
Abstract
Natural killer (NK) cells play a crucial role in immune surveillance by recognizing and eliminating tumor cells. However, tumors employ various mechanisms to evade NK cell-mediated immunity. NKp30 is a potent activating receptor on NK cells, but its function can be inhibited by specific ligands secreted by cancer cells. Here, we identified dipeptidase 1 (DPEP1) as a novel ligand for NKp30 in KM12C colon cancer cells, using co-immunoprecipitation, confocal microscopy, and flow cytometry. We examined how the DPEP1–NKp30 interaction affects NK cell activity and found that NK cytotoxicity increased in KM12C cells with DPEP1 knockdown but was significantly reduced in HCT116 cells overexpressing DPEP1. We further demonstrated that DPEP1 is secreted via extracellular vesicles and that its interaction with NKp30 suppressed the expression and secretion of perforin 1, granzyme B, CD107a, and interferon-γ in NK92 cells. In a xenograft mouse model treated with NK92 cells, tumors derived from HCT116/DPEP1 cells were significantly larger than those from HCT116/mock cells. Using peripheral blood-derived human NK cells, we confirmed that DPEP1 inhibited both cytotoxicity and granzyme B secretion. These findings suggest that disrupting the DPEP1–NKp30 interaction may enhance NK cell-mediated cytotoxicity and represent a novel therapeutic strategy for cancer immunotherapy.
Similar content being viewed by others


Introduction
Natural killer (NK) cells are innate lymphocytes that provide rapid and potent responses against virally infected or transformed cells. They eliminate harmful cells without the need for prior sensitization or antigen priming1,2,3. Owing to their robust cytolytic potential, NK cells are essential in limiting tumor progression4,5. Upon activation, NK cells release cytotoxic granules containing perforins and granzymes, leading to target cell lysis. Unlike T cells, NK cell activity is governed by a balance between activating and inhibitory receptors, such as killer cell immunoglobulin-like receptors (KIRs), natural cytotoxicity receptors (NCRs), and NKG2D6,7,8 .
NCRs consist of three type I transmembrane receptors: NKp46 (NCR1), NKp44 (NCR2), and NKp30 (NCR3)9,10,11. These receptors recognize ligands on target cells and trigger downstream signaling pathways that culminate in cytotoxic granule release. NKp30, encoded by the NCR3 gene, has three isoforms (a, b, and c) produced via alternative splicing of exon 4 9,12. While isoforms a and b promote cytolysis and cytokine production (e.g., IFN-γ, TNF-α), isoform c is associated with immunosuppressive IL-10 production, particularly in gastrointestinal stromal tumors13.
Multiple ligands for NKp30 have been identified, including B7-H6 (NCR3LG1)14,15,16,17,18galectin-3 (LGALS3)19BAT3 (BAG6)20,21and the cytomegalovirus pp65 protein22. Among these, B7-H6 is a membrane-bound B7 family member that activates NK cells via NKp30 and is expressed in several cancers, including lung, breast, and lymphoma14,16,23,24. BAG6, while initially identified as a membrane ligand that activates NK cells, can also act in soluble form to suppress NK function, particularly in chronic lymphocytic leukemia25. Similarly, galectin-3 contributes to immune evasion by binding NKp30 and impairing NK activity19. In cervical cancer patients, elevated levels of soluble B7-H6 and reduced expression of NKp30 isoform c have been observed26further suggesting that ligand–receptor dynamics can influence NK cell function in tumors.
Dipeptidase 1 (DPEP1), also known as membrane-bound dehydropeptidase-1, is a zinc-dependent metalloproteinase involved in glutathione metabolism and the hydrolysis of various dipeptides27. DPEP1 catalyzes the conversion of leukotriene D4 to E4, a process linked to inflammation and cancer progression28,29,30. Its expression varies by cancer type: it is downregulated in breast cancer and Wilms tumors31,32while in pancreatic ductal adenocarcinoma, it inhibits invasion and enhances chemosensitivity33. In contrast, DPEP1 promotes invasion and metastasis in colon, liver, and prostate cancers34,35,36,37. We previously demonstrated that DPEP1 enhances colon cancer metastasis by modulating E-cadherin expression29yet its immunoregulatory role within the tumor microenvironment remains unclear.
In this study, we identify DPEP1 as a novel NKp30 ligand in colon cancer and investigate how this interaction affects NK cell cytotoxicity. We show that DPEP1 is secreted via extracellular vesicles and binds to NKp30, thereby suppressing NK cell function. Moreover, tumors overexpressing DPEP1 exhibit increased resistance to NK-mediated cytotoxicity in both xenografted mice and human NK cell assays. Our findings suggest that targeting the DPEP1–NKp30 axis may represent a promising immunotherapeutic strategy.
Results
NKp30 on NK cells interacts with DPEP1 in KM12C colon cancer cells
To identify proteins interacting with NKp30, NK92 cells expressing C-terminal His-tagged NKp30 were generated. Following cell lysis, NKp30-His was immobilized on a Ni-NTA column, which was then incubated with lysates from KM12C colon cancer cells. The bound proteins were eluted and analyzed by liquid chromatography–tandem mass spectrometry (LC-MS/MS). Among the identified NKp30-binding candidates, dipeptidase 1 (DPEP1), a receptor involved in neutrophil recruitment38was selected for further analysis.
To examine the cellular localization of DPEP1, HCT116 cells were transfected with either a C-terminal GFP-tagged DPEP1 construct or a GFP control vector. While GFP alone was evenly distributed throughout the cytoplasm and nucleus, DPEP1-GFP localized predominantly to the plasma membrane, weakly in the cytoplasm, and strongly in an area consistent with the endoplasmic reticulum (ER) (Supplementary Fig. S1A). Immunoprecipitation with an anti-GFP antibody confirmed that DPEP1-GFP co-immunoprecipitated with NKp30 (Supplementary Fig. S1B).
To validate this interaction under more physiological conditions, lysates from co-cultured KM12C and NK92 cells were subjected to immunoprecipitation using anti-DPEP1 or anti-NKp30 antibodies. In both cases, reciprocal co-immunoprecipitation confirmed that NKp30 and DPEP1 interact (Fig. 1A). NKp30 is known to exist in three isoforms (a, b, and c), with isoforms a and b generally promoting NK activation, while isoform c has been reported to suppress NK cell activity13. Therefore, we expressed each NKp30 isoform with a C-terminal His tag in NK92 cells and examined their binding to DPEP1 via immunoprecipitation using an anti-DPEP1 antibody. All three isoforms co-immunoprecipitated with DPEP1, likely due to their identical extracellular domains (Fig. 1B). To further confirm the interaction, recombinant DPEP1 (rDPEP1) was purified from the culture medium of HEK293F cells (Fig. 1C). When NK92 cells were incubated with rDPEP1 and stained with anti-NKp30-APC and anti-DPEP1-FITC antibodies, approximately 83% of the NK92 cells showed dual positivity, indicating surface binding of rDPEP1. Although colocalization of NKp30 and DPEP1 in the co-culture of NK92 and KM12C cells was not clearly visualized, confocal microscopy revealed co-localization of NKp30 and rDPEP1 on the plasma membrane of NK92 cells (Fig. 1D).

NKp30 interacts with DPEP1. (A) Co-immunoprecipitation of NKp30 and DPEP1. After co-culturing NK92 and KM12C cells, cell lysates were immunoprecipitated (IP) using either anti-DPEP1 or anti-NKp30 antibodies. Immunocomplexes were analyzed by SDS–PAGE followed by western blotting. (B) Interaction of NKp30 isoforms with DPEP1. NK92 cells were transfected with His-tagged NKp30 isoforms (a/b/c), co-cultured with KM12C cells, and subjected to IP using anti-DPEP1 antibody. NS, not specific. (C) Binding of recombinant DPEP1 to NK92 cells. rDPEP1 was purified from HEK293F cells and visualized by Coomassie Brilliant Blue staining. NK92 cells were treated with rDPEP1, washed, and stained with anti-NKp30/APC and anti-DPEP1/FITC antibodies. NKp30⁺/rDPEP1⁺ NK92 cells were quantified by flow cytometry. Data represent mean ± SD from three independent experiments (n = 3), each performed in triplicate. Statistical significance was assessed using Student’s t-test (**P < 0.01). (D) Colocalization of rDPEP1 and NKp30 on NK92 cells. Cells treated with or without rDPEP1 were fixed and stained with anti-NKp30/FITC and anti-DPEP1/TRITC antibodies. Nuclei were counterstained with DAPI. Arrows indicate colocalization of both proteins on the plasma membrane (confocal microscopy).
Overexpression of DPEP1 attenuates NK cell cytotoxicity
Since NKp30 is a natural cytotoxicity receptor (NCR) that plays a key role in NK cell-mediated killing of target cells, we next investigated whether DPEP1 modulates NK cell cytotoxicity via interaction with NKp30. Western blot analysis of multiple colon cancer cell lines revealed that KM12C and SW620 cells strongly expressed DPEP1, whereas SW480 and HCT116 cells showed little to no expression (Fig. 2A). To evaluate whether DPEP1 affects tumor cell proliferation, DPEP1 was overexpressed in DPEP1-negative SW480 and HCT116 cells. However, no significant differences in cell proliferation were observed compared to vector control cells (Fig. 2B), consistent with our previous report29 that DPEP1 primarily promotes metastasis by regulating E-cadherin, rather than affecting cell proliferation.

DPEP1 overexpression in colon cancer cells decreases NK cell cytotoxicity, whereas its knockdown enhances NK cell cytotoxicity. (A) Lysates of 13 colon cancer cell lines were subjected to SDS–PAGE followed by western blotting using an anti-DPEP1 antibody. (B) Cell proliferation assay. DPEP1-negative HCT116 and SW480 cells were transfected with DPEP1 or vector control, and cell growth was measured using the WST-1 assay. (C, D) DPEP1 knockdown enhances NK cytotoxicity. KM12C (C) or SW620 (D) cells were transfected with DPEP1 siRNA. Half of the cells were used to confirm knockdown by western blotting, and the remaining cells were stained with Calcein-AM, co-cultured with NK92 cells (E: T = 2:1), and assayed for Calcein release. (E, F) DPEP1 overexpression suppresses NK cytotoxicity. SW480 (E) or HCT116 (F) cells were transfected with DPEP1 plasmid. Western blotting confirmed DPEP1 expression. Calcein-AM–labeled cells were co-cultured with NK92 cells (E: T = 2:1), and Calcein release was measured. (G) Attenuation of NK cell cytotoxicity by NKp30 knockdown. NK92 cells were transduced with shRNA targeting NKp30 (shNKp30) or control shRNA (shCtrl), and knockdown was confirmed by western blotting. Calcein-AM–labeled colon cancer cells were incubated with NK92/shNKp30 or NK92/shCtrl cells (E: T = 2:1), and Calcein release was measured. Data represent mean ± SD from three independent experiments (n = 3), each performed in triplicate. Statistical significance was assessed using Student’s t-test (*P < 0.05, **P < 0.01; NS, not significant).
To assess the impact of DPEP1 on NK cell cytotoxicity, NK92 cells were co-cultured with Calcein-AM–labeled target cells, and the release of Calcein from lysed cells was quantified. Knockdown of DPEP1 in KM12C and SW620 cells using siRNA significantly enhanced NK92-mediated cytotoxicity (Fig. 2C, D). Conversely, overexpression of DPEP1 in SW480 and HCT116 cells significantly reduced NK cell–mediated killing compared to vector-transfected controls (Fig. 2E, F). To confirm that this effect is mediated by NKp30, we generated NK92 cells with NKp30 knockdown using lentiviral shRNA. The knockdown was validated by western blotting (Fig. 2G), flow cytometry (Supplementary Fig. S2A), and confocal microscopy (Supplementary Fig. S2B). NK92/shNKp30#1 cells showed markedly reduced NKp30 expression and exhibited decreased cytotoxic activity against both KM12C and K562 target cells compared to NK92/shCtrl cells (Supplementary Fig. S2C, D). As shown in Fig. 2G, cytotoxicity assays revealed that NK92/shCtrl cells exhibited relatively lower cytotoxicity against DPEP1-positive cells (KM12C, SW620) compared with DPEP1-negative cells (SW480, HCT116), likely due to the inhibitory interaction between NKp30 and DPEP1. However, when NKp30-knockdown NK92 cells (NK92/shNKp30) were co-cultured with these targets, they showed reduced cytotoxicity against all four cell lines, regardless of DPEP1 expression. Since NK92/shNKp30 cells lack NKp30 and cannot interact with DPEP1, their reduced cytotoxicity across all targets suggests that DPEP1-overexpressing cancer cells may suppress NK92 cell activity not only through NKp30-mediated inhibition but also via additional mechanisms.
DPEP1 in extracellular vesicles inhibits NK cell cytotoxicity
Although DPEP1 is a membrane-bound protein, it is also secreted and detectable in extracellular vesicles (EVs)39,40,41,42,43. DPEP1, together with CD9 and GAPDH, was detected in cell lysates, conditioned media, and EVs derived from KM12C cells (Fig. 3A). Treatment with GW4869, an inhibitor of EV biogenesis and release, effectively blocked DPEP1 secretion. We further confirmed that DPEP1 binds to the NK cell receptor NKp30 when its extracellular release is not inhibited by GW4869 (Fig. 3B). Flow cytometry analysis showed that approximately 66% of NK92 cells treated with KM12C-conditioned media were positive for DPEP1/FITC staining along with NKp30/APC (Fig. 3C). Colocalization of DPEP1 released from KM12C cells and NKp30 on NK92 cells was also observed by confocal microscopy (Fig. 3D). This colocalization was markedly reduced when NK92 cells were treated with conditioned media from KM12C cells pretreated with GW4869.

Secreted DPEP1 attenuates NK cell cytotoxicity by binding to NKp30. (A) DPEP1 release via EVs. KM12C cells were cultured with or without the EV secretion inhibitor GW4869. Cell lysates, conditioned media, and EVs isolated from the media were analyzed by western blotting. CD9 and GAPDH were used as EV markers, and PARP1 as a control for cell lysates. (B) Interaction between secreted DPEP1 and NKp30. NK92 cells were treated with conditioned media from KM12C cells (± GW4869), and cell lysates were immunoprecipitated with anti-NKp30, followed by western blotting. (C) Flow cytometric analysis of NK92 cells treated with KM12C conditioned media and stained with anti-NKp30/APC and anti-DPEP1/FITC antibodies. (D) Confocal imaging showing colocalization of extracellular DPEP1 and NKp30 on NK92 cells treated with KM12C conditioned media (± GW4869). Cells were stained with anti-NKp30/FITC and anti-DPEP1/TRITC; DAPI was used for nuclear staining. Arrows indicate colocalization at the plasma membrane. (E) Increased NK cytotoxicity in response to DPEP1 knockdown. KM12C/shDPEP1 cells (± GW4869) were labeled with Calcein-AM, cocultured with NK92 cells, and Calcein release was measured. (F) Suppressed NK cytotoxicity by DPEP1 overexpression. HCT116 cells overexpressing DPEP1 (± GW4869) were labeled with Calcein-AM, cocultured with NK92 cells, and Calcein release was measured. (G) Conditioned media of KM12C cells (± GW4869, ±rDPEP1) were assessed as in (E) and (F). (H) Conditioned media from KM12C/shDPEP1 and shCtrl cells were immunoprecipitated with anti-DPEP1 or control IgG. The DPEP1-depleted media were then applied to NK92 cells, followed by NKp30 immunoprecipitation and western blotting. (I) DPEP1 depletion increases IFN-γ release from NK cells. NK92 cells were treated with DPEP1-suppressed or DPEP1-depleted conditioned media, and IFN-γ levels were quantified. Data represent mean ± SD from three independent experiments (n = 3), each performed in triplicate. Statistical significance was assessed using Student’s t-test (*P < 0.05, **P < 0.01, ***P < 0.001; NS, not significant).
To evaluate the functional relevance of DPEP1 in NK cell regulation, we generated DPEP1-knockdown KM12C cells (Fig. 3E) and DPEP1-overexpressing HCT116 cells (Fig. 3F), and tested NK cell cytotoxicity in the presence or absence of GW4869. DPEP1 knockdown enhanced NK cell cytotoxicity, whereas DPEP1 overexpression reduced it. Furthermore, blocking EV secretion with GW4869 increased NK92 cytotoxicity by approximately 8%, and this effect was reversed by the addition of rDPEP1 (Fig. 3G). As CD9 in EVs has also been implicated in suppressing NK cell function44we next examined whether DPEP1 influences CD9 secretion and IFN-γ release by NK92 cells. Conditioned media from KM12C/shDPEP1 and KM12C/shCtrl cells were subjected to immunoprecipitation with anti-DPEP1 or control IgG to remove DPEP1. The resulting supernatants were then added to NK92 cell cultures. NK92 cell lysates were subsequently immunoprecipitated with an anti-NKp30 antibody, and interaction between DPEP1 and NKp30 was detected only in lysates treated with media that had not been depleted of DPEP1 (Fig. 3H–I). While CD9 secretion was unaffected by DPEP1 knockdown, IFN-γ secretion from NK cells increased by approximately 23% when treated with DPEP1-deficient or DPEP1-depleted conditioned media. Consistent with these findings, NK92 cells treated with conditioned media from KM12C/shDPEP1 cells showed no detectable DPEP1 and lacked colocalization with NKp30 (Supplementary Fig. S3), whereas colocalization was evident in NK92 cells treated with conditioned media from KM12C/shCtrl cells.
DPEP1 inhibits the production of cytotoxic granules in NK cells
To determine whether the interaction between NKp30 and DPEP1 affects the secretion of cytotoxic granules in NK cells, rDPEP1 was added to the conditioned medium. As shown in Fig. 4A, coculture of NK92 cells with HCT116 cells increased the mRNA expression of perforin 1 and granzyme B, while the addition of rDPEP1 to the coculture reduced their mRNA levels. Western blot analysis further confirmed the reduction in perforin 1 and granzyme B protein levels, and the presence of rDPEP1 was verified in the cell lysate (Fig. 4B). To visualize these effects, NK92 cells were treated with resveratrol—a known enhancer of NK cytotoxicity45—alone or in combination with rDPEP1, and perforin 1 distribution was assessed via confocal microscopy (Fig. 4C). Perforin 1 expression was enhanced by resveratrol but significantly diminished in cells treated with rDPEP1. CD107a, a marker of NK cell degranulation that correlates with the release of perforin 1 and granzyme B46,47was measured in cocultures of NK92 and HCT116 cells. Flow cytometry revealed that the resveratrol-induced increase in CD107a expression was suppressed by rDPEP1, while cilastatin—an enzymatic inhibitor of DPEP1—had no such effect (Fig. 4D). Similarly, rDPEP1 significantly reduced IFN-γ secretion from NK92 cells stimulated with resveratrol, whereas cilastatin did not (Fig. 4E). In addition to resveratrol, IL-15 was used to activate NK92 cells, given its ability to promote NKp30 and NKp46 expression and enhance cytotoxic function. When NK92 cells were treated with IL-2 and IL-15, the secretion of IFN-γ (Fig. 4F) and granzyme B (Fig. 4G) was significantly increased. However, the addition of rDPEP1 markedly reduced the secretion of both cytokines.

Inhibition of cytotoxic granule secretion in NK cells by DPEP1. (A) rDPEP1 suppresses the mRNA expression of perforin 1 and granzyme B. NK92 and HCT116 cells were cocultured with or without rDPEP1, and qRT-PCR was performed to assess mRNA levels. (B) Western blot analysis of perforin 1 and granzyme B protein levels after coculture of NK92 and HCT116 cells in the presence or absence of rDPEP1. (C) Representative confocal microscopy images showing the effect of rDPEP1 on perforin 1 localization in NK92 cells treated with resveratrol alone or in combination with rDPEP1. Cells were stained with perforin 1/FITC and DAPI. (D) CD107a expression is inhibited by rDPEP1. NK92 and HCT116 cells were cocultured and treated with resveratrol, rDPEP1, or cilastatin (alone or in combination). CD107a and CD56 expression was analyzed by flow cytometry. (E) Inhibition of IFN-γ secretion by rDPEP1. NK92 cells were treated with resveratrol, rDPEP1, or cilastatin, and IFN-γ levels in the culture medium were measured by ELISA. (F, G) rDPEP1 inhibits IFN-γ and granzyme B secretion induced by IL-2 and IL-15. NK92 cells were treated with IL-2 and/or IL-15, with or without rDPEP1. Secreted IFN-γ (F) and granzyme B (G) were quantified. Data are presented as mean ± SD from three independent experiments (n = 3), each performed in triplicate. Statistical significance was determined using Student’s t-test (**P < 0.01, ***P < 0.001; NS, not significant).
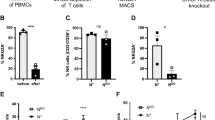
We also investigated whether DPEP1 binding to NKp30 alters the expression of NKp30 isoforms (Supplementary Fig. S4). As previously reported13NKp30 isoforms a and b are immunostimulatory, while isoform c is immunosuppressive. rDPEP1 treatment increased the expression of isoform c and significantly reduced isoform b mRNA in both NK92 cells and human peripheral blood NK (PBNK) cells. In addition, expression levels of TNF-α, IFN-γ, perforin 1, and granzyme B were decreased. A slight increase in CD3ζ expression and minimal changes in CD16a, CD56, NKG2D, and IL-10 were observed in both cell types. Although PBNK cells generally exhibit stronger cytotoxicity than NK92 cells, rDPEP1 treatment suppressed NK cell activity to a similar extent in both cell types.
DPEP1 increases resistance of HCT116 xenografts to NK92 cytotoxicity
To investigate whether DPEP1 expression confers resistance to NK cell–mediated cytotoxicity in vivo, we employed a mouse xenograft model. HCT116 colon cancer cells overexpressing DPEP1 (HCT116/DPEP1) or carrying a control vector (HCT116/mock) were subcutaneously injected into the dorsal flanks of athymic nude mice. Tumor growth was monitored over time using calipers. Under basal conditions, tumors formed by the two groups showed no significant differences in size (Fig. 5A, B) or weight (Fig. 5C). However, when NK92 cells were administered intratumorally three times at 3-day intervals starting on day 10 after tumor inoculation, tumors derived from HCT116/mock cells exhibited a significant reduction in size by day 25 compared with those from HCT116/DPEP1 cells, indicating that DPEP1 impairs NK cell–mediated tumor suppression. To confirm the presence of DPEP1 and NKp30 in the tumors, excised tumor tissues were homogenized and analyzed by western blotting (Fig. 5D). DPEP1 was detected in tumors from the HCT116/DPEP1 group, whereas NKp30 expression was not observed, likely due to the transient survival of NK92 cells, which typically persist only for a few days before undergoing apoptosis or being cleared by the host immune system. Collectively, these findings suggest that DPEP1 functions as an immunomodulatory factor that suppresses NK cell cytotoxic activity within the tumor microenvironment, and that targeting DPEP1 may enhance NK cell–mediated antitumor immunity.

Attenuation of NK cell cytotoxicity by DPEP1 overexpression in a xenograft model. HCT116/mock or HCT116/DPEP1 cells were subcutaneously injected into the dorsal flank of athymic nude mice. On day 10 post-injection, NK92 cells were administered intratumorally three times at 3-day intervals. Tumor dimensions were measured every 5 days using a digital caliper, and mice were euthanized on day 25 for tumor collection and analysis. (A) Tumor volume was calculated at the indicated time points using caliper measurements. (B) Excised tumors were photographed for visual comparison. (C) Tumor weights were recorded post-sacrifice. (D) Tumor tissues were homogenized, and protein lysates were analyzed by western blotting to assess DPEP1 expression. Each group contained 5 mice, and data are presented as mean ± SD from three independent experiments (n = 3). Statistical significance was determined using Student’s t-test (*P < 0.05, **P < 0.01; NS, not significant).
DPEP1 inhibits the cytotoxicity of human peripheral blood NK (PBNK) cells
To determine whether the inhibitory effect of DPEP1 on NK cell cytotoxicity also applies to primary human NK cells, we evaluated its function in peripheral blood NK (PBNK) cells, rather than the NK92 cell line. Consistent with our findings in NK92 cells, PBNK cells were co-cultured with target cells either expressing or lacking DPEP1, and cytotoxicity was assessed (Fig. 6A). Target cells such as HCT116 and K562, which do not endogenously express DPEP1, were more susceptible to PBNK-mediated killing compared to cells treated with recombinant DPEP1 (rDPEP1) or transfected to overexpress DPEP1 (E: T ratio = 2:1). In contrast, KM12C cells, which naturally express DPEP1, exhibited increased susceptibility to PBNK-mediated cytotoxicity when DPEP1 was knocked down. These results further support that DPEP1 suppresses the cytotoxic activity of primary human NK cells.

DPEP1 suppresses the cytotoxic function of human peripheral blood NK (PBNK) cells. (A) Target cells (K562, HCT116, and KM12C) were either treated with rDPEP1, transfected to overexpress DPEP1, or subjected to DPEP1 knockdown, then labeled with Calcein-AM. Labeled cells were co-cultured with human PBNK cells at an E: T ratio of 2:1. Cytotoxicity was assessed by quantifying Calcein released into the culture medium. (B) PBNK cells were co-cultured with SW480, HCT116, or KM12C cells with DPEP1 overexpression or knockdown, in the presence or absence of rDPEP1 (0.1 µg/mL). After 12 h, granzyme B levels in the culture medium were measured. (C) HCT116 cells were co-cultured with PBNK cells with or without rDPEP1. After incubation, cell lysates were collected, and western blotting was performed to detect granzyme B and perforin 1 protein levels. Data represent mean ± SD from three independent experiments (n = 3), each performed in triplicate. Statistical significance was assessed using Student’s t-test (*P < 0.05, **P < 0.01, P***<0.001).
To further validate the immunosuppressive effect of DPEP1, we measured granzyme B secretion from PBNK cells after co-culture with SW480, HCT116, or KM12C cells manipulated to overexpress or knock down DPEP1, with or without rDPEP1 treatment (Fig. 6B). Overexpression of DPEP1 in SW480 and HCT116 cells significantly reduced granzyme B secretion from PBNK cells, whereas knockdown of DPEP1 in KM12C cells enhanced it. Notably, treatment with rDPEP1 reversed this increase, confirming that DPEP1 inhibits granzyme B release from primary NK cells. Lastly, we examined intracellular expression of granzyme B and perforin 1 in HCT116 cells co-cultured with PBNK cells, in the presence or absence of rDPEP1 (Fig. 6C). Co-culture with PBNK cells increased granzyme B and perforin 1 protein levels in HCT116 cells, whereas rDPEP1 treatment attenuated this upregulation.
Collectively, these findings demonstrate that DPEP1 suppresses both cytotoxic activity and effector molecule release from primary human NK cells, supporting its potential role as a novel immune checkpoint regulator.
Discussion
NK cells serve as a first line of defense in immune surveillance and tumor elimination, relying on a fine-tuned balance of activating and inhibitory receptor–ligand interactions1,3,48. In this study, we identify DPEP1 as a novel ligand for the activating receptor NKp30 and demonstrate that its interaction suppresses NK cell cytotoxicity.
While NKp30 ligands such as B7-H6, Gal3, and BAG6 are well-established, their functional roles vary depending on whether they are membrane-bound or secreted. For instance, soluble B7-H6, elevated in cervical cancer patients, impairs NK cell cytotoxicity despite increased NK cell numbers26. Likewise, Gal3 and BAG6, when secreted, suppress NK function by binding NKp30 19,20,21. These examples highlight how ligand secretion and tumor microenvironmental context determine NKp30-mediated outcomes.
DPEP1 has been reported to promote invasion and metastasis in several cancers, including colon34,35liver36prostate37ovarian49and renal cancers50and its overexpression is generally associated with poor prognosis. Recently, Liu et al.51 demonstrated that DPEP1 binds to MYC and promotes cell proliferation in colon cancer, with overexpression in HCT116 cells enhancing proliferation both in vitro and in vivo. In contrast, our results showed that DPEP1 overexpression in HCT116 cells led to only a slight, non-significant increase in proliferation in both in vitro assays and mouse xenografts compared with controls. This discrepancy suggests that the proliferative effects of DPEP1 may be context dependent, requiring additional oncogenic signals or tumor microenvironmental factors that were different in our experimental models. Previously, we reported29 that DPEP1 contributes to colon cancer metastasis by regulating E-cadherin expression rather than by directly promoting cell proliferation. Our present findings extend this oncogenic role by demonstrating that DPEP1 also facilitates immune evasion through direct binding to NKp30, thereby suppressing NK cell cytotoxicity. Notably, DPEP1 secreted via extracellular vesicles (EVs) was sufficient to inhibit the secretion of perforin-1, granzyme B, and IFN-γ from NK cells—mimicking the immunosuppressive effects of established NKp30 ligands such as Gal3 and BAG6.
However, DPEP1’s function appears context-dependent. It is downregulated in cancers such as breast31Wilms tumor32and pancreatic ductal adenocarcinoma36suggesting tumor-specific roles. Interestingly, Gal3 is upregulated in several cancers where DPEP1 is suppressed, including breast, gastric, and pancreatic cancers52,53,54indicating possible redundancy or compensation among NKp30 ligands. We also observed that DPEP1 binds all three NKp30 isoforms, which have distinct roles in immune regulation: isoforms a and b promote cytotoxicity, while isoform c is immunosuppressive13. Treatment with recombinant DPEP1 decreased expression of isoform b and reduced IFN-γ and TNF-α secretion in NK92 and primary NK cells, raising the possibility that DPEP1 influences isoform expression or downstream signaling to favor immune suppression.
Despite these insights, several limitations should be acknowledged. First, most functional assays were performed in vitro using NK92 or primary NK cells, which may not fully reflect NK cell complexity in the human tumor microenvironment. Second, the xenograft model—although supportive—lacks the full diversity and regulation of immune cell populations seen in human cancers. Third, we did not perform in vivo rescue experiments (e.g., DPEP1 blockade or NKp30 inhibition) to confirm the specificity of the DPEP1–NKp30 interaction. Fourth, tumor heterogeneity in ligand/receptor expression and EV composition could influence the extent of DPEP1-mediated suppression. Fifth, DPEP1 expression varies widely across tumor types, and its metabolic functions related to peptide and glutathione metabolism suggest that therapeutic targeting must be approached cautiously. Finally, the mode of DPEP1 presentation (membrane-bound vs. EV-associated) and NKp30 isoform distribution in different tumor contexts may alter the immunological outcome.
In summary, our study identifies DPEP1 as a novel immunosuppressive NKp30 ligand secreted via EVs by colon cancer cells. By attenuating NK cell cytotoxicity, DPEP1 may contribute to immune evasion within the tumor microenvironment. Targeting the DPEP1–NKp30 axis could represent a new strategy to boost innate immune responses against cancer. Future work should focus on mapping the structural interaction site, dissecting isoform-specific signaling pathways, evaluating the impact of DPEP1 inhibition in patient-derived tumor models, and exploring its broader effects on other immune cell subsets to better assess therapeutic potential.
Methods
Cell culture
NK-92 cells were cultured in α-MEM (LM008-02; Welgene, Gyeongbul, South Korea) supplemented with 12.5% fetal bovine serum (FBS; Gibco, NY, USA), 12.5% horse serum (Gibco), 0.2 mM inositol, 0.02 mM folic acid, 0.1 mM 2-mercaptoethanol (all from Sigma-Aldrich, St. Louis, MO, USA), and 20 ng/mL IL-2 (200-02; PeproTech, Rocky Hill, NJ, USA). Human peripheral blood NK (PBNK) cells were obtained from Stemcell Technologies (70036; Vancouver, BC, Canada) and expanded using the ImmunoCult NK Cell Expansion Kit, following the manufacturer’s instructions. PBNK cells were maintained in RPMI-1640 (LM011-01; Welgene) supplemented with 10% FBS, 40 ng/mL IL-2, and 20 ng/mL IL-15 (PeproTech).
Colon cancer cell lines (Caco2, Colo205, HT29, HCT116, LoVo, KM12C, KM12SM, HCT15, LS174T, RKO, DLD1, SW480, and SW620) were obtained from the Korean Cell Line Bank (KCLB; Seoul National University, Seoul, South Korea) and the American Type Culture Collection (ATCC; Manassas, VA, USA). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Gibco) with 10% FBS and antibiotics (100 U/mL penicillin, 100 µg/mL streptomycin) at 37 °C in a humidified 5% CO₂ incubator.
RNA interference and qRT-PCR
DPEP1-specific and control siRNAs were purchased from Bioneer (Daejeon, South Korea). The DPEP1 siRNA sequence was 5′-CAG UUC UGG UCC GUG UAC AdTdT-3′. Colorectal cancer cells were transfected with siDPEP1 or control siRNA using Lipofectamine RNAiMAX (Invitrogen, Carlsbad, CA, USA). Short hairpin RNAs (shRNAs) targeting DPEP1 or NKp30 were purchased from Sigma-Aldrich. Lentiviral particles were produced by transfecting Lenti-X 293T cells (Takara Bio, CA, USA) with the shRNA vector. Viral supernatants were collected 48 h post-transfection and used to transduce target cells. Stable cell lines were selected using 2 µg/mL puromycin (Clontech, CA, USA). Total RNA was extracted using an RNA extraction kit (Biofact, South Korea), and cDNA was synthesized using the GoScript reverse transcription system (Promega, Madison, WI, USA). Quantitative real-time PCR (qRT–PCR) was performed using AccuPower 2× GreenStar qPCR Master Mix (Bioneer, Daejeon, Korea) on a QuantStudio 3 Real-Time PCR System, and data were analyzed with QuantStudio Design & Analysis Software v1.4 (Applied Biosystems, Foster City, CA, USA). Relative gene expression was calculated using the 2^–ΔΔCt method. Primer sequences were: β-actin (F: 5′-caaacatgatctgggtcatcttctc-3′, R: 5′-gctcgtcgttcgacaacggct-3′), Perforin (F: 5′-atgtaaccagggccaaagtca-3′, R: 5′-gtgccgtagttggagataagc-3′), Granzyme B (F: 5′-gcagatgcagacttttccttc-3′, R: 5′-cacagggataaactgctgggt-3′).
Plasmids, transfection, immunoprecipitation, and Recombinant protein production
Plasmids encoding DPEP1-Flag (HG13543-CH) and NKp30-His (RC201964) were obtained from Sino Biological (Beijing, China) and OriGene (Rockville, MD, USA), respectively. In addition, DPEP1 (NM_004413) was cloned into the pEGFPN2 or pEGFPC2 vector. The NKp30 isoforms NKp30-a (NM_147130), NKp30-b (NM_001145466), and NKp30-c (NM_001145467) were amplified from NK-92 cell cDNA and subcloned into the pcDNA3.1/Myc-His vector (Invitrogen). In this study, NKp30-a isoform was used to achieve NKp30 overexpression. All constructs were verified by DNA sequencing and validated by western blot analysis. Cells were transfected using Lipofectamine 3000 (Invitrogen) according to the manufacturer’s instructions. For immunoprecipitation, cells were lysed in CHAPS buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% CHAPS, 5% glycerol, 2 mM EGTA, 1 mM DTT, protease inhibitor cocktail (Sigma-Aldrich)] for 20 min on ice. Lysates were incubated with specific antibodies and Protein A/G agarose beads (sc-2003; Santa Cruz Biotechnology, Dallas, TX, USA) for 6 h at 4 °C, followed by SDS–PAGE and western blotting. Recombinant DPEP1 (rDPEP1) was produced by transfecting FreeStyle 293 F cells (Thermo Fisher Scientific) with a pCMV6-DPEP1 plasmid (OriGene). Culture supernatants were collected and rDPEP1 was purified using a protein purification kit (Thermo Fisher Scientific).
Culture media concentration and extracellular vesicle (EV) isolation
Conditioned culture medium (CM) was collected and sequentially centrifuged at 300 × g for 10 min and 2,000 × g for 30 min to remove cell debris. The resulting supernatant was concentrated using Amicon Ultra-15 centrifugal filter units with a 30 kDa molecular weight cut-off (Millipore, USA) at 4,000 × g until the desired volume was reached, according to the manufacturer’s instructions. For EV isolation, KM12C cell culture supernatants were processed using either the Total Exosome Isolation Reagent (Invitrogen) or ultracentrifugation. For chemical precipitation, the supernatant was incubated overnight with the isolation reagent and centrifuged at 10,000 × g for 1 h. For ultracentrifugation, the supernatant was centrifuged at 120,000 × g for 2 h at 4 °C (Beckman Coulter). The EV pellets were resuspended in PBS or SDS sample buffer for western blot analysis.
Western blotting
Cells were lysed in RIPA buffer [50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% NP-40, 0.25% sodium deoxycholate, 1 mM PMSF, 1 mM sodium orthovanadate, protease inhibitors]. Protein lysates were separated by SDS–PAGE, transferred to PVDF membranes, and probed with the following primary antibodies: β-actin, GAPDH, His-tag, and NKp30 (clone CLH9) (Santa Cruz Biotechnology); DPEP1 (clone E4O6J), PARP1 (#9542), CD9 (clone D3H4P), perforin (clone E7D8R), and granzyme B (clone D6E9W) (Cell Signaling Technology, Danvers, MA, USA); and NKp30 (clone P30-15) (BioLegend, San Diego, CA, USA). Signals were detected using enhanced chemiluminescence (ECL) reagents (EMD Millipore, Burlington, MA, USA) and visualized with an Ez-Capture MG imaging system (Atto, Tokyo, Japan).
Flow cytometry and CD107a degranulation assay
NK92 cells (5 × 10⁴ cells/well) were pretreated with recombinant DPEP1 (rDPEP1; 0.1 µg/mL) for 2 h, washed, and subsequently stained on ice for 30 min with PE- or APC-conjugated anti-NKp30 (W20238A, clone P30-15), FITC-conjugated anti-CD107a (clone H4A3), or APC-conjugated anti-CD56 (clone QA17A16) (all from BioLegend). Flow cytometry data were acquired using a FACSVerse cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and analyzed with FlowJo software v7.6.1 (BD Biosciences). For the degranulation assay, NK92 cells (2 × 10⁴/well) were co-cultured with target cells at different E: T ratios for 6 h in the presence of anti-CD107a-FITC and anti-CD56-APC antibodies before flow cytometry.
Cytotoxicity assay
Target cells were labeled with 2 µM Calcein-AM (Invitrogen) for 30 min at 37 °C, washed three times with PBS, and seeded at 1 × 10⁴ cells/well in U-bottom 96-well plates. Cells were co-cultured with NK92 or primary human NK (PBNK) cells at various effector-to-target (E: T) ratios for 4–8 h. Plates were centrifuged at 300 × g for 10 min, and the supernatant was carefully transferred to a black 96-well plate. Fluorescence was measured using a FilterMax F3 plate reader (Molecular Devices, Sunnyvale, CA, USA) with SoftMax Pro software v6.5.1 (Molecular Devices), at an excitation wavelength of 485 nm and an emission wavelength of 535 nm. Specific lysis (%) was calculated as: (Experimental − Spontaneous) / (Maximum − Spontaneous) × 100.
Enzyme-linked immunosorbent assay (ELISA)
NK92 or PBNK cells (1 × 10⁵) were cultured in 24-well plates for 24 h in the presence or absence of recombinant DPEP1 (rDPEP1; 0.1 µg/mL), resveratrol (10 µM), cilastatin (20 µM), IL-2 (20 ng/mL), and/or IL-15 (20 ng/mL). Following incubation, culture supernatants were collected and analyzed for granzyme B (R&D Systems) and IFN-γ (BD Biosciences) using ELISA kits, according to the manufacturers’ instructions. Absorbance was measured at 450 nm using a microplate reader.
Cell proliferation assay
To assess cell proliferation, 5 × 10³ cells were seeded in 96-well plates. After incubation for the indicated time points, WST-1 reagent (Roche Applied Science, Penzberg, Germany) was added to each well and incubated for 2 h at 37 °C. Absorbance was measured at 450 nm using a microplate reader.
Confocal microscopy
Cells were cultured in 8-well ibidi chambers (ibidi GmbH, Germany) and transfected with pEGFP or pEGFP-DPEP1. NK92 cells were seeded onto poly-L-lysine-coated plates and treated with rDPEP1 (0.1 µg/mL) or EVs (1 µg/mL) for 2 h. Cells were fixed with Cytofix (BD Biosciences), blocked with 1% BSA, and stained with anti-DPEP1 and anti-NKp30 antibodies. Imaging was performed using a laser scanning confocal microscope (LSM 800; Carl Zeiss Microscopy GmbH, Jena, Germany) and analyzed with ZEN software v2.5 (Carl Zeiss Microscopy).
Animal experiments
All animal protocols were approved by the KRIBB Institutional Animal Care and Use Committee (KRIBB-AEC-24317). Female athymic nude (nu/nu) mice (DBL, South Korea), 6–8 weeks old, were subcutaneously injected with HCT116/mock or HCT116/DPEP1 cells (7 × 106) in 200 µL PBS into the dorsal flank. At 10 days post-implantation, when tumors reached approximately 80 mm³ and became palpable, NK-92 cells (1 × 10⁷) were administered intratumorally three times at 3-day intervals. To support NK-92 cell activity, IL-2 (30,000 IU) and IL-15 (0.5 µg/kg) were co-administered. Tumor dimensions were measured every 5 days using calipers, and volumes were calculated as: Volume (mm³) = Length × Width × Height × 0.52. Since tumor volumes typically exceeded 1,500 mm³ by day 30, mice were euthanized on day 25 for tumor collection and weighing. Euthanasia was performed according to institutional animal care guidelines using CO₂ inhalation by the gradual fill method, in accordance with the AVMA Guidelines for the Euthanasia of Animals (2020)55.
Statistical analysis
All experiments were independently performed at least three times, with each experiment including triplicate technical measurements. Data are presented as mean ± standard deviation (SD) of biological replicates, unless otherwise stated. Statistical significance was assessed using one-way ANOVA or Student’s t-test (GraphPad Prism 9.4 and Microsoft Excel). P values were considered significant as follows: P < 0.05 (*), P < 0.01 (**), and P < 0.001 (***).
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Abbreviations
- B7H6:
-
B7 homolog 6
- DPEP1:
-
Dipeptidase 1
- ER:
-
Endoplasmic reticulum
- EV:
-
Extracellular vesicle
- Gal3:
-
Galectin-3
- GFP:
-
Green fluorescent protein
- GRZM:
-
Granzyme B
- IFN-γ:
-
Interferon-γ
- IL2:
-
Interleukin 2
- IL10:
-
Interleukin 10
- IL15:
-
Interleukin 15
- NCR3:
-
Natural cytotoxicity receptor 3
- NK:
-
Natural killer
- NKG2D:
-
Natural killer group 2 member D
- TNF-α:
-
Tumor necrosis factor-α
References
Guillerey, C., Huntington, N. D. & Smyth, M. J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 17, 1025–1036 (2016).
Waldhauer, I. & Steinle, A. NK cells and cancer immunosurveillance. Oncogene 27, 5932–5943 (2008).
Hamerman, J. A., Ogasawara, K. & Lanier, L. L. NK cells in innate immunity. Curr. Opin. Immunol. 17, 29–35 (2005).
Kim, H. R. et al. Anti-cancer activity and mechanistic features of a NK cell activating molecule. Cancer Immunol. Immunother. 58, 1691–1700 (2009).
Muntasell, A. et al. Targeting NK-cell checkpoints for cancer immunotherapy. Curr. Opin. Immunol. 45, 73–81 (2017).
Sivori, S. et al. Human NK cells: surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 16, 430–441 (2019).
Biassoni, R. & Malnati, M. S. Human natural killer receptors, Co-Receptors, and their ligands. Curr. Protoc. Immunol. 121, e47 (2018).
Sivori, S. et al. TLR/NCR/KIR: which one to use and when?? Front. Immunol. 5, 105 (2014).
Pende, D. et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J. Exp. Med. 190, 1505–1516 (1999).
Barrow, A. D., Martin, C. J. & Colonna, M. The natural cytotoxicity receptors in health and disease. Front. Immunol. 10, 909 (2019).
Kruse, P. H., Matta, J., Ugolini, S. & Vivier, E. Natural cytotoxicity receptors and their ligands. Immunol. Cell. Biol. 92, 221–229 (2014).
Sivakamasundari, R., Raghunathan, A., Zhang, C. Y., Chowdhury, R. R. & &Weissman, S. M. Expression and cellular localization of the protein encoded by the 1C7 gene: a recently described component of the MHC. Immunogenetics 51, 723–732 (2000).
Delahaye, N. F. et al. Alternatively spliced NKp30 isoforms affect the prognosis of Gastrointestinal stromal tumors. Nat. Med. 17, 700–707 (2011).
Brandt, C. S. et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J. Exp. Med. 206, 1495–1503 (2009).
Joyce, M. G. et al. Crystal structure of human natural cytotoxicity receptor NKp30 and identification of its ligand binding site. Proc. Natl. Acad. Sci. 108, 6223–6228 (2011).
Li, Y., Wang, Q. & Mariuzza, R. A. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J. Exp. Med. 208, 703–714 (2011).
Kaifu, T., Escalière, B., Gastinel, L. N., Vivier, E. & Baratin, M. B7-H6/NKp30 interaction: a mechanism of alerting NK cells against tumors. Cell. Mol. Life Sci. 68, 3531–3539 (2011).
Ponath, V. et al. Secreted ligands of the NK cell receptor NKp30: B7-H6 is in contrast to BAG6 only marginally released via extracellular vesicles. Int. J. Mol. Sci. 22, 2189 (2021).
Wang, W. et al. Tumor-released Galectin-3, a soluble inhibitory ligand of human NKp30, plays an important role in tumor escape from NK cell attack. J. Biol. Chem. 289, 33311–33319 (2014).
von Pogge, E. et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 27, 965–974 (2007).
Binici, J. et al. A soluble fragment of the tumor antigen BCL2-associated athanogene 6 (BAG-6) is essential and sufficient for Inhibition of NKp30 receptor-dependent cytotoxicity of natural killer cells. J. Biol. Chem. 288, 34295–34303 (2013).
Arnon, T. I. et al. Inhibition of the NKp30 activating receptor by pp65 of human cytomegalovirus. Nat. Immunol. 6, 515–523 (2005).
Chen, H. et al. Deficiency of N-linked glycosylation impairs immune function of B7-H6. Front. Immunol. 14, 1255667 (2023).
Schlecker, E. et al. Metalloprotease-mediated tumor cell shedding of B7-H6, the ligand of the natural killer cell-activating receptor NKp30. Cancer Res. 74, 3429–3440 (2014).
Reiners, K. S. et al. Soluble ligands for NK cell receptors promote evasion of chronic lymphocytic leukemia cells from NK cell anti-tumor activity. Blood 121, 3658–3665 (2013).
Gutierrez-Silerio, G. Y. et al. Non-fitness status of peripheral NK cells defined by decreased NKp30 and perforin, and increased soluble B7H6, in cervical cancer patients. Immunology 168, 538–553 (2023).
Nakagawa, H. et al. Assignment of the human renal dipeptidase gene (DPEP1) to band q24 of chromosome 16. Cytogenet. Cell. Genet. 59, 258–260 (1992).
Yamaya, M. et al. Dipeptidase inhibitor and epithelial removal potentiate leukotriene D4-induced human tracheal smooth muscle contraction. Respir Physiol. 111, 101–109 (1998).
Park, S. Y. et al. Dehydropeptidase 1 promotes metastasis through regulation of E-cadherin expression in colon cancer. Oncotarget 7, 9501–9512 (2016).
Samuelsson, B. Leukotrienes: mediators of immediate hypersensitivity reactions and inflammation. Science 220, 568–575 (1983).
Green, A. R. et al. Loss of expression of chromosome 16q genes DPEP1 and CTCF in lobular carcinoma in situ of the breast. Breast Cancer Res. Treat. 113, 59–66 (2009).
Austruy, E. et al. Isolation of kidney complementary DNAs down-expressed in wilms’ tumor by a subtractive hybridization approach. Cancer Res. 53, 2888–2894 (1993).
Zhang, G. et al. DPEP1 inhibits tumor cell invasiveness, enhances chemosensitivity and predicts clinical outcome in pancreatic ductal adenocarcinoma. Plos One. 7, e31507 (2012).
McIver, C. M., Lloyd, J. M., Hewett, P. J. & Hardingham, J. E. Dipeptidase 1: a candidate tumor-specific molecular marker in colorectal carcinoma. Cancer Lett. 209, 67–74 (2004).
Eisenach, P. A. et al. Dipeptidase 1 (DPEP1) is a marker for the transition from low-grade to high-grade intraepithelial neoplasia and an adverse prognostic factor in colorectal cancer. Br. J. Cancer. 109, 694–703 (2013).
Chen, J., Jin, H., Zhou, H., Hei, X. & Liu, K. Research into the characteristic molecules significantly affecting liver cancer immunotherapy. Front. Immunol. 14, 1029427 (2023).
Xiang, C. et al. Joint analysis of WES and RNA-Seq identify signature genes related to metastasis in prostate cancer. J. Cell. Mol. Med. 27, 1947–1958 (2023).
Choudhury, S. R. et al. Dipeptidase-1 is an adhesion receptor for neutrophil recruitment in lungs and liver. Cell 178, 1205–1221 (2019).
Welsh, J. A. et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J. Extracell. Vesicles. 13, e12404 (2024).
Dash, M., Palaniyandi, K., Ramalingam, S., Sahabudeen, S. & Raja, N. S. Exosomes isolated from two different cell lines using three different isolation techniques show variation in physical and molecular characteristics. Biochim. Biophys. Acta Biomembr. 1863, 183490 (2021).
van Niel, G. et al. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 121, 337–349 (2001).
Liu, Z. et al. Isolation and characterization of human urine extracellular vesicles. Cell. Stress Chaperones. 23, 943–953 (2018).
Zhang, Q. et al. Supermeres are functional extracellular nanoparticles replete with disease biomarkers and therapeutic targets. Nat. Cell. Biol. 23, 1240–1254 (2021).
Gonzalez, V. D. et al. High-grade serous ovarian tumor cells modulate NK cell function to create an immune-tolerant microenvironment. Cell. Rep. 36, 109632 (2021).
Lee, Y. J. & Kim, J. Resveratrol activates natural killer cells through Akt- and mTORC2-Mediated c-Myb upregulation. Int. J. Mol. Sci. 21, 9575 (2020).
Alter, G., Malenfant, J. M. & Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods. 294, 15–22 (2004).
Gómez-Lomelí, P. et al. Increase of IFN-gamma and TNF-alpha production in CD107a + NK-92 cells co-cultured with cervical cancer cell lines pre-treated with the HO-1 inhibitor. Cancer Cell. Int. 14, 100 (2014).
Smyth, M. J., Hayakawa, Y., Takeda, K. & Yagita, H. New aspects of natural-killer-cell surveillance and therapy of cancer. Nat. Rev. Cancer. 2, 850–861 (2002).
Okamoto, T. et al. Distinguishing primary from secondary mucinous ovarian tumors: an algorithm using the novel marker DPEP1. Mod. Pathol. 24, 267–276 (2011).
Lau, A. et al. Dipeptidase-1 governs renal inflammation during ischemia reperfusion injury. Sci. Adv. 8, eabm0142 (2022).
Liu, Q. et al. DPEP1 promotes the proliferation of colon cancer cells via the DPEP1/MYC feedback loop regulation. Biochem. Biophys. Res. Commun. 532, 520–527 (2020).
Shalom-Feuerstein, R., Cooks, T., Raz, A. & Kloog, Y. Galectin-3 regulates a molecular switch from N-Ras to K-Ras usage in human breast carcinoma cells. Cancer Res. 65, 7292–7300 (2005).
Ajani, J. A. et al. Galectin-3 expression is prognostic in diffuse type gastric adenocarcinoma, confers aggressive phenotype, and can be targeted by YAP1/BET inhibitors. Br. J. Cancer. 118, 52–61 (2018).
Zhang, L. et al. RN1, a novel galectin-3 inhibitor, inhibits pancreatic cancer cell growth in vitro and in vivo via blocking galectin-3 associated signaling pathways. Oncogene 36, 1297–1308 (2017).
Leary, S. et al. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition (American Veterinary Medical Association, 2020).
Funding
This work was supported by the National Research Foundation of Korea (NRF) and the National Research Council of Science and Technology (NST), funded by the Ministry of Science and ICT (MSIT) (Grant No. 2020R1A2C2010321), and by the KRIBB Research Initiative Program (Grant No. 1711196163).
Author information
Authors and Affiliations
Contributions
J.T.K., H.J.C., and H.G.L. conceived and designed the study. J.T.K., E.S.P., and Y.S.H. performed the experiments. J.T.K., H.R.Y., S.R.Y., and H.J.C. provided essential reagents or tools. J.T.K., E.S.P., H.J.C., and H.G.L. analyzed the data. J.T.K., E.S.P., H.J.C., and H.G.L. wrote the manuscript. All authors reviewed and critically revised the manuscript and approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
All animal procedures were approved by the KRIBB Institutional Animal Care and Use Committee under approval number KRIBB-AEC-24317. All methods were carried out in accordance with relevant guidelines and regulations, including the ARRIVE 2.0 (Animal Research: Reporting of In Vivo Experiments) guidelines.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kim, JT., Park, E.S., Hwang, Y.S. et al. Attenuation of natural killer cell cytotoxicity by interaction between NKp30 of NK cells and dipeptidase 1 of colon cancer cells. Sci Rep 15, 34777 (2025). https://doi.org/10.1038/s41598-025-18475-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-18475-z
