
Abstract
Given the widespread conflicts between humans and birds in contexts such as agricultural production, airports, and new energy installations, the development and application of bird deterrence technologies hold significant importance for both safeguarding human interests and advancing the cause of wildlife conservation. Artificial sounds are one of the most widely used methods for bird deterrence; however, there is a lack of systematic research on the effects of sound frequency and their combination. The black-necked crane (Grus nigricollis), a first-class nationally protected wild animal of China, shares some of its habitats with human agricultural activities. This study designed sounds of various frequencies and combinations and conducted deterrence experiments on black-necked cranes at their wintering sites in northeastern Yunnan Province, exploring how sound frequency and combinations affect the behavioral responses, escape distances, and escape speeds of the cranes. Our results show that: (1) in terms of behavioral response intensity, escape distances, and escape speed, sounds with high frequency and high variability significantly outperformed other sounds (n = 479, p < 0.001); (2) there were no significant differences in the responses of black-necked cranes of different flock types and age combinations to sounds. The study recommends using high-frequency, highly variable sounds for short-term control of black-necked cranes. Additionally, this research demonstrates that using either high-frequency or high variable sound combinations can achieve efficient bird deterrence in the short term, and provides a scientific basis for developing and refining bird deterrence strategies for other bird species.
Similar content being viewed by others

Introduction
In pursuit of global sustainable development, “protecting animals to maintain harmonious coexistence between humans and nature” has emerged as a prerequisite1. With the expansion of human activities, the living spaces of wildlife have been compressed, leading to intensified conflicts between humans and animals2. This not only jeopardizes the conservation of wildlife but also threatens the safety of human production and livelihoods. Birds are a class of animals intricately linked to human life, profoundly influencing fields such as aviation, agriculture, and urban development3. For example, birds may consume large portions of crops, potentially causing total harvest failures4; birds may also spread diseases and affect environmental hygiene, posing a threat to people’s health5,6. The adoption of avian deterrent methods enables the mitigation of avian-induced disturbances and economic losses while simultaneously supporting bird conservation efforts7. With technological advancements, the approaches used for bird deterrence have become increasingly diverse. Currently, bird deterrence technologies mainly include visual deterrents, acoustic devices, lighting systems, olfactory repellents, and protective netting8,9. Among these, acoustic deterrence stands as one of the most widely adopted strategies10. Sounds employed in bird repellents include gunshots11,12,13, firework and firecracker explosions14, propane cannons15,16, predator vocalizations specific to target species17, conspecific alarm or distress calls18, as well as irregular electromagnetic sounds, and other aversive high-frequency sounds19. Although birds have been found to quickly adapt or return to the area once the sound stops20, acoustic deterrence remains a prevalent countermeasure due to its immediate efficacy and operational practicality21,22.
Previous studies have found that many factors can influence the response patterns of birds to sound. Some studies have indicated that many birds are sensitive to sound frequency, and high-frequency sounds close to the auditory limits of birds—even ultrasonic waves—have been widely incorporated into the design of bird repellent devices23. Conversely, other research has shown that birds exhibit strong reactions to low-frequency sounds, such as fire alarms24 or the sounds that overlap with the frequency range of bird vocalizations25. The pattern of sound combinations, including the extent of frequency change, the transition speed between different frequencies, and the presence of regularity, may all affect bird repellent effectiveness23. In addition, bird repellent devices are typically designed to target multiple species; however, variations in species-specific responses26, flock composition, and age cohorts27,28 can result in significantly different reactions to the same auditory stimuli. Therefore, it is necessary to conduct experiments targeting specific species in particular bird deterrence scenarios to test the repellent effects of different sound frequency combinations, thereby providing valuable references for the design of bird repellent devices.
The black-necked crane (Grus nigricollis) is a Class I protected wild animal in China and is categorized as Near Threatened (NT) by the International Union for Conservation of Nature (IUCN)29. It is mainly distributed in the Qinghai-Tibet Plateau and Yunnan-Guizhou Plateau30, and inhabit at plateau marshes, lakes, riverbanks, and agricultural land31,32. During the wintering period, black-necked cranes often feed in farmland to consume potatoes, barley, buckwheat, oats, radishes, and grass roots33. This foraging behavior can lead to crop damage, particularly as the population size increases30 and the activity range expands34. Consequently, the conflict between humans and cranes over food and land is increasingly prominent35. In addition, during the migration season, cranes face the risk of collisions with wind turbines and high-voltage power lines commonly found on the plateau36. Therefore, it is necessary to adopt appropriate measures to encourage avoidance behaviors during critical sowing and migration periods, thereby alleviating conflicts between humans and black-necked cranes.
We designed a series of sound frequencies and combinations and conducted behavioral experiments in a black-necked crane wintering area in northeastern Yunnan Province. This study aims to explore how the frequency and combination of sounds affect the behavior, escape time, escape distance, and escape speed of black-necked cranes. Additionally, we explore whether different flock types and ages exhibit varied responses to these auditory stimuli. This study will provide scientific recommendations for the local conservation of black-necked cranes and also offer a reference for the development and improvement of bird repellent strategies for other bird species.
Methods
Study area
The study area is located in Daxiandang Village, Wuzhai Township, Yongshan County, Zhaotong City, northeastern Yunnan Province, China (N27°36′32″-27°35′18″, E103°24′2″-103°27′16″) (Fig. 1, created using ArcGIS 10.8, available at: https://www.esri.com/). The region exhibits a typical alpine climate and is prone to frost and ice, with an annual average temperature of 6.2 °C. The average temperature in the hottest month, July, is 20 °C, while falls to -5 °C in the coldest month, january. The area experiences an average of 60.6 frosty days and a frost period of 242 days, and 184.8 foggy days annually. The predominant vegetation type in the region is alpine meadow, with buckwheat, oats, and potatoes being the primary crops cultivated.

Map of the study area in Daxiandang Village, Wuzhai Township, Yongshan County, Yunnan Province, China (27°36′32″–27°35′18″N, 103°24′2″–103°27′16″E).
This location serves as a stable wintering ground for black-necked cranes, with the wintering period extending from late October to mid-April of the following year35. The experiments for this study were conducted from March 10 to March 22, 2024. During this period, we observed between 30 and 85 black-necked cranes (with a typical daily average of 57 individuals) residing in the area each day. During the day, the cranes predominantly engaged in activities as either non-breeding flocks (flock) or family groups. The Flock typically consisted of 10 to 55 individuals, all of which were non-breeding individuals aged 2 years or older. The family groups ranged from 2 to 5 individuals, composed of two adult cranes and one to three subadults (Juvenile). The flock cranes frequently split and reformed, with variable numbers and unstable activity range. In contrast, family groups generally had stable membership compositions and home range.
Sound design
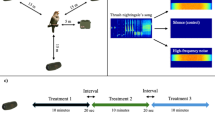
The auditory thresholds of birds encompass a frequency range from 1 to 15,000 Hz. In this study, three pure tone frequencies were selected: 200 Hz, 6,400 Hz, and 12,800 Hz. The 200 Hz frequency corresponds to the lower threshold of avian hearing and simulates noise produced by wind turbine infrastructure, which is characterized by dominant frequencies below 2,000 Hz. The 6,400 Hz frequency falls within the peak sensitivity range of birds, corresponding to the frequency band where the energy of natural alarm calls is most concentrated and overlapping with the spectral range of anthropogenic noise pollution. The 12,800 Hz frequency was chosen to evaluate the upper functional limit of avian hearing.
A total of four types of sounds were prepared (Fig. 2) Type A represents a continuous constant frequency sound, for which we selected three frequencies in ascending order: 200 Hz - Constant (Group A1), 6400 Hz - Constant (A2), and 12,800 Hz - Constant (A3). Type B was designed to assess the impact of frequency range. It included three subgroups where frequencies of 200 and 6400 Hz - Alternating (B1), 6400 and 12,800 Hz - Alternating (B2), and 200 and 12,800 Hz - Alternating (B3), with alternated every 5 s (Fig. 2B). Type C featured audio fluctuating within a specific range. The three subgroups were 200–6400 Hz - Rapid Sweep, 6400–12,800 Hz - Rapid Sweep, and 200–12,800 Hz - Rapid Sweep. Within each range, five frequencies were selected from low to high, each continued for 1/6 of a second. Group D used the same frequency combinations as Group C, but each audio was played for one second, with each set lasting 5 s followed by a 10-second pause before repeating the cycle. The three subgroups were 200–6400 Hz - Stepped Frequency, 6400–12,800 Hz - Stepped Frequency, and 200–12,800 Hz - Stepped Frequency.Sound designs were created using Adobe Audition 2022 software, which involved combining single audio tracks.

Acoustic spectrograms of experiment sound types. A1.200 Hz - Constant; A2.6400 Hz - Constant; A3.12800 Hz - Constant; B1.200–6400 Hz - Alternating; B2.6400–12,800 Hz - Alternating; B3.200–12,800 Hz - Alternating; C1.200–6400 Hz - Rapid Sweep; C2.6400–12,800 Hz - Rapid Sweep; C3.200–12,800 Hz - Rapid Sweep; D1.200–6400 Hz - Stepped Frequency; D2.6400–12,800 Hz - Stepped Frequency; D3.200–12,800 Hz - Stepped Frequency.
Field experiments
The experiment was conducted in agriculture land and wetlands where black-necked cranes were foraging. The audio was played using a standard portable loudspeaker (Shenzhen Oppo Technology Co., Ltd., Shenzhen, China). Prior observations were used to predict the areas where cranes would forage, and the device was positioned within the area before the cranes entered. During the placement of the device, efforts were made to utilize concealed routes to avoid entering the visual field of the black-necked cranes. The sound was initiated when black-necked cranes were within 10 m of the speaker, with each sound played for one minute, with a minimum interval of 30 min between experiments. The experimental groups were conducted in a crossover manner.
Prior to sound playback, the type of sound, time, group type, and the number of adults and juveniles were recorded. When the sound playback commenced, observations were made on the age characteristics of the subjects, all behaviors and events from the moment the audio began to the end of playback, and the escape distance. Crane behavioral responses were categorized into six types: flying, walking, jumping, wing-flapping, alerting, and foraging. The response time refers to the interval between sound start to a specific behavior. Movement is defined when ① habitat type changes; ② the movement distance without intermediate behaviors exceeds 30 m; ③ flying behavior is involved. The new location is defined as the position where black-necked cranes stop moving and start another behavior, such as foraging. The escape distance is defined as the straight-line distance from the original location to the new location. If the black-necked cranes disappear from the observer’s field of view due to reasons such as crossing over a ridge, the escape distance is defined as the distance between the last visible position and the original position.
Statistical analysis
This study conducted experimental with 143 trials from March 10–22, 2024. A total of 5150 individual records were obtained during the experiment, including 4197 adults and 953 juveniles.
In this study, behavioral response, escape distance, and escape speed were used as metrics to evaluate the effectiveness of the audio stimuli. Based on the intensity of the response, behaviors such as flying, walking, jumping, wing-flapping, alerting, and foraging were assigned scores of 10, 7, 5, 5, 3, and 0 respectively, with the highest observed score taken as the measure. In order to analyze the response of the black-necked crane to different sounds, the Kruskal-Wallis test was used to analyze whether there were significant differences between the behavior assignment, reaction time and escape distance of the black-necked crane, since the data did not satisfy the normal distribution hypothesis. To explore differences in responses between adults and juveniles, as well as between family groups and flocks to different sounds, the Wilcoxon rank-sum test was employed to test for significant differences in behavior scores, escape distances, and escape speed due to data distribution. Additionally, multiple comparisons were conducted using the Least Significant Difference (LSD) method. The escape speed was calculated as the ratio of distance to time. All statistical analyses were performed using R software, version 4.4.337.
The datasets generated and/or analysed during the current study are not publicly available due ongoing utilization of the data for follow-up studies but are available from the corresponding author on reasonable request.
Results
Impact of different sound types on the behavior, escape distance, and escape speed of black-necked cranes
Different audio sounds elicit varying behavioral responses in black-necked cranes (Fig. 3a). Among the 12 sound types, 12,800 Hz - Constant (A3), 200–12,800 Hz - Alternating (B3), 200–12,800 Hz - Rapid Sweep (C3), and 200–12,800 Hz - Stepped Frequency (D3), which included the highest frequencies, elicited the strongest behavioral responses, with 6400–12,800 Hz - Stepped Frequency (D2) ranking next. The behavioral responses to the seven sound types 200 Hz - Constant (A1), 6400 Hz - Constant (A2), 200–6400 Hz - Alternating (B1), 6400–12,800 Hz - Alternating (B2), 200–6400 Hz - Rapid Sweep (C1), 6400–12,800 Hz - Rapid Sweep (C2), and 200–6400 Hz - Stepped Frequency (D1) were not pronounced and remained at a lower level. In the analysis of significant differences, the results indicated that 200 Hz - Constant (A1) significantly differed from 6400 to 12,800 Hz - Alternating (B2) and 200–6400 Hz - Rapid Sweep (C1), and there was also a significant difference between 200 and 12,800 Hz - Alternating (B3) and 200–6400 Hz - Rapid Sweep (C1). No significant differences were observed among the other groups.
Different audio sounds result in varying escape distances in black-necked cranes (Fig. 3b). 12,800 Hz - Constant (A3) and 200–12,800 Hz - Rapid Sweep (C3) exhibited superior performance in terms of escape distance, showing significant differences compared to 200 Hz - Constant (A1), 200–6400 Hz - Alternating(B1), 200–6400 Hz - Rapid Sweep (C1), 200–6400 Hz - Stepped Frequency (D1), 6400 Hz - Constant (A2), 6400–12,800 Hz - Alternating (B2), 6400–12,800 Hz - Rapid Sweep (C2), 6400–12,800 Hz - Stepped Frequency (D2), and 200–12,800 Hz - Alternating (B3). However, overall, the C sound type performed the best.
Different audio sounds lead to varying escape speeds in black-necked cranes (Fig. 3c). The two sound types 12,800 Hz - Constant (A3) and 200–12,800 Hz - Stepped Frequency (D3) showed significant differences in escape speed compared to the other 10 sound types. Among these, 12,800 Hz - Constant (A3) and 200–12,800 Hz - Stepped Frequency (D3) demonstrated the fastest escape speeds, followed by 200–12,800 Hz - Rapid Sweep (C3), 6400–12,800 Hz - Rapid Sweep (C2), and 200–12,800 Hz - Alternating (B3).

Effect of different sound types on behavior assignment, escape distance, and escape speed.
The impact of different sound types on juveniles and adult
No significant differences responses between juvenile and adults were observed (Fig. 4b)d)g)h)i)j)k)). In juveniles and adults, there were significant differences in behavior and escape distance in response to 6400 Hz - Constant (A2) and 12800 Hz - Constant (A3) sound (Fig. 4ae). In type B sounds, both juveniles and adults showed significant differences only in escape distance (Fig. 4f) between 200 and 6400 Hz - Alternating (B1) and 200–12800 Hz - Alternating (B3). For type C sounds, both juveniles and 200–6400 Hz - Rapid Sweep (C1) adults exhibited significant differences only in behavior scores (Fig. 4c) between 200 and 6400 Hz - Rapid Sweep (C1) and 200–12800 Hz - Rapid Sweep (C3). In type D sounds, both juveniles and adults demonstrated significant differences in both escape distance (Fig. 4h) and escape speed (Fig. 4l) between 200 and 6400 Hz - Stepped Frequency (D1) and 200–12800 Hz - Stepped Frequency (D3).

Effects of different sound types on juveniles and adults (Red and blue represent juvenile and adult groups, respectively).
The impact of different sound types on family groups and flocks
Family groups and flocks exhibited no significant differences in behavior assignment, escape distance or escape speed within each sound type(Fig. 5bgk)). Under type A sounds, both family groups and flocks demonstrated significant differences in behavior (Fig. 5a), escape distance (Fig. 5e), and escape speed (Fig. 5i) between 12800 Hz - Constant (A3) and both 200 Hz - Constant (A1) and 6400 Hz - Constant (A2). In type B sounds, significant differences were observed in escape distance (Fig. 5f) and escape speed (Fig. 5j) between 200 and 12800 Hz - Alternating (B3) and both 200–6400 Hz - Alternating(B1) and 6400–12800 Hz - Alternating (B2). For type C sounds, significant differences were only observed in behavior scores (Fig. 5c) between 200 and 12800 Hz - Rapid Sweep (C3) and both 200–6400 Hz - Rapid Sweep (C1) and 6400–12800 Hz - Rapid Sweep (C2). Under type D sounds, neither family groups nor flocks showed significant differences in behavior (Fig. 5d), escape distance (Fig. 5h), or escape speed (Fig. 5l).

Effect of different sound types on family groups and flocks (Green and blue represent family groups and flocks, respectively).
Discussion
The pitch and range of sound frequencies significantly impact on the response of black-necked cranes
This study demonstrates that high-frequency sounds are markedly more effective than low- and mid-frequency sounds in repelling black-necked cranes under single-frequency stimuli, which is consistent with the findings of many previous studies as well as with the design principles of bird deterrent devices23. Sound combinations encompassing a high frequency range manifest a substantially superior deterrence effect compared to those within low or medium frequency ranges. However, the temporal characteristics and variations in these sound combinations exert minimal influence on the birds’ behavioral response intensity, escape distance, or escape velocity. The enhanced deterrence effect observed with high-frequency sounds may arise from avian sensitivity to such frequencies, potentially rendering these auditory stimuli more effective in eliciting alert and evasive behaviors in birds38. While the temporal aspects and variations in sound combinations did not notably impact the escape behaviors of black-necked cranes in this study, sound combinations with a higher frequency range yielded significantly improved bird deterrence outcomes.
Black-necked cranes of different flock types and ages respond similarly to sound
This study revealed that both family groups and flocks, as well as juveniles and adults, displayed consistent responses to sound stimuli. We hypothesize that, as a typical flocking species, the behavioral and physiological reactions of black-necked cranes are shaped by their natural proclivity for consistency. Such flocking behavior not only facilitates rapid responses to predators but also enhances cooperation during activities like foraging and migration. Furthermore, this research was conducted towards the end of the wintering period, during which juveniles had progressively acclimated to the winter environment. Under adult guidance, their behavioral patterns matured, increasingly mirroring those of the adults.
Application recommendation
This study indicated that artificially designed sounds can be highly effective for the short-term displacement of black-necked cranes and potentially other bird species. The findings underscore the importance of considering not only the frequency but also the variations in sound frequency when designing bird-deterring sounds. It is noteworthy that, in the presence of food resources, displaced black-necked cranes may return to the site in less than 20 min following the cessation of sound playback. So in addition to individual sound avoidance techniques, we also advocate interdisciplinary collaboration, such as combining sound design with visual or chemical deterrence to make the avoidance effect more obvious.
Noise pollution not only alters the distribution of wild birds but also directly leads to a decline in their reproductive rates and changes in community structure39. Therefore, we must pay close attention to the impacts of using sound deterrents on other wildlife. We recommend that sound devices should only be used in areas and during periods where strict deterrence is necessary, such as in farmlands that have just been sown and are at risk of direct crop predation by black-necked cranes, or in wind farms during major migratory seasons where there is a greater risk of collision with wind turbines.
Limitations and perspective
The results of this study may be influenced by various factors, including differences in the experimental environment, individual variations, and the performance of the sound equipment utilized. This study focused solely on the effects of different sound frequencies and combinations, without evaluating variables such as sound intensity and playback duration. Future research could explore varying sound intensities and durations, and further investigate ultrasonic frequencies and random noise.
With ongoing technological advancements and deeper research, we can develop a clearer understanding of how sound influences species behavior. This knowledge provides a scientific foundation for devising more effective conservation measures and designing species-specific bird deterrence strategies. Additionally, sound-related research can offer valuable tools and methodologies for fields such as ecology and ethology, further enriching our understanding of animal behavior and ecosystem dynamics.
This study demonstrates that regulating species distribution through sound holds significant potential for promoting human-wildlife coexistence. By improving our understanding of how birds perceive and respond to auditory cues, we can develop more humane and sustainable methods to mitigate conflicts between humans and wildlife. Such approaches will help foster greater harmony between human activities and wildlife habitats.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
Verkuijl, C., Sebo, J. & Green, J. Animal welfare matters for sustainable development: UNEA 5.2 is an opportunity for governments to recognize that. https://sdg.iisd.org/commentary/guest-articles/animal-welfare-matters-forsustainable-development-unea-5-2-is-an-opportunity-for-governments-torecognize-that (International Institute for Sustainable Development, 2022).
Redpath, S. M. et al. Understanding and managing conservation conflicts. Trends Ecol. Evol. 28(2), 100–109 (2013).
AC No: 150/5200–32B FAA Advisory Circular on Reporting Wildlife Aircraft Strikes. https://www.faa.gov/airports/resources/advisory_circulars/index.cfm/go/document.current/documentNumber/150_5200-32 (2013).
Niksson, L. et al. Large grazing birds and agriculture—predicting field use of common cranes and implications for crop damage prevention. Agric. Ecosyst. Environ. 219, 163–170 (2016).
Zhao, B. et al. Searching a site for a civil airport based on bird ecological conservation: An expert-based selection (Dalian, China). Glob. Ecol. Conserv. 20, e00729 (2019).
Li, W. D. et al. Avian diversity and bird strike risk evaluation in Ninglang Luguhu Airport, Yunnan. J. Southwest For. Univ. (Nat. Sci.) 40(2), 96–102 (2020).
Frings, H. Behavioral manipulation (visual, mechanical, and acoustical). Pest Control Biol. Phys. Sel. Chem. Methods. 387–454 (1967).
Bhusal, S. et al. Automated execution of a pest bird deterrence system using a programmable unmanned aerial vehicle (UAV). Comput. Electron. Agric. 198, 106972 (2022).
Liu, H. J. et al. Research and practice of bird repellent equipment based on ultrasonic technology. Mod. Agric. Res. 002, 030 (2024).
Sun, P., Wang, G. Y. & Zhu, W. Y. Progress of research on integrated control technology for airport flying birds. Sci. Technol. Innov. Herald 9, 2 (2008).
Defusco, R. P. Frightening Devices for Airfifield Bird Control. (Colorado State University, 1983).
Nomsen, D. E. Preventing waterfowl crop damage. (1991).
Lagler, K. F. The control of fish predators at hatcheries and rearing stations. J. Wildl. Manag. 3(3), 169–179 (1939).
Andelt, W. F. et al. Effectiveness of barriers, pyrotechnics. Wildl. Soc. Bull. 25(3), 686–694 (1997).
Feare, C. J., Dunnet, G. M. & Patterson, I. J. Ecological studies of the rook (Corvus frugilegus L.) in North-East Scotland: Food intake and feeding behaviour. J. Appl. Ecol. 867–896 (1974).
Nelson, P. Serious pests need serious treatment. Orchardist N. Z. 63(10), 25–27 (1990).
Thompson, R. D. et al. Differential heart rate response of starlings to sound stimuli of biological origin. J. Wildl. Manag. 888–893 (1968).
Spanier, E. The use of distress calls to repel night herons (Nycticorax nycticorax) from fish ponds. J. Appl. Ecol. 287–294 (1980).
Ming, L. & Hu, W. Contrast of several bird scaring modes. Agric. Eng. S1, 2 (2014).
Cook, A. et al. An evaluation of techniques to control problem bird species on landfill sites. Environ. Manag. 41, 834–843 (2008).
Tao, J. R. Design and Application of the Intellect Wireless Driving-Bird System Based on A Voice and Ultrasound Wave (Dalian Maritime University, 2011).
Conniff, R. Why catfish farmers want to throttle the crow of the sea. Smithsonian 22(4), 44–53 (1991).
Ogochukwu, E. S., Okechukwu, A. D. & Nnaegbo, O. G. Construction and testing of ultrasonic bird repeller. J. Nat. Sci. Res. 2, 8–17 (2012).
Corbani, T. L., Martin, J. E. & Healy, S. D. The impact of acute loud noise on the behavior of laboratory birds. Front. Vet. Sci. 7, 607632 (2021).
Swaddle, J. P. et al. A sonic net excludes birds from an airfield: implications for reducing bird strike and crop losses. Ecol. Appl. 339–345 (2016).
Jin, L. Y. & Li, D. L. Comparative evaluation of efficient bird repeller distance between gas gun and sound bird repeller. Chin. J. Appl. Ecol. 32(1), 7 (2021).
Wang, K. et al. Relations of daily activity patterns to age and flock of wintering black-necked crane (Grus nigricollis) at Napa Lake, Shangri-La in Yunnan. Zool. Res. 030(001), 74–82 (2009).
Che, Y. & Li, Z. Q. Vigilance behavior of animals: overview and perspective. Sichuan J. Zool. 1(7) (2014).
BirdLife International. Grus nigricollis. The IUCN Red List of Threatened Species 2020: e.T22692162A180030167. https://www.iucnredlist.org/species/22692162/180030167 (Accessed 7 Oct 2024).
Chen, J. et al. Global distribution and number of overwintering black-necked crane (Grus nigricollis). Biodivers. Sci. 31(6), 22400 (2023).
Ci, R. et al. Relationship between wintering habitat protection of the blacknecked crane and local agricultural activities. J. Tibet Univ. 5, 7 (2009).
Jia, R. et al. Population dynamics and habitat use of the black-necked crane (Grus nigricollis) in the Yarlung Tsangpo River basin, Tibet, China. Avian Res. 10, 1–8 (2019).
Dong, H. Y. et al. Winter diet and food selection of the Black-necked Crane Grus nigricollis in Dashanbao, Yunnan, China. PeerJ 4, e1968 (2016).
Liu, W. et al. Home range and habitat use of breeding black-necked cranes. Animals 10(11), 1975 (2020).
Zhong, L. et al. Local farmers’ perceptions of ecosystem services and disservices provided by the Black-necked Crane (Grus nigricollis) and their conservation implications. Glob. Ecol. Conserv. 46, e02614 (2023).
Pearse, A. T. et al. Migrating Whooping Cranes avoid wind‐energy infrastructure when selecting stopover habitat. Ecol. Appl. e02324 (2021).
Li, Z. H. M. & Li, F. S. Black-Necked Crane Research (Shanghai Science and Technology Education Press, 2005).
R Core Team. R: A Language and Environment for Statistical Computing. https://www.R-project.org/ (R Foundation for Statistical Computing, 2024).
Francis, C. D., Ortega, C. P. & Cruz, A. Noise pollution changes avian communities and species interactions. Curr. Biol. 1415–1419 (2009).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (32360274). The research was conducted with the substantial support and assistance from Datang Yongshan Wind Power Co., Ltd., specifically through the Laoluliangzi Wind Farm Winter Black-necked Crane Monitoring Service Project. We also thank the staff members and students who participated in the fieldwork.
Author information
Authors and Affiliations
Contributions
Conceptualization, Wen Xiao; Methodology, Wen Xiao; Resources, Na Li; Software, Chang-Jin Liu; Visualization, Hong-Bin Ma; Writing – original draft, Zi-Juan Dong; Writing – review & editing, Kun Tan.All authors will be informed about each step of manuscript processing including submission, revision, revision reminder, etc. via emails from our system or assigned Assistant Editor.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Dong, ZJ., Tan, K., Ma, HB. et al. High-frequency and high-amplitude sounds enhance bird deterrence. Sci Rep 15, 30945 (2025). https://doi.org/10.1038/s41598-025-13737-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-13737-2
