
Abstract
Riparian ecosystems are highly diverse and dynamic, but effects of fire on riparian vegetation are poorly understood. In 2020, widespread wildfires impacted forests across the western Oregon Cascades, including riparian areas. To investigate riparian plant community recovery, we quantified riparian vegetation responses to wildfire and forest management. We determined that vegetation response to burn severity varied by structural layer and was dynamic across the first three years post-fire. Overstory mortality after wildfire varied by species. In the understory, forb cover recovered rapidly; shrub cover and richness showed some recovery within three years. Indicator species highlighted compositional differences between sites that burned and those that did not. Although riparian zones are thought to be resilient to wildfire, our results demonstrate megafires can significantly alter them, resulting in extensive initial and delayed mortality, and dynamic regrowth. Globally, riparian zones are increasingly exposed to wildfire, and understanding factors influencing their recovery is critical.
Similar content being viewed by others

Introduction
Riparian areas are often characterized by high plant diversity and structural complexity but can be affected by forest management1. Although the width of riparian buffers and specific operating restrictions differ in managed forests across the globe2,3, the broader goal is often to insulate aquatic ecosystems from upslope disturbance. The efficacy of riparian buffers in protecting aquatic ecosystems from disturbance likely differs between anthropogenic and natural disturbances, and unique responses may occur when multiple disturbances occur in sequence. Natural disturbance (e.g., wind, fire, flood) can disrupt the regulating efficacy of riparian vegetation. However, riparian vegetation can also help regulate fire intensity or extent due to higher fuel moisture content and relative humidity4.ss
While ecosystem response after wildfire has been examined, the response of riparian areas to wildfire, specifically that of understory riparian vegetation, has not been well-studied. Understory vegetation in riparian areas may be less affected by wildfire than adjacent uplands, even with similar overstory fire severity5. In the western US, greater water availability in riparian areas often supports deciduous trees and shrubs with greater levels of stem and foliar moisture content, particularly in dry periods6, although larger streams tend to have more hardwoods than smaller streams that support more conifers5. Therefore, riparian areas may act as refugia for fire-sensitive species typically found in upland forests4. If most of the riparian buffer is wet with a deep soil organic layer, understory community composition may be minimally affected by fire compared to unburned stands7. Even if affected by wildfire, riparian plant species may be uniquely resilient to disturbances given their adaptations to seasonal flooding, seasonal drought, and wildfire1,8.
The speed and nature of riparian vegetation regrowth following disturbance are important because they affect recovery of stream ecosystem characteristics, such as stream temperature, bank stability, and sediment delivery. Without an intact overstory, riparian understory vegetation may serve a critical function in the post-fire recovery of the aquatic ecosystem by providing shade to the stream, intercepting precipitation and limiting overland runoff, and stabilizing banks with root development. In Mediterranean ecosystems, riparian zones can achieve rapid re-growth, particularly when riparian zones may have greater moisture content and nutrient availability9, or when disturbance-adapted species, with traits such as rapid sprouting or suckering, can quickly re-colonize10,11,12.
In September 2020, widespread forest fires affected a mosaic of federal and private managed forests in the western Oregon Cascades, USA. Five mega-fires burned approximately 3440 km2, with more than 80 percent of that area burning in the first 48 h of the wind-driven event13. Early in the event, these wildfires burned with high severity, affecting numerous forested riparian areas along headwaters streams4,14. To better understand how these wildfires affected aquatic ecosystem processes, we applied an established, large-scale replicated study design, used to quantify environmental responses to variation in watershed forest composition (see Coble, et al.15), to examine site and vegetation characteristics following mixed-severity wildfire within riparian management zones adjacent to small perennial streams. Our objectives were to determine whether riparian vegetation recovery after severe wildfires varied with burn severity, years since fire, pre-fire watershed age, or riparian stand structure. We explored these objectives for each layer of riparian vegetation, including (1) overstory (2) shrub and (3) forb layers using mortality, cover, and species richness as response variables. We hypothesized that burn severity would affect mortality and cover for all vegetation layers with delayed overstory mortality contributing to reduced canopy cover with time since fire, and rapid re-growth and establishment for shrub and forb layers contributing to increasing understory cover and species richness with time since fire.
Methods
Our study was designed to compare stream and riparian responses to wildfires in the western Oregon Cascades, USA15,16. This area has a maritime climate regime, with mild wet winters and warm, dry summers, soils that are moderately deep, well drained, and loamy. This results in highly productive tree species, like Pseudotsuga menziesii, and a long history of timber production by various forest industry landowners17. This ecoregion supports vegetation within the Tsuga heterophylla forest zone, which is dominated not only by Pseudotsuga menziesii, but also by Tsuga heterophylla and Thuja plicata. The successional trend for this region typically begins as forest reestablishment after fire and/or harvest, when weedy species dominate, and succeeds into a shrub-dominated community that supports hardwood species like Alnus rubra and Acer macrophyllum as the common conifer species develop17.
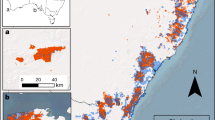
Vast stream networks and their riparian zones burned in the three fires included in our study (Riverside, Beachie Creek, and Holiday Farm), where 2044 km2 area burned. This included 4,020 km of stream length and their associated riparian areas; over 50% of which resulted in moderate soil burn severity and < 5% at high soil burn severity. For our study, we had 30 study sites; 25 sites from original project design also used by others (see Coble et al.15) For the original 25 sites, we used a stratified random sampling design to select 4th order watersheds within 6 km of the Beachie Creek, Riverside, and Holiday Farm fire boundaries (Fig. 1), with mean watershed pre-fire stand age (using data available at https://lemma.forestry.oregonstate.edu/data/structure-maps, based on 2017 data, which was the most recent pre-fire stand data available) and watershed fire extent (more details available in Coble, et al.15).

Study sites (watersheds denoted in black polygons) across western Oregon, USA Cascade Mountains, within and near the Riverside, Beachie Creek, and Holiday Farm fires of 2020. Map was created using ArcPro v.3.4.2.
We considered watershed fire extent as one of four categories: unburned (0%), minority (< 50%) of watershed burned, majority (> 50% and < 99%) of watershed burned, and complete (> 99%) watershed burned. Mean watershed pre-fire stand age was split into < 85y or > 85y, based on the distribution of mean watershed stand age from all 4th order watersheds within 6 km of these fires. From the available 4th-order watersheds, we randomly selected three streams for each of the eight categories, with one additional site selected for one category combination. For the vegetation sampling, we sampled an additional five streams within our larger study area, generally located further up in the stream network, including two 3rd order and three 2nd order streams, to capture more severely burned riparian areas and monitor their recovery over time. This was done because riparian responses were expected to differ with watershed size and geomorphology of hydrologic features that also control riparian species composition11. To estimate the length of stream and associated riparian areas burned in these three wildfires, we clipped the National Hydrography Dataset plus version 2 (NHDPlus V2) stream layer to the fire boundaries for Beachie Creek, Riverside, and Holiday Farm and summed total length of stream. We obtained soil burn severity (SBS) from Burned Area Emergency Response (BAER) database18, and evaluated riparian areas as within 30.48 m of stream. Data collected from these sites were combined with all other data, and analyzed together. Study sites were located on various forest landowners: 18 sites were on private large industrial landowners, 1 on small private, and 11 were on public lands (United States Forest Service and Bureau of Land Management).
Field sampling
We collected data on overstory, shrub, and forb layer during summers 2021–2023 (after 2020 wildfires) at 30 different subbasin watersheds stratified across gradients of fire severity and prior forest management intensity. Each study site was comprised of a 100 m stream reach located at the downstream end of a 4th order stream basin. We placed four transects at 20 m intervals on alternating sides of each stream, extending 50 m perpendicular to each stream. We relocated and resampled these transects each year using field marking (flagging and tree tags) and GPS coordinates. We collected data on overstory condition and species composition in 10 × 20 m (200 m2 plots), placed at 5–25 m along each transect, recording the diameter and species of every standing tree or snag (> 10 cm diameter) within each tree plot. Using a line-intercept method along each transect, we recorded cover of each shrub species19. For forb layer species, we placed 1 m2 quadrats every 5 m along each transect and recorded cover class of each species: 1 = 0–1%, 2 = 1–5%, 3 = 5–25%, 4 = 25–50%, 5 = 50–75%, 6 = 75–95%, or 7 = > 95% cover. We sampled canopy density at each quadrat using a spherical densiometer by calculating an average of 4 readings (one reading facing each cardinal direction) at each quadrat20.
As a measure of burn severity, we used a qualitative scale (0–3) to quantify evidence of fire/burn at each quadrat (e.g., 0 = no evidence of fire, 3 = the quadrat is entirely covered by charred organic matter or other signs of fire) during the first sample year. We calculated mean burn severity for each site by averaging the quadrat-level numeric assessments of burn severity.
To include a measure of riparian stand structure in our statistical models, we defined dominant canopy trees as those representing the largest 50 percent of total plot basal area (m2/ha), and calculated the mean diameter at breast height (DBH (cm)) of those dominant trees for each site.
We averaged canopy cover measurements for each vegetation quadrat and then summarized these to the site level for each sample year. We calculated overstory mortality annually as the proportional mortality (number of dead trees divided by number of both live and dead trees) by tree plot, and then calculated mean mortality across the four tree plots at each site. We summarized shrub cover as the total distance (m) along each transect covered by each unique species. We then calculated shrub richness as the count of unique species recorded per transect each sample year, and then calculated a mean across the four transects for each site. We summarized forb cover and richness by species each sample year at the quadrat level and then calculated means to the site level.
Statistical analyses
To assess whether burn severity influenced mortality and cover for all vegetation layers, we developed generalized linear mixed models with quadrat-level burn severity (hereafter referred to as burn severity), year sampled, watershed average stand age (as a categorical variable of < 85 or > 85 yrs old), average DBH of dominant riparian trees, and selected interactions as fixed effects (burn severity × year, burn severity × age, burn severity × DBH, and age × DBH), and Site or Transect within a Site as random effect(s). We specified a binomial or quasibinomial distribution for canopy cover, overstory mortality, shrub cover and forb cover. For shrub richness, we specified a Poisson distribution, and forb richness, we specified a Gamma distribution. For each model, we assessed model fit by examining the standardized residuals against the fitted values. We evaluated the significance of terms using an Analysis of Deviance and estimated least squared means using a significance level of 0.1.
To examine the association between species patterns and fire severity in the shrub and forb layers, we performed an indicator species analysis using the multipatt function in R, specifying the IndVal index and 999 permutations21. We subset the data by mean watershed stand age (Old or Young) and grouped by mean plot-level fire severity (None (0–0.05), Mid (0.5–1), or High (1–1.5). We calculated an Indicator Value (IV) for each significant species (ISA A × ISA B × 100) and considered any species with an IV > 25 to be an important indicator of a group, sensu Dufrêne and Legendre22.
Results
Riparian overstory cover was correlated with quadrat-level burn severity and exhibited some delay in overstory mortality. Canopy cover was negatively related to year since fire (p < 0.001) and burn severity × year since fire (p < 0.01), with canopy cover decreasing with increasing burn severity and decreasing each year since fire (Fig. 2A). At no/low and high fire severity, canopy cover was consistently very high or very low each year, but at mid-level burn severity, canopy cover greatly varied by year. Conversely, overstory mortality increased with increasing quadrat-level burn severity (p < 0.001), year since fire (p < 0.001), and by burn severity × year since fire (p = 0.01); mortality increased 2–3 years after fire compared to the first year, especially at sites with mid-level burn severity (Fig. 2B). We observed a greater decline in canopy cover through time for moderately burned sites (44%) than severely burned sites (25%). Moderately burned sites also exhibited greater magnitude shrub cover increase (3X) relative to most severely burned sites. In contrast, the most severely burned sites had the greatest magnitude of increases in forb cover. Burn severity and years since fire had heterogenous effects on overstory mortality across tree species: e.g., Thuja plicata had higher mortality immediately after fire, even in stands with lower burn severity, while Tsuga heterophylla and especially Alnus rubra showed higher survival the first year after fire but had high mortality two- and three-years post-fire (Fig. 3).

Canopy cover (A) and overstory mortality (B) by quadrat-level burn severity and year sampled. Quadrat-level burn severity was a qualitative scale of evidence of burn, calculated for each quadrat, and a mean was calculated for each site. Data lines were calculated from least squared means at fire severity intervals of 0.005, with shading denoting 90% confidence intervals.

Overstory mortality by quadrat-level burn severity and year. Quadrat-level burn severity was a qualitative scale of evidence of burn, calculated for each quadrat, and a mean was calculated for each site. Data lines were calculated from least squared means of proportional mortality (number of dead trees / total number of trees) at fire severity intervals of 0.005, with shading denoting 90% confidence intervals.
Ecosystem response to fire was more variable in the understory than overstory vegetation layers. Shrub cover was correlated with quadrat-level burn severity (p = 0.001) and the interaction of burn severity × year sampled (p < 0.001). In general, shrub cover decreased with increasing burn severity but showed some increases in cover by year three (Fig. 4A). Shrub species richness also decreased with increasing burn severity (p < 0.001) and burn severity × year since fire (p < 0.001) with similar increases in species richness three years after fire (Fig. 4B). Forb cover and species richness also varied by quadrat-level burn severity × year since fire (p < 0.001 for both). There appears to be some recovery in forb cover two and three years after fire (Fig. 5A), but the effect of year since fire for forb species richness is less clear (Fig. 5B).

Shrub cover (A) and richness (B) by quadrat-level burn severity and year sampled. Quadrat-level burn severity was a qualitative scale of evidence of burn, calculated for each quadrat, and a mean was calculated for each site. Data lines were calculated from least squared means at fire severity intervals of 0.005, with shading denoting 90% confidence intervals.

Forb cover (A) and richness (B) by quadrat-level burn severity and year sampled. Quadrat-level burn severity was a qualitative scale of evidence of burn, calculated for each quadrat, and a mean was calculated for each site. Data lines were calculated from least squared means at fire severity intervals of 0.005, with shading denoting 90% confidence intervals. Forb layer cover could exceed 100% because of multiple layers of vegetation.
For both shrub and forb species, indicators of wildfire were pioneer or early seral species, while species indicative of no wildfire tended to be mid- to late seral species (Table 1). In addition, most significant indicator species were native; the only significant exotic indicator species was a forb species indicative of high severity wildfire at older sites (Lactuca serriola).
Discussion
Changes in riparian cover across vegetation layers
Globally, megafires (> 40,500 ha) have been increasing, creating greater exposure for riparian ecosystems. These three megafires resulted in a range of fire severities at a patch scale that contributed to significant initial overstory mortality throughout entire watersheds, including riparian forests where canopy cover continued to decrease each year for the first three years after wildfire. While soil burn severity in adjacent riparian areas is often less burned than uplands5,23, we found riparian overstory mortality was extensive—exceeding 73% within three years post-fire in mostly burned and completely burned watersheds.
Although our related research found that initial overstory mortality in riparian areas was lower for Alnus rubra than Thuja plicata one year post-fire15, our analyses revealed overstory mortality was dynamic in the first three years post-fire, with evidence of both delayed mortality and rapid re-sprouting. Mortality of deciduous species was particularly dynamic with Alnus rubra experiencing delayed mortality and Acer macrophyllum, which can resprout after wildfire, initially showing delayed mortality from year one to two and then reviving again by year three. Species that re-sprout after apparent top kill can survive even high severity wildfire24 and, in our study, Acer macrophyllum apparent mortality reversed by over half in moderately burned sites between years two and three post-fire, due to epicormic sprouting in the canopy. Both Tsuga heterophylla and Alnus rubra experienced delayed mortality two and three years after wildfire, with the greatest change in delayed mortality observed in more moderately burned sites for both species, although Alnus rubra also experienced substantial delayed mortality in the most severely burned sites. Alnus rubra mortality increased from 66% in year one to 100% in years two and three in the most severely burned sites. At moderately burned sites, Alnus rubra mortality more than tripled from 23% in year one to 82% in year two and 89% in year three. In the most severely burned sites, Tsuga heterophylla already experienced high mortality (95%) in year one, which increased to 100% in years two and three. In moderately burned sites, Tsuga heterophylla mortality increased 30% from 56% in year one to 88% in years two and three. Previous research also found that Tsuga heterophylla had high mortality after fire compared to other common species, like Pseudotsuga menziesii, partly due to differences in tree diameter, as Pseudotsuga menziesii tended to be larger than Tsuga heterophylla25.
The magnitude of delayed mortality observed with these riparian species exceeded estimated delayed mortality in the literature, likely because much of that literature focuses on conifer mortality and reflects a lack of information in riparian areas where initial survival may be greater for some riparian species. For example, using a remote sensing analysis across the western U.S., Busby, et al.26 estimated delayed mortality of conifers reached 5–25% at the scale of a fire perimeter five years after wildfire. This is similar to the 30% increase in mortality in moderately burned sites of the only conifer (Tsuga heterophylla) that experienced delayed mortality in our study. Brown et al.25 found that 44% of the trees alive initially after fire had died 6–7 years later. Others found that most mortality occurred within two years of fire, although only conifers were considered in their study27. Other studies of riparian areas found that 94% of Alnus rubra and Salix spp. (willow) trees were killed within three years by a wildfire in a Chapparral landscape in southern California, USA28 and canopy cover was reduced 48–63% post-fire compared to pre-fire29.
Pre-fire drought can increase tree mortality susceptibility to wildfire30,31 and post-fire drought may compound impacts24,32. Water years 2019–2021 (Oct 1 to Sep 30) were drought years for northwest Oregon (includes Riverside and Beachie Creek fires) and 2018, 2020, and 2021 in southwest Oregon (includes Holiday Farm fire) based on the SPI calculated from PRISM data33. Thus, drought conditions before, during, and after these wildfires may have contributed to initial and delayed fire-induced mortality. In the first summer following the fires (2021), there was an early summer period of extreme temperatures caused by an anomalous heat-dome, and periods of extreme heat have been shown to be more impactful for delayed tree mortality than cooler drought periods34,35. Fire-induced injuries that lead to cambium necrosis and limit hydraulic conductivity of the xylem also contribute to delayed mortality24,36, likely limiting ability to survive future stressful conditions in addition to fire, such as flooding, winds, ice storms, and forest pests. Forest structure, including tree density, may influence mortality after fire, particularly as it affects fuels, moisture content of vegetation, and interacts with fire behavior24. Pre-fire watershed stand age also encompasses differences in forest ownership and management practices that lead to a divergence in forest stand structure across our study sites. However, we did not find overstory mortality response to differ by pre-fire stand age; rather, only fire severity explained mortality, indicating riparian tree mortality in these fires was similar regardless of pre-fire stand conditions.
Riparian vegetation response to wildfire was dynamic across vegetation layers: forb cover showed some recovery by the second year after fire across all burn severities, and forb richness temporarily increased during the first two years following fire; shrub recovery was delayed to the third-year post-fire when both shrub cover and richness increased three years after wildfire across all burn severities, with substantial increases in richness in more severely burned sites. Simultaneous with understory recovery was the delayed mortality of Alnus rubra and Tsuga heterophylla [primarily two years after wildfire], and continued reductions in canopy cover each year, which may have contributed to the timing and success of forb and shrub recovery by opening the canopy. Similar to our findings, previous research also showed increases in species richness, which they attributed to both persistence of existing species and invasion of new species after disturbances like logging and burning37,38. The delay in shrub cover recovery we found is similar to previous research that also showed slower responses in the shrub layer to disturbance37.
Severity of fire, and time since fire, are important factors that drive complex responses in various layers of riparian vegetation. Therefore, continued monitoring of recovery after wildfire in these riparian areas is critical not only to understand terrestrial ecosystems, but also for aquatic ecosystems that rely on vegetation for shade, organic matter and nutrient inputs, and bank stability. Coble, et al.15, and Swartz, et al.16 show that simultaneous monitoring of canopy cover from measurements taken in these streams revealed similar declines in overstory canopy cover each year since the wildfire, with high overstory mortality contributing to higher stream temperature maxima. In-stream monitoring indicated that understory re-growth, while extensive, did not effectively shade or limit thermal increases in the first three years post-fire15,16. Despite these elevated summer stream temperatures, fish biomass and densities were elevated in exposed and severely burned watersheds, presumably due to greater basal resource availability, while amphibian biomass and densities were unaffected16. Dead standing trees, their root structures, and roots of understory shrubs and forbs, may provide important functions for stream and riparian ecosystems such as bank stability and microhabitats28. Fallen overstory trees can also contribute to riparian function as coarse wood on forest floor or as large wood in streams15,23.
Indicator species
Studies from other land uses have revealed anthropogenically disturbed riparian zones may represent unique ecosystems, hosting different species and combinations of species than less disturbed systems39. Although we are unaware of prior documentation of this effect in forests, we expected forest management-related disturbances either pre-fire or post-fire (including a greater proportion of recently harvested stands, salvage, higher road density and stream crossings) to have been of greater magnitude or frequency in the watersheds with younger pre-fire stand age. In our study, older mean watershed stand ages generally indicated less anthropogenic disturbance and often included a component of old-growth. Importantly, legacy effects of prior historical harvest activities likely affected most/all sites (i.e., splash dams and log drives were once prevalent throughout much of western Oregon streams)40. Riparian zones historically impacted by anthropogenic disturbance may be more likely to have invasive plant species in their seed banks41,42. Examining differences in the vegetation community composition after wildfire allows us to explore whether novel species combinations occur in riparian systems that had more frequent or greater magnitude anthropogenic disturbance before also being confronted with wildfire.
We expected to find exotic indicator species in younger sites, but only documented Lactuca serriola, an exotic forb species indicative of wildfire in older sites. Similar research also found Lactuca serriola after disturbance, and classified this species as an invader (rather than residual) with low cover37. Anthropogenic disturbance gradients in forested riparian areas did not contribute to novel riparian communities (i.e., there were no invasives in more disturbed sites), but did have unique native species indicators. We found only two indicator species shared by both young and old unburned riparian areas (Acer circinatum and Tsuga heterophylla). In burned sites, Rubus spectabilis was an indicator of both young and old sites, and no other indicator species were found in young sites. This indicates that while a unique combination of mid- to late-seral species present in younger unburned sites can be used to identify riparian areas draining younger stands, in our study, only presence of Rubus leucodermis may differentiate an older burned site from younger burned sites.
Only two species of shrubs were indicators of wildfire: Rubus leucodermis in younger mean watershed stand ages, and Rubus spectabilis in both younger and older stand age watersheds. In a prior study of clearcut followed by broadcast burning in northern Idaho, USA, Rubus leucodermis occurred more frequently in less severely burned sites than high burn severity sites and also varied by time since wildfire, maintaining cover in the first five years and then reduced 15 years post-fire43. Other research also found Rubus leucodermis invaded after disturbance, peaking after 5–10 years, but maintained low cover37. Both of these native shrub species are rhizomatous, and may have established quickly post-fire via dormant seeds44 or sprouted from belowground perennating roots.
Implications & conclusions
Our findings indicate understory vegetation-initiated recovery within three years after wildfire. Although individual species may re-establish quickly after wildfire, recovery of vegetation structure and composition takes more time45. In the absence of overstory canopy, understory vegetation layers provided little shade to streams even after three years post-fire, but this will likely change in coming years. Early seral or pioneer species were indicative of wildfire effects which align with previous research showing disturbance-associated species to be more common or abundant after wildfire46. Other studies have shown wildfire in riparian areas reduced native species richness, although the responses were variable depending on the season of burning47, but our data do not reflect that pattern during the first three years post-fire. Longer term study is essential to fully understand recovery of vegetation in these riparian forest stands.
Our results demonstrate that megafires can significantly alter riparian zones, even in relatively large 4th order streams. While the three wildfires included in our study were extensive, these three wildfires represent only a fraction of area burned in Oregon’s 2020 wildfires, which encompassed 4900 km2 across the state48,49. Furthermore, the three megafires in our study burned > 2000X the stream length of what had burned over a 30-year period (1984–2014) in this region50. Similarly, megafires in Australia burned 126,000 km2, in a brief 7 month period in 2019–2020, affecting 81,304 km of rivers and their riparian areas, and 29% of that river’s riparian network burned at extreme or high burn severity39. Although riparian areas comprise a small proportion of an area burned, the extent of riparian vegetation burned, and therefore the wildfire return intervals of the riparian areas themselves, are extremely varied between different ecoregions51. Globally, riparian zones are increasingly exposed to wildfire across broad scales, and understanding their recovery is important for water quality, nutrient cycling, and providing habitat structure for in-stream and riparian associated wildlife50.
Data availability
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
References
Stella, J. C., Rodríguez-González, P. M., Dufour, S. & Bendix, J. Riparian vegetation research in Mediterranean-climate regions: Common patterns, ecological processes, and considerations for management. Hydrobiologia 719, 291–315 (2013).
Boisjolie, B. A., Santelmann, M. V., Flitcroft, R. L. & Duncan, S. L. Legal ecotones: a comparative analysis of riparian policy protection in the Oregon Coast Range, USA. J. Environ. Manage. 197, 206–220 (2017).
Lee, P., Smyth, C. & Boutin, S. Quantitative review of riparian buffer width guidelines from Canada and the United States. J. Environ. Manage. 70, 165–180 (2004).
Riis, T. et al. Global overview of ecosystem services provided by riparian vegetation. Bioscience 70, 501–514 (2020).
Halofsky, J. E. & Hibbs, D. E. Determinants of riparian fire severity in two Oregon fires, USA. Can. J. For. Res. 38, 1959–1973 (2008).
Dwire, K. A. & Kauffman, J. B. Fire and riparian ecosystems in landscapes of the western USA. For. Ecol. Manage. 178, 61–74 (2003).
Day, N. J. et al. Fire characteristics and environmental conditions shape plant communities via regeneration strategy. Ecography 43, 1464–1474 (2020).
D’Souza, L. E., Reiter, M., Six, L. J. & Bilby, R. E. Response of vegetation, shade and stream temperature to debris torrents in two western Oregon watersheds. For. Ecol. Manage. 261, 2157–2167 (2011).
Verkaik, I. et al. Fire as a disturbance in Mediterranean climate streams. Hydrobiologia 719, 353–382 (2013).
Naiman, R. J. & Decamps, H. The ecology of interfaces: riparian zones. Annu. Rev. Ecol. Syst. 28, 621–658 (1997).
Kobziar, L. N. & McBride, J. R. Wildfire burn patterns and riparian vegetation response along two northern Sierra Nevada streams. For. Ecol. Manage. 222, 254–265 (2006).
Keeley, J. E. & Zedler, P. H. Reproduction of chaparral shrubs after fire: A comparison of sprouting and seeding strategies. Am. Midl. Nat. 99, 142–161 (1978).
NCASI, I. The Labor Day Fires of 2020. (National Council for Air and Stream Improvement (NCASI), Rsearch Triangle Park, NC, 2021).
Reilly, M. J. et al. Cascadia burning: The historic, but not historically unprecedented, 2020 wildfires in the Pacific Northwest, USA. Ecosphere 13, e4070 (2022).
Coble, A. A., Penaluna, B. E., Six, L. J. & Verschuyl, J. Fire severity influences large wood and stream ecosystem responses in western Oregon watersheds. Fire Ecol. 19, 34. https://doi.org/10.1186/s42408-023-00192-5 (2023).
Swartz, A. et al. Withstanding megafires: fishes thrive and amphibians persist in severly burned watersheds. Communications Earth & Environment (In review).
Franklin, J. F. & Dyrness, C. T. Natural vegetation of Oregon and Washington. 452 (Oregon State University Press, 1988).
MTBS Project: MTBS Data Access: Fire Level Geospatial Data, <http://mtbs.gov/direct-download> (2021 (last accessed December 2021)).
Canfield, R. H. Application of the line interception method in sampling range vegetation. J. For. 39, 388–394 (1941).
Strickler, G. S. Use of the densiometer to estimate density of forest canopy on permanent sample plots. PNW Old Series Research Notes No. 180, p. 1–5 180 (1959).
Bakker, J. D. Increasing the utility of indicator species analysis. J. Appl. Ecol. 45, 1829–1835 (2008).
Dufrêne, M. & Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–366 (1997).
Pettit, N. E. & Naiman, R. J. Fire in the riparian zone: Characteristics and ecological consequences. Ecosystems 10, 673–687 (2007).
Hood, S. M., Varner, J. M., Van Mantgem, P. & Cansler, C. A. Fire and tree death: Understanding and improving modeling of fire-induced tree mortality. Environ. Res. Lett. 13, 113004 (2018).
Brown, M. J., Kertis, J., Huff, M. H. Natural tree regeneration and coarse woody debris dynamics after a forest fire in the western Cascade Range. Res. Pap. PNW-RP-592. Portland, OR: US Department of Agriculture, Forest Service, Pacific Northwest Research Station. 50 p. 592 (2013).
Busby, S., Evers, C. & Holz, A. Patterns, drivers, and implications of postfire delayed tree mortality in temperate conifer forests of the western United States. Ecosphere 15, e4805 (2024).
Hood, S. M., Smith, S. L. & Cluck, D. R. Predicting mortality for five California conifers following wildfire. For. Ecol. Manage. 260, 750–762 (2010).
Bendix, J. & Cowell, C. M. Fire, floods and woody debris: Interactions between biotic and geomorphic processes. Geomorphology 116, 297–304 (2010).
Rehn, A. C. The effects of wildfire on benthic macroinvertebrates in southern California. (San Diego, CA, 2010).
van Mantgem, P. J. et al. Climatic stress increases forest fire severity across the western United States. Ecol. Lett. 16, 1151–1156 (2013).
Van Mantgem, P. J., Falk, D. A., Williams, E. C., Das, A. J. & Stephenson, N. L. Pre-fire drought and competition mediate post-fire conifer mortality in western US National Parks. Ecol. Appl. 28, 1730–1739 (2018).
Furniss, T. J., Das, A. J., van Mantgem, P. J., Stephenson, N. L. & Lutz, J. A. Crowding, climate, and the case for social distancing among trees. Ecol. Appl. 32, e2507 (2022).
O’Neill, L. W., Koszuta, M., Siler, N. & Fleishman, E. Historical and projected future drought in Oregon (Oregon State University, 2023).
Still, C. J. et al. Causes of widespread foliar damage from the June 2021 Pacific Northwest Heat Dome: More heat than drought. Tree Physiol. 43, 203–209 (2023).
Cansler, C. A. et al. Drought before fire increases tree mortality after fire. Ecosphere 15, e70083 (2024).
Bär, A., Michaletz, S. T. & Mayr, S. Fire effects on tree physiology. New Phytol. 223, 1728–1741 (2019).
Halpern, C. B. Early successional patterns of forest species: Interactions of life history traits and disturbance. Ecology 70, 704–720. https://doi.org/10.2307/1940221 (1989).
Halpern, C. B. et al. Response of forest vegetation to varying levels and patterns of green-tree retention: An overview of a long-term experiment. Northwest Sci. 73, 27–44 (1999).
Fryirs, K., Zhang, N., Duxbury, E. & Ralph, T. Rivers up in smoke: Impacts of Australia’s 2019–2020 megafires on riparian systems. Int. J. Wildland Fire 31, 720–727 (2022).
Steel, Z. L., Safford, H. D. & Viers, J. H. The fire frequency-severity relationship and the legacy of fire suppression in California forests. Ecosphere 6, 1–23 (2015).
Moffatt, S. F. & Mclachlan, S. M. Effects of land use disturbance on seed banks of riparian forests in southern Manitoba. Ecoscience 10, 361–369 (2003).
O’Donnell, J., Fryirs, K. A. & Leishman, M. R. Seed banks as a source of vegetation regeneration to support the recovery of degraded rivers: A comparison of river reaches of varying condition. Sci. Total Environ. 542, 591–602 (2016).
Morgan, P. & Neuenschwander, L. F. Shrub response to high and low severity burns following clearcutting in northern Idaho. West. J. Appl. For. 3, 5–9 (1988).
Morgan, P. & Neuenschwander, L. Seed-bank contributions to regeneration of shrub species after clear-cutting and burning. Can. J. Bot. 66, 169–172 (1988).
DeBano, L. F. & Neary, D. G. Effects of fire on riparian systems. United States Department of Agriculture Forest Service General Technical Report RM, 69–76 (1996).
Donato, D. C., Fontaine, J. B., Robinson, W. D., Kauffman, J. B. & Law, B. E. Vegetation response to a short interval between high-severity wildfires in a mixed-evergreen forest. J. Ecol. 97, 142–154 (2009).
Hankins, D. L. The effects of indigenous prescribed fire on riparian vegetation in central California. Ecol. Process. 2, 1–9 (2013).
Mass, C. F., Ovens, D., Conrick, R. & Saltenberger, J. The september 2020 wildfires over the Pacific Northwest. Weather Forecast. 36, 1843–1865 (2021).
Council, O. W. E. R. Recovering & Rebuilding from Oregon’s 2020 Wildfires. Governor, O., Ed, 32 (2021).
Ball, G., Regier, P., González-Pinzón, R., Reale, J. & Van Horn, D. Wildfires increasingly impact western US fluvial networks. Nat. Commun. 12, 2484. https://doi.org/10.1038/s41467-021-22747-3 (2021).
Bendix, J. & Commons, M. G. Distribution and frequency of wildfire in California riparian ecosystems. Environ. Res. Lett. 12, 075008 (2017).
USDA, N. The PLANTS database http://plants.usda.gov (2017).
Cook, J. E. & Halpern, C. B. Vegetation changes in blown-down and scorched forests 10–26 years after the eruption of Mount St. Helens, Washington, USA. Plant Ecol. 219, 957–972 (2018).
Priebe, J. E. Silvicultural treatment impacts on understory trees and 20-year understory vegetation dynamics in mature Douglas-fir forests. (2016).
Hitchcock, C. L., Cronquist, A., Ownbey, M. & Thompson, J. Vascular plants of the pacific northwest (University of Washington Press, 1969).
Acknowledgements
We thank M. Barnett, R. Boucher, A. Clements, E. Cruz, A. Hammond, A Lamet, B. Hobscheid, G. Demorest, A. Owens, M. Thomas, and especially G. Broyles and L. Koerner for assisting with field data collection. Bureau of Land Management, Campbell Global, LLC, Fruit Growers Supply, Giustina Resources, Port Blakely, Rayonier, State of Oregon, United States Forest Service, and Weyerhaeuser kindly allowed land access and provided assistance. Statistical support was provided by J. Thornton-Frost, J. Jones, and M. Fix. J. Homyack and D. Miller provided revisions to an earlier version of this manuscript. This research was funded by the National Council for Air and Stream Improvement, Inc., and Weyerhaeuser.
Funding
Weyerhaeuser Company, National Council for Air and Stream Improvement.
Author information
Authors and Affiliations
Contributions
All authors (L.S., J.V., and A.C.) contributed to writing the manuscript, while L.S. completed the analyses and most figures/tables, except for Fig. 1, which A.C. contributed. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Six, L.J., Verschuyl, J. & Coble, A.A. Riparian vegetation layers differ three years after mixed severity wildfire in Oregon. Sci Rep 15, 36247 (2025). https://doi.org/10.1038/s41598-025-20242-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-20242-z
