
Abstract
Colombia’s strategic location, encompassing diverse climates and ecosystems, enables abundant flora. This environment offers bioprospecting opportunities, emphasizing sustainable plant use. Our main objective was to expand the knowledge of the chemical, pharmacological, and cosmetic potential of three cultivated species, namely, T. pumilea, H. lantanifolia, and A. popayanensis. UHPLC-ESI-Orbitrap-HRMS and quantification of total flavonoid, phenolic, and condensed tannin contents were employed to elucidate the chemical composition. The antioxidant activity and photoprotective capacity were evaluated by spectrophotometry. Calu-1 and HepG2 cells were used to assess cytotoxicity by MTT assay, and early intracellular ROS production was evaluated using an H2-DCFDA probe. The results demonstrate the richness of the chemical composition of the studied species, identifying compounds with therapeutic properties, including gallic acid, p-coumaric acid, caffeic acid, rosmarinic acid, chlorogenic acid, apigenin, and rutin, which are recognized as antioxidant and ultraviolet protective agents, supporting their uses in traditional medicine. The cytotoxicity evaluation on Calu-1 and HepG2 cells revealed the antiproliferative effects of these plants, especially the T. pumilea extract, on Calu-1 cells, and the possible influence of ROS generation, which could explain the observed effects. This research underlines the need for thorough evaluations of toxicological effects and the characterization of the mechanism of action involved to support the results obtained.
Similar content being viewed by others


Introduction
Colombia ranks second in South America in botanical diversity, consolidating itself as a megadiverse country1 that remains largely underexplored2. Its strategic geographical position, which encompasses a wide variety of climates and the coexistence of diverse ecosystems, plays a crucial role in the richness and abundance of its flora3. This privileged environment opens significant opportunities for bioprospecting studies, highlighting the need to promote the sustainable use of plant resources.
However, in the Colombian territory, unsustainable agricultural practices, climate change, and sociocultural factors that induce population displacements threaten the conservation of species and ancestral knowledge about their medicinal properties. This situation puts at risk not only biodiversity, but also cultural heritage linked to native flora4. Faced with this panorama, the search for economic development based on biodiversity promotes recognition of plants’ potential and the adoption of sustainable agricultural practices, to preserve the vegetable wealth of the country and guarantee its long-term responsible use5. Thus, the conservation and management of natural resources are presented as fundamental pillars to maintain the biological diversity that characterizes Colombia and to ensure that future generations can benefit from its multiple uses, both in traditional medicine and in scientific and technological innovation. Research initiatives in this field facilitate the discovery of valuable bioactive compounds and emphasize the urgent need for responsible management of natural resources (https://bio-reto-xxi.uis.edu.co/ accessed March 10, 2024).
Within this context, the chemical analysis and biological activities identified in extracts from grown plants play a fundamental role in the valorisation of biodiversity, serving as a basis for the sustainable development of products intended for human consumption, such as cosmetics, cosmeceuticals, and pharmaceuticals6,7,8,9. To this end, three plant species cultivated in Bucaramanga, Colombia: Turnera pumilea, Hyptis lantanifolia, and Ageratina popayanensis, have been selected based on their notable chemical and biological characteristics, which hold potential relevance for the cosmetic and pharmaceutical industries.
Turnera pumilea L. is one of 140 accepted species of the genus Turnera in Plants of the World Online of Royal Botanic Gardens [https://powo.science.kew.org/ accessed March 10, 2024], sometimes identified as part of the Turneraceae family. However, the Angiosperm Phylogeny Group (APG) III belongs to the Passifloraceae family10so the APG III guidelines will be considered in this research. The common name of this species is “Damiana” or “Yellow Damiana”. It is described as an annual subshrub that grows mainly in the seasonally dry tropical biome, with a native distribution area ranging from Mexico to Tropical America. Species of this genus have a wide range of traditional uses, including abortive properties, and the treatment of anemia, diabetes, respiratory tract, gastrointestinal, and skin disorders10.
Hyptis lantanifolia Poit. (Lamiaceae) is a species popularly known as “Santa Maria Bush” or “Coquito de Monte” that grows in neotropical regions with ample distribution in Colombia and Mexico [https://www.gbif.org/es/species/2926810 accessed March 16, 2024]. Species within this genus have been utilized in traditional medicine for treating viral processes, infections, fever, cramps, and disorders in the liver, gastrointestinal tract, and skin11. In recent years, scientific evaluation of the genus Hyptis has revealed its potential as a hormone modulator, and anti-inflammatory, anticancer, antimicrobial, insecticidal, molluskicidal, and acaricidal agent12. The water extract obtained from the whole plant of H. lantanifolia has shown potent inhibition of human immunodeficiency virus-reverse transcriptase (HIV-RT); hence, its anti-HIV effects are presently under investigation13.
Ageratina popayanensis (Hieron.) R.M.King & H.Rob. The so-called “Chilco Negro” or “Amargoso” is a species of the Asteraceae family native to Colombia with distribution in the northern region of the Andes, including Venezuela [https://www.gbif.org/es/species/5400619 accessed March 16, 2024]. Different studies on extracts and phytocompounds identified in species of this genus have revealed a wide spectrum of bioactivities, including anti-inflammatory, antiviral, antimicrobial (relief of onychomycosis), antiprotozoal, antioxidant, antidiabetic, antinociceptive, antifeedant, larvicidal, acaricidal, and wound-healing activities14.
This research aims to generate valuable insights into the hydroalcoholic extracts of T. pumilea, H. lantanifolia, and A. popayanensis, focusing on their chemical composition and potential applications in the pharmaceutical and skincare industries. Key areas of investigation include the in vitro antioxidant capacity, photoprotective effects, cytotoxicity against human cancer cell lines (Calu-1 and HepG2), and early reactive oxygen species (ROS) generation. These findings will contribute to understanding the traditional uses and the sustainability of Colombia’s rich botanical resources.
Materials and methods
Vegetal material
Aerial parts of T. pumilea L. (Passifloraceae), H. lantanifolia Poit (Lamiaceae), and A. popayanensis (Hieron) R. King & H. Rob (Asteraceae) were obtained from the garden of the National Center for Agroindustrialization of Aromatic and Medicinal Tropical Plant Species (CENIVAM) at the Industrial University of Santander (UIS, Bucaramanga, Colombia). Botanical identification was carried out at the UIS Herbarium (national registry RNC:030) under the supervision of the curator, Prof. Dr. Andrés Felipe Castaño González. The process was based on morphological comparison with reference specimens and the use of standard taxonomic keys for the families Passifloraceae, Lamiaceae, and Asteraceae. The corresponding exsiccatae and voucher specimens were deposited at the UIS Herbarium under the following accession codes: T. pumilea (UIS 21972), H. lantanifolia (UIS 21974), and A. popayanensis (UIS 22040).
Hydroalcoholic extracts
The aerial parts of the plants (100 g), either fresh before distillation or residual biomass was dried and ground, were combined with a 70% aqueous ethanol solution (2 L) and placed in an ultrasonic bath (Elmasonic S15H, Singen, Germany) at 50 °C for 1 h. Afterward, the extracts were filtered, concentrated using a Heidolph rotary evaporator (Hei-VAP, Advantage HL, Chicago, IL, USA), and then lyophilized (VirTis AdVantage Plus, SP Scientific, Gardiner, NY, USA). The final products were stored at 4 °C, in the dark. Solvent extractions were carried out in triplicate.
Sample preparation for UHPLC-ESI-Orbitrap-HRMS analysis
The samples were weighed (0.1 mg) and dissolved in a mobile phase of methanol (MeOH): water 50:50 with 0.1% formic acid (FA) and 5 mM ammonium formate (1 mL) to achieve a final concentration of 100 mg/L. Phenolic compounds were analyzed using an ultra-high performance liquid chromatography system on a VanquishTM instrument (Thermo Scientific, Waltham, MA, USA), equipped with a thermostatically controlled column compartment (40 °C). Chromatographic separation was performed on a Zorbax Eclipse XDB C18 column (Sigma Aldrich, St. Louis, MO, USA) 50 mm L × 2.1 mm I.D. with a particle size of 1.8 μm. The flow rate of the mobile phase, containing (A) water (0.1% FA + 5 mM FA) and (B) MeOH (0.1% FA + 5 448 mM FA), was 300 µL/min. The initial gradient conditions were 100% A, which was linearly changed to 100% B over 8 min, held for 4 min, returned to 100% A in 1 min, and held for 3 min. The injection volume was 2 µL. The liquid chromatograph was connected to a Q Exactive Plus Orbitrap mass spectrometer (Thermo Scientific, Bremen, Germany) with a heated electrospray ionization source (HESI-II). C-Trap ions were transferred to a high-energy collisional dissociation (HCD) cell and fragmented with collision energies ranging from 10 to 70 eV. Data were acquired and analyzed using Thermo Scientific™ Dionex™ Chromatography Data System (CDS) software, version 7.2, and Thermo Xcalibur 3.1 software (Thermo Scientific, San Jose, CA, USA). Compound identification was performed by comparing retention times (tR), exact masses, isotopic patterns, and MS spectra with those of standard substances, and by consulting spectral databases [e.g., HMDB (Wishart, D.S., Guo, A., Oler, E. & Wang, F. HMDB 5.0: The human metabolome database for 2022. Nucleic Acids Research. 2022. Available on-line: www.hmdb.ca(open in a new window) and MassBank (MassBank. Available on-line: www.massbank.eu/MassBank/(open in a new window)] and relevant literature. The confidence level for each compound was assigned according to the Metabolomics Standards Initiative (MSI) and is indicated in the table footnotes as follows: (a) confirmed identification using authentic standards (MSI Level 1); (b) tentative identification based on literature reports for species of the genera Turnera, Hyptis, and Ageratina (MSI Level 2); and (c) tentative annotation based solely on exact mass and isotopic pattern (MSI Level 3).
Total flavonoid, phenolic, and tannin contents
For the quantification of total flavonoid, phenolic, and condensed tannin contents the methodology of Sánchez-Gutiérrez et al.15 was followed. Absorbance measurements were acquired using a Varioskan™ LUX Multimode Microplate Reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
The total flavonoid content was quantified by constructing a standard curve of the flavonoid rutin (A415 nm = 0.0089x + 0.0017, R² = 0.9994) at five concentrations ranging from 0.002 to 0.0315 mg/mL. Flavonoid levels were expressed as milligrams of rutin equivalents per gram of dry tissue (mg RE/g DT), with the extract concentration for analysis set at 1.0 mg/mL.
Similarly, the total phenolic content was determined using a gallic acid standard curve (A760 nm = 0.0046x + 0.044, R² = 0.9966) established from seven concentrations ranging from 0.003 to 0.200 mg/mL. The phenolic content was reported as milligrams of gallic acid equivalents per gram of dry tissue (mg AGE/g DT), with the extract concentration for analysis set at 1.0 mg/mL.
The total condensed tannin content was assessed through a (+) catechin standard curve (A500 nm = 0.0037x + 0.1181, R² = 0.9873) derived from eight concentrations ranging from 0.0078 to 1.00 mg/mL. Condensed tannin levels were expressed as milligrams of (+) catechin equivalents per gram of dry tissue (mg CE/g DT), with the extract concentration for analysis set at 1.0 mg/mL.
Antioxidant activity
The DPPH assay, a widely recognized and used method in antioxidant research, was used to determine the antioxidant capacity of the extracts4,16,17. This assay allows estimating the efficacy of antioxidant compounds in neutralizing superoxide anion (O₂•−), hydroxyl (•OH), alkoxyl (RO•), and peroxyl radicals (ROO•). These molecules can cause cellular damage and contribute to the development of various pathologies associated with the generation of ROS, such as cancer18 and skin disorders8.
The principle of the DPPH assay is based on the reduction of the free radical DPPH (2,2-diphenyl-1-picrylhydrazyl), which presents an intense purple color, to a non-radical form, which progressively fades upon interaction with antioxidants. The antioxidant capacity evaluated in this assay reflects a mixed mechanism involving both electron transfer and hydrogen atom transfer. The reaction rate of the DPPH radical depends largely on the steric accessibility of the radical site, which influences the efficiency with which antioxidants can donate electrons or hydrogen atoms to neutralize it19.
This study employed the kit from Bioquochem (Llanera, Asturias, Spain) to determine the DPPH radical scavenging activity17. Each extract was tested in duplicate at five different concentrations (1:2 dilution), ranging from 31.23 to 500 µg/mL. The extract (20 µL) was subsequently reacted with DPPH solution (200 µL) in the dark at room temperature for 30 min. A Trolox standard curve was prepared at the various concentrations (500, 400, 300, 200, 100, and 0 µg/mL). The absorbance data at 517 nm were acquired with a Varioskan™ LUX Multimode Microplate Reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The scavenging DPPH radical percentage (%) for each standard point was obtained with the following formula: % inhibition = [1 − (Abs Sn/Abs S1)] × 100, where Abs S1 is the DPPH•+ radical absorption without inhibition and Abs Sn is the DPPH•+ radical absorption of the corresponding standard. The Trolox equivalent antioxidant capacity (TEAC) of the tested concentrations of extracts was subsequently calculated via the following formula: TEAC (µM) = % inhibition − intercept/slope.
Photoprotective capacity
Based on its wavelength, the ultraviolet (UV) radiation emitted by sunlight is classified into three categories: UVA (320–400 nm), UVB (280–320 nm), and UVC (100–280 nm). While UVC rays are largely absorbed by the ozone layer and do not reach the Earth’s surface, approximately 95% and 5% of UVA and UVB rays penetrate the atmosphere, respectively. These UV radiations pose a significant health risk when overexposed, as they can trigger various conditions, such as photoaging, immune system suppression, and tumor development. Currently, the use of plant extracts as natural sunscreens is being explored to counteract these harmful effects. Plant extracts are an interesting source of bioactive compounds (flavonoids and phenolic compounds) that exhibit a remarkable capacity to absorb radiation in the wavelength range between 280 and 400 nm, corresponding to the UVB and UVA regions of the UV spectrum. This property makes them valuable ingredients for incorporation into cosmetic and cosmeceutical formulations, where they act as natural filters that contribute to skin protection. In addition to their absorption capacity, these compounds possess antioxidant activities that help neutralize free radicals generated by sun exposure20,21.
In this study, the photoprotective capacity of the extracts was evaluated by spectrophotometry with a Varioskan™ LUX Multimode Microplate Reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA)7. The absorbance data at different wavelengths and optical path lengths were recorded to determine the sun protection factor (290–320 nm; 5 nm), critical wavelength (290–400 nm; 1 nm), UVA/UVB ratio (290–400 nm; 5 nm), and erythema transmission/pigmentation transmission (292–372 nm; 5 nm). The UV protection criteria used were those described previously3,7.
Sun protection factor (SPF)
SPF values were obtained by applying the Mansur equation22:: \(\:\text{S}\text{P}\text{F}=\text{C}\text{F}\:\times\:{\sum\:}_{290\:nm}^{320\:nm}\text{E}\text{E}\:\left({\uplambda\:}\right)\times\:\:\text{I}\left({\uplambda\:}\right)\times\:\:\text{A}\text{b}\text{s}\:\left({\uplambda\:}\right),\:\) where CF is the correction factor (equal to 10), EE (λ) × I(λ) are constants at each wavelength obtained from the correlation between EE (erythematous effect of radiation of wavelength λ) and I (the solar intensity at wavelength λ) (Supplementary Table S1), and Abs is the absorbance of the solution at wavelength λ.
Critical wavelength (λc)
λc is the wavelength at which 90% of the area under the absorbance curve is found, considering the integral of the absorption spectrum (290 nm to 400 nm)23. This value was determined via the following equation: \(\:{\int\:}_{290\:\text{n}\text{m}}^{{\uplambda\:}\text{c}}\text{A}\:\left({\uplambda\:}\right)\text{d}{\uplambda\:}\:=0.9\:{\int\:}_{290\:\text{n}\text{m}}^{400\:\text{n}\text{m}}\text{A}\left({\uplambda\:}\right)\text{d}{\uplambda\:}\:,\) where A is the absorbance at wavelength λ3,7.
UVA/UVB ratio
To determine the UVA/UVB ratio, the average UVA absorbance was related to the average UVB absorbance using the following equation: \(\:\text{U}\text{V}\text{A}/\text{U}\text{V}\text{B}=\:\frac{\sum\:_{320\:\text{n}\text{m}}^{400\:\text{n}\text{m}}\text{A}\left({\uplambda\:}\right),\:\text{d}\left({\uplambda\:}\right)}{\sum\:_{290\:\text{n}\text{m}}^{320\:\text{n}\text{m}}\text{A}\left({\uplambda\:}\right),\:\text{d}\left({\uplambda\:}\right)}\)
, where A(λ) is the effective absorbance related to the transmittance of the sunscreen23.
Percentages of erythema transmission and pigmentation transmission
The ability to induce redness (erythema transmission) or sunspots on the skin (pigmentation transmission), was determined from the transmission (T), to then apply the following Eq24.:
-
a)
\(\:\text{E}\text{r}\text{y}\text{t}\text{h}\text{e}\text{m}\text{a}\:\text{t}\text{r}\text{a}\text{n}\text{s}\text{m}\text{i}\text{s}\text{s}\text{i}\text{o}\text{n}\:\left(\text{\%}\right)=\frac{\text{E}\text{e}}{\sum\:\text{F}\text{e}}=\sum\:\frac{(\text{T}\:\times\:\:\text{F}\text{e})}{\sum\:\text{F}\text{e}}\), where Fe (erythema flux) is a constant that corresponds to wavelengths ranging from 292 to 338 nm (Supplementary Table S2).
-
b)
\(\:\text{P}\text{i}\text{g}\text{m}\text{e}\text{n}\text{t}\text{a}\text{t}\text{i}\text{o}\text{n}\:\text{t}\text{r}\text{a}\text{n}\text{s}\text{m}\text{i}\text{s}\text{s}\text{i}\text{o}\text{n}\:\left(\text{\%}\right)=\frac{\text{E}\text{p}}{\sum\:\text{F}\text{p}}=\sum\:\frac{\left(\text{T}\:\times\:\:\text{F}\text{p}\right)}{\sum\:\text{F}\text{p}},\) where Fp (pigmentation flux) is a constant that corresponds to wavelengths ranging from 322 to 372 nm (Supplementary Table S2).
Cytotoxicity and early intracellular ROS production
In this study, the MTT assay was employed to evaluate the in vitro cytotoxicity of plant extracts by measuring cell viability. MTT (3-(4, 5-dimethylthiazol-2-yl)−2,5-diphenyltetrazolium bromide), a water-soluble tetrazolium salt, is selectively absorbed by metabolically active cells and enzymatically reduced by mitochondrial succinate dehydrogenase to form insoluble purple formazan crystals. These crystals are subsequently dissolved using an organic solvent, typically dimethyl sulfoxide (DMSO), and quantified spectrophotometrically. The intensity of the purple color directly correlates with the number of viable cells, providing a reliable indicator of cellular metabolic activity25.
The human lung epidermoid carcinoma cells (Calu-1) and human hepatocellular carcinoma cell line (HepG2) obtained from the American Type Tissue Culture Collection (ATCC, USA) and ScienCell (Carlsbad, CA, USA) were cultured in Eagle’s minimum essential medium (EMEM; Quality Biological, Gaithersburg, MD, USA) supplemented with 10% FBS (Biowest, Riverside, MO, USA) and 1% penicillin/streptomycin (Sigma-Aldrich, St. Louis, MI, USA), and maintained in a humidified atmosphere with 5% CO2 at 37°C4.
The cells were seeded in 96-well plates at a density of 2 × 104 cells per well and incubated at 37 °C in 5% CO2 for 24 h. The cells were then treated with the extract at concentrations ranging from 3.9 to 500 µg/mL for 24 h. Subsequently, MTT solution was added to each well at a concentration of 5 mg/mL, and the plates were further incubated at 37 °C for 4 h. After incubation, 200 µL of DMSO (Panreac Applichem®, Barcelona, Spain) was added to dissolve the formazan crystals in each well, and the absorbance was measured at 570 nm using a spectrophotometric microplate reader (Varioskan™ LUX, Thermo Fisher Scientific, Inc., Waltham, MA, USA). Cell viability was determined by comparing the absorbance of treated cells to that of untreated cells. Three independent experiments were performed with four replicates for each treatment5.
Finally, the percentages of cell viability were normalized and calculated with the following equation: cell viability (%) = (ODtest/ODcontrol)×100, where OD is the optical density or absorbance of the test or control, respectively.
To quantify the early ROS production of the three extracts, hydrogen peroxide (H2O2) at a concentration of 200 µM was used as a positive control, which resulted in high intracellular ROS production at each recorded time interval (up to 120 min)3. In this study, the results revealed that the hydroalcoholic extracts from T. pumilea, H. lantanifolia, and A. popayanensis evaluated in HepG2 and Calu-1 cells showed no significant differences in the levels of ROS detected compared with those in the control.
Data analysis
Data were obtained in triplicate (n = 3) for the total content of flavonoids, phenolics, and condensed tannins, as well as for the photoprotective activity, cytotoxicity, and early intracellular ROS, and in duplicate (n = 2) for antioxidant activity. The results are expressed as the mean ± SD (standard deviation). The normality of the data obtained from the cell viability assays employing Calu-1 and HepG2 cell lines was assessed using the Shapiro-Wilk test. Significant differences between experimental groups were evaluated using a one-way analysis of variance (ANOVA), followed by Dunnett’s test. Each treatment’s mean inhibitory concentration at 50% (IC50 values) was calculated using a non-linear sigmoid curve fitting with a four-parameter logistic model. For each IC50 estimate, 95% confidence intervals were reported. This fit was performed on the cell proliferation data versus the logarithm of the concentration of the compound. The results were analyzed using GraphPad Prism 8.0.
Results
Hydroalcoholic extracts of the plant species T. pumilea, H. lantanifolia, and A. popayanensis were analyzed by UHPLC-ESI+/−-Orbitrap-MS. The extracted ion currents (EICs), presented in Fig. 1A and C, allowed the precise detection and quantification of the specific ions present in each extract, filtered in SIM mode to identify both protonated [M + H] + and deprotonated [M − H] − species. The analysis facilitated the chemical characterization of the compounds in the evaluated extracts. The initial identification was based on comparing exact masses and isotopic patterns, allowing the assignment of molecular formulas. These data were subsequently compared with information available in scientific literature, especially in studies related to species of the genera Turnera, Hyptis, and Ageratina, providing taxonomic and chemical support for the assignments made. Finally, the identification was confirmed by comparing retention times (tR) and mass spectra with reference standards, ensuring the correct assignment of key metabolites. Furthermore, a high-energy collision dissociation (HCD) cell was used to generate characteristic fragments that further elucidated the structure of the detected compounds. The complete results of this analysis are summarized in Tables 1 and 2, and 3.

Extracted ion currents (EICs), obtained using UHPLC-ESI+/−-Orbitrap-MS, operated in SIM mode to filter the protonated [M + H]+ or deprotonated [M − H]− molecules of substances present in the hydroalcoholic extract of (A) T. pumilea, (B) H. lantanifolia, and (C) A. popayanensis.
Total flavonoid, phenolic, and condensed tannin contents of hydroalcoholic extracts
The total contents of flavonoids, phenolics, and condensed tannins in the hydroalcoholic extracts of the three plant species evaluated are presented in Table 4. The hydroalcoholic extract of H. lantanifolia exhibited the highest flavonoid content (175.9 ± 12.4 mg RE/g DT), whereas A. popayanensis had the lowest (108.8 ± 0.7 mg RE/g DT). Turnera pumilea showed the highest phenolic content (238.3 ± 1.3 mg GAE/g DT), with H. lantanifolia displaying the lowest phenolic content (191.3 ± 0.5 mg AGE/g DT). In terms of condensed tannins, Ageratina popayanensis had the highest content (43.0 ± 0.2 mg CE/g DT), whereas H. lantanifolia had the lowest (22.0 ± 0.7 mg CE/g DT).
DPPH radical scavenging activity
The antioxidant activities of the three hydroalcoholic extracts measured using the assay of free radical scavenging (DPPH) are presented in Table 4. Ageratina popayanensis showed the highest TEAC value (226), indicating strong antioxidant activity, followed by T. pumilea (105) and H. lantanifolia (93). For the scavenging of DPPH radicals, Ageratina popayanensis exhibited the highest percentage (52%), while T. pumilea and H. lantanifolia showed values of 18% and 22%, respectively. The IC50 values (concentrations required for 50% inhibition) for DPPH scavenging were > 500 µg/mL for T. pumilea (Supplementary Table S3), 496 µg/mL for H. lantanifolia (Supplementary Table S4), and 313 µg/mL for A. popayanensis (Supplementary Table S5), indicating that A. popayanensis had the most potent DPPH scavenging activity among the three extracts.
Photoprotective capacity of hydroalcoholic extracts
The in vitro photoprotective capacity of hydroalcoholic extracts from T. pumilea, H. lantanifolia, and A. popayanensis is summarized in Table 5. The sun protection factor (SPF) values were 41.9 ± 0.1, 38.6 ± 0.7, and 38.2 ± 0.2, respectively, indicating their potential as sunscreens. The extracts also provided long-wavelength UVA protection, with λc values of 385.2 ± 0.1 nm, 380.1 ± 0.2 nm, and 392.4 ± 0.1 nm. The UVA/UVB ratios were 1.6 ± 0.2, 1.5 ± 0.2, and 1.1 ± 0.2, respectively, with A. popayanensis being the most effective at providing maximum protection. The extracts also exhibited low erythema and pigmentation transmission percentages, indicating their potential for quick tan prevention and further supporting their use as sunscreen agents. To validate the results, the photoprotective activity of two sunscreens (commercial products claiming to have SPF 50+) was determined using a 1:1 ratio of sunscreen and absolute ethanol (Supplementary Table S6).
Cytotoxicity and early intracellular ROS in human cancer cell lines
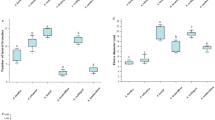
The antiproliferative effects of the extracts were evaluated in the Calu-1 and HepG2 cell lines, and the results are presented in Fig. 2; Table 6. The A. popayanensis extract did not exhibit significant cytotoxicity in either studied cell lines. The IC50 values were 444 µg/mL for Calu-1 cells and 387 µg/mL for HepG2 cells. On the other hand, the IC50 values of 201 µg/mL and 216 µg/mL obtained in HepG2 cells indicated that both the T. pumilea and H. lantanifolia extracts did not exert a relevant antiproliferative effect. However, a more notable antiproliferative effect was observed in Calu-1 cells, with IC50 values of 97 µg/mL for T. pumilea and 116 µg/mL for H. lantanifolia. The IC10 and IC25 values are shown in Supplementary Tables S7-S10.
The evaluation of early intracellular ROS generation (Fig. 3) revealed that, compared with the positive control (H2O2, 200 µM), the extracts did not induce the production of ROS at the concentration evaluated (15.63 µg/mL). However, when comparing the levels of ROS generated by the different extracts, it was evident that after 45 min of exposure, the T. pumilea extract slightly increased the production of intracellular ROS in Calu-1 cells.

Cytotoxicity in Calu-1 and HepG2 cells exposed to hydroalcoholic extracts from T. pumilea, H. lantanifolia, and A. popayanensis for 24 h. *. There was significant difference in viability compared with that of the control group (C-) (p < 0.05). Data are the mean ± SEM (n = 3).

Early intracellular ROS levels in hydroalcoholic extracts of T. pumilea, H. lantanifolia, and A. popayanensis at a concentration of 15.63 µg/mL on Calu-1 (A) and HepG2 (B).
Discussion
The hydroalcoholic extract obtained from the aerial parts of T. pumilea underwent UHPLC-ESI-Orbitrap-HRMS analysis, which led to the identification of various phenolic compounds, flavones, and their glycosylated derivatives. A robust analytical strategy was employed, combining the use of reference substances, literature reports, and spectral databases to achieve a high level of confidence in compound identification. According to the Metabolomics Standards Initiative, five compounds were confirmed using commercial reference standards: gallic acid (≥ 97%), p-coumaric acid (≥ 98%), apigenin-7-glucoside (≥ 90%), apigenin (≥ 95%), and cirsimaritin (≥ 98%). Additional flavones and glycosides were tentatively identified based on literature reports for the genus Turnera and MS spectral interpretation.
To establish the differences and similarities between T. pumilea and its related species, a literature review was conducted. This review led to the identification of T. diffusa as the most prominent species in terms of chemical characterization within the genus, facilitating comparative analysis of the species and enhancing our understanding of their flavonoids and phenolic profiles. The compounds identified include apigenin, pinocembrin, velutin, gonzalitosin I, apigenin 7-O-glucoside, apigenin 7-O-(6”-O-p-E-coumaroyl)-glucoside, apigenin 8-C-(2-O-rhamnosyl)-glucoside, luteolin 8-C-(2-O-rhamnosyl)-glucoside, luteolin 8-C-(2-O-rhamnosyl)-quinovoside, diosmetin 8-C-(2-O-rhamnosyl)-glucoside, chrysoeriol 7-O-glucoside, and a range of other flavonoids and glycoside derivatives43,44,45,46. Additionally, phenolics, phenolic acids, and maltol derivatives such as arbutin, ellagic acid, 4-O-β-D-glucopyranosyl-p-coumaric acid, and maltol 3-O-glucoside have been identified from various sources46,47.
Other interesting compounds reported in T. pumilea (not identified in our study) are cyanogenic glycosides, which constitute a structurally less diverse yet significant class of natural products48. Spencer et al.49identified four cyanogenic glycosides derived from cyclopentenyl glycine: deidaclin, volkenin, tetraphyllin A, and tetraphyllin B. These are defense agents against herbivores and play crucial roles in the early stages of plant development by releasing toxic hydrogen cyanide upon tissue damage48.
According to UHPLC-ESI-Orbitrap-HRMS analysis of compounds in T. pumilea, gallic acid and p-coumaric acid have emerged as noteworthy natural products recognized for their anti-inflammatory and anti-cancer properties50,51and antimelanogenic effects50respectively. Apigenin, a significant flavone in cosmetology, has properties that contribute to the effective treatment of skin inflammatory conditions, UV-induced skin damage, skin aging, and vitiligo52,53along with its anticancer53,54and antiviral attributes55. Chrysoeriol is recognized for its diverse effects including anti-inflammatory, anti-osteoporosis, antibacterial, antifungal, neuroprotective, and anti-cancer properties56. Cirsimaritin, another flavone, demonstrates antioxidant, antimicrobial, antiparasitic, anticancer, antidiabetic, and anti-inflammatory effects57. Apigenin 7-glucoside has been associated with skin anti-inflammatory activity in rats58antimicrobial effects against Staphylococcus aureus and Enterococcus faecalis, as well as inhibitory effects on tyrosinase (TYR-), Acetylcholinesterase (AChE-), and butyrylcholinesterase (BuChE-)59.
A review of T. diffusa highlights the relevance of this species in popular medicine, as it is considered an important therapeutic agent for the ancient Mayan civilization60. In this review, basic information on the in vitro and in vivo biological activities of the extracts, essential oils, and phytocompounds studied by different authors is summarized and presented as antioxidant, antiphotoaging, antimicrobial, cytotoxic, anti-inflammatory, hepatoprotective, nephroprotective, testicular protective, neuroprotective, hypoglycemic/antidiabetic/antihyperglycemic, anti-obesity, analgesic, antidepressant and anxiolytic, aphrodisiac, potent activator of the pregnane-xenobiotic and aryl-hydrocarbon receptors, anti-glycosylation, monoamine oxidases inhibitor, phosphodiesterase 5 inhibitor, anti-aromatase and estrogenic, skin penetration enhancement, and smooth muscle relaxant.
The hydroalcoholic extract of T. pumilea evaluated in our study showed the ability to neutralize DPPH radicals, although with moderate efficacy, given that the concentration required to achieve 50% inhibition (IC50) exceeded 500 µg/mL. At a concentration of 500 µg/mL, the extract achieved 39% radical scavenging, which, although lower than the IC50 threshold, is consistent with the activity reported in previous studies. In particular, Wong-Paz et al.61 observed greater antioxidant capacity (77.9–79.7%) at a higher concentration (1 mg/mL), suggesting an expected dose-response relationship for this type of extract. However, it is important to note that variability in antioxidant activity can be influenced by factors such as the extraction method, the specific chemical composition of the extract, and the total concentration of phenolic compounds present. In this regard, Wong-Paz et al.61 reported a phenolic content of 101.5 mg GAE/g with an EC50 of 305.4 µg/mL, while Tsaltaki et al.62 indicated a maximum phenolic content of 183.7 mg GAE/g DW. Our study recorded a phenolic content of 238.3 mg GAE/g DW at 1 mg/mL, exceeding those previously reported and could reflect differences in the plant source, extraction conditions, or analytical methods used. This disparity between phenolic content and antioxidant capacity suggests that the quantity, quality, composition, and synergy of the phenolic compounds present in the extract influence this activity. Therefore, although the T. pumilea extract demonstrates antioxidant potential, the efficacy observed in the DPPH assay indicates that its activity is moderate compared to other extracts with similar or lower phenolic profiles.
This study presents the in vitro photoprotective efficacy of T. pumilea at a concentration of 0.75 mg/mL. The findings revealed that the extract displayed an SPF of 41.9 and a λc of 385.2 nm, indicative of its broad-spectrum photoprotective capabilities. These results complement the research conducted by Wong-Paz et al.61who investigated the anti-photoaging effects of T. pumilea in HaCaT and HDF cells exposed to UVB radiation. Their study demonstrated the ability of the hydroalcoholic extract to regulate key pathways involved in photoaging, including MMP-1/procollagen type 1, TGF-β1/Smad, and MAPK pathways61.
We evaluated the cytotoxic potential and early intracellular ROS of the hydroalcoholic extract derived from the leaves of T. pumilea against HepG2 (liver cancer) and Calu-1 (lung cancer) cell lines. Our findings revealed that the extract exhibited cytotoxic activity in both cell lines, with IC50 values of 201 µg/mL and 97 µg/mL for HepG2 and Calu-1 cells, respectively. The results indicate that the extract was more potent against Calu-1 cells than against HepG2 cells under the tested conditions. The literature concerning the cytotoxicity of T. pumilea extracts in HepG2 and Calu-1 cells is limited. However, we found two relevant studies that employed different extraction methods. A previous study used aqueous extracts obtained from the leaves and reported a CC50 value of 43.87 µg/mL63. Another study utilized essential oils extracted from the plants and reported CC50 values ranging from 186.5 to 199.3 µg/mL64. Interestingly, these latter results are comparable to our observations using the hydroalcoholic extract. Furthermore, no early intracellular ROS production due to oxidative stress induced by the extract was detected at a concentration of 15.63 µg/mL in HepG2 cells; however, a slight increase in the generation of reactive species was observed in Calu-1 cells. Previous studies have shown that this lung carcinoma cell line responds to various treatments with a significant increase in intracellular ROS, which is associated with the induction of cell death and apoptosis, through the modulation of pathways related to cancer growth, survival, and angiogenesis65. Furthermore, the sensitivity of Calu-1 to cytotoxic agents, such as ABT-263, has been reported to correlate with intracellular ROS levels, suggesting a regulatory role for these species in the response to antitumor treatments66. In this context, moderate ROS generation may act as a physiological signal that modulates oxidative stress and apoptosis pathways, indicating a possible early activation of oxidative stress mechanisms that could contribute to the cytotoxicity of the T. pumilea extract67.
Within the genus Hyptis, the most studied species have been H. verticillata, H. radicans, and H. multibracteata. The limited bibliography about H. lantanifolia leads us to highlight the potent inhibitory activity of the cytopathic effect induced by HIV in cultured cells employing the aqueous extract obtained from its aerial parts, as well as its activity in essential enzymes (HIV-reverse transcriptase and HIV protease enzymes)13,68.
Studies conducted on the genus Hyptis have shown its potential in Brazilian folk medicine11including its anti-inflammatory, antimicrobial, antisecretory properties, anticancer, hepatoprotective, antiviral, acaricidal, insecticidal, molluskicide, and hormonal modulating activities. In addition, many chemical components have been identified, including lignans, triterpenes, diterpenes, sesquiterpenes, monoterpenes, flavonoids, polyphenols, and alkaloids11,12.
UHPLC-ESI-Orbitrap-HRMS analysis of the hydroalcoholic extract of H. lantanifolia leaves led of the identification of caffeic acid (≥ 98%), rosmarinic acid (≥ 97%), and cirsimaritin (≥ 98%) with reference substances. Other flavonoids such as vicenin 2 (apigenin 6,8-C-diglucoside), jaceidin, casticin, sarothrin, isorhamnetin rutinoside, and brickellin were tentatively identified (Table 2). Among of the mentioned compounds, caffeic acid and rosmarinic acid have been previously identified in species within this genus and are considered major constituents of the Lamiaceae family11.
Molecules identified in the H. lantanifolia leaves recognized for their antioxidant, antimicrobial, and anticancer potential. In addition, caffeic acid affects diabetes, atherosclerosis, and Alzheimer’s disease69. Danshensu properties are remarkable in cardiovascular disease, cerebral lesions, and different health conditions, including tumorigenesis and pancreatitis70. Rosmarinic acid affects diabetes, inflammatory and neurodegenerative disorders, and liver diseases71. Vicenin 2 prevents Helicobacter pylori-associated infection and protects the gastric cells72; jaceidin affects Ehrlich’s ascites carcinoma, attenuating tumor progression73; cirsimaritin has remarkable antidiabetic, antiparasitic, and anti-inflammatory effects57; casticin ameliorates scopolamine-induced cognitive dysfunction74 and protects against inflammation in human osteoarthritis chondrocytes75isorhamnetin rutinoside induces the apoptosis of human myelogenous erythroleukemia cells76; and brickellin reduces both the formation of advanced glycation end products (AGEs) and oxidative damage to the proteins77.
The antibacterial activity of different species of the genus Hyptis has been evaluated911. However, H. lantanifolia is highlighted as an antiretroviral agent since the water extract of the aerial parts of this species acts as a potent inhibitor of HIV-RT, with an IC50 value of 7 µg/mL13. In our study, the results of the quantitative analysis of the polyphenols in the hydroalcoholic extract of H. lantanifolia leaves revealed the following total phenolic content: 191.3 mg GAE/g DT > flavonoids: 175.9 mg RE/g DT > condensed tannins: 22.0 mg CE/g DT. Furthermore, the extract showed antioxidant activity, with an IC50 value of 496 µg/mL and sunscreen properties, with an SPF of 38.6 and a λc of 380.1 nm. Additionally, the assessment of their cytotoxic potential revealed that the ability of Calu-1 cells (non-small cell lung cancer) to inhibit proliferation was greater than that of HepG2 cells (human hepatoma), with IC50 values of 116 µg/mL and 216 µg/mL, respectively.
Chemical analysis of the evaluated extract of A. popayanensis revealed the presence of phenolic acids (caffeoyl quinic acid, chlorogenic acid), hydroxycinnamic acid (p-coumaric acid), phenylpropanoids (3,4-di-caffeoylquinic acid and its isomer), a trihydroxy flavone (kaempferol 3-O-rutinoside), and rutin, a glycoside that combines quercetin (flavonol) and rutinoside (Table 3). Some of the identified compounds in this extract are recognized as antioxidant agents, including caffeoyl quinic acid783,4-di-caffeoylquinic acid79and chlorogenic acid, which are also anti-inflammatory, antimicrobial, hypoglycemic, immunomodulatory, anticardiovascular, and antimutagenic80. P-coumaric acid is effective in treating inflammation, diabetes, cardiovascular, and nervous system diseases. Furthermore, p-coumaric acid has shown a positive effect on the prevention and treatment of diseases by inhibiting oxidative stress and exerting anti-aging effects on the skin81. Kaempferol 3-O-rutinoside acts on different targets and is useful in wound healing82. The activities of rutin highlight this glycoside compound as an antimicrobial, antifungal, and antiallergic agent83.
In the evaluation of the three plant species, the hydroalcoholic extract derived from A. popayanensis leaves presented a notably higher concentration of condensed tannins (43.0 ± 0.2 mg CE/g DT). Additionally, this extract exhibited superior scavenging activity against DPPH radicals, with a lower IC50 value of 313 µg/mL and a scavenging rate of 44%. Furthermore, it had minor inhibitory effects on the proliferation of Calu-1 and HepG2 cells.
The cellular permeability of the compounds identified in A. popayanensis has not been extensively studied in cell lines such as Calu-1 or HepG2. However, scientific literature provides relevant data on the permeability of these compounds in Caco-2 cells, a widely used model for simulating the human intestinal barrier84,85,86,87,88,89,90.
These studies demonstrate that some compounds have low apparent permeability, such as chlorogenic acid, caffeoylquinic acid isomers, and 3,4-dicaffeoylquinic acid. Other compounds, including caffeoylquinic acid, caffeic acid, dicaffeoylquinic acid, and rutin, show moderate permeability, while p-coumaric acid exhibits high permeability. It is important to note that the biological effects of these compounds do not depend exclusively on their cellular permeability91,92.
For instance, 3,4-dicaffeoylquinic acid can exert relevant actions through interactions on the cell surface or through its high affinity for plasma proteins, which modulates its bioavailability and pharmacological activity. Furthermore, the same compound (rutin) can exhibit different levels of permeability depending on the cell type and the transport mechanisms involved. In certain cell lines, rutin can demonstrate high permeability mediated by transporters such as P-glycoprotein (P-gp) and the multidrug resistance proteins MRP2 and MRP3, while in Caco-2 cells, the permeability of the same compound may be moderate due to differences in the expression and activity of these transporters. A study of the extract’s permeability in Calu-1 and HepG2 cells will lead to a general permeability profile and allow analysis of the results compared with the existing scientific literature.
Finally, T. pumilea, H. lantanifolia, and A. popayanensis did not exhibit early intracellular ROS production compared with the positive control (H2O2, 200 µM). However, among these species, T. pumilea demonstrated intracellular ROS generation in Calu-1 cells, which could be linked to the observed cytotoxic properties, as reflected by an IC50 value of 97 µg/mL.
Conclusions
The hydroalcoholic extracts of T. pumilea, H. lantanifolia, and A. popayanensis, which belong to the Passifloraceae, Lamiaceae, and Asteraceae families, respectively, were cultivated, collected, and processed following standardized procedures4. The chemical composition of the studied extracts was characterized using UHPLC-ESI-Orbitrap-HRMS analysis. Their activities, including antioxidant capacity, photoprotective effects, cytotoxicity in human cancer cells, and early reactive oxygen species (ROS) production, were assessed through in vitro assays. Collectively, these results highlight the potential of the three evaluated species as promising candidates for antioxidant, photoprotective, and anti-aging applications, particularly in protecting against UV-induced skin damage. Notably, the antiproliferative effects of T. pumilea on non-small cell lung cancer cells underscore its anticancer potential. Further research is needed to elucidate the underlying mechanisms and optimize the use of these botanical extracts in skin care formulations and pharmaceutical products.
Future directions include the evaluation of the antioxidant capacity of the extracts using complementary methods such as ferric reducing antioxidant power (FRAP) and oxygen free radical absorbance capacity (ORAC), which will further demonstrate their applicability in the cosmetic, pharmaceutical, and food industries. Furthermore, the inclusion of normal and cancerous human cell lines, as well as in vivo studies, is crucial to identify potential adverse effects and ensure therapeutic safety and efficacy. The moderate ROS generation induced by T. pumilea extract in Calu-1 cells suggests a controlled pro-oxidant mechanism that triggers oxidative stress and apoptosis. Future studies should focus on the regulation of these pathways and their impact on cell viability to clarify the role of ROS in the observed cytotoxic effects.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Stashenko, E. & Martínez, R. Study of essential oils obtained from Tropical plants grown in Colombia. In: Essent Oils-Oils Nat. Intechopen, London United Kingdon. pp. 403–451. (2019). https://doi.org/10.5772/intechopen.87199
Quintero-Rincón, P. et al. Exploring the potential of extracts from Sloanea medusula and S. calva: formulating two skincare gels with antioxidant, sun protective factor, and anti-Candida albicans activities. Pharmaceuticals 16, 990. https://doi.org/10.3390/ph16070990 (2023).
Quintero-Rincón, P. et al. Sloanea Chocoana and S. pittieriana (Elaeocarpaceae): chemical and biological studies of ethanolic extracts and skincare properties. Plants 12, 3953. https://doi.org/10.3390/plants12233953 (2023).
Caballero-Gallardo, K., Alvarez-Ortega, N. & Olivero-Verbel, J. Cytotoxicity of nine medicinal plants from San Basilio de Palenque (Colombia) on HepG2 cells. Plants 12, 2686. https://doi.org/10.3390/plants12142686 (2023).
Huddart, J. E. A., Crawford, A. J., Luna-Tapia, A. L., Restrepo, S. & Di Palma, F. EBP-Colombia and the bioeconomy: genomics in the service of biodiversity conservation and sustainable development. Proc. Natl. Acad. Sci. U.S.A. 119 (4), e2115641119. https://doi.org/10.1073/pnas.2115641119 (2022).
Stashenko, E. & Martínez, J. R. The expression of biodiversity in the secondary metabolites of aromatic plants and flowers growing in Colombia. Potential of essential oils, Hany A. El-Shemy, Chapther 4. pp 59–86 (2018). https://doi.org/10.5772/intechopen.78001
Caballero-Gallardo, K., Quintero-Rincón, P., Stashenko, E. E. & Olivero-Verbel, J. Photoprotective agents obtained from aromatic plants grown in Colombia: Total phenolic content, antioxidant activity, and assessment of cytotoxic potential in cancer cell lines of Cymbopogon flexuosus L. and Tagetes lucida Cav. essential oils. Plants 11, 1693 (2022). https://doi.org/10.3390/plants11131693
Olivero-Verbel, J., Quintero-Rincón, P. & Caballero-Gallardo, K. Aromatic plants as cosmeceuticals: benefits and applications for skin health. Planta 260, 132. https://doi.org/10.1007/s00425-024-04550-8 (2024).
Olivero-Verbel, J., Alcala-Orozco, M., Caballero-Gallardo, K. & Stashenko, E. & Gonzalez Mina, R. Catálogo de plantas de la flora colombiana con propiedades cosmecéuticas. Editorial Universitaria. ISBN: 978-958-5439-65-8 (2024).
Szewczyk, K. & Zidorn, C. Ethnobotany, phytochemistry, and bioactivity of the genus Turnera (Passifloraceae) with a focus on damiana–Turnera diffusa. J. Ethnopharmacol. 152 (3), 424–443. https://doi.org/10.1016/j.jep.2014.01.019 (2014).
Sedano-Partida, M. D., Santos, K. P., Sala-Carvalho, W. R., Silva-Luz, C. L. & Furlan, C. M. Anti-HIV-1 and antibacterial potential of Hyptis radicans (Pohl) Harley & J.F.B. Pastore and Hyptis multibracteata Benth. (Lamiaceae). J. Herb. Med. 20,100328 (2020). https://doi.org/10.1016/j.hermed.2019.100328
Picking, D., Delgoda, R., Boulogne, I. & Mitchell, S. Hyptis verticillata jacq: a review of its traditional uses, phytochemistry, Pharmacology and toxicology. J. Ethnopharmacol. 147 (1), 16–41. https://doi.org/10.1016/j.jep.2013.01.039 (2013).
Matsuse, I. T., Lim, Y. A., Hattori, M., Correa, M. & Gupta, M. P. A search for anti-viral properties in Panamanian medicinal plants. The effects on HIV and its essential enzymes. J. Ethnopharmacol. 64 (1), 15–22. https://doi.org/10.1016/S0378-8741(98)00099-3 (1999).
Rivero-Cruz, I., Gutiérrez-González, J. A., Pérez-Vásquez, A., Villaseñor, J. L. & Mata, R. The genus Ageratina (Asteraceae) in america: an insight into its chemistry and Pharmacological potential. Comb. Chem. High. Throughput Screen. 26, 1629–1659. https://doi.org/10.2174/1386207325666220908093541 (2023).
Sánchez-Gutiérrez, J. A., Moreno-Lorenzana, D., Álvarez-Bernal, D., Rodríguez-Campos, J. & Medina-Medrano, J. R. Phenolic profile, antioxidant and anti-proliferative activities of methanolic extracts from Asclepias linaria cav. Leaves. Molecules 25 (1), 54. https://doi.org/10.3390/molecules25010054 (2019).
Quintero-Rincón, P., Pájaro-González, Y. & Diaz-Castillo, F. Maclura tinctoria (L.) D. Don ex steud. (Moraceae): a review of the advances in ethnobotanical knowledge, phytochemical composition, and Pharmacological potential. Adv. Tradit Med. 25, 331–351. https://doi.org/10.1007/s13596-024-00764-z (2025).
Alvarez-Ortega, N. et al. Protective effects of the hydroethanolic extract of Fridericia Chica on undifferentiated human neuroblastoma cells exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL). Toxins 13, 748. https://doi.org/10.3390/toxins13110748 (2021).
Quintero-Rincón, P., Caballero-Gallardo, K. & Olivero-Verbel, J. Natural anticancer agents: prospection of medicinal and aromatic plants in modern chemoprevention and chemotherapy. Nat. Prod. Bioprospect. 15 (1), 25. https://doi.org/10.1007/s13659-025-00511-0 (2025).
Gulcin, İ. & Alwasel, S. H. DPPH radical scavenging assay. Processes 11 (8), 2248. https://doi.org/10.3390/pr11082248 (2023).
Li, L. et al. Natural products and extracts from plants as natural UV filters for sunscreens: A review. Anim. Models Exp. Med. 6 (3), 183–195. https://doi.org/10.1002/ame2.12295) (2023).
Moskwa, J., Bronikowska, M. & Socha, K. Markiewicz-Żukowska, R. Vegetable as a source of bioactive compounds with photoprotective properties: implication in the aging process. Nutrients 15 (16), 3594. https://doi.org/10.3390/nu15163594 (2023).
Mansur, J., Breder, M., Mansur, M. & Azulay, R. Determination of sun protecting factor in human beings and by spectrophotometry: comparison between of the two methods. Bras. Dermatol. 61, 167–172 (1986).
Panyakaew, J. et al. Kaempferia sp. extracts as UV protecting and antioxidant agents in sunscreen. J. Herbs Spices Med. P. 27, 37–56. https://doi.org/10.1080/10496475.2020.1777614 (2021).
Karunarathne, W. et al. Protective effect of anthocyanin-enriched polyphenols from Hibiscus syriacus L. (Malvaceae) against ultraviolet B-induced damage. Antioxidants 10 (4), 584. https://doi.org/10.3390/antiox10040584 (2021).
Jain, A. K. et al. Chapter 3 - Models and methods for in vitro toxicity. In vitro toxicology (pp. 45–65). Academic Press. ISBN 9780128046678, (2018). https://doi.org/10.1016/B978-0-12-804667-8.00003-1
Sinosaki, N. et al. Structural study of phenolic acids by triple quadrupole mass spectrometry with electrospray ionization in negative mode and H/D isotopic exchange. J. Braz Chem. Soc. 31 (2), 402–408. https://doi.org/10.21577/0103-5053.20190197 (2020).
Geng, P. et al. Comprehensive characterization of C-glycosyl flavones in wheat (Triticum aestivum L.) germ using UPLC‐PDA‐ESI/HRMSn and mass defect filtering. J. Mass. Spectrom. 51, 914–930. https://doi.org/10.1002/jms.3803 (2016).
Colombo, R., Yariwake, J. H. & McCullagh, M. Study of C-and O-glycosylflavones in sugarcane extracts using liquid chromatography: exact mass measurement mass spectrometry. J. Braz Chem. Soc. 19 (3), 483–490. https://doi.org/10.1590/S0103-50532008000300016 (2008).
Abad-García, B., Garmón‐Lobato, S., Berrueta, L. A., Gallo, B. & Vicente, F. New features on the fragmentation and differentiation of C‐glycosidic flavone isomers by positive electrospray ionization and triple quadrupole mass spectrometry. Rapid Commun. Mass. Spectrom. 22 (12), 1834–1842. https://doi.org/10.1002/rcm.3560 (2008).
Quirantes-Piné, R. et al. HPLC–ESI–QTOF–MS as a powerful analytical tool for characterising phenolic compounds in olive‐leaf extracts. Phytochem Anal. 24 (3), 213–223. https://doi.org/10.1002/pca.2401 (2013).
March, R. E. et al. A comparison of flavonoid glycosides by electrospray tandem mass spectrometry. Int. J. Mass. Spectrom. 248 (1–2), 61–85. https://doi.org/10.1016/j.ijms.2005.09.011 (2006).
Bezerra, A. G., Negri, G., Duarte-Almeida, J. M., Smaili, S. S. & Carlini, E. A. Phytochemical analysis of hydroethanolic extract of Turnera diffusa willd and evaluation of its effects on astrocyte cell death. Einstein 14 (1), 56–63. https://doi.org/10.1590/S1679-45082016AO3386 (2016).
Córdoba, Y. & Stashenko, E. E. Estudio de la composición química y las actividades antioxidante y fotoprotectora de los extractos de damiana y orégano de monte, cultivadas de Santander. Tesis doctoral. Universidad Industrial de Santander. Tesis doctoral. Universidad Industrial de Santander. Repositorio Educativo Digital UIS, 96–101. (2019).
Wishart, D. S. et al. The human metabolome database. Nucleic Acids Res on-line: (2009). https://hmdb.ca/
Sen, A., Ozbas Turan, S. & Bitis, L. Bioactivity-guided isolation of anti-proliferative compounds from endemic Centaurea Kilaea. Pharm. Biol. 55 (1), 541–546. https://doi.org/10.1080/13880209.2016.1255980 (2017).
Silva, J. N. B. et al. Caryocar brasiliense camb. Fruit Peel butanolic fraction induces antiproliferative effects against murine melanoma cell line. Phytomed Plus. 2 (2), 100273. https://doi.org/10.1016/j.phyplu.2022.100273 (2022).
MoNA. Mass Bank of North America. Disponible on-line: https://mona.fiehnlab.ucdavis.edu/
NIST. The NIST mass spectral search program. Standard reference data program of the national institute of standards and technology, USA. (2017).
Falcao, R. A. et al. Antileishmanial phenylpropanoids from the leaves of Hyptis pectinata (L.) Poit. Evid. Based Complement. Alternat. Med. 460613. (2013). https://doi.org/10.1155/2013/460613 (2013).
Zhang, M. et al. Bioactive Quinic acid derivatives from Ageratina adenophora. Molecules 18 (11), 14096–14104. https://doi.org/10.3390/molecules181114096 (2013).
Arciniegas, A. et al. Chemical composition and antimicrobial activity of Ageratina deltoidea. Chem. Biodivers. 15 (3), e1700529. https://doi.org/10.1002/cbdv.201700529 (2018).
Dokalahy, E. E., El-Seedi, H. & Farag, M. A. Soft coral biodiversity in the red sea family Alcyoniidae: A biopharmaceutical and ecological perspective. In: Ramawat, K. (eds) Biodiversity and Chemotaxonomy. Sustainable development and biodiversity, Springer, Cham. vol 24. pp. 55–85. (2019). https://doi.org/10.1007/978-3-030-30746-2_4
Piacente, S. et al. Flavonoids and arbutin from Turnera diffusa. Naturforsch C J. Biosci. 57 (11–12), 983–985. https://doi.org/10.1515/znc-2002-11-1204 (2002).
Kumar, S., Taneja, R. & Sharma, A. The genus turnera: A review update. Pharm. Biol. 43, 383–391 (2005). https://doi.org/10.1080/13880200590962926 (2002).
Kumar, S., Sharma, A. & Apigenin The anxiolytic constituent of Turnera aphrodisiaca. Pharm. Biol. 44, 84–90. https://doi.org/10.1080/13880200600591758 (2006).
Zhao, J., Pawar, R. S., Ali, Z. & Khan, I. A. Phytochemical investigation of Turnera diffusa. J. Nat. Prod. 70, 289–292. https://doi.org/10.1021/np060253r (2007).
Aguilera-Carbo, A., Augur, C., Prado-Barragan, L., Aguilar, C. & Favela-Torres, E. Extraction and analysis of ellagic acid from novel complex sources. Chem. Pap. 62, 440–444. https://doi.org/10.2478/s11696-008-0042-y (2008).
Yulvianti, M. & Zidorn, C. Chemical diversity of plant cyanogenic glycosides: an overview of reported natural products. Molecules 26 (3), 719. https://doi.org/10.3390/molecules26030719 (2021).
Spencer, K., Seigler, D. & Fraley, S. Cyanogenic glycosides of the Turneraceae. Biochem. Syst. Ecol. 13, 433–435. https://doi.org/10.1016/0305-1978(85)90088-2 (1985).
Boo, Y. C. -Coumaric acid as an active ingredient in cosmetics: A review focusing on its antimelanogenic effects. Antioxidants 8, 275. https://doi.org/10.3390/antiox8080275 (2019).
Jiang, Y. et al. Gallic acid: a potential anti-cancer agent. Chin. J. Integr. Med. 28, 661–671. https://doi.org/10.1007/s11655-021-3345-2 (2022).
Majma Sanaye, P., Mojaveri, M. R., Ahmadian, R., Sabet Jahromi, M. & Bahramsoltani, R. Apigenin and its dermatological applications: A comprehensive review. Phytochemistry 203, 113390. https://doi.org/10.1016/j.phytochem.2022.113390 (2022).
Yoon, J. H., Kim, M. Y., Cho, J. Y. & Apigenin A therapeutic agent for treatment of skin inflammatory diseases and cancer. Int. J. Mol. Sci. 24, 1498. https://doi.org/10.3390/ijms24021498 (2023).
Imran, M. et al. Apigenin as an anticancer agent. Phytother Res. 34, 1812–1828 (2020).
Lee, I. G., Lee, J., Hong, S. H. & Seo, Y. J. Apigenin’s therapeutic potential against viral infection. Front. Biosci. 28, 237. https://doi.org/10.1002/ptr.6647 (2023).
Aboulaghras, S. et al. Health benefits and Pharmacological aspects of chrysoeriol. Pharmaceuticals 15, 973. https://doi.org/10.3390/ph15080973 (2022).
Benali, T. et al. The current Etate of knowledge in biological properties of cirsimaritin. Antioxidants 11, 1842. https://doi.org/10.3390/antiox11091842 (2022).
Fuchs, J. & Milbradt, R. Skin anti-inflammatory activity of apigenin-7-glucoside in rats. Arzneimittelforschung 43, 370–372 PMID: 7683883 (1993).
Sezen Karaoğlan, E., Hancı, H., Koca, M. & Kazaz, C. Some bioactivities of isolated apigenin-7-O-glucoside and luteolin-7-O-glucoside. Appl. Sci. 13 (3), 1503. https://doi.org/10.3390/app13031503 (2023).
Parra-Naranjo, A., Delgado-Montemayor, C. & Salazar-Aranda, R. Waksman-Minsky, N. Bioactivity of the genus turnera: A review of the last 10 years. Pharmaceuticals 16, 1573. https://doi.org/10.3390/ph16111573 (2023).
Wong-Paz, J. E., Muñiz-Márquez, D. B., Aguilar-Zárate, P., Rodríguez-Herrera, R. & Aguilar, C. N. Microplate quantification of total phenolic content from plant extracts obtained by conventional and ultrasound methods. Phytochem Anal. 25, 439–444. https://doi.org/10.1002/pca.2512 (2014).
Tsaltaki, C., Katsouli, M., Kekes, T., Chanioti, S. & Tzia, C. Comparison study for the recovery of bioactive compounds from Tribulus terrestris, Panax ginseng, Gingko biloba, Lepidium meyenii, Turnera diffusa and Withania somnifera by using microwave-assisted, ultrasound-assisted and conventional extraction methods. Ind. Crops Prod. 142, 111875. https://doi.org/10.1016/j.indcrop.2019.111875 (2019).
Bernardo, J., Ferreres, F., Gil-Izquierdo, Á., Valentão, P. & Andrade, P. B. Medicinal species as mtdls: Turnera diffusa willd. Ex Schult inhibits CNS enzymes and delays glutamate excitotoxicity in SH-SY5Y cells via oxidative damage. Food Chem. Toxicol. 106, 466–476. https://doi.org/10.1016/j.fct.2017.06.014 (2017).
Velandia, S. A., Flechas, M. C., Stashenko, E. E. & Ocazionez, R. E. Proposal to select essential oils from Colombian plants for research based on its cytoxicity. Vitae 23 (1), 18–29. https://doi.org/10.17533/vitae.v23n1a03 (2016).
Zhou, Y., Zhang, Y., Bao, J., Chen, J. & Song, W. Low temperature plasma suppresses lung cancer cells growth via VEGF/VEGFR2/RAS/ERK axis. Molecules 27 (18), 5934. https://doi.org/10.3390/molecules27185934 (2022).
Ohgino, K. et al. Intracellular levels of reactive oxygen species correlate with ABT-263 sensitivity in non-small-cell lung cancer cells. Cancer Sci. 111 (10), 3793–3801. https://doi.org/10.1111/cas.14569 (2020).
An, X. et al. Oxidative cell death in cancer: mechanisms and therapeutic opportunities. Cell. Death Dis. 15 (8), 556. https://doi.org/10.1038/s41419-024-06939-5 (2024).
Lim, Y. et al. Inhibitory effects of some Panamanian plants on human immunodeficiency viral reverse transcriptase and protease. J. Trad Med. 14, 54–58 (1997).
Pavlíková, N. Caffeic acid and diseases-mechanisms of action. Int. J. Mol. Sci. 24, 588. https://doi.org/10.3390/ijms24010588 (2022).
Zhang, J., Zhang, Q., Liu, G. & Zhang, N. Therapeutic potentials and mechanisms of the Chinese traditional medicine Danshensu. Eur. J. Pharmacol. 864, 172710. https://doi.org/10.1016/j.ejphar.2019.172710 (2019).
Noor, S. et al. Biomedical features and therapeutic potential of Rosmarinic acid. Arch. Pharm. Res. 45, 205–228. https://doi.org/10.1007/s12272-022-01378-2 (2022).
Zhang, Y., Sun, J., Dong, Y., Shen, X. & Zhang, Z. Vicenin-2 inhibits the Helicobacterium pylori infection associated gastric carcinogenic events through modulation of PI3K/AKT and Nrf2 signaling in GES-1 cells. J. Biochem. Mol. Toxicol. 35 (3), e22680. https://doi.org/10.1002/jbt.22680 (2021).
Elhady, S. S. et al. Jaceidin flavonoid isolated from Chiliadenus Montanus attenuates tumor progression in mice via VEGF inhibition: in vivo and in Silico studies. Plants 9, 1031. https://doi.org/10.3390/plants9081031 (2020).
Kim, J. et al. Casticin ameliorates scopolamine-induced cognitive dysfunction in mice. J. Ethnopharmacol. 259, 112843. https://doi.org/10.1016/j.jep.2020.112843 (2020).
Mu, Y., Hao, W. & Li, S. Casticin protects against IL-1β-induced inflammation in human osteoarthritis chondrocytes. Eur. J. Pharmacol. 842, 314–320. https://doi.org/10.1016/j.ejphar.2018.10.051 (2019).
Boubaker, J. et al. Ethyl acetate extract and its major constituent, Isorhamnetin 3-O-rutinoside, from Nitraria retusa leaves, promote apoptosis of human myelogenous erythroleukaemia cells. Cell. Prolif. 44, 453–461. https://doi.org/10.1111/j.1365-2184.2011.00772.x (2011).
Rodrigues, W. D., Cardoso, F. N., Baviera, A. M. & Dos Santos, A. G. In vitro antiglycation potential of erva-baleeira (Varronia Curassavica Jacq). Antioxidants 12, 522. https://doi.org/10.3390/antiox12020522 (2023).
Hung, T. M. et al. Antioxidant activity of caffeoyl Quinic acid derivatives from the roots of Dipsacus Asper wall. J. Ethnopharmacol. 108, 188–192. https://doi.org/10.1016/j.jep.2006.04.029 (2006).
Nakajima, Y., Shimazawa, M., Mishima, S. & Hara, H. Water extract of propolis and its main constituents, caffeoylquinic acid derivatives, exert neuroprotective effects via antioxidant actions. Life Sci. 80, 370–377. https://doi.org/10.1016/j.lfs.2006.09.017 (2007).
Miao, M. & Xiang, L. Pharmacological action and potential targets of chlorogenic acid. Adv. Pharmacol. 87, 71–88. https://doi.org/10.1016/bs.apha.2019.12.002 (2020).
Chen, F., Zhang, X., Wang, J., Wang, F. & Mao, J. -coumaric acid: advances in Pharmacological research based on oxidative stress. Curr. Top. Med. Chem. 24, 416–436. https://doi.org/10.2174/0115680266276823231230183519 (2024).
Petpiroon, N., Suktap, C., Pongsamart, S., Chanvorachote, P. & Sukrong, S. Kaempferol-3-O-rutinoside from Afgekia mahidoliae promotes keratinocyte migration through FAK and Rac1 activation. J. Nat. Med. 69, 340–348. https://doi.org/10.1007/s11418-015-0899-3 (2015).
Negahdari, R. et al. Therapeutic benefits of Rutin and its nanoformulations. Phytother Res. 35, 1719–1738. https://doi.org/10.1002/ptr.6904 (2021).
Botti, G. et al. Chlorogenic acid permeation across intestinal cell monolayers: influence by circadian rhythms in the presence of other natural polyphenols and by dopaminergic neuronal-like cells. J. Funct. Foods. 119, 106331. https://doi.org/10.1016/j.jff.2024.106331 (2024).
Li, X. et al. Antioxidant and cytoprotective effects of the Di-O-Caffeoylquinic acid family: the mechanism, structure-activity relationship, and conformational effect. Molecules 23 (1), 222. https://doi.org/10.3390/molecules23010222 (2018).
Hong, Y. J., Yang, S. Y., Nam, M. H., Koo, Y. C. & Lee, K. W. Caffeic acid inhibits the uptake of 2-amino-1-methyl-6-phenylimidazo [4, 5-b] pyridine (PhIP) by inducing the efflux transporters expression in Caco-2 cells. Biol. Pharm. Bull. 38 (2), 201–207. https://doi.org/10.1248/bpb.b14-00495 (2015).
Konishi, Y., Kobayashi, S. & Shimizu, M. Transepithelial transport of p-coumaric acid and Gallic acid in Caco-2 cell monolayers. Biosci. Biotechnol. Biochem. 67 (11), 2317–2324. https://doi.org/10.1271/bbb.67.2317 (2003).
Zhang, X. et al. Absorption and metabolism characteristics of Rutin in Caco-2 cells. ScientificWorldJournal 2013, 382350. https://doi.org/10.1155/2013/382350 (2013).
Hufnagel, M., Rademaekers, A., Weisert, A., Häberlein, H. & Franken, S. Pharmacological profile of Dicaffeoylquinic acids and their role in the treatment of respiratory diseases. Front. Pharmacol. 15, 1371613. https://doi.org/10.3389/fphar.2024.1371613 (2024).
Ma, Y. et al. Intestinal absorption and neuroprotective effects of kaempferol-3-O-rutinoside. RSC Adv. 7 (50), 31408–31416. https://doi.org/10.1039/C7RA05415G (2017).
Jana, S. & Rastogi, H. Effects of caffeic acid and Quercetin on in vitro permeability, metabolism and in vivo pharmacokinetics of melatonin in rats: potential for Herb-Drug interaction. Eur. J. Drug Metab. Pharmacokinet. 42 (5), 781–791. https://doi.org/10.1007/s13318-016-0393-7 (2017).
Hsieh, C. F. et al. 3, 4-Dicaffeoylquinic acid from the medicinal plant Ilex Kaushue disrupts the interaction between the five-fold axis of enterovirus A-71 and the Heparan sulfate receptor. J. Virol. 96 (7), e00542–e00521. https://doi.org/10.1128/jvi.00542-21 (2022).
Funding
This research was funded by the Ministry of Science, Technology and Innovation (Minciencias), the Ministry of Education, the Ministry of Industry, Commerce and Tourism, and ICETEX, Programme Ecosistema Científico–Colombia Científica, from the Francisco José de Caldas Fund, Grant RC-FP44842-212-2018; The Ministry of Environment and Sustainable Development of Colombia supported the Universidad Industrial de Santander through access permits to genetic resources and derivatives for bioprospecting (Contract No. 270–2019); and Minciencias - Programa Orquídeas, mujeres en la ciencia: agentes para la paz (Call No. 935–2023) through the Francisco José de Caldas Fund and from the municipality of Zambrano Bolivar, (Grant RC 112721-262-2023); and University of Cartagena (Resolution No.02-2024).
Author information
Authors and Affiliations
Contributions
Conceptualization, K.C.-G. and P.Q.-R.; methodology, E.E.-S., K.C.-G., J.O.-V., and P.Q.-R.; formal analysis, P.Q.-R., K.C.-G. and J.O.-V.; investigation, J.O.-V., and E.E.-S., and P.Q.-R.; resources, K.C.-G. and J.O.-V.; data curation, P.Q.-R.; writing—original draft preparation, P.Q.-R.; writing—review and editing, P.Q.-R., K.C.-G. and J.O.-V.; supervision, K.C.-G. and J.O.-V.; project administration E.E.-S. and J.O.-V.; funding acquisition, E.E.-S. and J.O.-V. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that there are no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Caballero-Gallardo, K., Quintero-Rincón, P., Stashenko, E.E. et al. Chemical composition and in vitro bioactivities of hydroalcoholic extracts from Turnera pumilea, Hyptis lantanifolia, and Ageratina popayanensis cultivated in Colombia. Sci Rep 15, 31189 (2025). https://doi.org/10.1038/s41598-025-12778-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12778-x
