
Abstract
Nature-based solutions are now a key part in climate change adaptation, particularly for urban environments. Urban forests are one of the most used methods for adding ecosystem services to an urban environment and at the same time address urban-specific climate change challenges such as heat-island effect, intense rainfall and water management. However, the effects of climate change in the long-term on urban forests are seldom taken into account when planning interventions such as afforestation. Species selection for urban forests should, among other factors, be based on an assessment of local present and future climatic conditions, so to ensure the long-term viability of the adaptation action. Here we propose a methodology to aid the species selection process, which is readily applicable to any place in Europe. We use data from publicly available gridded datasets of climate (ClimateEU) and tree species distribution (European Tree Atlas) in order to estimate realised trees’ climatic niches. These are then compared to local climatic data, in order to obtain vulnerability scores that ranks the species according to their vulnerability to projected future climate conditions. As an example, we apply the method to a pilot city in central Italy, L’Aquila, where planning of urban afforestation is undergoing. Our analysis suggests that some species considered locally native will be less vulnerable than others (e.g. Quercus pubescens, Castanea sativa), while some species not locally native will result even less vulnerable (e.g. Quercus ilex, Pinus nigra). This information may be used as an initial input to the tree species selection decision process.
Similar content being viewed by others


Introduction
Urban environments across the globe are significant contributors to anthropogenic climate change and some of the most affected by it. Cities are particularly vulnerable to increases in intensity and frequency of storms, heatwaves, coastal flooding, and heavy rainfall. Such extreme weather events pose great risk to urban environments as multiple climate and non-climate hazards can interact and affect human health, livelihoods and infrastructure at large scale1,2. It is therefore vital to consider urban environments as not only contributors to climate change but also as primary actors in mitigation and adaptation actions.
Many climate adaptations for infrastructure have been proposed and enacted, but growing consensus favours a combination of nature-based solutions (NBS) which attempt to create resilience through the creation and incorporation of natural ecosystems within the urban fabric3. This approach provides, in principle, resilience and the provision of multiple ecosystem services at once4,5. The popularity and effectiveness of NBS as an instrument to increment climate change resilience of urban areas has been vastly explored and documented6. However the implementation of such solutions is not always straightforward and needs to be thoroughly assessed5,7. The need for continuous research in support of NBS is paramount in order to assess their sustainability and long term viability7,8,9. Problems may arise when such solutions are not carefully planned in consideration of future climatic models that may affect their viability, sustainability and effectiveness in the long term10,11.
The use of urban forest for climate change mitigation and adaptation in urban environments, shows promising results. Urban forests have the potential to mitigate the effects of intense rainfall, heat-island effects and overall provide an efficient tool for carbon storage and sequestration12 and water collection, storage and filtration13,14. In addition, they provide a series of ecosystem services to the public and have the potential to preserve local biodiversity15,16.
It is important to clearly define the objectives of urban forestry initiatives for climate adaptation and to further assess whether the goal will actually be met. Various elements might significantly reduce the effectiveness of urban forests for obtaining desired outcomes. Latitude, for example, has been shown to have a significant effect on urban forests’ potential to reduce land surface temperature, with southern European cities showing lower benefits compared to northern European ones14. Climate change in itself could pose significant strain on the survival of forests and consequently on the effectiveness of forest centred NBSs. The vulnerability of forests to climate change is often overlooked by practitioners17. Tree vulnerability to extreme weather events, habitat degradation, invasive species, disease and changes in pest life-cycle are all exacerbated by climate change10,18,19. It is important for practitioners to thoroughly assess the future conditions these forests will need to survive in, ensuring that the benefits they provide will be maintained when most needed.
An assessment of future environmental conditions should inform the selected species composition of the urban forest, i.e. whether it is possible for such a forest to not only survive but thrive. Attempts have been made to provide frameworks to produce climate change impact assessments for urban forests17,20,21,22. This has resulted primarily in an assessment of species and habitat dynamics in relation to climate change exposure risks such as temperature, precipitation and wind22. Social aspects also shouldn’t be overlooked, as access and education, economic value and political engagement could directly affect the long term management and ultimate success of the forest22.
The study of species realised climatic niches has been carried out by several authors to assess the vulnerability of urban forests to climate change. This has been done primarily through the initial quantification of such niches through the use of species occurrence data and gridded climate datasets. However, how this data was then used, depends primarily on the objective of the study. Most of these studies tend to provide large scale insight into vulnerability of urban forests. Whether through the study of urban forest composition23exposure, safety margins and risk24or comparison of urban vs. native climate niches25. However, they do not try to provide a ranked species list of vulnerability in relation to climate change, something that can be used for an initial assessment of species selection by local authorities. Khan and Conway17 provide a more replicable species selection methodology which ranks species based on their vulnerability to climate change, and it is therefore what has been adapted by us for this investigation.
In this work, we combine tree species distribution from the European Tree Atlas project26 with climate information of the past and the future, with the aim of ranking those species in terms of their vulnerability to climate change. Here we define an urban forest, as a single plantation spatially localized in a well delimited area inside the city’s boundaries, and not as the sum total of all public and private trees across the city, consequently urban afforestation is the act of planting such a forest. We test our methodology in the municipality area of L’Aquila, a middle-sized city in Central Italy, which is including urban afforestation, as one possible action in its climate adaptation strategy. The method developed may be readily applied to other cities and places in Europe.
Data and methods
European tree atlas
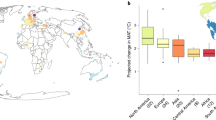
The European Tree Atlas collates and analyses forest tree species occurrence data from various sources across European countries. The results are used to produce a series of species distribution maps. The model-derived maps produced are for Relative-probability of presence (Fig. 1A) and Maximum Habitat Suitability (de Rigo et al., 2016). The latter highlights areas across Europe where tree species could potentially survive, based on many climatic variables. This would theoretically allow us to verify for a given location which species are more likely to survive. However, the lack of some key species for which this data is unavailable and the lack of a temporal dimension that doesn’t allow an assessment of suitability in future climatic conditions, limits its use for species selection based on climate change vulnerability, hence the motivation for the analysis proposed here.
The reason for using such probability of presence data compared to occurrence data, falls on the fact that the latter offers an often biased distribution that requires modelling for it to be a closer representation of the species true distribution, as presence or absence cannot be assumed where occurrence is not recorded27,28. One of the aims of the European Tree Atlas was to eliminate as much as possible the limitations due to the sparse and incomplete nature of multi-source occurrence data. Furthermore, being probability of presence data, it provides information of not only presences, but also absences, useful for quantifying a species climatic niche.
European climate
The ClimateEU project provides gridded climate data for Europe through aggregation and enhancement of data from several publicly and not publicly available databases. It provides monthly, annual, decadal, and 30-year normal climate data for the years 1901 to 2019 as well as multi-model (CIMP5) climate change projections, which are not used here. Gridded bioclimatic variable data is available at a resolution of 1 km2 allowing its use for spatial data analysis (Fig. 1B)29.
In order to check the robustness of our results, we also use an alternative climate gridded dataset, WorldClim version 2.1. Thissimilarly offers gridded bioclimatic variables for the global land surface, averaged over the 1970–2000 period, at an approximate resolution of 1 km2. This gridded product was generated through the interpolation of weather station data from a dataset comprising up to 60,000 weather stations, in conjunction with satellite data30. Climate EU provides a higher quality dataset, as it uses data from WorldClim combining it with data from other packages and processing it to make it more relevant to Europe.
The choice of using publicly available datasets is aimed at enhancing the reproducibility of our work, and it is an improvement with respect to our main referenced previous work17which used a commercial software to produce the gridded climate dataset (ANUCLIM).

Pilot area
L’Aquila is a medium-size city with a population of approximately 70,000, located in the region of Abruzzo in Central Italy. It is located in the Middle Aterno Valley, which extends from north-west to south-east between the Gran Sasso Massif to the North, the largest and highest mountain massif of the Apennines mountain range, and the Velino-Sirente Massif to the South. The altitude of the main urbanized are is between 650 and 720 m a.s.l. One proposed pilot forest site is located in the western part of the city (Fig. 2), characterized mostly by the presence of industrial and commercial warehouses, residential neighbourhoods and sparse unmanaged patches of vegetation.
The composition of the native flora of the surrounding area, because of its mountainous terrain, is primarily defined by altitudinal gradients. Natural and semi-natural areas tend to be well preserved with most of them falling inside protected national parks. Agricultural habitats (pastures and crops) are concentrated in the flats closest to the city’s urban area, while woodland becomes the primary habitat type as the elevation gradient changes. Two main types of woodlands are mostly observed: native Quercus dominated natural and semi-natural deciduous forests and locally non-native Pinus nigra coniferous plantations32. These two main forest types are often merged together, particularly at higher elevations. The city’s climate is classified as continental temperate without dry season and warm summer or Cfb according to the Köppen-Geiger climate classification33. The city’s Mean Annual Temperature is 12.5 °C (1991–2020) and has ranged from 10.7 °C to 13.4 °C in the years 1974 to 2022. The Mean Annual Precipitation is 650 mm/year (1991–2020) and has ranged 440–951 mm/year in 1974–2022. The highest temperatures occur in August with a mean maximum temperature of 31 °C. Lowest temperatures tend to occur mostly in January with the mean minimum temperature being − 2.0 °C.

Satellite image of the city of L’Aquila, the location of the proposed urban forest and the city’s geographical collocation. Map produced with QGIS31.
Local climate observations
A long-term time series of daily temperature and precipitation from 1974 to 2022 is taken from previous work33. The dataset was carefully checked for data quality and has been subject to a homogenization procedure, in order to correct the unavoidable discontinuities (e.g. due to instrumental updates or site relocation) affecting multi-decadal weather station observations. The corrected time series show an increasing trend of temperature in the recent decades, in the order of about + 6 °C/century for daily maximum temperature, + 4 °C/century for daily minimum temperature, while for precipitation the trend is less clear.
We use this local climate dataset also to bias correct future climate projections for the scenarios SSP 126 and SSP 585. The method used for bias correction is the ISIMIP3BASD v1.034, which is a quantile-based adjustment method for climate projections developed for the Inter-Sectoral Model Intercomparison Project (ISIMIP) that is aimed at preserving trends in the original projection across quantiles, in order to have a robust correction for both central and extreme values. The correction for precipitation is multiplicative, while that for temperature is additive. In a recent intercomparison of bias adjustment methods, ISIMIP3BASD was found to preserve better the raw signals of the original variables together with other two methods35. We selected ISIMIP3BASD for our work, because it is more recent in the development and because we found its installation and usage more straightforward than the other two algorithms.
In the present work, we need information on maximum and minimum daily temperature values in addition to the mean, thus we need this kind of information in the future projections. We explored the latest climate projection dataset available, namely the Coupled Model Intercomparison Project Phase 636, using the Earth System Grid Federation facility37: we found only one model providing maximum and minimum temperature (tasmax and tasmin) at daily resolution for both historical and future periods, namely EC-Earth v3 model38. We thus selected this model and downloaded the variables tasmax, tasmin and pr (precipitation) for the period 1970–2100 and for the available SSP scenarios.
We applied a point-wise bias correction in order to avoid complications of generating a gridded dataset39which is not explicitly needed and left to future development of this work. We interpolated the raw EC-Earth gridded timeseries over the point of interest, i.e. the same location of the station for which we have a climatological series in the area33and applied only the bias adjustment step of the procedure: the statistical downscaling is needed only when producing a gridded dataset, useful for increasing the spatial resolution of the grid. The daily timeseries obtained from the procedure were thus employed to calculate the values of the local thresholds used to build the ranking of tree species vulnerability.
Tree climate vulnerability score
Tree species distribution data was obtained from the European Tree Atlas26. The most common and dominant tree species native to Italy were chosen from those available (24 species). Italy’s complex geomorphology and extent allowing for a diverse range of habitat types and bioclimatic zones, makes such a list of species a good starting point as it encompasses trees adapted for a multitude of different climatic conditions. No exotic species have been used as one of the main scopes of urban afforestation should be to also increase the nature conservation value of a city by using native species40,41,42. However, we do include species which are not locally native, i.e. occurring naturally, without human intervention, in the natural habitats characteristic of the ecological identity of the area where the city is located.
The data provided in form of relative probability of presence raster files, was combined with spatial climate data obtained from the ClimateEU project29 (Fig. 1). A series of bioclimatic indicators were obtained. These are Mean Annual Temperature (MAT), Maximum Summer Temperature (Tmax_sm), Minimum Winter Temperature (Tmin_wt) and Summer Precipitation (PPT_sm). The choice of these indicators was made based on the projected changes in climate expected for the study area, which has experienced throughout the past decades an increase in minimum and maximum temperatures and a decrease in precipitation particularly during the summer season33. We chose to include variables of both temperature and precipitation which show extremes but also general conditions, in order to provide a more complete representation of species climatic niches, compared to other studies using less variables only showing one or the other17,23. All the variables used are all known to influence tree species distribution43,44.
Data were processed and analysed using R Statistics software45. Species realised climatic niches were obtained from layering raster files from chosen ClimateEU bioclimatic variables with Relative Probability of Presence raster files for each tree species. In this way, tree probability of presence was associated with each bioclimatic variable in each grid cell. Probability of presence equal and greater to 5% was used as cut-off and was considered presence. This value was chosen based on the metrics used by the European Tree Atlas to define species occurrence, with values of 5% defined as low presence. As we are interested in the species realised niche, even low presence of a species in a given area, will result in valuable information for assessing the niche thresholds for each species.
Climate niche range for each species for a given bioclimatic variable were defined by the 95% and 5% percentile of the species full climatic niche obtained. These extreme limits were used as an indication for niche tolerance. From here on, the niche calculated between these two extreme limits will be named “core range”.
In order to assess the species vulnerability to climate change a scoring system was used, building on the method developed by Khan and Conway17as summarised in Table 1 and as explained in what follows. For each climatic variable, a score was given to each species based on whether the local value (in our illustrative case, the city of L’Aquila) of historical, SSP 126 and SSP 585 future projection of the bioclimatic variable fell within the species core range or not. When the local value of a bioclimatic variable does not fall into the species core range, a point of 1 was given. Since we are considering the local values of three periods (historical, SSP 126 and SSP 585), each species could score a minimum of 0 and a maximum of 3 for each bioclimatic variable. The higher the score summing on the variables, i.e. the less present and future conditions fell into the core range, the more vulnerable the species is considered. The historical climatic conditions were taken into consideration as baseline conditions, as species will have to be planted now, therefore it is important to not only guarantee future success but also short-term success, as tree species are more vulnerable during their early stages of growth46.
In order to differentiate species with smaller core ranges which are better adapted to specific narrower conditions, in addition to the scores associated to the four bioclimatic variables, an adaptability index was also created. We define the “core spread” of a species for a given climatic variable as the breadth of its core range (max – min). For each climatic variable, each species core spread was calculated. Considering the relative magnitude of the core spread across all the species, a score ranging 0–3 was given. Higher values indicate narrower spread hence lower adaptability. If the core spread of a species fell into the 1 st, 2nd, 3rd, or 4th quartile a score of 3, 2, 1, or 0 was assigned. This was done for all climatic variables. To provide the final adaptability score, the scores for all climatic variables were added giving a total value for each species. The ensemble of these totals was then divided again in 4 quartiles, and according to which quantile the species total score fell into, the final adaptability score (3, 2, 1 or 0) was assigned. This makes it possible to assess a species based not only on its climatic range but also on its potential to live in a wider range of conditions.
In order to check whether results are independent of the climatic dataset used, core ranges and a vulnerability matrix were generated again by using rasters from theWorldClim v2.1 dataset30with the same horizontal resolution of 1 km2. The bioclimatic variables used by WorldClim are slightly different from those used by ClimateEU. Max Temperature of Warmest Month (BIO 5) and Minimum Temperature of Coldest Month (BIO 6) were used as substitutes for Summer Mean Max Temperature and Winter Mean Min Temperature respectively. While Precipitation of Warmest Quarter (BIO 18) was used in contrast with Summer Precipitation. The chosen rasters for these bioclimatic variables were projected onto the ClimateEU rasters in order to produce equivalent files and match geographic extent. To generate core climatic ranges and a vulnerability matrix, the same method described above was used.
The study assumes that the current distribution of trees across Europe is entirely dependent on historical climatic conditions. While this is one of the main factors defining distribution, it is important to keep in mind that other factors such as historical events, human interference and biotic factors such as competition and community dynamics, may affect geographical restrictions47.
Results and discussion
European tree climatic ranges
In Fig. 3 we can observe differences in core ranges for the 24 tree species analysed. Through these it is possible to differentiate species adapted to higher temperatures and drier conditions from those more adapted to colder and wetter environments. For example, Quercus suber as a Mediterrenean evergreen oak, is the most warm adapted and drought tolerant species in the list. With our results setting its median Mean Annual Temperature at approximately 15 °C, as well in line with current literature48. On the other hand of the spectrum we have Pinus sylvestris showing clear preference for lower temperatures with a median Mean Annual Temperature of approximately 5 °C with lower limits < 0 °C. These results support current knowledge of the species optimal temperature ranges which dictate fundamental physiological processes49. In addition, the large breadth of its temperature-related climatic range support the wide geographical extent of this species and its ability to live in diverse climates50. In general, the core ranges calculated with our method appear to be well in line with the pre-existing knowledge about the species considered.


Species core ranges for each of the four bioclimatic variables used. Bar height indicates 95 and 5 percentile of the entire species range, and middle bar the median. Horizontal lines supply the historical value for L’Aquila, calculated on the period 1973–2003, and the projected SSP 126 and 8.5 values.
Tree climate change vulnerability ranking in the pilot area (L’Aquila)
The majority of the species explored show high vulnerability to projected climatic conditions in the test area (Fig. 3), in particular to Summer Mean Maximum Temperature (Tmax_sm) and Mean Annual Temperature (MAT). For MAT, no species core range overlap the predicted SSP 585 local value, and only seven overlap the SSP 126 one. These species are Quercus suber, Quercus ilex, Quercus pubescens, Quercus cerris, Pinus nigra and Fraxinus ornus. Eleven species overlap the historic value for MAT. These include the ones just mentioned except Q. suber.
With regards to Tmax_sm, no species core range overlaps with the SSP 585 and only two overlap the SSP 126 local value: Q. suber and Q. ilex. P. nigra is the only one, together with the two just mentioned, to overlap the historical value.
With regards to Minimum Mean Winter Temperature (Tmin_wt) eight species overlap the SSP 585 scenario, with twenty-one overlapping the SSP 126 value. The same species mentioned above are the ones that show the least vulnerability to predicted Tmin_wt, together with Castanea sativa and Quercus robur. Q. suber shows the lowest levels of vulnerability to the highest predicted Tmin_wt, but does not show overlap for the SSP 126 value nor the historic value, further indicating vulnerability to present local conditions. Q. ilex being a less specialized species, on the other hand, shows both the lowest levels of vulnerability, but also an overlap with historic values, together with the species above mentioned whose ranges are shifted lower into colder temperature, in comparison, but still manage to encompass the SSP 585 worst case scenario value.
With regards to Summer Precipitation (PPT_sm), seven species overlap the SSP 585 value, C. sativa, F. ornus, P. nigra, Q. cerris, Q. ilex, Q. pubescens and Q. suber. Popolus nigra is the only one in addition to these that reaches the SSP 126 scenario. Only eleven of the species encompass the historic value.
In Table 2, we show the calculated vulnerability scores associated with each bioclimatic variable, that reflect the observations illustrated above. Moreover, we also report the calculated adaptability score, which is based on the width of the core ranges and it is useful to identify species which seem to perform well, but that may not be good candidates because associated with a relatively narrow climatic niche. Q. suber shows in fact low vulnerability to predicted climatic conditions, but very low adaptability, being a highly specialist species. P. sylvestris shows high adaptability, but mainly to cold environments and therefore shows also high vulnerability to climate change.
Species with high adaptability with also an overall low vulnerability score are P. nigra, Q. ilex, Q. pubescens, C. sativa and F. ornus. They are the ones the rank first in the area of interest of L’Aquila and are thus the first that should be considered for the urban afforestation action. These species are widely distributed across the local natural landscape. The fact that these species are also the less vulnerable shows that the method used has real life applications and that it matches what is locally observed, serving as first line of evidence in support of our analysis. Q. pubescens, C. sativa and F. ornus occur naturally and are considered locally native, composing most of the natural and seminatural woodland in the area outside the city. Pinus nigra is widespread both in the city and in the surrounding mountains. Its occurrence is primarily due to afforestation and natural expansion from the first plantations. The species is not considered to be locally native. Nevertheless, it thrives in the current ecological and climatic conditions. Q. ilex, on the other hand, is more widespread at lower altitudes and in warmer areas of central Italy. As an evergreen Mediterranean oak, it is highly adapted to dry and hot conditions year-round. Our analysis shows that it can also sustain moderately cold climates, in fact the species is presently planted ornamentally in the city even if it does not occur naturally in the surrounding natural landscape.
Compared to the method developed by Kahn and Conway17our work uses input gridded data at a finer resolution of 1 km2 against 100 km2allowing for greater specificity of topographical variation in species distribution. Furthermore, it uses freely available climate and species distribution data. It takes advantage of the large work conducted by the European Tree Atlas project, which was able to produce species distribution data from the mostly sparse and inconsistent datasets available. It relies only on climate data and does not require qualitative measures of species characteristics such as those used by Khan and Conway17as these are often not available. Most importantly, the method differs in the use of observational data to provide local climatological values on which the reliability of these results is highly dependent. Accuracy increases when homogenised climatological data based on local observations is used instead of simply extracting values from publicly available climate packages (e.g. ClimateEU & WorldClim). These work well on large scale and in fact, as we have demonstrated, they allow us to successfully define species climatic ranges, but they tend to show significant bias when single points are extracted. We therefore recommend those wanting to apply this method to other cities, to work on obtaining reliable climatological data from local observations, as results will be greatly affected by this input.
The comparison with the results obtained when using a different climate database, serve as further evidence for reliability of the method. Species’ scores for each bioclimatic variable and their overall rank, obtained from running the analysis on two different databases (ClimateEU and WorldClim) (Fig. 1S), are nearly identical (Table S1). The proposed approach therefore allows for an applied use of climate change vulnerability assessment for application in urban forestry. Of course, species selection should not be based entirely on climate change vulnerability, but rather on a complete evaluation of ecological and sociological factors. We further elaborate on these aspects in the next section.
Validation against independent information
Previous work used species distribution modelling in combination with climate projections to predict the tree species composition changes expected under ongoing climate change51,52,53. Although a direct validation is difficult for these models, we may compare qualitatively the information that can be gathered from them with that that can be inferred from our simplified method.
In particular, the EU-Trees4F Dataset52 contains images of the tree species modelled until the end of the century (2100) and for the emission scenarios RCP 4.5 and 8.5. The images provide the expected change with respect to present, using an intuitive classification that the authors define as follows: “‘Stable presence’ denotes areas that will remain suitable habitats from the present until the end of the century. ‘Decolonized’, indicates areas that will become climatically unsuitable by the end of the century. ‘Suitable but not occupied’, represents areas that will become climatically suitable by the end of the century but will not be naturally colonized due to dispersal limitations.”. The images for the species relevant for our study are reported in the updated supplement of our manuscript in Figure S2.
In order to compare with the EU-Trees4F projections and also to check the consistency of our ranking methodology with the current climate of L’Aquila, we recalculated the vulnerability score, based on three climatic values (in the primary analysis: one for the present and two for the future climate scenarios) by only using the value for the present climate: that way, we rank tree species vulnerability only based on current climate. We recall that the species selected are among those available in the European Tree Atlas and naturally already present in the Mediterranean area.
In Table S2, we report the difference of the scores calculated with present climate only and with present and future climates. When a difference is positive (negative), it means that in the future the score of that species is getting “worse” (“better”), i.e. more (less) vulnerable to future conditions. The first consideration is that all species but two (Q. ilex and Q. suber) have an increase in the total sum of their scores, confirming that in the present all those species are already mostly adapted to the local climate, which is the reason why we selected them for the study. In this sense, the method already demonstrates a certain degree of sensitivity to input climate and results change in the direction expected.
Looking at some species in particular, we notice that Pinus nigra stays in the first place, although it will suffer more from increased summer temperature in the future. According to the EU-Trees4F classification the species is suitable and remains suitable in the future: it is indeed not a native species, but a tree that was planted in a wide afforestation program at the beginning of the 20th century in Italy.
Q. ilex and Q. suber are expected to suffer less from minimum winter temperature, which are also projected to increase: indeed, they both climb up the ranking with respect to the present. Consistently, in the EU-Trees4F projection Q. ilex is considered a stable species in the area, while Q. suber is an emerging one. Due to increased winter temperature, also other species gain positions, e.g. Acer pseudoplatanus, Corylus avellana, Pinus sylvestris. In these cases, the results of EU-Trees4F are not always consistent with ours, such as for Corylus avellana, which is for the most part a cultivated species.
Species such as F. sylvatica, Acer campestre and Populus nigra will become less suitable, because of the decreased water availability due to increased evaporation (MAT increases) and to decreased summer precipitation. A few species become less suitable because of the increased winter temperature, e.g. Sorbus aucuparia and Larix decidua, which are species that need a certain amount of chilling accumulation. The projections of EU-Trees4F are consistent with our results, indicating a decolonization in the more pessimistic RCP 8.5 scenario.
Overall, we may conclude that our method proves to be sensitive to the input climate and that the information from the resulting vulnerability ranking is consistent with other independent projections.
Further considerations on urban afforestation
The planting of locally non-native species poses a series of questions. For example, let us suppose that Q. ilex is selected to be planted for the proposed project. The future climatic conditions predicted for the area suggest that it would be a good candidate for future forests to thrive in future conditions. Its introduction though could pose a serious change to local biodiversity, as sclerophyllous Mediterranean woodlands are not to be expected in the area. The challenge with wanting to maintain ecological coherence during planning for future climatic change is not to be taken for granted. How accurate are our predictions for ecosystem change and how much should we accommodate for such predictions in spite of concerns for local biodiversity conservation?
Beyond climatic conditions, the success of an urban forest project relies on a series of other parameters that need to be considered related to the species compatibility with the specifics of the site. Expert horticultural knowledge would be necessary to further assess site specific parameters, such as gradient, sun exposure, soil composition, quality and permeability54,55. Integration with its use and anthropogenic disturbance predicted throughout the stages of growth should also be considered. Vulnerability of young trees is considerable; therefore, all sources of stress should be thoroughly taken into account. Management considerations are furthermore paramount especially in light of the climatic changes discussed in this study. Water availability throughout the driest seasons needs to be guaranteed and its long-term requirements thoroughly calculated based on predicted changes in rainfall and evapotranspiration.
As this project’s objectives are to provide natural based solutions to expected climatic changes in rainfall and temperature it is important that its desired benefits overcompensate the resources needed for maintaining the forest. Rainwater collection and storage during extreme weather events can be an important asset to counteract drought events. Furthermore, considerations for species choice can be made with the goal of implementing specific objectives. Such considerations could go beyond that of choosing species solely based on biodiversity conservation. For example, the choice between conifer and broadleaved dominant forest may have implications on water retention and loss. Conifers have higher evapotranspiration rates than broadleaf trees particularly during the dormant season56 in high winter precipitation regions57. The extra water that reaches the ground with broadleaf trees during rainier months could be captured and stored for later use during drier months. On the other hand, the presence of coniferous trees during extreme rain events could reduce the amount of overflow and of flooding by capturing and slowing down more water than would broadleaved trees especially during dormancy57,58. This argument could be extended to evergreen broadleaved trees as well57. Water use by trees may have implications on water availability overall59though considering the relatively small size of the proposed project, this aspect might be irrelevant for the pilot study, but important if large scale application is foreseen.
Species diversity is also an important aspect to consider. Species diversity has been shown to be an important factor when trying to maximise forest resilience in light of disturbances. In particular to those exacerbated by climate change60,61. This would imply better results if a variety of species would be chosen to be planted instead of just one62. This poses another element to be considered, should the community being planted be based on natural forest communities already native to the area, or should a selection of best trees be based solely on their predicted performance? Eventually it all comes down to the objectives behind the proposed project. If creating a natural native habitat with the potential to also benefit local biodiversity conservation, the former should be done. On the other hand, if the objective is to plant the most efficient and least vulnerable forest to climate change, necessary to mitigate its effects, then the latter would be more adequate. Of course, there could be a solution that would produce both desired outcomes. Even though this study focuses on single species and not communities, results could be extrapolated to comprehend the communities associated with the least vulnerable species and propose a diverse plantation that mimics such communities. This could potentially provide a solid basis for planting a forest that resembles closely local native woodland and therefore increase the nature value of the city.
Conclusions
Thanks to the use of quality publicly available datasets and homogenised climate data from local observations we were able to produce an easily reproducible methodology for the rapid assessment of tree species vulnerability to climate change, and illustrated its application for the city of L’Aquila, Italy. Climate vulnerability is not often considered when planting urban forests for climate change adaptation, and therefore a simple way to readily make this assessment across European cities, is needed. We provide in the supplement the R script and the basic input data to calculate the tree vulnerability ranking for other locations. Stakeholders might often overlook important aspects if information is not readily available, hence the necessity for such a method. The ability to obtain such information in a simple and practical manner can be of great benefit to stakeholders. As noted, the success of urban forests relies on a multitude of diverse biological, ecological and socio-economic factors which should all be taken into account, for which the proposed method represents one starting input.
Data availability
The R script and the basic input data to calculate the tree ranking is attached in the online supplement of this article. The Tree Atlas data are publicly available at the link (last access 29 Apr 2025): https://forest.jrc.ec.europa.eu/en/european-atlas/atlas-data-and-metadata/. ClimateEU dataset is publicly available at the link (last access 29 Apr 2025): https://sites.ualberta.ca/~ahamann/data/climateeu.html. The other data and materials are available upon request to the authors.
References
Gasper, R., Blohm, A. & Ruth, M. Social and economic impacts of climate change on the urban environment. Curr. Opin. Environ. Sustain. 3, 150–157 (2011).
IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability. (2022).
European Environment Agency. Nature-Based Solutions in Europe Policy, Knowledge and Practice for Climate Change Adaptation and Disaster Risk Reduction (Publications Office, 2021).
Epelde, L. et al. Quantification of the environmental effectiveness of nature-based solutions for increasing the resilience of cities under climate change. Urban Urban Green 67, (2022).
European Commission. Evaluating the Impact of Nature-Based Solutions: A Handbook for Practitioners (Publications Office of the European Union, 2021).
Oral, H. V. et al. A review of nature-based solutions for urban water management in European circular cities: a critical assessment based on case studies and literature. Blue-Green Syst. 2, 112–136 (2020).
Seddon, N. et al. Understanding the value and limits of nature-based solutions to climate change and other global challenges. Philos. Trans. R Soc. B Biol. Sci. 375, 20190120 (2020).
Frantzeskaki, N. & McPhearson, T. Mainstream Nature-Based solutions for urban climate resilience. BioScience 72, 113–115 (2022).
Turner, B. et al. The role of Nature-Based solutions in supporting Social-Ecological resilience for climate change adaptation. Annu. Rev. Environ. Resour. 47, 123–148 (2022).
Anderegg, W. R. L. et al. Climate-driven risks to the climate mitigation potential of forests. Science 368, 7005 (2020).
Gómez Martín, E., Máñez Costa, M., Egerer, S. & Schneider, U. A. Assessing the long-term effectiveness of Nature-Based solutions under different climate change scenarios. Sci. Total Environ. 794, 148515 (2021).
Nowak, D. J., Greenfield, E. J., Hoehn, R. E. & Lapoint, E. Carbon storage and sequestration by trees in urban and community areas of the united States. Environ. Pollut. 178, 229–236 (2013).
Kuehler, E., Hathaway, J. & Tirpak, A. Quantifying the benefits of urban forest systems as a component of the green infrastructure stormwater treatment network. Ecohydrology 10, e1813 (2017).
Schwaab, J. et al. The role of urban trees in reducing land surface temperatures in European cities. Nat. Commun. 12, 6763 (2021).
Carrus, G. et al. Go greener, feel better? The positive effects of biodiversity on the well-being of individuals visiting urban and peri-urban green areas. Landsc. Urban Plan. 134, 221–228 (2015).
Roy, S., Byrne, J. & Pickering, C. A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different Climatic zones. Urban Urban Green. 11, 351–363 (2012).
Khan, T. & Conway, T. M. Vulnerability of common urban forest species to projected climate change and practitioners perceptions and responses. Environ. Manage. 65, 534–547 (2020).
Allen, C. D. et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Ecol. Manag. 259, 660–684 (2010).
Lindner, M. et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Ecol. Manag. 259, 698–709 (2010).
Foden, W. B. et al. Climate change vulnerability assessment of species. WIREs Clim. Change. 10, e551 (2019).
Ordóñez, C. & Duinker, P. N. Climate change vulnerability assessment of the urban forest in three Canadian cities. Clim. Change. 131, 531–543 (2015).
Ordóñez, C. & Duinker, P. N. Assessing the vulnerability of urban forests to climate change. Environ. Rev. 22, 311–321 (2014).
Esperon-Rodriguez, M. et al. Assessing the vulnerability of australia’s urban forests to climate extremes. PLANTS PEOPLE PLANET. 1, 387–397 (2019).
Esperon-Rodriguez, M. et al. Climate change increases global risk to urban forests. Nat. Clim. Change. 12, 950–955 (2022).
Kendal, D. et al. A global comparison of the Climatic niches of urban and native tree populations. Glob Ecol. Biogeogr. 27, 629–637 (2018).
de Rigo, D., Caudullo, G., Houston Durrant, T. & San-Miguel-Ayanz, J. Publications Office of the European Union, Luxembourg,. The European Atlas of Forest Tree Species: modelling, data and information on forest tree species. in European Atlas of Forest Tree Species e01aa69+ (2016).
Bracken, J. T. et al. Maximizing species distribution model performance when using historical occurrences and variables of varying persistency. Ecosphere 13, e3951 (2022).
Franklin, J. & Miller, J. A. Mapping Species Distributions: Spatial Inference and Prediction (Cambridge University Press, 2009).
Marchi, M. et al. ClimateEU, scale-free climate normals, historical time series, and future projections for Europe. Sci. Data. 7, 428 (2020).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1-km Spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
QGIS.org. QGIS Geographic Information System (QGIS Association, 2025).
Giglio, E., Pace, L. & Tammaro, F. Lineamenti Del Paesaggio vegetale Della Conca Aquilana (Italia Centrale). G Bot. Ital. 130, 487–487 (1996).
Curci, G. et al. Building a local climate reference dataset: application to the Abruzzo region (Central Italy), 1930-2019. Int. J. Climatol. joc.7081 (2021). https://doi.org/10.1002/joc.7081
Lange, S. Trend-preserving bias adjustment and statistical downscaling with ISIMIP3BASD (v1.0). Geosci. Model. Dev. 12, 3055–3070 (2019).
Casanueva, A. et al. Testing bias adjustment methods for regional climate change applications under observational uncertainty and resolution mismatch. Atmospheric Sci. Lett. 21, e978 (2020).
CMIP6. CMIP Phase 6 (CMIP6) - Coupled Model Intercomparison Project. (2022). https://wcrp-cmip.org/cmip6/
ESGF. cmip6 - Home | ESGF-CoG. (2024). https://esgf-node.llnl.gov/projects/cmip6/
EC-Earth. EC-Earth – EC-Earth a Global Climate Model. (2024). https://ec-earth.org/
Herrera, S. et al. Uncertainty in gridded precipitation products: influence of station density, interpolation method and grid resolution. Int. J. Climatol. 39, 3717–3729 (2019).
Jang, J. & Woo, S. Y. Native trees as a provider of vital urban ecosystem services in urbanizing new zealand: status quo, challenges and prospects. Land 11, 92 (2022).
Sjöman, H., Morgenroth, J., Sjöman, J. D., Sæbø, A. & Kowarik, I. Diversification of the urban forest—Can we afford to exclude exotic tree species? Urban Urban Green. 18, 237–241 (2016).
Varricchione, M. et al. Urban woods in a small mediterranean city: are native vs alien woods lonely together? Preprint at https://doi.org/ (2024). https://doi.org/10.21203/rs.3.rs-4357615/v1
O’Donnel, M. S. & Ignizio, D. A. Bioclimatic Predictors for Supporting Ecological Applications in the Conterminous United States. Data Series https://pubs.usgs.gov/publication/ds691 (2012). https://doi.org/10.3133/ds691
Woodward, F. I. & Williams, B. G. Climate and plant distribution at global and local scales. Vegetatio 69, 189–197 (1987).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2023).
Au, T. F. et al. Younger trees in the upper canopy are more sensitive but also more resilient to drought. Nat. Clim. Change. 12, 1168–1174 (2022).
Woodward, F. I. Climate and Plant Distribution (Cambridge University Press, 1987).
Aronson, J., Pereira, J. S. & Pausas, J. G. Cork Oak Woodlands on the Edge: Ecology, Adaptive Management, and Restoration (Island, 2012).
Li, J. et al. Critical temperatures controlling the phenology and radial growth of Pinus sylvestris var. Mongolica on the Southern margin of a cold temperate coniferous forest. Ecol. Indic. 126, 107674 (2021).
Matías, L. & Jump, A. S. Interactions between growth, demography and biotic interactions in determining species range limits in a warming world: the case of Pinus sylvestris. Ecol. Manag. 282, 10–22 (2012).
Buras, A. & Menzel, A. Projecting tree species composition changes of European forests for 2061–2090 under RCP 4.5 and RCP 8.5 scenarios. Front Plant. Sci 9, (2019).
Mauri, A. et al. EU-Trees4F, a dataset on the future distribution of European tree species. Sci. Data. 9, 37 (2022).
Mauri, A., Strona, G. & San-Miguel-Ayanz, J. EU-Forest, a high-resolution tree occurrence dataset for Europe. Sci. Data. 4, 160123 (2017).
Pauleit, S. Urban street tree plantings: identifying the key requirements. Proc. Inst. Civ. Eng. - Munic Eng. 156, 43–50 (2003).
Pregitzer, C. C., Sonti, N. F. & Hallett, R. A. Variability in urban soils influences the health and growth of native tree seedlings. Ecol. Restor. 34, 106–116 (2016).
Bosch, J. M. & Hewlett, J. D. A review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration. J. Hydrol. 55, 3–23 (1982).
Komatsu, H., Kume, T. & Otsuki, K. Increasing annual runoff-broadleaf or coniferous forests? Hydrol. Process. 25, 302–318 (2011).
Cape, J. N., Brown, A. H. F., Robertson, S. M. C., Howson, G. & Paterson, I. S. Interspecies comparisons of throughfall and stemflow at three sites in Northern Britain. Ecol. Manag. 46, 165–177 (1991).
Nisbet, T. Water Use by Trees. (2005).
Dymond, C. C. et al. Diversifying managed forests to increase resilience. Can. J. Res. 44, 1196–1205 (2014).
Messier, C. et al. For the sake of resilience and multifunctionality, let’s diversify planted forests! Conserv. Lett. 15, e12829 (2022).
Paquette, A. et al. Praise for diversity: A functional approach to reduce risks in urban forests. Urban Urban Green 62, (2021).
Acknowledgements
This work was partly funded by the Experimental Programme for Intervention on Climate Change Adaptation in Urban Areas of the Italian Ministry for the Environment and led by the Municipality of L’Aquila. We are grateful to developers of the European Tree Atlas, ClimateEU WorldClim and R programming environment for making their valuable products freely available.
Author information
Authors and Affiliations
Contributions
CG conceived and designed the work and acquired, analysed and interpreted the data GC conceived and designed the work and acquired, analysed and interpreted the dataLP revised the workAM revised the workFF revised the workDDT revised the work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Gala, C., Curci, G., Pace, L. et al. A method for preliminary assessment of the vulnerability to climate change of tree species for urban afforestation. Sci Rep 15, 31362 (2025). https://doi.org/10.1038/s41598-025-13806-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-13806-6
