
Abstract
Eld’s deer (Rucervus eldii) is a rare and globally endangered tropical Southeast Asian deer species. There is no research on pathogens in Eld’s deer in Hainan, China. This study aimed to understand the virus diversity and novel viruses in Eld’s deer, and provided important epidemiological baseline information for conservation of this endangered species. 33 nasal swabs, 33 anal swabs, and 9 wound (bitten by ticks) swabs were collected from 33 wild Eld’s deer in a nature reserve in Hainan, which constituted into 5 pools. Based on next-generation sequencing (NGS) and macrogenomic analysis, there were differences in the 5 pools of viral reads, while the overall viral reads were closely related to mammals. The novel papillomavirus (PsPV-HMU-1) and Circular Rep-encoding (replication-associated protein encoding) single-stranded DNA (CRESS DNA) virus (PsaCV-HMU-1) were identified in Eld’s deer, with amino acid homology of the less than 77.20% of the L1 and less than 45.43% of the rep, respectively. PsPV-HMU-1 and PsaCV-HMU-1 are relatively independent on their phylogenetic trees, and with the overall prevalence of 24.24% (8/33) and 3.03% (1/33) in Eld’s deer, respectively. Our results expanded the viral genomic information and host range, and implied that it is necessary for continued epidemiological surveillance in order to understand pathogenicity and the potential for cross-species transmission of viruses in wild Eld’s deer.
Similar content being viewed by others

Introduction
Eld’s deer (Rucervus eldii) is a rare and globally endangered tropical Southeast Asian deer species, belonging to Artiodactyla, Family Cervidae and Subfamily Cervinae. It has been listed in Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) and classified as endangered on the Red List of Threatened Species by the International Union for Conservation of Nature (IUCN) and the class I national key protected wildlife in China1. Only over 360 Eld’s deer were surviving in India in 20032. This species saw a 90% decline in Cambodia between 1998 and 2008 and a 50% decline in the ensuing 10 to 15 years. The number of Eld’s deer in Laos and Myanmar is also declining annually, and the wild population of Eld’s deer are thought to be regional extinct in Thailand and Vietnam2. Eld’s deer on Hainan Island is the only island subspecies, which once was widely distributed in Hainan, China. However, due to factors like habitat loss and destruction, blind hunting, and illegal trade, their population has declined sharply and is in danger of going extinct3. There were only 26 Eld’s deer remaining in Hainan in the 1970s4. At present, Eld’s deer are mainly distributed on the Hainan Datian National Nature Reserve and Hainan Bangxi Provincial Nature Reserve in China. Notably, the higher population density in the latter, with near 300 deer, could lead to the increasing risk of virus transmission among wildlife5. The outbreaks and prevalence of viral infectious diseases among endangered wildlife could lead to population collapse and impact on their survival.
Pathogenic organisms can cause outbreaks and even lead to population collapse in wild animals. Virus infections in Cervidae are a serious threat for health and survival of some deer species worldwide, such as peste des petits ruminants virus (PPRV) in water deer6, hog deer, thamin, and mouse deer7, herpesviruses in red deer, sambar, milu, reindeer and mule deer8,9,10, epizootic hemorrhagic disease virus (EHDV) and bluetongue virus (BTV) in mule deer and white-tailed deer11,12, and Bovine Viral Diarrhea Virus (BVDV) in wild cervid species of South America13. Papillomaviruses (PVs) and Circular Rep-encoding (replication-associated protein encoding) single-stranded DNA (CRESS DNA) viruses also have been identified in deer species. PVs constitute a diverse group of small non-enveloped viruses, with double-stranded DNA genomes, and belong to the family Papillomaviridae, which included two subfamilies, Firstpapillomavirinae and Secondpapillomavirinae, and more than 50 virus genera14. PVs can infect the stratified squamous epithelia of skin and mucosa of humans and various animals15,16, in which they can appear asymptomatic or cause different neoplastic changes, ranging from self-limited warts to malignant carcinomas, such as cervical and genital cancers14,17,18. In cervids, most of the detected PVs belong to the Deltapapillomavirus genus and cause neoplastic changes, predominately on the neck, head, abdomen and extremities, which were commonly described as endemic disease in the host population19. Larger tumours can become ulcerated and eroded, even form metastases in the lungs and eyes15,20,21. To date, PVs have been detected in papillomas of cervid species wordlwide, including RalPV1 in Visayan spotted deer from UK18, RtPV2 in reindeer from Norway21, RtimPV1 and RtimPV2 in Timor deer from Germany22, CcaPV1-type in roe deer, CePV1v-type variant in red deer from Slovenia19, Hungary, Austria, Croatia20 and Italy16,17, CePV2 in red deer from New Zealand23, and RtPV4 in caribou from USA24.
The Circular Rep-encoding (replication-associated protein encoding) single-stranded DNA (CRESS DNA) viruses associated with eukaryotic hosts have been classified by the International Committee on Taxonomy of Viruses (ICTV), namely Circoviridae, Genomoviridae, Geminiviridae, Nanoviridae, Bacilladnaviridae, and Smacoviridae25,26, which are the varied collection of tiny viruses and identified in prokaryotes, eukaryotes, and environmental samples27,28. It was reported that infection of CRESS DNA viruses was the significant risk factor for diarrheal disease in domestic animals, such as Porcine Circovirus (PCs) in diarrheal pigs, cattle and donkeys from China29,30,31 and Dominican Republic32. Furthermore, porcine circoviral infections were the important trigger for various clinical manifestations in pigs, including post-weaning multi-systemic wasting disease syndrome (PMWS), porcine dermatitis and nephropathy syndrome (PDNS), respiratory distress, reproductive failure, systemic inflammation, vasculitis and myocarditis, respiratory and enteric signs33,34,35. Although the reports on deer’s clinical presentations caused by CRESS DNA viruses are still very limited, these viruses have been identified in captive and wild cervids, including red deer36, sika deer37 and forest musk deer38 in China, roe deer in USA39 and Italy40, Rocky Mountain elk in Canada41 and fallow deer in New Zealand42 and Spain43. Therefore, it is necessary to take attention on the cross-species transmission and clinical pathogenic mechanisms of viruses among artiodactyl.
Complete genome and characteristic of new viruses have been identified and described from animal fecal successfully by viral metagenomic techniques, such as next-generation sequencing (NGS). So far, there is no study on papillomaviruses and CRESS DNA viruses in Eld’s deer worldwide. In this study, two complete virus sequences of papillomavirus and CRESS DNA virus from Elder’s deer were obtained for the first time, which were subsequently identified and described as PsPV-HMU-1 and PsaCV-HMU-1. It is crucial to protect Hainan Eld’s deer by comprehending the diversity, evolution and variation patterns of viruses in this endangered wildlife, and their potential risk for health of Eld’s deer (Fig. 1).

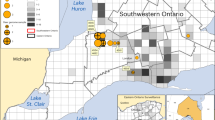
Distribution of sampling sites of Eld’s deer in the Hainan Bangxi Provincial Nature Reserve in the present study. The specific sampling points (Fence No.1, 2 and 8) have been marked on the map.
Results
Viral metagenome of Eld’s deer
In present study, 33 nasal and anal swabs, and 9 swabs of wound bitten by ticks of wild Eld’s deer were obtained from the nature reserve in Hainan, China. Efforts were made to eliminate bacteria, microbial eukaryotes, archaea, and organisms that showed no discernible relationship to any amino acid sequences in the viral non-redundant (NR) protein database. Five pools yielded 59.50 GB of nucleotide data (58,402,837 valid reads, 150 bp in length). The sequences of less than 50 bp, archaea, bacteria, microbial eukaryotes (such as fungi), and those that had no significant similarity with any amino acid sequences in the viral NR protein database were eliminated (Table S1). Retrotranscribing viruses, ssRNA, double-stranded (ds) DNA, single-stranded (ss) DNA, and ssRNA made up the viromes. There were differences and tissue tropism in viral composition and distribution among different type of samples. The virus-related reads in 2 pools with nasal swabs (Pool 1 and 2) were mainly in Ackermannviridae and Schitoviridae families, while the viruses in Picornavirdae and Parvoviridae families were not identified in samples from Pool 1 and 2, respectively. The majority of viruses in 2 pools with anal swabs (Pool 3 and 4) were belonged to Myoviridae, Siphoviridae, and Microviridae families, and a large number of unclassified reads (Table S1). Significantly, low nucleotide and amino acid sequence identities were observed between most reads of sequence and known viruses. In Pool 4 and 5, the proportion of virus-associated reads was 2,039,836 (16.97%) and 3,128,644 (28.15%), including that corresponded to CRESS DNA virus and papillomavirus were 0.13% and 0.12% of all the viruses, respectively (Fig. 2A and B).

Next-generation sequencing (NGS) sequencing data of the virome profile of 5 pools. (A) Viral content in Pool 4 composed by anal swab samples. (B) Viral content in Pool 5 composed by tick bite wound swab samples.
Genomic characterization of novel PV
A complete genome of Papillomavirus (PsPV-HMU-1) (7379 bp in length) in Eld’s deer was obtained from Pool 5, with genbank accession PQ037580. A core collection of early (E6, E7, E1, and E2) and late (L2 and L1) open-reading frames (ORFs) was predicted using Pfam software (Fig. 3A). PsPV-HMU-1 also exhibits conserved LCR organizations among PV types belonging to the same genus. The non-coding LCR region of PsPV-HMU-1 (Fig. 3B), located between the late and early genes, contained a single polyadenylation (PolyA) signal, TATA box, nuclear factor (Nf1) (TTGGC), and four E2 binding sites, following the ‘traditional’ four E2-binding site (ACCN2-11GGT) pattern found in most mammalian papillomaviruses. The E7 and E6 protein upregulate DNA replication, inhibit cell checkpoints, and block apoptosis in mammalian hosts. The novel PV has a prototypical mammalian E6 protein consisting of two highly conserved zinc finger motifs (C-X2-C-X29-C-X2-C). The C-terminal zinc-binding domain suggests that the E7 protein conforms to the classical E7 protein observed in mammalian papillomaviruses (Fig. 3C). An ATP/GTP binding site (G-X4-GKS) was present in the E1 helicase domain.

Genomic structure and motif annotation of PsPV-HMU-1. (A) Using the ORF predictor, putative and confirmed early and late ORFs were found in the genomes of PsPV-HMU-1. LTR, long-terminal repeat; early (E6, E7, E1, E2); and late (L2, L1). (B) The motif structure diagram of LTR in PsPV-HMU-1. (C) The CR1, CR2, and N-terminal structure diagram of E7.
Phylogenetic classification of the genomes of novel PV
The mammalian PVs were used as references, which was primarily based on BLAST results, and the early evolutionary analysis of the ICTV novel PVs. A small number of sauropsid and fish PV clades were selected as outgroups. The phylogenetic tree is consistent with the evolutionary relationships among three distinct monophyletic branches: avian, sauropsid, and mammalian papillomavirus, as proposed by the ICTV. The PVs most closely related to PsPV-HMU-1 were those from the genus Dyokappapapillomavirus (Table S2), particularly bovine papillomavirus 22 (BVP 22) (NC035208) (Fig. 4).

Phylogenetic tree of PsPV-HMU-1, concatenated early (E1, E2) and late (L2, L1) papillomavirus nucleotide sequences. The maximum likelihood approach and a 1000 bootstrap replication were used to construct the tree. The scale bar indicates the number of amino acid changes per site, and bootstrap values < 50% are not shown.
Genomic characterization of novel CRESS DNA virus
The complete Eld’s deer-associated circular single-stranded (ssDNA) virus genome (PsaCV-HMU-1) was obtained from Pool 4, with GenBank accession: PQ119789. PsaCV-HMU-1 has genome length of 2,722 bp, with C + G content of 40.56%. The genome contains two major open reading frames (ORFs), encoding the replication-associated protein (Rep) and the capsid protein (Cp) on different strands of a double-stranded DNA (dsDNA) replicative form. The rolling circle replication (RCR) mechanism of circular CRESS DNA viruses is associated with conserved Rep motifs, including N-terminal RCR motifs (I, II, and III), C-terminal superfamily 3 helicase motifs (Walker-A, -B, and Motif C), and an arginine finger, which together have a role in the initiation, elongation, and termination of the RCR of CRESS DNA viruses of eukaryote origin. Similar to other CRESS DNA viruses, PsaCV-HMU-1 contains a conserved sequence (CAGTGTTAC), which was located in a putative stem loop structure at the origin of replication.
Phylogenetic classification of the genomes of novel CRESS DNA virus
Due to the diversity of species and genomes, it is a challenge to classify CRESS DNA virus effectively. The representative virus sequences from the Circoviridae, Smacoviridae, and CRESS DNA viruses were used to jointly establish a phylogenetic tree. Notably, although the phylogenetic trees of Rep and Cp are almost consistent with each other, the Rep of PsaCV-HMU-1 was segregated from the known CRESS DNA viruses at the deep root of the phylogenetic tree. PsaCV-HMU-1, only distantly related to the Circoviridae and Smacoviridae, was on the separate branch with unclassified CRESS virus 1 (CRESSV1) genomes together, particularly the camel-associated drosmacovirus (KM573766) (Fig. 5A and B). The Rep and Cp genome sequences of PsaCV-HMU-1 showed lower amino acid identities of 45.43% and 31.73%, respectively, compared to other known viruses (Table S3). Additionally, these sequences were difficult to compare with those of other known circular viruses at the nucleic acid level in the NCBI nucleotide database.

Phylogenetic tree based on the amino acid sequences of the complete Rep and Cp amino acids of PsaCV-HMU-1. Phylogenetic tree constructed by the maximum likelihood method using the best-fit models (LG + G + I). The scale bar indicates the number of amino acid changes per site, and bootstrap values < 50% are not shown. (A) Phylogenetic tree based on Rep amino acid sequence. (B) Phylogenetic tree based on the Cp amino acid sequence.
Prevalence of novel virus infection in Eld’s deer
Quick sampling within two months is necessary to track the percentage of deer that are viral carriers within the same season. Five pools were examined for positivity of PsPV-HMU-1 and PsaCV-HMU-1. For each individual sample in the positive-result pool, conduct a second nucleic acid extraction and a positive rate screening. PsPV-HMU-1 and PsaCV-HMU-1 positive samples yielded amplification products of 1529 bp and 1169 bp, respectively. In this study, the overall prevalence of PsPV-HMU-1 and PsaCV-HMU-1 in wild Eld’s deer in Hainan were 24.24% (8/33) and 3.03% (1/33), respectively (Table 1). Specifically, one female and two males in Fence No.1, and five females in Fence No.2 infected PsPV-HMU-1, but the samples of Eld’s deer in Fence No.8 were negative. PsPV-HMU-1 was identified in both nasal and wound samples in three females, one from Fence No.1 and two from Fence No.2, while that was found in both anal and wound samples in two females from Fence No.2. PsaCV-HMU-1 was identified in a female from Fence No.2. There was no mixed infection with two viruses in this study.
Discussion
The viral outbreaks in endangered cervid species, such as Visayan spotted deer in UK18, huemul deer in Chile44, water deer and forest musk deer in China6,45, marsh deer in Brazil46, barking deer, hog deer, thamin and mouse deer in India7, have temporarily reduced their populations and threatened survival of species. The past infection of H1N1pdm09 virus was determined in Eld’s deer in Thailand, which implied the susceptibility of this species to natural infection and potentiality of the cross-species transmission of human influenza virus to Eld’s deer47. So far, except for influenza virus, there are no more reports on others viruses in globally endangered Eld’s deer. In present study, 5 pools of samples from wild Eld’s deer, including 2 pools of nasal swabs (Pool 1 and 2), 2 pools of anal swabs (Pool 3 and 4), and a pool of wound bitten by ticks (Pool 5), mainly contained host carried eukaryote. There were differences and tissue tropism in viral composition and distribution among different type of samples, especially abundant unclassified reads and viruses in anal and wound swabs, which highlighted the importance for focusing on their characteristics, the potentiality for pathogenicity and cross-species transmission.
The evolutionary history of papillomaviruses extends back at least 400 million years ago (MYA) as evidenced by the recent discovery of fish papillomaviruses48. Osteichthyes and Amniota shared a common ancestor around 400 MYA49. The host range and virological categorization of this group of viruses have been broadened by the isolation and characterization of the novel viruses from species of the taxa Carnivora and Artiodactyla. The most PVs in cervids mainly induce fibropa pillomas but also malignant carcinomas50, such as CePV-1 variant in red deer in Portugal15, CcaPV1-type variant in roe deer in Slovenia19, PpudPV1 and CelaPV2 in Pudu puda and Cervus elaphus Deer51. In addition, based on observations of infection with BPV-1 and BPV-2 results in the most common skin tumor of horses and sheep52,53, and the detection of OaPVs in cattle from intensive livestock farming without any contact with sheep54, cross-species transmission of papillomaviruses among ruminant animals seems to be an ongoing process.
In this study, a novel papillomavirus (PsPV-HMU-1) was identified in both wound swabs and nasal or anal swabs of Eld’s deer, which indicated that the infection was persistent. Based on PsPV-HMU-1 and other recent studies, all mammalian papillomaviruses in the Dyokap genus contain seven typical sequences (LCR E6, E7, E1, E2, L2, and L1). The pairwise alignment of the L1 ORF showed a maximum nucleotide sequence homology of 76.11% with the most similar sequence of the BvPV 22. The amino acid identities of L1 between PsPV-HMU-1 and species within the Dyokap genus range from 56.47 to 77.20% (Table S2). PsPV is distantly related to human-associated papillomaviruses. Based on sequence identity criteria and the distance of PsPV-HMU-1 to other viruses in the Dyokap genus, PsPV-HMU-1 is likely to be classified as a new papillomavirus species within the Dyokap genus (Fig. 4). PsPV-HMU-1 shares many features with other viruses in the Dyokap genus, providing evidence for host-virus co-speciation. However, utilizing “molecular morphology” information can more accurately determine the phylogenetic placement of Fulmarus glacialis papillomavirus 1 (FgPV1)55, emphasizing the importance of molecular-level analysis, especially early protein analysis. Moreover, the potential threat of insect vectors for animal populations was emphasized by the high prevalence of PsPV-HMU-1 in samples of wound bitten by ticks (Table 1). Significantly, compared with the Fence No.2 and 8, Fence No.1 was a completely independent habitat, in order that there was no contact and communication between Eld’s deer population in Fence No.1 and others. However, PsPV-HMU-1 was identified in populations both in Fence No.1 and 2. The viral infection in wild animals was influenced by living conditions, biogeographic distributions, age, gender, susceptibilities and health status of individuals56,57. Thus, it is necessary to continue surveillance with a larger sample size, in order to analyze the risk of pathogenic potential, prevalent factors and possibility of cross-species transmission of PsPV-HMU-1 in Eld’s deer.
CRESS DNA viruses are highly diverse, and have a broad range of hosts and widely prevalent in domestic animals in China58. In this study, a novel CRESS DNA virus, named PsaCV-HMU-1, was obtained in snal swab of a wild female healthy Eld’s deer from a nature reserve in Hainan, China. Not only it is difficult to compare the rep of PsaCV-HMU-1 with other CRESS DNA viruses at the nucleotide level, but also its amino acid homology with the closest Dromedary stool-associated circular ssDNA virus is 45.43% (Table S3). In the phylogenetic tree, PsaCV-HMU-1 was grouped with CRESSV1, but it belonged to separate groups of the Cirlivirales order (Fig. 5). Additionally, there is no species demarcation criteria for defining CRESSV1 and CRESSV2. Based on the distance between PsaCV-HMU-1 and known sequence on the phylogenetic tree, it is suggested that the new virus may belong to a different virus species. Therefore, it is hard to assign PsaCV-HMU-1 to the appropriate viral family. These findings provide some insight into the ecology of tiny circular DNA virus genomes and aid in the construction of data sets related to them. Closely related CRESS DNA viruses of the Cirlivirales and Cremevirales orders, such as those in the Circoviridae and Smacoviridae families, may be present in the intestines of swine and other mammals (such as bovines, fur seals, or primates), indicating that multiple host species can be infected despite differences in genomic sequences35. The known and novel CRESS DNA viruses have been identified in captive and wild deer, especially a novel circovirus (ElkCV) in a wild elk in Canada41, who displayed labored breathing, multisystemic hemorrhage, splenomegaly and pulmonary edema. Although the relationship between ElkCV and the lesions in elk remains unclear, in pigs and cattle, there has been a proposed association between infection of circoviruses and varying degrees of lesions, even more severe outcomes59. It is indicated that the continued epidemiological surveillance of CRESS DNA viruses would be useful to establish a potential association between the presence of the virus in cervids and clinical disease.
Gaining more insight into the genomic characteristics, genetic diversity, evolutionary processes and history of the virus carried by Eld’s deer will be possible through extensive genomic and phylogenetic analysis of PsPV-HMU-1 and PsaCV-HMU-1, by comparded with other mammalian and non-mammalian viruses. The results of this study are helpful for epidemiological surveillance and the potential for cross-species transmission of PVs and CRESS DNA viruses in wild Eld’s deer, and will provide a reference for the protection of this globally endangered species.
Conclusion
In summary, the virome in nasal, anal and wound swab samples of wild globally endangered Eld’s deer in Hainan was identified in detail for the first time in our study. The contents include genome protein structure, stem-loop structure and rep protein phylogenetic analysis. Two novel viruses show sequence similarity to papillomavirus and CRESS-DNA viruses, and named as PsPV-HMU-1 and PsaCV-HMU-1, respectively. PsPV-HMU-1 is prevalent in Eld’s deer, while PsaCV-HMU-1 is obtained in only one individual, their pathogenicity has not been known. This study increased the knowledge of the diversity of viruses in Eld’s deer.
Materials and methods
Swab sample collection
All sampling procedures and methods, without employed anesthesia, euthanasia or hunting, were carried out in accordance with relevant guidelines and regulations of the Ethics Committee of the Hainan Medical University (approval no. HMUEC20180059). During the translocation process of Eld’s deer between the nature reserves, thirty-three wild Eld’s deer (20 females and 13 males) were captured from three isolated fenced areas (Fence No.1, 2 and 8) in the Bangxi Provincial Nature Reserve in Hainan Province (Fig. 1) by experienced reserve staff with net in December 2023 and January 2024. All Eld’s deer are healthy, including nine of them have been bitten by ticks on their ears, and were protected in accordance with the biosafety commitment letter signed before sampling. After ticks were picked away by tweezers from these wild Eld’s deer, we collected 33 nasal swabs, 33 anal swabs, and 9 swab samples of wound on ears bitten by ticks. All swab samples were promptly submerged in virus-sampling tubes (Yocon Biology, Beijing, China) that will not inactivate the virus, kept in liquid nitrogen and sent to the laboratory within 24 h. Then the swab samples were split and stored at -80 °C. In this study, we only conducted microbiological identification in the swab samples by NGS, not involved any experiments on live animals. Combined type of swabs and location of Eld’s deer’s habitats, 5 pools were obtained, including 2 nasal pools (Pool 1 with nasal swabs from Fence No.1, Pool 2 with nasal swabs from Fence No.2 and 8), 2 anal pools (Pool 3 with anal swabs from Fence No.1, Pool 4 with anal swabs from Fence No.2 and 8), and a pool of wound bitted by ticks (Pool 5 with the wound swabs from Fence No.1, 2 and 8).
Viral nucleic acid library construction and NGS
The combined sample was ultracentrifuged and the precipitates from the samples were resuspended in 100 µL of Hank’s balanced salt solution and digested with a cocktail ofenzymes to decompose, as previously reported14. Viral DNA and RNA were extracted using QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). The first strand of cDNA was synthesized using Superscript III Reverse Transcriptase (Invitrogen, Thermo Fisher Scientific), and the double-stranded DNA was synthesized in compliance with previous guidelines14,60,61. The nucleic acid library was then constructed. The amplified viral nucleic acid libraries were sequenced, as previously reported. The valid sequence data that have been cleaned and filtered were deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive under the accession number PRJNA1138552.
Taxonomic assignment
The methods and parameters for extracting effective sequences after cleaning and screening the original sequences were applied using the previously described criteria, followed by sequence similarity-based taxonomic assignments14,62. First, we used Trinity (version 2.5.1) to assemble the randomly distributed reads across the entire viral genome into contigs, and compared them with the National Center for Biotechnology Information (NCBI) nonredundant protein (NR) database using basic local alignment search tool (BLAST) (E-value < 10− 5, -F: filter query sequence, default = T), and then with our in-house database (http://ftp.ncbi.nih.gov/pub/taxonomy/accession2taxid/) for classification annotation and other information obtained from the NCBI Entrez server.
Genome sequencing and annotation of novel virus
Based on the results of the virus categorization annotation, extract pertinent CRESS DNA virus and papillomavirus reads, as described previously. Sequence the amplified products and then splice them using SeqMan with the default settings (Version 7.1.0.44). A list of primers used to amplify the sequences found during this inquiry is provided in Table S4. Use EditSeq (Version 7.1.0.44) to predict the position of the open reading frame, as described previously. Conserved protein families and domains were predicted, as described previously.
Phylogenetic and data analyses
The ICTV was used to obtain the representative reference sequences required to construct the phylogenetic tree (https://ictv.global/report/genome). The reference sequences with close genetic relationships were obtained from GenBank. Alignments were prepared with MEGAX using the MUSCLE package with default parameters, and the best substitution model was evaluated using the Model Selection tool. To determine the origins and evolutionary history of novel PV, the complete E1-E2-L2-L1 nucleotide sequences of 42 representative viruses were randomly selected and used for phylogenetic analysis. The phylogenetic tree was constructed using the maximum likelihood method (GTR + G) with 1,000 bootstrap replicates. The classification and phylogeny of novel circular virus are based on the available replicase (Rep) and capsid (Cp) sequences using the maximum likelihood method (LG + F + G + I) with 1,000 bootstrap replicates.
Calculation of prevalence
RNA was extracted from all individual samples using the QIAamp Virus RNA Mini Kit (Qiagen, Hilden, Germany). Specific PCR primers were designed (Table S4) to amplify the conserved regions of L1 in PV and Rep in CRESS DNA viruses. Sequencing results revealed > 99% similarity between the amplified products and the novel viruses, which can be used for positive rate detection. PCR was performed using GoTaq Colorless Master Mix (Promega). Nested PCR used two microliters of the first-round PCR product as the template for the second round of PCR. The thermal cycling conditions for both PCRs were as follows: 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 57 °C for 35 s, 72 °C for 30 s, and a final elongation step at 72 °C for 10 min. If the product is consistent with expectations60, it is determined to be a positive sample.
Data availability
The datasets presented in this study can be found in online repositories of the National Center for Biotechnology Information (NCBI). The accession number(s) was PRJNA1138552.
References
Ghazi, M. G. et al. Population genetics and evolutionary history of the endangered Eld’s deer (Rucervus eldii) with implications for planning species recovery. Sci. Rep. 11, 2564 (2021).
Cardoso, P., Stoev, P., Georgiev, T., Senderov, V. & Penev, L. Species conservation profiles compliant with the IUCN red list of threatened species. Biodivers. Data J. e10356 (2016).
Liu, H. Y., Xue, F., Wan, Q. H. & Ge, Y. F. MHC class II Β genes in the endangered Hainan Eld’s deer (Cervus Eldi hainanus). J. Hered. 104, 874–880 (2013).
Pan, D., Song, Y. L., Zeng, Z. G. & Bravery, B. D. Habitat selection by Eld’s deer following relocation to a patchy landscape. PLoS One 9, e91158 (2014).
Zakharova, O. I. & Liskova, E. A. Patterns of animal rabies in the Nizhny Novgorod region of Russia (2012–2022): the analysis of risk factors. Front. Vet. Sci. 11, 1440408 (2024).
X Y, Z. et al. First report of peste des petits ruminants virus lineage II in Hydropotes inermis, China. Transbound. Emerg. Dis. 65, e205–e209 (2018).
Barman, N. N. et al. First incidence of peste des petits ruminants virus in Cervidae family from state zoo of Assam, India. Viruses 16, 1829 (2024).
Gutiérrez, J. F. M. et al. Infectious keratoconjunctivitis in free-ranging mule deer in Wyoming: a retrospective study and identification of a novel alphaherpesvirus. J. Vet. Diagn. Invest. 30, 663–670 (2018).
Hongwei, Z. et al. Evidence of two genetically different lymphotropic herpesviruses present among red deer, Sambar, and Milu herds in China. J. Vet. Sci. 19, 716–720 (2018).
Javier, S. R., Torill, K. S. K. K. L. A. & Morten, T. M. Ocular histopathological findings in semi-domesticated Eurasian tundra Reindeer (Rangifer tarandus tarandus) with infectious keratoconjunctivitis after experimental inoculation with cervid herpesvirus 2. Viruses 12, 1007 (2020).
Annette, R., Justin, S., Kent, H., Wade, H. & Arnaud, W. Investigation into causes of antler deformities in mule deer (Odocoileus hemionus) bucks in Southern Utah, USA. J. Wildl. Dis. 58, 222–227 (2021).
Cottingham, S. L., White, Z. S., Wisely, S. M. & Campos-Krauer, J. M. A. Mortality-based description of EHDV and BTV prevalence in farmed White-Tailed deer (Odocoileus virginianus) in Florida. USA Viruses 13, 1443 (2021).
Rodrigo, S., Ezequiel, H. H. & José, P. L. Detection of persistent pestivirus infection in Pudú (Pudu puda) in a captive population of artiodactyls in Chile. BMC Vet. Res. 14, 37 (2018).
Li, Y. et al. Diversity and independent evolutionary profiling of rodent-borne viruses in Hainan, a tropical Island of China. Virol. Sin. 38, 651–662 (2023).
Andreia, G., Isabel, P., Federica, S., Alessandra, S. & Laura, G. Cutaneous fibropapilloma in a red deer (Cervus elaphus) associated with Cervus elaphus papillomavirus in Portugal. J. Wildl. Dis. 56, 636–639 (2020).
Gallina, L. et al. Epitheliotropic infections in wildlife ruminants from the central alps and Stelvio National park. Front. Vet. Sci. 7, 229 (2020).
Scagliarini, A. et al. Cervus elaphus papillomavirus (CePV1): new insights on viral evolution in deer. Vet. Microbiol. 165, 252–259 (2013).
Fux, R. et al. Rusa alfredi papillomavirus 1—a novel deltapapillomavirus inducing endemic papillomatosis in the endangered Visayan spotted deer. J. Gen. Virol. 97, 128–133 (2016).
Kmetec, J., Kuhar, U., Fajfar, A. G., Vengust, D. Z. & Vengust, G. A. Comprehensive atudy of cutaneous fibropapillomatosis in free-ranging Roe deer (Capreolus capreolus) and Red deer (Cervus elaphus): from clinical manifestations to whole-genome sequencing of papillomaviruses. Viruses 12, 1001 (2020).
Erdélyi, K. et al. Endemic papillomavirus infection of roe deer (Capreolus capreolus). Vet. Microbiol. 138, 20–26 (2009).
Donlin, M. J. et al. Identification and characterization of two novel viruses in ocular infections in reindeer. PLoS ONE. 8, e69711 (2013).
Mengual-Chuliá, B., Wibbelt, G., Gottschling, M. & Bravo, I. G. Two novel, distantly related papillomaviruses isolated from healthy skin of the Timor deer (Rusa timorensis). Genome Announc. 6, e00505–e00518 (2018).
Munday, J. S., Vaatstra, B. L., Dunowska, M., Laurie, R. E. & Hills, S. Genomic characterization of a novel epsilonpapillomavirus associated with pigmented papillomas in a red deer (Cervus elaphus). Virus Genes 52, 633–639 (2016).
Schürch, A. C. et al. Metagenomic survey for viruses in Western Arctic Caribou, Alaska, through iterative assembly of taxonomic units. PLoS One 9, e105227 (2014).
Varsani, A. & Krupovic, M. Smacoviridae: a new family of animal-associated single-stranded DNA viruses. Arch. Virol. 163, 2005–2015 (2018).
Ladner, J. T. et al. A multicomponent animal virus isolated from mosquitoes. Cell. Host Microbe 20, 357–367 (2016).
Kazlauskas, D., Varsani, A., Koonin, E. V. & Krupovic, M. Multiple origins of prokaryotic and eukaryotic single-stranded DNA viruses from bacterial and archaeal plasmids. Nat. Commun. 10, 3425 (2019).
Zhao, L., Rosario, K., Breitbart, M. & Duffy, S. Eukaryotic circular Rep-Encoding Single-Stranded DNA (CRESS DNA) viruses: ubiquitous viruses with small genomes and a diverse host range. Adv. Virus Res. 103, 71–133 (2019).
Liu, X. et al. Emergence of Porcine circovirus-like viruses associated with Porcine diarrheal disease in China. Transbound. Emerg. Dis. 68, 3167–3173 (2021).
Chengyuan, J. et al. Circular replication-associated protein-encoding single-stranded DNA virus with risk of spillover is widely prevalent in domestic animals in China. Virus Res. 339, 199204 (2023).
Tongtong, W. et al. First detection and phylogenetic analysis of Porcine circovirus 3 in female donkeys with reproductive disorders. BMC Vet. Res. 17, 308 (2021).
Kerry, G. et al. Detection and complete genome analysis of Porcine circovirus 2 (PCV2) and an unclassified CRESS DNA virus from diarrheic pigs in the Dominican Republic: first evidence for predominance of PCV2d from the Caribbean region. Viruses 14, 1799 (2022).
Xifeng, H. et al. Frequency detection of Porcine circovirus-like viruses in pigs with Porcine respiratory disease. Vet. Microbiol. 275, 109581 (2022).
Agm, R. & Sheela, R. Comparative Immunopathogenesis and biology of recently discovered Porcine circoviruses. Transbound. Emerg. Dis. 68, 2957–2968 (2021).
Fehér, E. et al. Genomic Diversity of CRESS DNA Viruses in the Eukaryotic Virome of Swine Feces. Microorganisms 2021, 9 (2021).
Jiamin, P. et al. Metagenomic analysis of herbivorous mammalian viral communities in the Northwest plateau. BMC Genom. 24, 568–568 (2023).
Jun, J. et al. Molecular characterization of the Gyrovirus Galga 1 strain detected in various zoo animals: the first report from China. Microbes Infect. 24, 104983 (2022).
Liu, Q. et al. Viral metagenomics revealed diverse CRESS-DNA virus genomes in faeces of forest musk deer. Virol. J. 17, 61 (2020).
Molly, K., Gun, T. & E, P. P. Five years of Porcine circovirus 3: what have we learned about the clinical disease, immune pathogenesis, and diagnosis. Virus Res. 314, 198764 (2022).
Franzo, G. et al. A wild circulation: high presence of Porcine circovirus 3 in different mammalian wild hosts and ticks. Transbound. Emerg. Dis. 66, 1548–1557 (2019).
Fisher, M. et al. Discovery and comparative genomic analysis of elk circovirus (ElkCV), a novel circovirus species and the first reported from a Cervid host. Sci. Rep. 10, 19548 (2020).
Steel, O. et al. Circular replication-associated protein encoding DNA viruses identified in the faecal matter of various animals in new Zealand. Infect. Genet. Evol. 43, 151–164 (2016).
Czyżewska-Dors, E. et al. Detection of Porcine circovirus 3 in wildlife species in Spain. Pathogens 9, 341 (2020).
Roper, R. L. et al. Putative parapoxvirus-associated foot disease in the endangered Huemul deer (Hippocamelus bisulcus) in Bernardo O’Higgins National park, Chile. Plos One 14, e0213667 (2019).
Tang, J. et al. Transcriptome profiling of lung immune responses potentially related to acute respiratory distress syndrome in forest musk deer. BMC Genom. 23, 701 (2022).
Araujo, J. P., Nogueira, M. F. & Duarte, J. M. B. Survey for foot-and-mouth disease in the endangered marsh deer (Blastocerus dichotomus) from marshlands of the Parana river basin, Brazil. J. Wildl. Dis. 46, 939–943 (2010).
Chaiwattanarungruengpaisan, S. et al. Serologic evidence of pandemic (H1N1) 2009 virus infection in camel and Eld’s deer, Thailand. Vet. World 14, 2596–2601 (2021).
López-Bueno, A. et al. Concurrence of iridovirus, polyomavirus, and a unique member of a new group of fish papillomaviruses in lymphocystis Disease-Affected Gilthead sea Bream. J. Virol. 90, 8768–8779 (2016).
dos Reis, M. et al. Uncertainty in the timing of origin of animals and the limits of precision in molecular timescales. Curr. Biol. 25, 2939–2950 (2015).
Marta, P. et al. Ovine papillomaviruses: diversity, pathogenicity, and evolution. Vet. Microbiol. 289, 109955 (2023).
Mengual-Chuliá, B., Wittstatt, U., Olias, P. & Bravo, I. G. Genome sequences of two novel papillomaviruses isolated from healthy skin of Pudu puda and Cervus elaphus deer. Genome Announc. 6, e00298–e00218 (2018).
Gysens, L., Vanmechelen, B., Maes, P., Martens, A. & Haspeslagh, M. Complete genomic characterization of bovine papillomavirus type 1 and 2 strains infers ongoing cross-species transmission between cattle and horses. Vet. J. 298-299, 106011 (2023).
Roperto, S., Cutarelli, A., Corrado, F., De Falco, F. & Buonavoglia, C. Detection and quantification of bovine papillomavirus DNA by digital droplet PCR in sheep blood. Sci. Rep. 11, 10292 (2021).
Falco, F. D. et al. Evidence of a novel cross-species transmission by ovine papillomaviruses. Transbound. Emerg. Dis. 69, 3850–3857 (2022).
Van Doorslaer, K. et al. Unique genome organization of non-mammalian papillomaviruses provides insights into the evolution of viral early proteins. Virus Evol. 3, vex027 (2017).
Despres, H. W. et al. Surveillance of Vermont wildlife in 2021–2022 reveals no detected SARS-CoV-2 viral RNA. Sci. Rep. 13, 14683 (2023).
Ernst, S. et al. Prion protein gene (PRNP) variation in German and Danish cervids. Vet. Res. 55, 98 (2024).
Ji, C. et al. Circular replication-associated protein-encoding single-stranded DNA virus with risk of spillover is widely prevalent in domestic animals in China. Virus Res. 339, 199204 (2024).
Gao, Y. Y. et al. Genomic composition and pathomechanisms of Porcine circoviruses: a review. Virulence 15, 2439524 (2024).
Wu, Z. et al. Detection of hantaviruses and arenaviruzses in three-toed Jerboas from the inner Mongolia autonomous region, China. Emerg. Microbes Infect. 7, 35 (2018).
Wang, G. et al. Identification and genome analysis of a novel picornavirus from captive Belugas (Delphinapterus leucas) in China. Sci. Rep. 11, 21018 (2021).
Yang, J. et al. Unbiased parallel detection of viral pathogens in clinical samples by use of a metagenomic approach. J. Clin. Microbiol. 49, 3463–3469 (2011).
Acknowledgements
We would like to extend our gratitude to all the institution and individuals who participate and provided their kind assistance, especially generous permission and collaboration in the sample collection process from the Hainan Bangxi Provincial Nature Reserve Administration and the Hainan Tropical Infectious Diseases Biobank.
Funding
This work was supported by the Key Research and Development Plan of Hainan Province (No. ZDYF2022SHFZ085), Hainan Provincial Natural Science Foundation of China (No. 822RC695), National Natural Science Foundation of China (Nos. 32060015 and 82060378), Hainan Medical University Talent Development Project (No. XRC2021002), Science and Technology Planning Project of Guizhou Province (No. qkhjc-zk2022-yb435), Gradutae student innovation grant of Hainan Medical University (No. HYYS2020-33), Research Project of the Hainan Academician Innovation Platform (No. YSPTZX202004), and Hainan Talent Development Project (No. SRC200003).
Author information
Authors and Affiliations
Contributions
Y.Z., G.L. and Y.L. (Youyou Li) designed the study. Y.Z., X.Q., Y.C., Q.L., G.R., R.P., G.W., X.H. and Y.H. collected the specimens. H.S., Z.L., J.D., M.Z. and X.T., performed the experiments. D.Z., J.D., Y.L. (Youyou Li), C.T. (Chuanning Tang), X.C., L.N., J.Y., F.Y., G.L., J.L., Y.L. (Yu Li) and C.T. (Chuan Tian) analysed the data. Y.L. (Youyou Li) wrote the draft manuscript. D.Z., F.Y. and Y.Z. edited the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
The sampling strategy in this study did not involve hunting or euthanized method. During the translocation process in the nature reserves where Eld’s deer live, we collected throat swabs, anal swabs, and swab samples from tick bite sites. After sample collecting, all Eld’s deer were released on-site as soon as possible. We have no experiments on live vertebrates, but only identified the microorganisms in swab samples. All methods were carried out in accordance with relevant guidelines and regulations of the Ethics Committee of the Hainan Medical University (approval no. HMUEC20180059).
Ethical statement
In this study, there was no experiments in vivo, and all methods and procedures were conducted in accordance and compliance with the ARRIVE guidelines (https://arriveguidelines.org).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Su, H., Zheng, D., Li, Z. et al. The virome investigation of the globally endangered Eld’s deer (Rucervus eldii) on Hainan Island, China. Sci Rep 15, 8384 (2025). https://doi.org/10.1038/s41598-025-92781-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-92781-4
