
Abstract
Climate change is shifting optimal habitats for medicinal plants, potentially compromising the efficacy and therapeutic value of herbal remedies. Global warming and increased extreme weather events threaten the sustainability and pharmaceutical integrity of Angelica sinensis (Oliv.) Diels (A. sinensis). Despite its importance in traditional herbal medicine, there is limited research on adaptation of A. sinensis to climate challenges. This study systematically collected occurrence data of A. sinensis through field expeditions and online databases, using the Maxent ecological niche modeling tool and ArcGIS software to forecast suitable habitats. A total of 402 species occurrence points and 21 environmental variables were selected for modeling, resulting in 1,160 distribution models, of which only one met the stringent 5% odds ratio (OR) standard. The optimal model exhibited a pROC value of 0, an OR of 0.0196, and an AICc score of 9,287.133. The model, run ten times for robustness, showed an average AUC of 0.980, indicating high accuracy and reliability. Under current climate conditions, suitable habitats for A. sinensis cover approximately 13% of mainland China, primarily in Gansu (73.77%), Qinghai (14.73%), and Sichuan (11.18%) provinces. Environmental factors such as altitude, humidity, and temperature significantly influence the geographical distribution of A. sinensis. The future climate scenario predictions suggest that suitable habitats will generally shift towards higher latitudes, with areas of moderate to high suitability primarily distributed across the provinces of Gansu and Qinghai. The interactions between environmental factors, characterized by mutual and nonlinear enhancement, further influence the spatial differentiation of suitability zones. Overlay analysis with 2020 land cover data indicated that 861,437 km² of arable and forest land are suitable for A. sinensis cultivation. Future predictions under four SSP scenarios show varying changes in suitable habitat areas, with the most significant expansion under SSP370 between 2080 and 2100, covering 14.54% of mainland China. These findings provide critical insights for optimizing A. sinensis cultivation regions and quality assessments in response to climate change.
Similar content being viewed by others
Introduction
Climate change is widely regarded as one of the primary threats to global biodiversity in the 21st century1. According to the Third Global Biodiversity Report2, nearly one-quarter of all plant species are currently at risk of extinction, with the likelihood of species loss escalating sharply. Concurrently, the Fifth Assessment Report by the Intergovernmental Panel on Climate Change (IPCC) projects a continued trend of global warming3, with average global temperatures expected to rise between 0.3 °C and 4.8 °C by the end of this century. As a fundamental driver of species habitat distribution, shifts in climatic conditions will inevitably reshape the geographic distribution and ecological characteristics of species. With the ongoing evolution of global climate patterns, the spatial distribution of species is expected to undergo significant changes4. Consequently, investigating and accurately predicting the impacts of climate change on potential species distribution patterns is of paramount importance for understanding biodiversity dynamics and informing conservation strategies.
Species distribution models (SDMs) play a pivotal role in ecological and biogeographical research, acting as a bridge that tightly links specific species to their ecological niche factors. Among the various SDMs developed to predict species distribution patterns, including MaxEnt, GARP, BIOCLIM, and Climex, the MaxEnt model stands out for its outstanding performance. It has become one of the most widely favored tools for modeling species distribution and environmental niches. Notably, MaxEnt demonstrates superior predictive accuracy compared to other models when dealing with the challenges of sparse or incomplete species distribution data5. Consequently, it has been extensively adopted to forecast the long-term impacts of ecological niche factors, particularly climate change, on future habitat suitability. In recent years, the application of the MaxEnt model has continued to expand across key domains of plant biodiversity6. These include the scientific development of conservation strategies for endangered species, precise risk assessments of biological invasions, and the rational planning of crop cultivation areas. These advancements underscore critical role of MaxEnt in driving ecological conservation and promoting sustainable development.
Angelica sinensis (Oliv.) Diels (A. sinensis) is a perennial herb belonging to the family Umbelliferae. It has held a pivotal role in traditional Chinese medicine (TCM) for thousands of years7. Its dried root, formally recognized as Angelica sinensis Radix in pharmacology, has been included in the Chinese Pharmacopoeia as a core medicinal ingredient. Modern pharmacological studies have comprehensively revealed the diverse therapeutic potential of A. sinensis, including cardioprotection, immune enhancement, anti-tumor properties, radiation protection, and anti-arrhythmic effects8. In addition, due to its unique health benefits, A. sinensis has gained increasing popularity as a functional food ingredient. Statistics indicate that over six thousand traditional Chinese medicine prescriptions and one hundred and fifty-one health products incorporate A. sinensis as a primary ingredient9.This plant is primarily distributed in Gansu, Sichuan, Yunnan, and Hubei provinces in China, with the “Mingui” variety from Min County, Gansu Province, particularly renowned for its strong aroma, rich oil content, high yield, and superior quality10. Despite the remarkable application potential of A. sinensis in health and wellness, current research predominantly focuses on its chemical composition11,12,13 and pharmacological activities14,15,16, leaving significant gaps in understanding its ecological requirements and ideal habitats. Given the dual challenges posed by climate change and increasing market demand, addressing these knowledge gaps is essential for ensuring the sustainable production of high-quality A. sinensis. We hypothesize that with global warming, certain regions may no longer be suitable for the growth of A. sinensis due to excessively high temperatures, while some high-altitude or cooler areas may become new suitable habitats.
Herein, based on the distribution data of Angelica sinensis, this study utilized the MaxEnt ecological niche model and the ArcGIS software platform to predict the potential distribution areas, spatial distribution patterns, and climate-adapted habitats of A. sinensis under current and future climatic conditions in China. The specific research objectives were as follows: (1) What is the potential distribution of A. sinensis in China under current and future climate scenarios? (2) How will climate change affect the future suitable distribution and spatial patterns of A. sinensis? (3) How do environmental factors influence the spatial distribution of A. sinensis habitats? Addressing these questions will provide a theoretical basis for the strategic planning of A. sinensis introduction and cultivation. Furthermore, the findings will offer scientific evidence and practical guidance for the conservation of medicinal plant resources and the establishment of nature reserves. This research ultimately aims to promote the sustainable development and conservation of this vital medicinal plant, ensuring its ecological and economic value is preserved amidst the challenges posed by climate change.
Materials and methods
Species occurrence record collection and processing
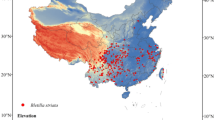
The occurrence records of A. sinensis in China were obtained through a combination of online database searches and field investigations. Specifically, the online search utilized the Global Biodiversity Information Facility (GBIF, https://www.gbif.org/) database, yielding a total of 180 records. Given that Gansu Province is the primary supplier of A. sinensis in China, contributing approximately 90% of the domestic market, extensive field surveys were conducted in this region. These surveys resulted in the collection of 611 distribution records. To prevent model overfitting, distribution data were filtered using the ENMTools software (http://enmtools.blogspot.com/), ensuring that only one record was retained for each 5 × 5 km grid. After filtering, 402 distribution points were selected for model construction (Fig. 1). The maps were generated following the standard mapping framework of the Ministry of Natural Resources of the People’s Republic of China (GS[2020]4619), with all boundary delineations remaining unchanged.

Occurrence records of A. sinensis in China.
Selection and processing of environmental variables
The ecological niche of a species is influenced by a variety of factors, including climate, topography, and biotic interactions. To comprehensively characterize the ecological niche of A. sinensis, we selected 104 environmental variables that reflect its climatic and topographic preferences. These include: 19 bioclimatic variables, 12 Precipitation Variables, 36 Temperature Variables (tmax12,tmin12,tavg12), 12 Solar Radiation, 12 Wind Speed, 12 Vapor Pressure, and Elevation. The biological and ecological relevance of these variables was determined based on the known habitat preferences and physiological constraints of A. sinensis, as supported by field observations. Solar radiation was included because it directly influences photosynthesis and the growth of medicinal plants, while wind speed was selected due to its impact on plant morphology, water retention, and the overall physiological stress that can affect plant growth and medicinal compound production. Since wind speed and solar radiation data are not available for future time periods, we assumed that these variables would remain constant in future climate scenarios.
The bioclimatic data for current conditions (1970–2000), early-century (2021–2040), mid-century (2041–2060), mid-late-century (2061–2080) and late-century (2081–2100) were obtained from the WorldClim database (http://www.worldclim.org) at a resolution of 2.5 arc minutes (~ 5 km). Future projections were derived from the Coupled Model Intercomparison Project Phase 6 (CMIP6) under four Shared Socioeconomic Pathways (SSPs): SSP1-2.6 (low radiative forcing), SSP2-4.5 (medium radiative forcing), SSP3-7.0 (medium-high radiative forcing), and SSP5-8.5 (high radiative forcing). Future climate data are from simulations. Recognizing the uncertainties associated with climate change projections, our initial study utilized the BCC-CSM2-MR global climate model (developed by the National Climate Center, Beijing) due to its strong performance in China-specific climatic predictions17. However, to enhance the robustness of our analysis, we expanded the models used for future predictions to include: EC-Earth3-Veg (Europe), MPI-ESM1-2-HR (Germany), and CMCC-ESM2 (CItaly). The final predictions for A. sinensis were based on the average results of these four models, with additional maps of estimation uncertainty18.
Multicollinearity among environmental variables, which can lead to model overfitting and detrimentally affect model output, prompted a correlation analysis of the environmental variables at the species distribution points using SPSS software (version 25.0). Environmental variables exceeding an absolute correlation coefficient value of 0.9 were scrutinized, and those making a significant contribution based on the initial calculation results of the MaxEnt model were retained, systematically excluding highly correlated variables. Ultimately, 21 variables were selected (Table 1).
MaxEnt model construction, optimization and evaluation
MaxEnt 3.4.1 was employed to predict the suitable habitat distribution of A. sinensis. The 75% occurrence points were randomly selected for model training, while the remaining 25% were utilized for validation19. Tenfold cross-validation, a method commonly employed in previous studies, was utilized to ensure the accuracy of the model. The robustness of predictions of the MaxEnt model is significantly impacted by the configuration of the feature class (FC) and regularization multiplier (RM), and the default parameter settings are often found to be suboptimal. Therefore, R 3.6.3 (https://www.r-project.org/) and the Kuenm package (https://github.com/marlonecobos/kuenm) were employed to optimize the MaxEnt model. Candidate models were evaluated by the Kuenm package through the calculation of Partial ROC (pROC), omission rate (OR), and AICc values, considering the statistical significance of the model, predictive power, and complexity. The optimal model parameters, based on statistical significance (pROC = 0), omission rate below the threshold (OR < 5%), and the smallest AICc value, were selected for the modeling analysis.
Redefinition of the current potential distribution areas
The habitat suitability for A. sinensis was redefined and categorized using the reclassification tool in ArcMap 10.8, based on model predictions. The suitability index was divided into four classes to reflect varying levels of suitability20: Unsuitable: Areas with a suitability index of 0 to 0.2; Poorly Suitable: Areas with a suitability index of 0.2 to 0.4; Moderately Suitable: Areas with a suitability index of 0.4 to 0.6; Highly Suitable: Areas with a suitability index of 0.6 to 1. In parallel, the land-use patterns were analyzed within high, medium, and low suitability zones to provide insights into potential cultivation strategies. Using the 2020 Global Land Cover Dataset with a 30 m resolution (GLC_FCS30-2020), provided by the Aerospace Information Research Institute of the Chinese Academy of Sciences, the distribution was mapped and proportion of cropland and forest in each suitability category.
Factors affecting the variability in the spatial distribution of potential suitable zones
The Geographical Detector is an innovative spatial data analytics tool, comprising a suite of statistical techniques designed to uncover spatial disparities and the underlying forces that drive them. This method is adeptly utilized to identify and quantify the influence and mechanisms of spatial factors on specific events or phenomena21. By amalgamating GIS, statistical analysis, and mathematical modeling, the Geographical Detector swiftly pinpoints key factors and assesses their interplay. The tool is structured around four specialized modules: the Ecological Detector, the Risk Detector, the Interaction Detector, and the Divergence and Factor Detector, each contributing to a robust framework for spatial data analysis. In this research, factor and interaction detectors were utilized to examine the spatial distribution of A. sinensis habitats. The Interaction Detector was employed to understand how environmental factors interact to influence habitat differentiation. Meanwhile, the Factor Detector assessed the spatial heterogeneity of potential habitats and quantified the impact of various factors on this spatial variation, as indicated by q-values. The q-values were expressed as:
The explanatory power of environmental factors on the spatial distribution of ecological niches is measured by q, which is scaled from 0 to 1, with higher values denoting a more significant influence. Values from 1 to L are taken by h, representing the stratification of the explanatory variable Y or factor X. The number of grid cells in category h and the number of grid cells in the whole region are represented by Nh and N, respectively. Additionally, the total variance and the variance of stratum h are represented by σ². The sum of the within-stratum variance and the total regional spatial variance are represented by WSS and TSS, respectively.
Results
Model optimization and accuracy evaluation
After screening, a total of 402 species occurrence points and 21 environmental variables were meticulously curated for the purpose of modelling. Employing the Kuenm package, a comprehensive suite of 1160 distribution models was generated, from which only one model conformed to the stringent 5% odds ratio (OR) standard. The optimal model exhibited a pROC value of 0, an OR of 0.0196, and an AICc score of 9287.133. The optimised model parameters were RM = 0.2 and FC = LQP (Fig. 2(A)). The distribution data of A. sinensis, along with the selected environmental factors, were imported into the MaxEnt software, which utilises the principles of maximum entropy to model species distributions. The model was run using filtered RM and FC parameters. This procedure was repeated 10 times to ensure the robustness of the model. The results show an average Area Under the Receiver Operating Characteristic Curve (AUC) of 0.980 with a standard deviation of 0.003 (Fig. 2(B)). These values indicate that the model has high accuracy and reliability, suggesting that it can be effectively used to predict the suitable habitat distribution of the species.

(A) Omission rate of candidate models and the natural logarithm of AICc values, (B) ROC curve of the predictive model.
Predicting distribution under current conditions
The simulated distribution of suitable habitats for A. sinensis under the current climate model is shown in Fig. 3(A). The logistic output from the MaxEnt model is expressed as a probability, with values ranging from 0 to 1 22. Employing the reclassification tool within ArcMap 10.8, the simulation outcomes were stratified into four distinct categories. The results show that the total area of suitable habitats for A. sinensis is approximately 1.25 × 106 km2, accounting for approximately 13% of mainland China’s area, distributed across 12 provinces. The most suitable regions were primarily located in the Gansu (73.77%), Qinghai (14.73%), and Sichuan (11.18%) provinces. Moderately suitable areas were mainly Gansu (33.24%), Sichuan (25.42%), Shaanxi (13.79%), Qinghai (10.03%), and Yunnan (5.77%). Overall, the areas of high, moderate, and low suitability are 1.66 × 104 km2, 2.87 × 105 km2, and 9.48 × 105 km2, respectively, accounting for 0.17%, 2.99%, and 9.88% of the area of mainland China.
An overlay analysis was carried out using the 2020 land cover data with the suitable distribution areas for A. sinensis under current climatic conditions to further test and obtain the actual possible distribution range of A. sinensis under current climatic conditions (Fig. 3(B)). The land cover type of A. sinensis is cultivated land or forest land, so only areas with a forest land cover type or cultivated land cover type have the potential for A. sinensis distribution (Figure S1(A) and (B)). The analysis indicates that the arable land area suitable for A. sinensis cultivation spans 319,596 km2, while the forest land area extends to 541,841 km2, totaling 861,437 km2. However, the current predicted potential distribution area amounts to 1,251,600 km2, suggesting that there is an area of 390,163 km2 that consists of other land cover types, which are not conducive to the cultivation of A. sinensis.

(A) Map for potential current habitat suitability and (B) the actual possible distribution of A. sinensis in China.
Predicted future potential distribution and suitable habitat changes
To minimize the uncertainty inherent in future climate predictions, data sets generated by four GCMs from different countries were used for integrated analysis under each SSP. The selected models include: EC-Earth3-Veg from Europe, MPI-ESM1-2-HR from Germany, CMCC-ESM2 from CItaly, and BCC-CSM2-MR from China. Simulations were carried out for each GCM, and the results are presented in the supplementary materials as Figures S4, S5, S6, and S7. The average values of the predictions from these four GCMs were then calculated and used as the basis for predicting the future distribution under different climate scenarios (Fig. 4). The estimated uncertainty maps for these four GCMs projections were shown in Figure S2. A statistical evaluation of the total predicted area was also conducted, as shown in Figure S3(A). Under SSP126, the proportion of the total suitable habitat area to the mainland area of China was 12.97%, 12.26%, 11.76%, and 12.05% across four time periods. For SSP245, the proportions were 12.13%, 13.19%, 11.48%, and 11.86%, while for SSP370, the respective values were 13.72%, 13.10%, 13.53%, and 14.54%. Under SSP585, the proportions were 14.05%, 12.58%, 10.75%, and 14.50%. Notably, under SSP370, the largest increase in total suitable habitat area was observed during 2081–2100, accounting for 14.54% of mainland area of China.

The potential suitable distribution of A. sinensis in China under different climate scenarios.
Changes in highly suitable habitat area were also examined. Under SSP126, the highly suitable habitat accounted for 0.53%, 0.38%, 0.12%, and 0.42% of mainland area of China in the four periods, respectively (Figure S3(B)). For SSP245, these proportions were 0.19%, 0.65%, 0.28%, and 0.51% (Figure S3(C)); for SSP370, they were 1.32%, 0.42%, 0.80%, and 0.73% (Figure S3(D)); and for SSP585, the values were 0.57%, 0.69%, 0.72%, and 1.19% (Figure S3(E)). The most significant increase in highly suitable habitat occurred during 2021–2040 under SSP370, reaching 1.32% of mainland area of China. Highly suitable regions are crucial for the sustainable production of A. sinensis, and their spatial dynamics under various SSP scenarios indicate a northward migration of currently optimal habitats, predominantly toward Gansu and Qinghai provinces, with some distribution in Ningxia and Shaanxi. Among these regions, Gansu is projected to retain the highest proportion of highly suitable habitat.
The analysis further revealed that climate change-induced shifts in the potential distribution of A. sinensis habitats will largely involve expansive changes, with localized areas experiencing reductions (Fig. 5). Under SSP126, the largest habitat expansion and reduction occurred during 2021–2040, reaching 166,266 km² and 87,797 km², respectively. For SSP245, the most significant expansion was 101,509 km² during 2041–2060, while the largest reduction, 91,599 km², occurred in 2021–2040. Under SSP370, the maximum expansion was 228,718 km² in 2021–2040, with a reduction of 88,735 km² in 2041–2060. For SSP585, the greatest expansion and reduction were both observed in 2021–2040, reaching 139,755 km² and 78,013 km², respectively. Geographically, suitable habitats for A. sinensis are expected to shift toward higher latitudes. For instance, the southern regions of Sichuan are projected to experience reductions in suitable habitat, while new areas may emerge in northeastern regions. Expansion zones are expected to be concentrated in Gansu, Qinghai, Shaanxi, Ningxia, and Shanxi provinces, while reduction zones will mainly occur in Sichuan and Yunnan.

Changes in the distribution of potential habitat of A. sinensis under different climate scenarios.
Influence of different environmental factors on the differentiation of habitat zones
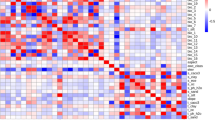
We utilized the Factor Detection method to evaluate spatial stratification heterogeneity within suitability zones and to quantify the influence of environmental factors on their spatial differentiation. Figure 6 illustrates the ‘q’ values and corresponding ‘p’ values for each environmental factor, where ‘q’ represents the correlation between these factors and the spatial differentiation of the fitness zone. The ‘p’ values determine the statistical significance of this correlation. The analysis revealed that among the environmental factors, Srad10, Vapr7, and Elev exhibited the strongest correlation with the spatial differentiation of suitability zones. Furthermore, the differentiation of the suitability zones for A. sinensis was found to be associated with several environmental factors, notably Tmin12, Bio15, Prec5, and Wind10. All environmental variables showed exceptionally low ‘p’ values, below 0.001, indicating that they were significantly correlated with fitness zone differentiation.
The interaction detector analysed the interplay among various environmental factors, delving deeper into how these interactions influenced the spatial differentiation of suitability zones. As depicted in Fig. 6, the q values for interactions between most environmental variables surpassed those of individual variables. These interactions were characterized by mutual enhancement and nonlinear effects. Specifically, variables such as Srad10, Bio4, and Srad5 exhibited interactions with Elevation, resulting in mutual and nonlinear enhancement effects. Furthermore, the interaction of Vapr7 with Bio4, Srad10, Srad3, and Srad5 also demonstrated mutual and nonlinear enhancement, significantly influencing the differentiation of habitat zones. In summary, the interactive impact of environmental variables on spatial differentiation within suitability zones is complex, characterized not by simple additive effects but rather by mutual or nonlinear enhancement.

The explanatory power and interaction of different environmental factors.
Discussion
Prediction of suitable habitat through modelling techniques became a powerful tool for predicting ideal habitats for medicinal plants, enabling the evaluation of habitat suitability and the effective conservation of medicinal plant resources. Moreover, analysing the impacts of climate change on the potential distribution of medicinal plants was indispensable for developing robust conservation strategies. As one of the most common and widely used medicinal plants in China, the conservation and sustainable development of A. sinensis were particularly critical. This study employed the MaxEnt model to systematically investigate, for the first time, the geographic distribution and environmental suitability of A. sinensis habitats in China under the context of global climate change. The findings provided a comprehensive understanding of the spatiotemporal dynamics of A. sinensis habitats, offering valuable insights for planning and implementing effective conservation strategies. This research held profound practical significance for ensuring the long-term sustainability of A. sinensis resources amidst a changing climate.
Previous studies demonstrated that the MaxEnt model had been widely applied in predicting the potential distribution of numerous medicinal plants in China, showcasing exceptional predictive performance23,24,25. Building on this foundation, this study refined the MaxEnt model using the ENMeval package by integrating environmental variables for optimization. This approach emphasized restricting the background dataset to a geographic range aligned with calibration sites, ensuring that the modeled potential geographic distribution accurately encompassed known occurrence points. To further enhance the performance of model, parameter adjustments were implemented, along with tests on varying levels of regularization as a means to reduce model complexity. The optimized MaxEnt model not only significantly reduced its complexity but also improved the consistency between predictions and actual observations. This refinement enabled the production of highly accurate predictions of species distribution, providing a robust basis for assessing and conserving the habitats of medicinal plants such as A. sinensis.
Environmental conditions play a critical role in the formation and transformation of chemical compounds in medicinal plants, and the quality and efficacy of these plants are closely linked to their geographical distribution. The results of this study indicate that the primary environmental factors influencing the distribution of A. sinensis are solar radiation in October, vapor pressure in July, and elevation, which aligns with the preference of species for cooler climates26,27,28. Previous research has shown that light intensity is closely related to the yield of A. sinensis, with the highest yield occurring at a shading rate of 50%, followed by 75% shading29. Additionally, studies by Lin et al.30demonstrated that reducing light intensity through 50–75% shading increases root yield of A. sinensis. During the growing season in July, the higher vapor pressure enhances the transpiration of plant, affecting stomatal regulation and soil moisture levels, which in turn reduces photosynthesis and negatively impacts growth. Therefore, an optimal vapor pressure is critical for the growth of A. sinensis. Elevation gradients alter environmental factors such as light, temperature, and humidity, which subsequently affect the growth and distribution of A. sinensis. Sayti et al. have pointed out that human activity also significantly impacts species distribution patterns31. However, this study focused primarily on environmental factors influencing the growth of species and did not account for human activity. Given that the intensity of human activity is uncertain in future projections, it was not included in this analysis. Thus, it can be inferred that the suitable habitat area identified in this study may be larger than the actual suitable area for A. sinensis.
Compared to the current climate scenario, the suitable habitat area for A. sinensis is projected to increase under future climate scenarios. A. sinensis is a typical habitat-dominated plant, and as a result of global climate change, the environmental conditions such as temperature, precipitation, air humidity, and light intensity in its growing regions will change. These alterations are expected to cause the suitable habitats of A. sinensis to gradually shift toward higher latitudes. This projection aligns with numerous previous studies, which indicate that climate change will significantly impact species’ geographic distributions. Global climate change is rapidly and substantially altering the ecological environment of high-altitude regions, leading to changes in the physiological and ecological adaptability of mountain plants, as well as shifts in their distribution boundaries32. Moreover, under the scenario of global warming, alpine plants are predicted to migrate toward higher altitudes, higher latitudes, and western regions. If the temperature continues to rise, species distributed at high altitudes and latitudes may face the threat of extinction33. In its Sixth Assessment Report released in 2021, the IPCC introduced the concept of SSPs, emphasizing the growing impact of climate change, including rising temperatures and increased precipitation. According to Thomas et al.34, under moderate carbon dioxide emission scenarios in the 2050s, approximately 15-37% of species are likely to face a heightened risk of extinction, while others might experience lower extinction probabilities. Interestingly, some species could even thrive under warming conditions. This highlights the complex and dual nature of influence of climate change on species distribution, where, on the one hand, certain species are at risk, but on the other, some may adapt or even benefit from the changing environment. The contrast in these outcomes underscores the need for more nuanced studies and strategies to address the diverse impacts of climate change on biodiversity.
Based on an in-depth analysis of current and future climate predictions, we propose a scientific classification of the suitable habitats for A. sinensis into three types, and suggest targeted management and conservation strategies for each type: protected areas, loss areas, and new development areas. Firstly, in the context of a diverse greenhouse gas concentration gradient, protected areas will play a key role as critical refuges for A. sinensis in response to climate change. Therefore, efforts should be focused on strengthening the protection and management of these regions to ensure they become the cornerstone for the stable survival of A. sinensis populations. Secondly, in addressing the potential loss of suitable habitats for A. sinensis, it is necessary to thoroughly analyse the underlying causes and, based on this analysis, develop effective in-situ conservation models. Under various climate change scenarios, certain regions will face varying degrees of habitat suitability decline, suggesting that these areas may no longer be suitable for the natural growth of A. sinensis in the future. In light of this, we strongly recommend increasing research investment in these areas, particularly in the fields of reproductive biology, population ecology, and artificial cultivation techniques, to mitigate the risk of genetic resource loss and lay a solid foundation for the long-term conservation and utilization of A. sinensis. Lastly, it is crucial to actively explore and develop new regions for the introduction and expansion of A. sinensis. Based on a thorough understanding of the biological and ecological characteristics of species, suitable locations should be selected for introduction and cultivation demonstration projects. Additionally, comprehensive and detailed spatial planning should be established to scientifically guide large-scale planting and sustainable utilization of A. sinensis in potential areas, ensuring its survival and development in the context of future climate change.
Conclusions
Understanding the potential impacts of climate change on the distribution of A. sinensis is critical for its conservation and sustainable utilization. This study demonstrated that the geographic distribution of A. sinensis in China is predominantly influenced by elevation, with highly suitable habitats concentrated in Gansu and Qinghai provinces, indicating its ecological niche is confined to high-altitude regions. The study results projected that the suitable habitats for A. sinensis would generally shift toward higher latitudes over time, accompanied by an overall expansion in habitat area. These results provide essential insights into the potential responses of species to climate change and offer a theoretical basis for developing conservation, cultivation management, and sustainable utilization strategies. The results of this study can serve as a reference for identifying high-priority areas for reintroduction and protection, enabling proactive measures to mitigate the projected impacts of climate change and ensure the long-term viability of A. sinensis resources. However, there are limitations to the current study, including the assumption that key environmental variables such as wind speed and solar radiation will remain constant under future climate scenarios, which may oversimplify the potential impacts of climate change on A. sinensis habitats. Additionally, the study did not incorporate projected changes in land cover, which could significantly alter habitat suitability and species distribution, warranting future research to refine these models and integrate more comprehensive data.
Data availability
The original contributions presented in this study are included in the article/supplementary material, and further inquiries can be directed to the corresponding author.
References
Dawson, T. P., Jackson, S. T., House, J. I., Prentice, I. C. & Mace, G. M. Beyond predictions: Biodiversity Conservation in a changing climate. Science 332, 53–58. https://doi.org/10.1126/science.1200303 (2011).
Glob, B. Outlook, 5(4) 267–284. (2010).
Stocker, T. F. et al. IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. (Cambridge University, Cambridge, New York, 2014).
Liu, L. et al. Modeling habitat suitability of Houttuynia cordata Thunb (Ceercao) using MaxEnt under climate change in China. Ecol. Inf. 63, 101324. https://doi.org/10.1016/j.ecoinf.2021.101324 (2021).
Wang, G. et al. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: suggestions for conservation of endangered red-crowned crane. Ecol. Ind. 116 https://doi.org/10.1016/j.ecolind.2020.106472 (2020).
Soilhi, Z., Sayari, N., Benalouache, N. & Mekki, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inf. 68, 101533. https://doi.org/10.1016/j.ecoinf.2021.101533 (2022).
Zhang, L. B., Lv, J. L. & Liu, J. W. Phthalide derivatives with anticoagulation activities from Angelica Sinensis. J. Nat. Prod. 79, 1857–1861. https://doi.org/10.1021/acs.jnatprod.6b00080 (2016).
Shen, J. et al. The angelica Polysaccharide: a review of phytochemistry, pharmacology and beneficial effects on systemic diseases. Int. Immunopharmacol. 133 https://doi.org/10.1016/j.intimp.2024.112025 (2024).
Hua, Y., Xue, W., Zhang, M., Wei, Y. & Ji, P. Metabonomics study on the hepatoprotective effect of polysaccharides from different preparations of Angelica Sinensis. J. Ethnopharmacol. 151, 1090–1099. https://doi.org/10.1016/j.jep.2013.12.011 (2014).
Wei, W. L., Zeng, R., Gu, C. M., Qu, Y. & Huang, L. F. AngSinensisnensis in China-A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis. J. Ethnopharmacol. 190, 116–141. https://doi.org/10.1016/j.jep.2016.05.023 (2016).
Wang, K. et al. Chemical composition analysis of Angelica Sinensis (Oliv.) Diels and its four processed products by ultra-high‐performance liquid chromatography coupled with quadrupole‐orbitrap mass spectrometry combining with nontargeted metabolomics. J. Sep. Sci. 46 https://doi.org/10.1002/jssc.202300473 (2023).
Zou, J. et al. Triligustilides A and B: two pairs of Phthalide trimers from Angelica sinensis with a Complex Polycyclic Skeleton and their activities. Org. Lett. 20, 884–887. https://doi.org/10.1021/acs.orglett.8b00017 (2018).
Xia, Z. et al. Diverse neolignans and lignans from an aqueous extract of the Angelica sinensis root head. J. Asian Nat. Prod. Res. 25, 919–940. https://doi.org/10.1080/10286020.2023.2173181 (2023).
Zhi, X. et al. Therapeutic potential of Angelica Sinensis in addressing organ fibrosis: a comprehensive review. Biomed. Pharmacother. 173 https://doi.org/10.1016/j.biopha.2024.116429 (2024).
Han, Y. et al. Overview of therapeutic potentiality of Angelica Sinensis for ischemic stroke. Phytomedicine 90 https://doi.org/10.1016/j.phymed.2021.153652 (2021).
Tuo, W. et al. Angelica Sinensis polysaccharide extends lifespan and ameliorates aging-related diseases via insulin and TOR signaling pathways, and antioxidant ability in Drosophila. Int. J. Biol. Macromol. 241 https://doi.org/10.1016/j.ijbiomac.2023.124639 (2023).
Wang, D. et al. Moving north in China: the habitat of Pedicularis kansuensis in the context of climate change. Sci. Total Environ. 697 https://doi.org/10.1016/j.scitotenv.2019.133979 (2019).
Puchałka, R. et al. Forest herb species with similar European geographic ranges may respond differently to climate change. Sci. Total Environ. 905, 167303. https://doi.org/10.1016/j.scitotenv.2023.167303 (2023).
Soilhi, Z., Sayari, N., Benalouache, N. & Mekki, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inf. 68, 101533. https://doi.org/10.1016/j.ecoinf.2021.101533 (2022).
Zheng, T. et al. Evaluation of climate factors affecting the quality of red huajiao (Zanthoxylum bungeanum maxim.) based on UPLC-MS/MS and MaxEnt model. Food Chemistry: X, 16, 100522. https://doi.org/10.1016/j.fochx.2022.100522 (2022)
Peng, W., Kuang, T. & Tao, S. Quantifying influences of natural factors on vegetation NDVI changes based on geographical detector in Sichuan, western China. J. Clean. Prod. 233, 353–367. https://doi.org/10.1016/j.jclepro.2019.05.355 (2019).
Gilani, H., Arif Goheer, M., Ahmad, H. & Hussain, K. Under predicted climate change: distribution and ecological niche modelling of six native tree species in Gilgit-Baltistan, Pakistan. Ecol. Ind. 111 https://doi.org/10.1016/j.ecolind.2019.106049 (2020).
Wan, G. Z., Guo, Z. H., Xi, S. Y., Jin, L. & Chen, J. Spatial variability and climate response characteristics of chemical components of Tussilago farfara L. Ind. Crops Prod. 204 https://doi.org/10.1016/j.indcrop.2023.117352 (2023).
Wan, G. Z., Wang, L., Jin, L. & Chen, J. Evaluation of environmental factors affecting the quality of Codonopsis pilosula based on chromatographic fingerprint and MaxEnt model. Ind. Crops Prod. 170, 113783. https://doi.org/10.1016/j.indcrop.2021.113783 (2021).
Li, J., Fan, G. & He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 698, 134141–134148. https://doi.org/10.1016/j.scitotenv.2019.134141 (2020).
Park, Y. et al. The characteristics of the growth and the active compounds of Angelica Gigas Nakai in Cultivation sites. Plants (Basel). 9 https://doi.org/10.3390/plants9070823 (2020).
Dong, H. et al. Cool temperature enhances growth, Ferulic Acid and Flavonoid Biosynthesis while inhibiting Polysaccharide Biosynthesis in Angelica Sinensis. Molecules 27 https://doi.org/10.3390/molecules27010320 (2022).
Chen, J. M. et al. Explore the interaction between root metabolism and rhizosphere microbiota during the growth of Angelica Sinensis. Front. Plant. Sci. 13, 1005711. https://doi.org/10.3389/fpls.2022.1005711 (2022).
Xiang, H., Li, Y. & Jin, Y. Effects of light intensity on yield and ferulic acid content in Angelica Sinensis. J. Traditional Chin. Veterinary Med. 34, 53–54. https://doi.org/10.13823/j.cnki.jtcvm.2015.02.017 (2015).
Lin, H. et al. Influence of sun shade cultivation on premature bolting in Angelica Sinensis and growth environment factors. Chin. J. Experimental Traditional Med. Formulae. 16, 79–83. https://doi.org/10.13422/j.cnki.syfjx.2010.04.003 (2010).
Sayit, H., Nurbay, A. & Arman, J. Impact of human activities on potential distribution of Solanum rostratum in Xinjiang. Acta Ecol. Sin. 39, 629–636 (2019).
Lawlor, J. et al. Mechanisms, detection and impacts of species redistributions under climate change. Nat. Reviews Earth Environ. 5, 351–368. https://doi.org/10.1038/s43017-024-00527-z (2024).
Lamsal, P. et al. Historical evidence of climatic variability and changes, and its effect on high-altitude regions: insights from Rara and Langtang, Nepal. Nat. Reviews Earth Environ. 24, 471–484. https://doi.org/10.1080/13504509.2016.1198939 (2017).
Thomas, C. et al. Extinction risk from climate change. Nature 427, 145–148. https://doi.org/10.1038/nature02121 (2004).
Acknowledgements
The author would like to acknowledge financial support from the Strategic Research and Consulting Project of the Chinese Academy of Engineering (No. GS2021ZDA06), and the Ecological Planting, and Quality Assurance Project of Daodi Medicinal Materials (China Science and Technology for Traditional Chinese Medicine (2020) 153), and Chinese Academy of Traditional Chinese Medicine Traditional Chinese Medicine Resource Center, Research on the Changes of Angelica sinensis Production Areas and Production Regionalization (NO.ZYZX-2023-KY-083).
Author information
Authors and Affiliations
Contributions
X.S. and G.X. wrote the main manuscript text and M.X. prepared figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xi, S., Guo, X., Ma, X. et al. Impacts of climate change on the suitable habitat of Angelica sinensis and analysis of its drivers in China. Sci Rep 15, 3508 (2025). https://doi.org/10.1038/s41598-025-87436-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-87436-3
Keywords
This article is cited by
-
Spatial distribution changes of cold-water Zoarces fish biodiversity and inter-species ecological competition replacement under climate change scenarios
Environmental Monitoring and Assessment (2026)
-
Predicting the potential distribution and climatic response of the endangered medicinal and edible species, Anoectochilus roxburghii, using an optimized MaxEnt model
Scientific Reports (2025)
