
Abstract
Motor sequence learning involves both oculomotor and manual motor systems, yet the role of the oculomotor system in the learning and execution of skilled arm movements remains underexplored. In the current work, the influence of sequence learning on the oculomotor system was investigated by testing 20 healthy adults for 3 days as they practiced an implicit motor learning task, the serial targeting task (STT). The STT contained a repeated sequence, which was interleaved with random sequences. This task was practiced on a KINARM robot that tracked both saccades and reaches. A delayed, 24-h retention test assessed sequence-specific motor learning. Sequence-specific changes across practice and learning were observed for both saccades and reaches; this was demonstrated by faster saccade and arm motor reaction times for the repeated sequence compared to random sequences. Notably, change in the oculomotor system occurred earlier in practice as compared to the manual motor system. Reaches were executed more quickly when led by express saccades (rapid eye movements occurring within 90–120 ms) compared to when they were preceded by regular latency (> 120 ms) saccades early in practice. Our findings highlight distinct yet interconnected functions between oculomotor and manual motor systems associated with implicit motor sequence learning.
Similar content being viewed by others


Introduction
Motor sequence learning is a process wherein the performance of repeated sequences of movements become automatized through practice. While research has elucidated the neural substrates and behavioural manifestations of motor sequence learning1,2,3, investigation of oculomotor behaviour in this context remains limited. The development of long-term memory can be stimulated by different inputs, leading to different types of learning, including explicit and implicit learning4,5,6. Implicit learning refers to the capacity to acquire skills through physical practice without direct conscious recollection that learning has occurred. In contrast, explicit learning involves the conscious memory of facts, events, and episodes, which can be directly assessed through recall or recognition tests7,8,9,10. Motor sequence learning tasks often involve manual motor responses, with performance measured by hand reaction times, and are typically designed to evaluate limb motor systems2,8,11,12,13. Implicit motor sequence learning can be tested by experimental paradigms that contain repeated and random sequences of movement. In these experiments, a repeated sequence is typically bracketed by random sequences. As the two types of sequences are not marked, participants may or may not become aware of their existence. With practice, participants show improvement on both types of sequences. However, since random sequences cannot be learned, improvements in their performance reflect changes in motor control. In contrast, repeated sequences can be learned, and implicit motor sequence learning is assessed following practice at a delayed retention test. This is done by comparing the magnitude of change in repeated sequences to that in random sequences. Differences between random and repeated sequences index the magnitude of implicit sequence-specific motor learning.
During motor sequence learning, saccades are typically triggered in response to the sudden appearance of a visual stimulus. Further, it is well-known that arm reaches to a target are led by saccadic eye movements14,15. Given the important role of saccades during sequential movement, implicit motor sequence learning likely extends beyond the manual motor system16,17,18,19 to the oculomotor system. Human motor control literature on eye-hand coordination demonstrates that these systems are indeed interconnected and their coordination may facilitate motor function. For example, reaching movements are more accurate when they are preceded by eye movements toward the same target20,21,22, and fixations on specific landmarks support hand movement planning by marking key positions for successful object manipulation23. Similarly, during tasks requiring novel visuomotor mappings, such as controlling a cursor using bimanual forces, changes in hand movement performance are accompanied by behavioural changes in eye movements24. Together, these findings suggest that motor sequence learning may extend beyond the manual motor system to include the oculomotor system.
Human oculomotor studies of implicit motor sequence learning provide some insight into the oculomotor system’s role in sequence learning; however they are sparse25,26,27,28. Marcus et al.25 showed that as hand reaction times improved with repetitive sequence practice, the percentage of anticipatory saccades also increased, suggesting a parallel improvement in predictive performance in these systems. However, performance was examined in a single session without a retention test, as such no conclusions can be drawn about oculomotor sequence learning that might occur during motor sequence learning. Another study of implicit oculomotor sequence learning (without manual movements) showed that saccade reaction times were reduced during practice, and performance was significantly enhanced after a 24-h time-delay (but not 30-min or 5-h after practice)29. While the authors highlighted some similarities between the oculomotor and manual motor systems, namely the time course of off-line performance improvements up to 24 h after training, the task involved only oculomotor movements. Thus, conclusions about the role of the oculomotor system in implicit motor sequence learning cannot be made. Recent studies also examined oculomotor performance using a type of serial reaction time task (similar to the original described by Nissen and Bullemer2), this time, together with manual motor responses26,27,28. However, performance was not investigated beyond a single session and therefore sequence-specific learning effects were not tested. To date, a comprehensive assessment of human oculomotor control in motor sequence learning remains unexplored, particularly regarding the retention of sequence-specific knowledge in the oculomotor system and its interaction with the manual motor system beyond a single session of practice.
In the current study, we investigated the effects of practice of an implicit motor sequence task on both oculomotor and manual motor systems. Importantly, we tested these effects across 3 days of practice and at a delayed retention test, allowing us to distinguish improvements related to practice from more lasting changes associated with motor sequence learning30. We hypothesized that 3 days of practice would result in: (1) sequence-specific learning for both saccades and arm reaches, evidenced by faster reaction times at a delayed retention test for the repeated sequence as compared to random sequences, (2) sequence-specific differences in implicit motor learning shown earlier in practice for saccades as compared to arm reaches, and (3) correlations between behavioural changes in saccades and reaches during practice and at the retention test. Additionally, we explored whether gaining explicit awareness of the repeated sequence through practice would influence either oculomotor or manual motor responses.
Methods
Participants
Twenty-four, healthy adults were recruited for this study (all right-handed and between the ages of 19 and 40). Individuals were excluded from participation for: (1) visual acuity < 20/60 (as determined using a Snellen Visual Acuity Chart), and (2) a history of head trauma, major psychiatric diagnosis, neurodegenerative disorder, visual disorders or substance abuse that affects visuomotor task performance. Four participants did not complete the study (due to personal reasons), leaving twenty who finished all testing sessions. The Research Ethics Board at the University of British Columbia approved all study procedures. All testing was conducted at the University of British Columbia. Informed consent from each participant was obtained prior to partaking in this study, in accordance with the principles of the Declaration of Helsinki.
Apparatus
Participants were seated in a chair and instructed to use their non-dominant (left) hand to grasp the handle of a robotic manipulandum that could move in a horizontal plane of a two-dimensional augmented reality environment (KINARM End-Point Laboratory, KINARM, Kingston, Ontario, Canada). Visual stimuli were projected at 60 Hz onto a semitransparent mirror from a monitor positioned above the workspace. This set-up allowed the stimuli to appear on the same horizontal plane as the handle while occluding the participant’s view of the hand (Fig. 1A). During task performance, hand position was sampled at 1000 Hz. Monocular (left eye) position of each participant was recorded at 500 Hz using a video-based, head-free remote eye-tracking system (Eyelink1000, SR Research, Ottawa, Ontario, Canada) integrated within the KINARM system.

Experimental procedure and task. The STT was practiced in the KINARM over three consecutive days (A). Participants saw only two targets at a time, the target they were currently positioned on, and the target they were required to move to. Participants’ view of their hand was occluded, and they controlled a white cursor to reach to the new target (B). The 8-element repeated sequence followed a unique pattern; numbers and grey circles depicted are for visualization purposes only (C).
Experimental design ands procedure
Participants practiced an implicit motor sequence learning task over 3 consecutive days (practice sessions were 24 h apart), followed by a delayed retention test administered 24 h after practice on day 331,32,33,34 (Fig. 1A). Eye position was calibrated at the beginning of each session, using the built-in 9-point calibration protocol. Eye position was monitored in real-time during all testing sessions, if the quality of the calibration diminished or the camera lost track of the pupil altogether, calibration was repeated between testing blocks. Before practice on day 1, a pre-test was completed to allow participants to become familiar with task demands. Immediately following the retention test, recognition tests were completed to determine whether participants gained explicit awareness of the repeated sequence during practice (described in the “Explicit awareness” section).
During testing, participants performed discrete whole-arm reaching movements in which they were instructed to move a cursor (white circle, 1 cm diameter; representing their veridical hand position) to the location of a suddenly appearing peripheral visual stimulus (yellow circle, 2 cm diameter) as quickly and accurately as possible. As soon as participants moved the cursor away from the current target (‘Start target’; Fig. 1B), it disappeared from the screen. Thus, during arm movement trajectory, participants could only see the target they were required to reach to (‘End target’; Fig. 1B). After the peripheral target was reached, visual feedback was provided to signal a successful reach (the stimulus changed from yellow to orange) and participants were required to hold their position for 500 ms, after which the next target appeared (Fig. 1B). Participants’ head movements were not restrained during testing due to the camera’s ability to compensate for head movements, however, participants were encouraged to rest their forehead on the KINARM head rest.
Each day consisted of 4 blocks of practice, lasting approximately 16 min total. Each block consisted of 153 trials where the 8-element repeated sequence appeared 8 times, which were flanked by 8-element random sequences. In total, participants performed 612 trials or reaches per day. The duration of each day was dependent on individual performance, however on average one block lasted approximately 4 min. Breaks between blocks were provided as needed. On practice day 1, participants were briefly exposed to the task which contained 27 trials; this pre-test took approximately 30 s and allowed participants to become familiar with the KINARM environment and task demands. The retention test contained 1 block of 153 trials; this block had the same characteristics of a practice block (see next section for details). Testing was conducted in a well-lit room and sessions were scheduled at roughly the same time of day for each participant (either morning, afternoon, or evening).
Implicit motor sequence learning task
The task was adapted from the traditional serial reaction time task2, and modified for the KINARM robot such that reaching movements instead of key-presses are used to respond to visual stimuli. The serial targeting task (STT) contains repeated and random sequences32,33,35,36. Unbeknownst to the participants, an 8-element sequence (Fig. 1C) was repeated which was flanked by 8-element random sequences, where targets appeared in random locations. This design was employed to separate performance or general practice-related effects from motor learning30. Typically, participants improve performance for both types of sequences. During practice, changes in repeated sequence performance reflect improvements in both motor learning and motor control. In contrast, the random sequences cannot be learned, as such changes during practice of these movements reflect only improved motor control. Because participants are not explicitly informed of the existence of the repeating sequence, difference between random and repeated sequences at a retention test reflects implicit sequence-specific motor learning.
During the STT, targets could appear in one of nine locations, eight peripheral targets forming an equidistant circular array (8 cm apart) with one target at its center (10.5 cm from peripheral targets) (Fig. 1B). The 8-element repeated sequence consisted of the following arrangement of targets: 8–9–6–7–4–8–2–6–3, with the number referring to its position on the screen (Fig. 1C). During practice, the 8-element repeated sequence appeared 8 times per block, and the 8-element random sequences appeared 9 times per block; participants always started and ended each block with a random sequence. The random sequences were created a priori to ensure they closely matched the repeated sequence in difficulty such that: (1) number of targets, and (2) average path length (Euclidean distance) in each sequence were matched. Further, to ensure the repeated and random sequences did not have easily recognizable patterns, we excluded familiar target configurations, such as alternating targets (i.e., 1–2–1; sequences of three or more targets that alternated between two locations), and the first target of each random sequence within a block started at a different spatial location.
Explicit awareness
A recognition test of repeated sequence composition was administered following the retention test to assess whether explicit awareness of the repeated sequence was gained. Recognition was assessed by presenting participants with 10 sequences played on the KINARM screen and asking whether they recognized each one. The test included 3 true sequences (same as the repeated sequence practiced during training) and 7 foil sequences. Participants responded verbally with “yes” or “no” to indicate recognition. All participants saw the same order of sequences, which was randomized a priori. To show recognition of the repeated sequence, participants had to correctly identify both of the repeated and random sequences at better than chance; 2 of 3 of the true sequences as having been seen before, and 4 of 7 foils as not having been seen37,38. Recognition test findings were used to group participants post-hoc into ‘explicit’ versus ‘implicit’ learners for our exploratory analyses of the effects of explicit awareness on oculomotor and manual motor responses.
Data analyses
Data were recorded using integrated KINARM Dexterit-E and SR Research software. Behavioural measures were extracted using custom MATLAB scripts, and all statistical analyses were performed in R/RStudio (v.4.3.3). Parametric statistical testing was conducted unless otherwise stated.
Gaze pre-processing and gaze-event identification
Gaze data were low-pass filtered at 20 Hz and preprocessed to remove blinks, one-sample spikes (due to incorrect detection of corneal reflection), and screen outliers (due to instances when gaze drifted outside of the workspace). Gaze angular kinematics were computed from filtered and artifact-free XY gaze position39. Saccades were identified using fixed velocity and acceleration thresholds. For each velocity peak that exceeded the threshold (20 deg/s), the peak acceleration leading up to the velocity peak had to also exceed the acceleration threshold (6000 deg2/s)40. If both thresholds were exceeded for at least 30 ms, the gaze event was classified as a saccade. If threshold criteria were not satisfied, then a saccade was not recorded for that trial.
Outcome measures
The primary dependent measure was reaction time (RT), calculated separately for saccades (‘saccade RT’) and reaches (‘reach RT’). RT was defined as the interval between stimulus onset and the onset of movement, recorded for each trial. The median RT for each repeated and random sequence was calculated to reduce the sensitivity of this measure to very large or very small values2,11,19,41,42,43,43,44. The mean of the median RT for each sequence (repeated or random) within each block was calculated. To facilitate comparison of sequence-specific practice effects across saccades and reaches, a change score was calculated as follows: RT change score = mean of the mean RT of all repeated sequences minus mean of the mean RT of random sequences. A change score for each participant was calculated for the first block of each training day to enable an accurate comparison with data from the retention block (data that did not contain impact of same day practice). A larger negative change score indicates greater change in RT for the repeated sequence relative to random sequences. Literature-defined cut-offs for saccade latencies were used to distinguish express saccades (90–120 ms) from regular latency, visually-guided saccades (> 120 ms)45; these data enabled us to examine relationships between saccade type and reach behaviour.
Data cleaning
The first trial of every block was removed (as is typical for reaching tasks; slower response is characteristic of inattention and does not accurately reflect motor control). Individual reach trials were discarded if RT was > 1000 ms or < 100 ms. For saccades that met threshold criteria, the primary saccade (first saccade after target onset) was considered in each trial. A valid trial for the task could include: (1) a reach (between 100 and 1000 ms), or (2) a saccade accompanied by a valid reach.
Statistical analyses
Sequence-specific motor learning
To determine sequence-specific implicit motor learning of the repeated sequence, paired t-tests separately assessed saccades and reaches using mean RT from repeated and random sequences at retention.
Patterns of change during practice
Data from practice were evaluated using linear mixed-effects (LME) models with mean RT as the dependent variable, sequence type (repeated vs. random) and day (Days 1–3) included as fixed effects, and participants as a random effect to account for individual variability. Separate models were conducted to assess practice data for saccades and reaches. Additionally, to determine whether explicit awareness affected performance, learner type (explicit vs. implicit) was added as an additional factor in the LME models.
Dose of practice required for sequence-specific changes
LME models were used to assess reaction time differences by sequence type across individual training blocks, to identify when performance for the repeated sequence became significantly faster than for the random sequences, indicating amount of practice needed to provoke sequence-specific differences. Twelve separate LME models were conducted, separately for saccades and reaches, with participant as a random effect. Results were corrected for multiple comparisons by adjusting the significance level to p < 0.004 (0.05/12).
Relationship between saccades and reaches:
Correlations between RT change scores examined the relationship between saccades and reaches during practice and at the 24-h retention test. To minimize the number of comparisons and ensure a more accurate comparison to the retention test, only the first block of each training day was included in the correlation analyses. Additionally, one-way ANOVAs were conducted to determine if reaches were faster if preceded by an express (90–120 ms) saccade compared to a regular latency (> 120 ms) saccade.
Statistical assumptions were checked for each test. Data normality was confirmed using Shapiro–Wilk tests, and visualized using Q–Q plots, boxplots and histograms. The alpha level for all tests was set at 0.05 unless otherwise stated.
Results
Twenty participants completed this study (15 female; mean age = 28 ± 5; age range 20–37 years). However, data from 19 participants were included in final analyses, and only 17 participants were included in retention test analyses. One participant was excluded altogether due to a significant amount of data loss resulting from issues with eye-tracking. Additionally, retention test data from two participants were excluded: one participant completed the retention test more than 24 h after the last practice session, and one had technical issues with the eye-tracker. In accordance with the minimal reporting guideline for research involving eye-tracking46, data loss was as follows: A total of 1046 reaching trials (2.5% of total trials) were considered outliers (RT > 1000 ms or < 100 ms) and removed from analyses. Following removal of saccade trials affected by technical issues with the eye-tracker and those that did not meet pre-processing criteria, saccade data were retained for 79% of valid reaching trials. Based on saccadic latency cut-off criteria, there were a total of 632 express saccades (1.6% of all saccade trials) and 35,213 regular saccades (91.8% of all saccade trials). All participants demonstrated high accuracy in their reaching movements; the average Euclidean distance between reach end-point and target position was 0.43 cm (SD = 0.09 cm). Post-hoc analysis of explicit awareness indicated that 42% of participants (8 out of 19 with practice data) demonstrated recognition of the repeated sequence by correctly identifying both the repeated and random sequences at levels above chance; the other participants did not meet these criteria and were deemed as having only implicit awareness of the repeated sequence.
Sequence-specific implicit motor learning
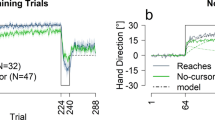
Sequence-specific implicit motor learning for saccades was demonstrated by a difference in mean saccade RT by sequence type (repeated vs. random), confirming that participants were faster for repeated compared to random sequences, as assessed at the 24-h retention test (ΔM = − 6.57 ms, 95% CI [− 10.23, − 2.91], t(16) = − 3.8, p = 0.0015, Cohen’s d = − 0.92). Similarly, sequence-specific implicit motor learning for reaches was shown by a difference in mean reach RT by sequence type (repeated vs. random), confirming that participants were also faster at executing reaches for repeated compared to random sequences at retention (ΔM = − 9.08 ms, 95% CI [− 14.18, − 3.98], t(16) = − 3.77, p = 0.0017, Cohen’s d = − 0.91). Data disaggregated by block, including the retention test block (“RET”), are plotted in Fig. 2.

Practice and retention test data. Plots show raw RT means with standard error of the mean for each block, for saccades (A) and reaches (B). Lines connect blocks of practice conducted within each testing session. RET: 24-h retention block.
Patterns of change during practice
LME models were used to investigate the effects of sequence type (random vs. repeated) and training day (Days 1, 2, and 3) on mean reach and mean saccade RTs, with a random intercept for each participant. Two sets of models were tested: one including only the main effects of sequence and day, and another including the interaction term (sequence × day) to assess whether sequence effects differed across days. The models were compared using a likelihood ratio test and Akaike Information Criterion (AIC) to evaluate whether adding the interaction term significantly improved model fit, and an ANOVA confirmed the significance of the fixed effects in the final model. For each model, the residuals were visually inspected using a histogram and Q–Q plot and appeared approximately normal, despite a significant Shapiro–Wilk test (p < 0.05). As the linear mixed-effects model is robust to minor deviations from normality, the analysis was deemed appropriate.
The inclusion of interaction terms in the models did not significantly improve model fit (saccades: χ2(2) = 1.7, p = 0.44; reaches: χ2(2) = 0.16, p = 0.93). Furthermore, the simpler models (main effects only) had a lower AIC compared to the interaction models (saccades: 3660.5 vs. 3662.9; reaches: = 4162.8 vs. 4166.6). Therefore, the simpler models were retained for interpretation. Both reach RT and saccade RT models, fitted using a restricted maximum likelihood (REML), revealed significant main effects of sequence and day. The model for mean reach RT showed that RTs were faster for repeated sequences compared to random sequences (Estimate = − 6.89 ms, SE = 1.96, t(434) = − 3.52, p = 0.0005). Both day 2 (Estimate = − 38.56 ms, SE = 2.4, t(434) = − 16.07, p < 0.001) and day 3 (Estimate = − 51.56 ms, SE = 2.4, t(434) = − 21.49, p < 0.001) were associated with significantly faster RTs compared to day 1. The model had a REML criterion of 4134.3, with the variance of the random intercept for participant estimated at 1818.8 (SD = 42.65), and the residual variance at 437.6 (SD = 20.92).
The model for mean saccade RT similarly showed that RTs were faster for repeated sequences compared to random sequences (Estimate = − 3.69 ms, SE = 1.13, t(434) = − 3.26, p = 0.001). Day 2 (Estimate = − 6.36 ms, SE = 1.39, t(434) = − 4.59, p < 0.001) and day 3 (Estimate = − 9.49 ms, SE = 1.39, t(434) = − 6.84, p < 0.001) showed significantly faster RTs compared to day 1. The model had a REML criterion of 3636.6, and the variance of the random intercept for participant was 533.5 (SD = 23.1), and the residual variance was 146.2 (SD = 12.09). Estimated marginal means for mean reach and saccade RT data by sequence and day are plotted in Fig. 3A and B.

Practice data illustrating differences in sequence type by day or block. Plots show estimated marginal means (EMMs) with 95% confidence intervals, as derived from the linear mixed-effects models, reflecting model-based differences in RT between random and repeated sequences across days (A,B), and across blocks for saccades (C) and reaches (D). Asterisks in plots C and D indicate statistically significant difference between sequence types, marking the first instance of sequence-specific change for saccades versus reaches.
Effect of explicit awareness
To examine whether learner type (explicit awareness vs. implicit) influenced saccade and reach RTs, expanded LME models including learner type as a fixed factor were tested. Adding learner type did not improve model fit for reaches (χ2(1) = 0.47, p = 0.49; AIC: 4162.8 vs. 4164.3) or saccades (χ2(1) = 0.49, p = 0.48; AIC: 3660.5 vs. 3662.0). The main effect of learner type was non-significant in both models (reaches: Estimate = 13.24 ms, p = 0.52; saccades: Estimate = − 7.29 ms, p = 0.51), indicating there was no effect of explicit awareness on implicit motor sequence practice.
Dose of practice required for sequence-specific changes
To investigate the amount of practice needed to provoke sequence-specific differences, separate LME models were conducted for each individual training block to examine the effects of sequence type (random vs. repeated) on mean reach and mean saccade RTs, with a random intercept for each participant. After correcting for multiple comparisons, LME models for mean saccade RTs revealed that participants were significantly faster on the repeated sequence compared to the random sequences starting at Block 7 (Estimate = − 5.59 ms, SE = 1.39, t(18) = − 4.01, p = 0.0008). In contrast, reaches showed a significant difference between sequence types at Block 12 (Estimate = − 8.53 ms, SE = 2.3, t(18) = − 3.71, p = 0.0016). Taken together, saccadic responses showed earlier sequence-specific changes as compared to manual responses during practice. Data are plotted in Fig. 3C and D.
Relationship between saccades and reaches
Magnitude of sequence-specific change between saccades and reaches
Pearson correlation analyses were conducted to examine the relationship between saccade and reach RT change scores across days; specifically, the first block of each day was assessed. No significant correlations were observed on Day 1 (r = − 0.012, p = 0.960), Day 2 (r = 0.021, p = 0.932), or at the 24-h retention test (r = 0.362, p = 0.153). However, a significant positive correlation was found on Day 3 (r = 0.689, p = 0.001), with a 95% confidence interval of [0.341, 0.871]; larger RT change scores for saccades were associated with larger RT change scores for reaches. Data were normally distributed, however, following visual inspection of correlation plots, we identified two potential outliers. After outlier removal, the significant correlation on Day 3 remained (r = 0.51, p = 0.03), and the non-significant correlation at retention remained (r = 0.48, p = 0.058). Data are plotted in Fig. 4.

Relationship between saccade and reach RT change scores. RT change scores represent the difference in performance between repeated and random sequences; a larger negative change score indicates greater change in RT for the repeated sequence relative to random sequences. Each dot represents a participant, demonstrating the relationship between saccade and reach RT change scores. Data from the first block of each practice day only were included in the analysis.
Effect of saccade type on reach behaviour
Separate one-way ANOVAs were conducted to examine differences in RT between reaches that were preceded by express (90–120 ms) compared to regular latency (> 120 ms) saccades. Data were examined across all trials, irrespective of sequence type. Reaction time significantly differed between saccade type on Days 1, 2, and 3, but not at the retention test. Welch’s one-way ANOVAs (not assuming equal variances) revealed that the express saccade group had significantly faster reaches than the regular latency saccade group on Day 1 (F(1, 212.57) = 10.87, p = 0.001, Δ M = 19.11 ms) and Day 2 (F(1, 199.83) = 3.97, p = 0.048, Δ M = 12.67 ms). On Day 3, however, the opposite effect was observed, where reaches were significantly slower when preceded by express saccades compared to regular latency saccades (F(1, 168.83) = 4.96, p = 0.027, Δ M = − 13.73 ms). No significant differences were found at retention (F(1, 42.39) = 0.033, p = 0.857, Δ M = 2.11 ms). Data are plotted in Fig. 5.

Relationship between saccade type and reach responses. Mean reach RT of all trials in which an express (90–120 ms) or regular latency (> 120 ms) saccade preceded the reach. Data are averaged over blocks for each practice day. Error bars represent standard error of the mean.
Discussion
This study investigated how sequence-specific implicit motor learning is supported by both the oculomotor and manual motor systems. Participants demonstrated implicit sequence-specific motor learning in both oculomotor and manual motor systems, as shown by faster reaction times for repeated sequences compared to random sequences at the 24-h retention test. These findings confirm implicit motor learning-related effects in both systems. Additionally, saccades showed change between repeated versus random sequences earlier in practice suggesting that implicit motor learning required less practice for the oculomotor as compared to the manual motor system. While sequence-specific learning was evident in both oculomotor and manual motor systems, we note that the magnitude of sequence-specific motor learning (shown by differences between repeated and random sequence RTs) was not correlated between systems at the 24-h retention test. While this might suggest that learning-related effects are not consistent across systems, our data also showed that reaches were typically faster to respond to stimuli when preceded by faster saccades. These findings suggest that while both systems might be learning independently, on a more fundamental level, they are coordinated. Overall, our data highlight the distinct yet interconnected involvement of the oculomotor and manual motor systems in implicit motor sequence learning. We present novel evidence that oculomotor behaviour is modulated over multiple days of motor sequence practice and that the oculomotor system participates in the retention of implicit sequence-specific knowledge when both systems are engaged during sequence practice. These findings provide insight into the development and relationship of behavioural changes across both motor systems, and confirm that sequence learning extends beyond the manual system to the oculomotor system.
Sequence-specific practice changes and motor learning
Our data show that behaviour in both oculomotor and manual motor systems is changed by implicit motor sequence learning, as seen by faster saccades and reaches during the repeated sequence compared to random sequences both across practice and at a delayed retention test. Use of both random and repeated sequences within each practice block allowed us to monitor the evolution of behavioural change within and between blocks, distinguishing improvements in general motor control from those specific to the sequence being learned by each participant. Participants reduced their oculomotor and manual motor RTs for random sequences showing that practice also improves general motor control, a finding that is consistent with prior work47,48. Novel to this study, saccades showed improvements in general motor control, with sequence-specific changes demonstrated throughout practice, and showed implicit motor learning as demonstrated at the retention test. While previous research demonstrated that oculomotor behaviour changes with practice of serial movements25,26, these past studies focused on a single session of practice. Without a delayed retention test, past work did not show implicit motor sequence learning in the oculomotor system. We employed both practice and a 24-h retention test to assess relatively permanent changes in behaviour after a delay (i.e., learning)30,49.
Notably, training led to sequence-specific changes in motor behaviour that was evident earlier in practice for saccades compared to reaches, suggesting that the oculomotor system may be acquiring implicit knowledge of the repeated sequence prior to the manual motor system. While it was not extensively investigated here, our data imply that the oculomotor system may facilitate learning in the manual motor system. Vieluf et al.50 provide supporting evidence for this idea, showing that allowing eye movements during motor sequence practice led to faster manual responses compared to participants who had to maintain fixation. This suggests that oculomotor engagement aids in extracting spatial and sequential information to enable faster motor sequence responses. However, Vieluf et al. could not directly address whether permitting eye movements specifically enhances motor sequence learning or merely supports general motor performance improvements. This is because the authors did not include a separate group or condition to contrast overall motor control improvements from sequence-specific motor learning. However, their findings strengthen the notion that the oculomotor system plays a functional role in guiding motor learning processes. This also aligns with Sailor et al.24, who demonstrated that eye-hand coordination evolves throughout learning of a novel visuomotor manipulation task (where participants were required to use bimanual manipulation of an object to control a cursor). They showed that during an initial exploratory stage, gaze follows the hand, followed by a shift to predictive gaze fixations as skill acquisition progresses. Sailor et al.’s findings suggest that early oculomotor strategies may contribute to the development of coordinated motor actions, supporting the idea that the oculomotor system plays a key role in motor sequence learning. Further, Laamerad et al.51 showed that eye movements facilitate the integration of visual information in a motion discrimination task, which can later be retained and utilized even without saccades. This past work aligns with the present study’s observations, suggesting that eye movements contribute to an internal representation that could benefit the manual motor system through mechanisms of visual-spatial integration. While the specific mechanisms underlying this facilitation remain unexplored, expansion of knowledge of these fundamental processes may serve as a basis for future studies that investigate elements of coordinated ocular-motor learning.
Relationship between oculomotor and manual motor systems
Our findings revealed both distinct and interconnected relationships between the oculomotor and manual motor systems during practice and at retention. Early in practice (Days 1 and 2), faster reaches were associated with quicker saccades, suggesting that these systems coordinate to support the acquisition of sequence-specific implicit motor knowledge. Although in the current study we examined behavioural manifestations of saccadic and reaching movements, we cannot infer from our study whether changes in one system cause or facilitate changes in the other. Nonetheless, temporal coordination between saccades and reaches observed here is in line with the eye-hand coordination literature, which commonly shows that the oculomotor and manual motor systems are temporally linked toward a common goal52,53,54. Interestingly, this relationship changed by Day 3, where express saccades were unexpectedly associated with slower reaches. While we might expect a positive relationship between systems during the entirety of implicit motor sequence learning, our findings suggest that this relationship is more dynamic. Sailor et al.24 demonstrated that gaze behaviour drastically evolves across different stages of learning a complex visuomotor manipulation task. The change in relationship observed in our study on Day 3 may suggest a restructuring of coordination strategies, where the early association between faster saccades and reach performance appears to change as implicit motor sequence knowledge evolves, potentially reflecting a shift toward more independent control of manual motor movements.
Further, the observed correlation between saccade and reach magnitude of sequence-specific changes on Day 3, but not earlier in training or during the retention test, underscores the dynamic relationship between these systems. The absence of a relationship during early practice aligns with our findings that sequence-specific implicit motor knowledge was acquired later during practice, and at different time-points for these systems; on Day 2 for saccades (Block 7) and Day 3 for reaches (Block 12). On Day 3, a correlation in the magnitude of sequence-specific changes suggests a convergence of learning processes between these systems. This suggests that while one system may acquire and demonstrate sequence-specific knowledge earlier, the underlying mechanisms of implicit learning may be shared across systems during certain timepoints in practice.
Considerations
While we lost data from three participants for the retention test analyses (due to technical issues and scheduling conflicts), the expected effects in sequence-specific learning were observed. Further, our primary focus was on saccadic latencies, however, predictive saccades (defined as latencies less than 90 ms and saccades occurring prior to target onset) could provide valuable insights into the dynamic changes observed within the oculomotor system during motor sequence learning. Additionally, the method used to group participants into explicit and implicit learners should be considered. Since this classification was done post-hoc, we do not know when explicit awareness developed during training. With this approach, the type of memory processes used by participants is only identifiable after training. However, employing recognition tests after training is a standard practice in implicit motor sequence learning paradigms, as it minimizes the influence of explicit awareness on learners during implicit motor sequence practice. Last, although there was a disproportionate number of data points between groups in the saccade-type analysis (~ 600 express vs. ~ 35,000 regular latency saccades), we still found a significant effect of saccade type on manual motor responses. However, future work may consider implementing different task demands to elicit more fast-latency saccades for further evaluation.
Conclusions
The current study suggests that both oculomotor and manual motor systems are key for implicit motor sequence learning. We demonstrated that the oculomotor system exhibits sequence-specific implicit motor learning following three days of skilled practice. Notably, sequence-specific changes occurred earlier in practice for the oculomotor system as compared to the manual motor system. Additionally, manual responses were generally faster when preceded by express saccades, demonstrating a dynamic relationship between oculomotor and manual motor systems during the implicit motor learning process. These findings underscore the distinct yet interconnected roles of these systems in implicit motor sequence learning.
Data availability
All data will be available from the corresponding author upon reasonable request.
References
Doyon, J., Penhune, V. & Ungerleider, L. G. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychologia 41, 252–262 (2003).
Nissen, M. J. & Bullemer, P. Attentional requirements of learning: Evidence from performance measures. Cogn. Psychol. 19, 1–32 (1987).
Robertson, E. M. The serial reaction time task: Implicit motor skill learning?: Figure 1. J. Neurosci. 27, 10073–10075 (2007).
Squire, L. R. Mechanisms of memory. Science 232, 1612–1619 (1986).
Squire, L. R. Memory and Brain (Oxford University Press, 1987).
Squire, L. R. Declarative and nondeclarative memory: Multiple brain systems supporting learning and memory. J. Cogn. Neurosci. 4, 232–243 (1992).
Boyd, L. A. & Winstein, C. J. Impact of explicit information on implicit motor-sequence learning following middle cerebral artery stroke. Phys. Ther. 83, 976–989 (2003).
Boyd, L. A. & Winstein, C. J. Explicit information interferes with implicit motor learning of both continuous and discrete movement tasks after stroke. J. Neurol. Phys. Ther. 30, 46–57 (2006).
Frensch, P. A., Lin, J. & Buchner, A. Learning versus behavioural expression of the learned: The effects of a secondary tone-counting task on implicit learning in the serial reaction task. Psychol. Res. 61, 83–98 (1998).
Frensch, P. A. One concept, multiple meanings: On how to define the concept of implicit learning. In The Handbook of Implicit Learning, 47–106 (Sage, Thousand Oaks, 1998).
Boyd, L. A. & Winstein, C. J. Providing explicit information disrupts implicit motor learning after basal ganglia stroke. Learn. Mem. 11, 388–396 (2004).
Boyd, L. A. & Winstein, C. J. Cerebellar stroke impairs temporal but not spatial accuracy during implicit motor learning. Neurorehabil. Neural Repair 18, 134–143 (2004).
Shea, C. H., Wulf, G., Whitacre, C. A. & Park, J.-H. Surfing the Implicit Wave. Q. J. Exp. Psychol. Sect. A 54, 841–862 (2001).
Land, M., Mennie, N. & Rusted, J. The roles of vision and eye movements in the control of activities of daily living. Perception 28, 1311–1328 (1999).
Neggers, S. F. W. & Bekkering, H. Coordinated control of eye and hand movements in dynamic reaching. Hum. Mov. Sci. 21, 37–64 (2002).
Mayr, U. Spatial attention and implicit sequence learning: Evidence for independent learning of spatial and nonspatial sequences. J. Exp. Psychol. Learn. Mem. Cogn. 22, 350–364 (1996).
Remillard, G. Pure perceptual-based sequence learning. J. Exp. Psychol. Learn. Mem. Cogn. 29, 581–597 (2003).
Schwarb, H. & Schumacher, E. Generalized lessons about sequence learning from the study of the serial reaction time task. Adv. Cogn. Psychol. 8, 165–178 (2012).
Willingham, D. B. Implicit motor sequence learning is not purely perceptual. Mem. Cognit. 27, 561–572 (1999).
Danion, F. R. & Flanagan, J. R. Different gaze strategies during eye versus hand tracking of a moving target. Sci. Rep. 8, 10059 (2018).
Fooken, J., Kreyenmeier, P. & Spering, M. The role of eye movements in manual interception: A mini-review. Vis. Res. 183, 81–90 (2021).
Vesia, M., Prime, S. L., Yan, X., Sergio, L. E. & Crawford, J. D. Specificity of human parietal saccade and reach regions during transcranial magnetic stimulation. J. Neurosci. 30, 13053–13065 (2010).
Johansson, R. S., Westling, G., Bäckström, A. & Flanagan, J. R. Eye-hand coordination in object manipulation. J. Neurosci. 21, 6917–6932 (2001).
Sailer, U., Flanagan, J. R. & Johansson, R. S. Eye-hand coordination during learning of a novel visuomotor task. J. Neurosci. 25, 8833–8842 (2005).
Marcus, D. J., Karatekin, C. & Markiewicz, S. Oculomotor evidence of sequence learning on the serial reaction time task. Mem. Cogn. 34, 420–432 (2006).
Tal, A. et al. Oculomotor anticipation reveals a multitude of learning processes underlying the serial reaction time task. Sci. Rep. 11, 6190 (2021).
Tal, A. & Vakil, E. How sequence learning unfolds: Insights from anticipatory eye movements. Cognition 201, 104291 (2020).
Vakil, E., Bloch, A. & Cohen, H. Anticipation measures of sequence learning: manual versus oculomotor versions of the serial reaction time task. Q. J. Exp. Psychol. 70, 579–589 (2017).
Albouy, G. et al. Implicit oculomotor sequence learning in humans: Time course of offline processing. Brain Res. 1090, 163–171 (2006).
Kantak, S. S. & Winstein, C. J. Learning-performance distinction and memory processes for motor skills: A focused review and perspective. Behav. Brain Res. 228, 219–231 (2012).
Greeley, B. et al. Multiple bouts of high-intensity interval exercise reverse age-related functional connectivity disruptions without affecting motor learning in older adults. Sci. Rep. 11, 17108 (2021).
Mang, C. S., Snow, N. J., Wadden, K. P., Campbell, K. L. & Boyd, L. A. High-intensity aerobic exercise enhances motor memory retrieval. Med. Sci. Sports Exerc. 48, 2477–2486 (2016).
Brodie, S. M., Meehan, S., Borich, M. R. & Boyd, L. A. 5 Hz repetitive transcranial magnetic stimulation over the ipsilesional sensory cortex enhances motor learning after stroke. Front. Hum. Neurosci. 8, 143 (2014).
Meehan, S. K., Dao, E., Linsdell, M. A. & Boyd, L. A. Continuous theta burst stimulation over the contralesional sensory and motor cortex enhances motor learning post-stroke. Neurosci. Lett. 500, 26–30 (2011).
Meehan, S. K., Randhawa, B., Wessel, B. & Boyd, L. A. Implicit sequence-specific motor learning after subcortical stroke is associated with increased prefrontal brain activations: An fMRI Study. Hum. Brain Mapp. 32, 290–303 (2011).
Pew, R. W. Levels of analysis in motor control. Brain Res. 71, 393–400 (1974).
Stadler, M. A. & Frensch, P. A. Handbook of Implicit Learning (Sage, 1997).
Vidoni, E. D. & Boyd, L. A. Achieving enlightenment: What do we know about the implicit learning system and its interaction with explicit knowledge?. J. Neurol. Phys. Ther. 31, 145–154 (2007).
Singh, T., Perry, C. M. & Herter, T. M. A geometric method for computing ocular kinematics and classifying gaze events using monocular remote eye tracking in a robotic environment. J. NeuroEng. Rehabil. 13, 10 (2016).
Ayala, N. & Niechwiej-Szwedo, E. Effects of blocked vs. interleaved administration mode on saccade preparatory set revealed using pupillometry. Exp. Brain Res. 239, 245–255 (2021).
Boyd, L. A. & Winstein, C. J. Implicit motor-sequence learning in humans following unilateral stroke: The impact of practice and explicit knowledge. Neurosci. Lett. 298, 65–69 (2001).
Grafton, S. T., Hazeltine, E. & Ivry, R. B. Motor sequence learning with the nondominant left hand: A PET functional imaging study. Exp. Brain Res. 146, 369–378 (2002).
Pohl, P. S., McDowd, J. M., Filion, D. L., Richards, L. G. & Stiers, W. Implicit learning of a perceptual-motor skill after stroke. Phys. Ther. 81, 1780–1789 (2001).
Willingham, D. B., Nissen, M. J. & Bullemer, P. On the development of procedural knowledge. J. Exp. Psychol. Learn. Mem. Cogn. 15, 1047–1060 (1989).
Munoz, D. P., Broughton, J. R., Goldring, J. E. & Armstrong, I. T. Age-related performance of human subjects on saccadic eye movement tasks. Exp. Brain Res. 121, 391–400 (1998).
Dunn, M. J. et al. Minimal reporting guideline for research involving eye tracking (2023 edition). Behav. Res. Methods 56, 4351–4357 (2023).
Howard, J. H. & Howard, D. V. Age differences in implicit learning of higher order dependencies in serial patterns. Psychol. Aging 12, 634–656 (1997).
Nemeth, D. et al. Sleep has no critical role in implicit motor sequence learning in young and old adults. Exp. Brain Res. 201, 351–358 (2010).
Schmidt, R. A. & Lee, T. D. Motor Control and Learning: A Behavioural Emphasis (Human Kinetics, 2005).
Vieluf, S., Massing, M., Blandin, Y., Leinen, P. & Panzer, S. The role of eye movements in motor sequence learning. Hum. Mov. Sci. 40, 220–236 (2015).
Laamerad, P., Guitton, D. & Pack, C. C. Eye movements shape visual learning. Proc. Natl. Acad. Sci. 117, 8203–8211 (2020).
Fisk, J. D. & Goodale, M. A. The organization of eye and limb movements during unrestricted reaching to targets in contralateral and ipsilateral visual space. Exp. Brain Res. 60, 159–178 (1985).
Fischer, B. & Rogal, L. Eye-hand-coordination in man: A reaction time study. Biol. Cybern. 55, 253–261 (1986).
Armstrong, I. T., Judson, M., Munoz, D. P., Johansson, R. S. & Flanagan, J. R. Waiting for a hand: Saccadic reaction time increases in proportion to hand reaction time when reaching under a visuomotor reversal. Front. Hum. Neurosci. 7, 319 (2013).
Funding
This work was funded by a Natural Sciences and Engineering Research Council of Canada (NSERC) / Discovery Grant (PI L.A.B.) RGPIN-2023-03647. CR received funding from the Natural Sciences and Engineering Research Council (NSERC) of Canada, Post-Graduate Scholarship (PGS-D).
Author information
Authors and Affiliations
Contributions
C.R. contributed to the conception and design of the study, as well as the acquisition, analysis, and interpretation of data. C.R. drafted the original manuscript and was involved in revisions and editing. A.H. assisted with the development of data pre-processing scripts, contributed to data interpretation, and participated in manuscript editing. L.A.B. was involved in the study’s conception and design, provided insights on data interpretation, and contributed to manuscript writing and editing. All authors approved the final version of the manuscript, and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved; and all persons designed as authors qualify for authorship, and all those who qualify for authorship are listed.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Rubino, C., Harrison, A.T. & Boyd, L.A. Oculomotor learning is evident during implicit motor sequence learning. Sci Rep 15, 9498 (2025). https://doi.org/10.1038/s41598-025-93498-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-93498-0
