreased the abundance of Bacteroidales30, which is a class belonging to the phylum Bacteroidetes, and a finding consistent with our results. We also found a significant decrease in the mean relative abundance of Muribaculum and Paramuribaculum in male and female rats combined. Additionally, when observing gut microbiome changes in male and female rats combined, we observed a significant increase in the mean relative abundance of Dubosiella, Parabacteroides, and Lachnotalea. These observations are consistent with another study that found DOXO-treated female mice displayed an increase in the relative abundance of Faecalibaculum, Dubosiella, and Lachnospiraceae. The same study also reported a decrease in the relative abundance of Allobaculum, Muribaculum, and Lachnoclostridium31. Overall, our results demonstrate that DOXO induces shifts in the gut microbiome composition, and this is consistent with prior studies of DOXO or other commonly used pediatric chemotherapies.
Moreover, our results provide insights into the associations between a few taxa and cognitive impairment; while the pathology may differ for these cognitive impairment studies, we are still able to make potential inferences. In a study evaluating how diet influences gut microbiome changes, researchers found that a decrease in Bacteroidales may be linked to poorer cognitive flexibility in male mice32. In Alzheimer’s disease patients, there was a higher relative abundance of Actinobacteria (also known as Actinomycetota) and a lower relative abundance of Bacteroidetes (also known as Bacteroidota) when compared to age and gender matched controls with normal cognition33. However, this is in contrast with many other published studies, which typically report that Alzheimer’s subjects with cognitive impairment have increased Bacteroidetes and decreased Firmicutes and Bifidobacterium34. Our results show that the mean relative abundance of species Duncaniella MGBC103508 and Porphyromonadaceae bacterium UBA7183 are increased in female and male rats, respectively; and both of these species belong to phylum Bacteroidetes. Further, we found that DOXO treatment dysregulates the fatty acid elongation pathway, suggesting that the gut-brain axis may play a role in the cognitive deficits. Although not the same chemotherapy, methotrexate treatment also induces an increase in the fatty acid elongation pathway19, supporting a shared metabolic response that requires further investigation. Our results may provide preliminary associations that cognitive impairment and gut microbiome changes may be linked, however, further exploration is needed.
We found evidence of DOXO-induced cognitive dysfunction, and specifically, we observed sex differences in the vulnerability to DOXO-induced behavioral changes and microbiome changes. Many preclinical studies investigating CICI focus on males or females exclusively, leaving an incomplete understanding of the mechanism underlying CICI and whether sex differences are reported. These sex differences are not well understood, yet women are more likely to report problems with memory and concentration35. In the current study, we reported that DOXO significantly induces visual memory deficits in female rats, while DOXO induces spatial memory deficits in male rats. This observation is consistent with other studies that show male rats treated with a similar regimen also display significant spatial memory impairments after DOXO treatment10,11. We observed significant changes in visual memory impairments and in the gut microbiome by DOXO treatment in female rats. While these results cannot prove causal associations, future work testing whether these visual memory deficits are a result of the DOXO-induced gut microbiome changes may result in a better understanding of the mechanistic link between the gut microbiome and CICI. Interestingly, sex differences play a role in the gut microbiome and are plausibly influenced by sex hormones and/or immune responses36. Moreover, sex hormones may modulate the gut-brain axis and play a role in cognitive functions37. Future studies will explore the role of estrogen and testosterone on both the gut microbiome and cognitive function, as others have highlighted the importance of these sex hormones in the gut microbiome38.
We found that DOXO-induced cognitive deficits do not persist 5 months after the last DOXO injection, in contrast to other chemotherapies like methotrexate, where rats display deficits persisting 1 year after the last dose24. However, different DOXO regimens may induce more persistent deficits. Approximately 35% of patients experience symptoms of CICI for months to years after completing treatment39, indicating that the quality of life for patients is significantly impacted. Moreover, our results clearly show that DOXO treatment led to significant weight loss in male rats. Additionally, we observed 50% survival rate of our DOXO-treated male rats at 20 weeks of age (12 weeks after the last DOXO injection). Our weight loss results are consistent with other studies that report weight loss in male DOXO-treated rats40,41. However, others have reported weight loss for both male and female DOXO-treated rats42, with a more pronounced effect in males43. Moreover, others also reported a decrease in the survival of DOXO-treated male rats43,44. Although in our study we did not conduct a necropsy to determine the cause of death, it is likely due to the systemic toxicity induced by DOXO treatment, including mucositis and cardiotoxicity.
While our pilot study sheds light on DOXO-induced cognitive impairment and DOXO-induced microbiome changes in both male and female rats, further investigations are required. In this study, our limited sample size prevented us from comparing microbiome changes between rats that developed cognitive impairment and those that did not following DOXO treatment. By increasing the number of animals per group, we can investigate microbiome alterations in DOXO-induced cognitive impairment. Additionally, we did not investigate the long-term effects of the microbiome, which would allow us to observe if these microbiome changes persist. Ongoing studies in our laboratory will involve conducting behavioral testing at intermediate timepoints to detect how long DOXO-induced cognitive impairment will persist. Future directions include investigating whether fecal microbiota transplantation or probiotics can rescue or prevent DOXO-induced cognitive impairment in rats. Lastly, our DOXO treatment regimen did not fully recapitulate a clinical regimen because DOXO is typically administered with the cardioprotectant, dexrazoxane45,46,47, which may change its toxicity profile.
Conclusion
Taken together, our findings indicate DOXO treatment induces cognitive impairment in a sex-dependent manner, where female rats are more likely to display visual memory impairments, whereas male rats are more likely to exhibit spatial memory impairment following DOXO treatment. Our results support that DOXO treatment induces alterations in the gut microbiome, with a profound effect in female rats. The observations noted in our pilot work warrant further investigations to delve into mechanistic studies of whether changes in the microbiome contribute to CICI or not. Our study sheds light on the development of potential therapeutic interventions for CICI that may improve the quality of life for patients.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Armenian, S. & Bhatia, S. Predicting and preventing Anthracycline-Related cardiotoxicity. Am. Soc. Clin. Oncol. Educ. Book. 38, 3–12. https://doi.org/10.1200/EDBK_100015 (2018).
Chabner, B. A. & Longo, D. L. Cancer Chemotherapy, Immunotherapy and Biotherapy: Principles and Practice, 6e. (Lippincott Williams & Wilkins, a Wolters Kluwer business, (2018).
Kotb, M. G., Soliman, A. E. R., Ibrahim, R. I., Said, R. M. M. & El Din, M. M. W. Chemotherapy-induced cognitive impairment in hematological malignancies. Egypt. J. Neurol. Psychiatry Neurosurg.55, 56. https://doi.org/10.1186/s41983-019-0104-9 (2019).
Ren, X. et al. Plausible biochemical mechanisms of chemotherapy-induced cognitive impairment (chemobrain), a condition that significantly impairs the quality of life of many cancer survivors. Biochimica et Biophysica Acta (BBA)1865, 1088–1097. https://doi.org/10.1016/j.bbadis.2019.02.007 (2019).
Argyriou, A. A., Assimakopoulos, K., Iconomou, G., Giannakopoulou, F. & Kalofonos, H. P. Either called chemobrain’’ or chemofog,‘’ the Long-Term Chemotherapy-Induced cognitive decline in Cancer survivors is real. J. Pain Symptom Manag. 41, 126–139. https://doi.org/10.1016/j.jpainsymman.2010.04.021 (2011).
van der Plas, E. et al. Neurocognitive late effects of chemotherapy in survivors of acute lymphoblastic leukemia: focus on methotrexate. J. Can. Acad. Child. Adolesc. Psychiatry. 24, 25–32 (2015).
Williams, A. M. & Cole, P. D. Biomarkers of cognitive impairment in pediatric cancer survivors. J. Clin. Oncol.39, 1766–1774. https://doi.org/10.1200/JCO.20.02436 (2021).
Van Dongen-Melman, J. E. Developing psychosocial aftercare for children surviving cancer and their families. Acta Oncol. 39, 23–31. https://doi.org/10.1080/028418600430932 (2000).
Askins, M. A. & Moore, B. D. 3 Preventing neurocognitive late effects in childhood cancer survivors. J. Child. Neurol. 23, 1160–1171. https://doi.org/10.1177/0883073808321065 (2008).
Ali, M. A., Menze, E. T., Tadros, M. G. & Tolba, M. F. Caffeic acid phenethyl ester counteracts doxorubicin-induced chemobrain in Sprague-Dawley rats: emphasis on the modulation of oxidative stress and neuroinflammation. Neuropharmacology 181, 108334. https://doi.org/10.1016/j.neuropharm.2020.108334 (2020).
Tong, Y., Wang, K., Sheng, S. & Cui, J. Polydatin ameliorates chemotherapy-induced cognitive impairment (chemobrain) by inhibiting oxidative stress, inflammatory response, and apoptosis in rats. Biosci. Biotechnol. Biochem.84, 1201–1210. https://doi.org/10.1080/09168451.2020.1722057 (2020).
Leung, W. S. et al. Protective effects of Diallyl trisulfide (DATS) against doxorubicin-induced inflammation and oxidative stress in the brain of rats. Free Radic Biol. Med. 160, 141–148. https://doi.org/10.1016/j.freeradbiomed.2020.07.018 (2020).
Alhowail, A. H. Pioglitazone ameliorates DOX-induced cognitive impairment by mitigating inflammation, oxidative stress, and apoptosis of hippocampal neurons in rats. Behav. Brain. Res. https://doi.org/10.1016/j.bbr.2023.114714 (2023).
Christie, L. A. et al. Impaired cognitive function and hippocampal neurogenesis following cancer chemotherapy. Clin. Cancer Res.18, 1954–1965. https://doi.org/10.1158/1078-0432.CCR-11-2000 (2012).
Bajic, J. E., Johnston, I. N., Howarth, G. S. & Hutchinson, M. R. From the Bottom-Up: chemotherapy and Gut-Brain Axis dysregulation. Front. Behav. Neurosci. 12, 104. https://doi.org/10.3389/fnbeh.2018.00104 (2018).
Ciernikova, S., Mego, M. & Chovanec, M. Exploring the potential role of the gut microbiome in chemotherapy-induced neurocognitive disorders and cardiovascular toxicity. Cancers (Basel)https://doi.org/10.3390/cancers13040782 (2021).
Subramaniam, C. B. et al. The microbiota-gut-brain axis: an emerging therapeutic target in chemotherapy-induced cognitive impairment. Neurosci. Biobehav Rev. 116, 470–479. https://doi.org/10.1016/j.neubiorev.2020.07.002 (2020).
Loman, B. R., Jordan, K. R., Haynes, B., Bailey, M. T. & Pyter, L. M. Chemotherapy-induced neuroinflammation is associated with disrupted colonic and bacterial homeostasis in female mice. Sci. Rep.9, 16490. https://doi.org/10.1038/s41598-019-52893-0 (2019).
Chen, Y. C. et al. The Impact of Gut Microbiota Changes on Methotrexate-Induced Neurotoxicity in Developing Young Rats.. >Biomedicines https://doi.org/10.3390/biomedicines12040908 (2024).
Otto-Dobos, L. D. et al. Chemotherapy-induced gut Microbiome disruption, inflammation, and cognitive decline in female patients with breast cancer. Brain Behav. Immun. 120, 208–220. https://doi.org/10.1016/j.bbi.2024.05.039 (2024).
Nair, A. B. & Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic. Clin. Pharm.7, 27–31. https://doi.org/10.4103/0976-0105.177703 (2016).
Mandal, S. et al. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis.26, 27663. https://doi.org/10.3402/mehd.v26.27663 (2015).
Wen, J. et al. Methotrexate causes persistent deficits in memory and executive function in a juvenile animal model. Neuropharmacology 139, 76–84. https://doi.org/10.1016/j.neuropharm.2018.07.007 (2018).
Wen, J. et al. Cognitive impairment persists at least 1 year after juvenile rats are treated with methotrexate. Neuropharmacology 206, 108939. https://doi.org/10.1016/j.neuropharm.2021.108939 (2022).
Ennaceur, A. & Meliani, K. A new one-trial test for neurobiological studies of memory in rats. III. Spatial vs. non-spatial working memory. Behav. Brain Res.51, 83–92. https://doi.org/10.1016/s0166-4328(05)80315-8 (1992).
Hattiangady, B. et al. Object location and object recognition memory impairments, motivation deficits and depression in a model of Gulf war illness. Front. Behav. Neurosci. 8, 78. https://doi.org/10.3389/fnbeh.2014.00078 (2014).
Denninger, J. K., Smith, B. M. & Kirby, E. D. Novel object recognition and object location behavioral testing in mice on a budget. J. Vis. Exp. https://doi.org/10.3791/58593 (2018).
Fekete, M. et al. Exploring the influence of Gut-Brain Axis modulation on cognitive health: A comprehensive review of prebiotics, probiotics, and symbiotics. Nutrients 16 https://doi.org/10.3390/nu16060789 (2024).
Lin, H. et al. Yellow wine polyphenolic compound protects against doxorubicin-induced cardiotoxicity by modulating the composition and metabolic function of the gut microbiota. Circ. Heart Fail.14, e008220. https://doi.org/10.1161/CIRCHEARTFAILURE.120.008220 (2021).
Zhou, B. et al. Induction and amelioration of Methotrexate-Induced Gastrointestinal toxicity are related to immune response and gut microbiota. EBioMedicine 33, 122–133. https://doi.org/10.1016/j.ebiom.2018.06.029 (2018).
Huang, J. et al. Involvement of abnormal gut microbiota composition and function in Doxorubicin-Induced cardiotoxicity. Front. Cell. Infect. Microbiol. 12, 808837. https://doi.org/10.3389/fcimb.2022.808837 (2022).
Magnusson, K. R. et al. Relationships between diet-related changes in the gut Microbiome and cognitive flexibility. Neuroscience 300, 128–140. https://doi.org/10.1016/j.neuroscience.2015.05.016 (2015).
Zhuang, Z. Q. et al. Gut microbiota is altered in patients with Alzheimer’s disease. J. Alzheimers Dis.63, 1337–1346. https://doi.org/10.3233/JAD-180176 (2018).
Vogt, N. M. et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep.7, 13537. https://doi.org/10.1038/s41598-017-13601-y (2017).
Kohli, S. et al. Self-reported cognitive impairment in patients with cancer. J. Oncol. Pract.3, 54–59. https://doi.org/10.1200/JOP.0722001 (2007).
Santos-Marcos, J. A., Mora-Ortiz, M., Tena-Sempere, M., Lopez-Miranda, J. & Camargo, A. Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases. Biology Sex. Differences. 14, 4. https://doi.org/10.1186/s13293-023-00490-2 (2023).
Leao, L., Miri, S. & Hammami, R. Gut feeling: exploring the intertwined trilateral nexus of gut microbiota, sex hormones, and mental health. Front. Neuroendocr. 76, 101173. https://doi.org/10.1016/j.yfrne.2024.101173 (2025).
Yoon, K. & Kim, N. Roles of sex hormones and gender in the gut microbiota. J. Neurogastroenterol Motil. 27, 314–325. https://doi.org/10.5056/jnm20208 (2021).
Janelsins, M. C., Kesler, S. R., Ahles, T. A. & Morrow, G. R. Prevalence, mechanisms, and management of cancer-related cognitive impairment. Int. Rev. Psychiatry26, 102–113. https://doi.org/10.3109/09540261.2013.864260 (2014).
Timm, K. N. et al. Early detection of doxorubicin-induced cardiotoxicity in rats by its cardiac metabolic signature assessed with hyperpolarized MRI. Commun. Biol.3, 692. https://doi.org/10.1038/s42003-020-01440-z (2020).
Podyacheva, E. et al. Modeling doxorubicin-induced cardiomyopathy with fibrotic myocardial damage in Wistar rats. Cardiol. Res.13, 339–356. https://doi.org/10.14740/cr1416 (2022).
Favreau-Lessard, A. J., Blaszyk, H., Jones, M. A., Sawyer, D. B. & Pinz, I. M. Systemic and cardiac susceptibility of immune compromised mice to doxorubicin. Cardiooncology 5, 2. https://doi.org/10.1186/s40959-019-0037-6 (2019).
Moulin, M. et al. Sexual dimorphism of doxorubicin-mediated cardiotoxicity: Potential role of energy metabolism remodeling. Circ. Heart Fail.8, 98–108. https://doi.org/10.1161/CIRCHEARTFAILURE.114.001180 (2015).
Zobeydi, A. M. et al. Publisher correction: Mitigating doxorubicin-induced hepatotoxicity in male rats: The role of aerobic interval training and curcumin supplementation in reducing oxidative stress, endoplasmic reticulum stress and apoptosis. Sci. Rep.15, 12004. https://doi.org/10.1038/s41598-025-95422-y (2025).
Cvetkovic, R. S. & Scott, L. J. Dexrazoxane: a review of its use for cardioprotection during anthracycline chemotherapy. Drugs 65, 1005–1024. https://doi.org/10.2165/00003495-200565070-00008 (2005).
Langer, S. W. Dexrazoxane for the treatment of chemotherapy-related side effects. Cancer Manag Res. 6, 357–363. https://doi.org/10.2147/CMAR.S47238 (2014).
Lipshultz, S. E. Exposure to anthracyclines during childhood causes cardiac injury. Semin. Oncol.33, 8–14. https://doi.org/10.1053/j.seminoncol.2006.04.019 (2006).
Acknowledgements
We would like to acknowledge Denise Lynch, scientific implication lead at One CodeX (Wilmington, DE, USA) for assistance with the One CodeX platform.
Funding
We acknowledge funding from the New Jersey Commission of Cancer Research Predoctoral Fellowship COCR24PRF009, the Biotechnology Training Program NIH T32 GM135141, and the New Jersey Pediatric Hematology and Oncology Research Center of Excellence (NJ PHORCE) Pilot Grant.
Author information
Authors and Affiliations
Contributions
Conceptualization: C.P, P.C.; Methodology: C.P.; Formal analysis: C.P.; Investigation: C.P, F.D., J.W.; Data curation: C.P.; Figure Preparation: C.P.; Writing—original draft: C.P.; Writing—review & editing: C.P., P.C., J.W., F.D.; Supervision: P.C.; Funding acquisition: C.P., P.C.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
41598_2025_12352_MOESM1_ESM.pdf
Supplementary Material 1
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Patel, C., Willekens, J., Diglio, F. et al. Doxorubicin causes cognitive impairment and alters gut microbiota in both male and female juvenile rats. Sci Rep 15, 27448 (2025). https://doi.org/10.1038/s41598-025-12352-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12352-5
Keywords
This article is cited by
-
Changes in gut, microbiome, and cognition after doxorubicin, cyclophosphamide, and paclitaxel chemotherapy treatment
Scientific Reports (2026)
-
Influence of ApoE genotype on doxorubicin-induced cognitive impairment in juvenile rats
Scientific Reports (2025)
