
Abstract
Gap junction protein B5 (GJB5, also known as Connexin 31.1) has recently been reported to be downregulated in several cancer types, where it functions primarily as a tumor suppressor in cancers such as melanoma and non-small cell lung cancer (NSCLC). However, there no reports describing its prognostic and immunological roles in pan-cancer. This study evaluated the association of GJB5 in various cancer types by a comprehensive pan-cancer analysis. The differential GJB5 expression in tumor and adjacent tissues acquired from The Cancer Genome Atlas (TCGA) databases was compared. Furthermore, univariate Cox regression and Kaplan-Meier survival analyses were performed to assess the influence of GJB5 on the disease-specific survival (DSS), disease-free interval (DFI), clinical stage, progression-free interval (PFI), and overall survival (OS) in various cancers. Moreover, the levels of GJB5 and its activity in the tumor microenvironment were assessed via the Tumor Immune Single-cell Hub (TISCH). In addition, the biological importance of GJB5 levels in various cancers was further assessed via Gene Set Enrichment Analysis. Tumor-Immune System Interactions Database (TISIDB) and Tumor Immune Estimation Resource Database 2.0 (TIMER2.0) tools indicated that GJB5 affected the tumor’s immune infiltration potential. This research also evaluated the association of GJB5 with immune features: immune modulatory genes, tumor mutational burden (TMB), and microsatellite instability (MSI). The data indicated that enhanced GJB5 level was linked to worse DFI, OS, PFI, and DSS in some cancers. Additionally, GJB5 level was positively related to immune modulatory genes, TMB, immune cell infiltration, immunological checkpoints, and MSI in malignancies. Furthermore, our study demonstrated that GJB5 was upregulated in colorectal cancer tissues compared to normal tissues. We also assessed GJB5 expression across various pancreatic cell lines. Notably, GJB5 was highly expressed in pancreatic cancer cells relative to normal pancreatic epithelial cells. Additionally, GJB5 knockdown in pancreatic cancer cells resulted in a significant reduction in cell proliferation. In summary, the findings indicated the potential of GJB5 as a prospective prognostic indicator and immunological biomarker.
Similar content being viewed by others

Introduction
Cancer is responsible for significant mortality and imposes a considerable burden on public health1. It was estimated that in 2024, there will be 611,720 deaths and 2,001,140 new cases related to cancer will occur in the USA2. Furthermore, the combined 5-year survival rates for all cancers have risen from 49% in the mid-1970s to 69% between 2013 and 20192. Currently, cancers with the lowest survival rates include pancreatic cancer (13%), followed by esophagus and liver (22%), and then lung cancer (25%)2. The cancer burden continues to increase with the increasing number of new diagnoses and death cases. Unfortunately, new and reliable biomarkers for cancer prognosis and treatment are still lacking. With advancements in technology, the discovery of biomarkers using computers has now become a hotspot in biomedical sciences. Therefore, in this research, potential therapeutic targets and biomarkers were identified by pan-cancer analysis for efficient tumor prognostic.
Gap junction protein B5 (GJB5) or Cx31.1 is a 31.088 kD protein comprising 273 amino acids and is primarily expressed in the skin epidermis and testes3,4. Recently, two studies have indicated that GJB5 serves as an essential modulator of invasion and migration in cutaneous malignant melanoma and lung cancer cells5,6. Scatolini et al. reported significant downregulation of GJB5 expression in metastatic melanoma lesions compared to primary tumors, as well as in BRAF-mutated versus BRAF-wild type (BRAF-WT) melanomas. Similarly, GJB5 expression is lower in BRAF-V600E cell lines compared to BRAF-WT cells, with a notable increase following treatment with mitogen-activated protein kinase (MAPK) inhibitors (MAPKi). In MAPKi-resistant melanoma cells, GJB5 expression remains elevated, correlating with morphological changes. Moreover, in clinical melanoma samples, GJB5 underexpression is linked to poorer overall survival, particularly in early-stage disease5. Thus, while high GJB5 expression is associated with drug resistance, it also correlates with better patient prognosis, underscoring the need for further research to clarify GJB5’s multifaceted role in tumors. Additionally, the study suggested that GJB5 exhibits tumor-suppressive properties in lung cancer, being downregulated in non-small cell lung cancer (NSCLC) cell lines. Overexpression of GJB5 in H1299 cells reduced cell proliferation and tumorigenicity in xenografts. GJB5 overexpression also caused a G1 phase delay, mediated by cyclin D3, inhibited anchorage-independent growth, and suppressed cell migration and invasion. These effects may result from increased expression of epithelial markers, such as cytokeratin 18, and decreased expression of mesenchymal markers, such as vimentin, suggesting a Cx31.1-mediated shift from a mesenchymal to an epithelial phenotype6. However, comprehensive research on the activity of GJB5 in various malignant tumors is still lacking.
Therefore, this research comprehensively investigated different cancers to identify the association of GJB5 in tumorigenesis and tumor advancement. The levels of GJB5 in different cancers acquired from the Cancer Genome Atlas (TCGA) and the Genotype-Tissue Expression (GTEx) databases were evaluated to assess its significance in prognosis and immune system participation. Furthermore, the associations of GJB5 levels with various factors, including tumor mutational burden (TMB), immune checkpoints, immune regulatory genes, and microsatellite instability (MSI), were also elucidated. Moreover, GJB5 expression levels and cellular functions were also analyzed. The data will provide evidence for the multifaceted roles of GJB5 and its efficiency as a biomarker for the prognostic prediction and immunotherapy of different cancers.
Methods
Bioinformatic analysis and RNA-sequencing data
The Tumor Immune Estimation Resource 2.0 (TIMER2.0, http://timer.cistrome.org)7 was employed to assess the GJB5 levels in pan-cancer. Furthermore, GJB5 levels and immune scores in different cancers were identified via Gene Expression Profiling Interactive Analysis (GEPIA, http://gepia.cancer-pku.cn/)8,9. TIMER 2.0 is an advanced repository comprising expression profiles of genes and generalized tumor-infiltrating immune cell functions of 23 types of cancer patients from TCGA10. GEPIA is an e-tool that provides a robust algorithm for 23 different cancers and has been employed to evaluate clinicopathological data and pan-cancer survival based on UCSC (http://xena.ucsc.edu/cite-us)11.
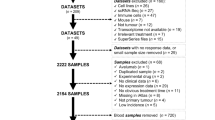
To assess the clinicopathological data and survival in various cancers, RNA-seq data and related prognostic profiles for 23 different cancers were acquired from TCGA. Individuals lacking complete clinical information were removed from the study. Moreover, survival data for 23 types of cancer were assessed using the survival packages in R version 4.1.1, which we previously published12. Spearman’s rank correlations were used to evaluate the links between GJB5 levels and cancer stages via the ‘stats’ package, whereas the statistical significance was measured via the ‘cor.test’ function. The Sangerbox (http://www.sangerbox.com/home.html) bioinformatics tool was utilized to examine the prognostic impact of various factors on disease-specific survival (DSS), disease-free interval (DFI), progression-free interval (PFI), and overall survival (OS). Moreover, the univariate Cox proportional hazards regression analysis (https://www.jstor.org/stable/2240714) was carried out as described in the previous literature13. The statistical importance was assessed via the Log-Rank test, and statistically, the p-value of 0.05 indicated the significance threshold. The TISIDB (http://cis.hku.hk/TISIDB/index.php) was employed to assess GJB5 levels in different molecular pan-cancer subtypes.
Single-cell expression of GJB5 in pan-cancer
To assess GJB5 levels in the major cell types in the tumor microenvironment (TME), 22 datasets were obtained from the TISCH database (http://tisch1.comp-genomics.org/home/). GJB5 expression in different cell types was visualized via heatmaps.
Tumor microenvironment and biological assessment
The stromal/immune cells infiltrating various tumors were identified via the Estimation of Stromal and Immune Cells in Malignant Tumor Tissues from Expression Data (ESTIMATE) algorithm. The TME scores were processed and analyzed using R, whereas tumor purity, as well as stromal and immune scores from the expression data, were assessed via the ‘estimate’ package. In addition, the ‘estimateScore’ function was utilized to measure the ESTIMATE scores. The relation between TME scores and GJB5 expression was elucidated via Spearman’s rank correlation method, while the statistical strength and significance of these correlations were identified using the ‘cor.test’ function. For data visualization, the ‘ggpubr’, ‘ggplot2’, ‘and ‘ggExtra’ packages were employed. Scatter plots were created to visualize the association between GJB5 levels and TME scores.
For each cancer type, the association of GJB5 levels with MSI and TMB was identified via Spearman’s rank correlations, using the ‘cor.test’ function for determining significance. Visualization was conducted using the ‘fmsb’ package to create radar charts. GJB5 expression in different immune subtypes in different cancers was assessed by the TISIDB.
The relation between GJB5 levels and immune-related genes in various cancers was also analyzed via Pearson’s correlation. The ‘ggplot2’ package was employed for generating heatmaps, and the significant p-values were annotated with asterisks. Moreover, the association of GJB5 levels with MMR-related genes, KRAS, TP53, and BRAF was similarly assessed.
Analysis of GJB5 genomic mutations in pan-cancer
To evaluate different cancer genomic mutational materials, cBio Cancer Genomics Portal (c-BioPortal) (http://cbioportal.org) was employed. c-BioPortal is an automatic deduction e-site for assessing overall cancer genomic characters datasets14.
Pathway enrichment analysis
The association between GJB5 levels and “c5.all.v7.1.symbols.gmt” gene sets in pan-cancer was evaluated via the Gene Set Enrichment Analysis (GSEA) using the ‘limma’, ‘clusterProfiler’, ‘org.Hs.eg.db’, and ‘enrichplot’ packages in R. Furthermore, the GSCA (https://guolab.wchscu.cn/GSCA/#/) was employed to assess the relation of GJB5 expression with pathway activity and Gene Set Variation Analysis (GSVA) score. In GSCA, reverse phase protein array (RPPA) data from the TCPA (https://www.tcpaportal.org/) database were employed to measure pathway activity scores for 10 cancer-related pathways in 7876 TCGA samples of 32 different cancers.
Cell lines, transfection, culture, and infection
The pancreatic normal cell lines (HPDE and HPNE) and pancreatic cancer cell lines (AsPC-1, Capan-1, Capan-2, CFPAC-1, MIA PaCa-2, PANC-1) were obtained from the American Type Culture Collection (ATCC). PANC-1 and HPDE cells were cultured in DMEM, while the other pancreatic cancer cell lines were cultured in RPMI-1640, both supplemented with 1% penicillin-streptomycin and 10% fetal bovine serum (FBS). All cells were maintained at 37 °C in a 5% CO₂ incubator. For transient transfection, OriFect Transfection Reagent (ORISCIENCE) was used according to the manufacturer’s instructions. PANC-1 cells were transfected with short interfering RNAs (siRNAs) and harvested 48 h post-transfection. The siRNAs used were synthesized by GenePharma (Suzhou, China) as follows: siRNA Negative Control (NC), UUCUCCGAACUGUCACGU; siGJB5-1, GCAGAUCCAUGUCCCAAUAtt; siGJB5-2, UUGAGCUUUCCUUGCUGCCtt; siGJB5-3, UUUCUUCACAUGGUCUCGGtt.
RNA isolation and quantitative real-time PCR
Total RNA from the stably transfected cells was isolated using the Cell Total RNA Isolation Kit (FOREGENE, Catalog Number: RE-03113). The isolated RNA was reverse transcribed into cDNA following the instructions provided in the RT Master Mix Kit (RK20433 | ABScript Neo RT Master Mix for qPCR with gDNA Remover). For PCR amplification, the Universal SYBR qPCR Master Mix Kit (RK21203 | 2 × Universal SYBR Green Fast qPCR Mix) was used. The relative expression of target genes was measured in triplicate and calculated using the comparative CT method 2(−ΔCT), where ΔCT = CT (internal control) − CT(gene of interest)). The primers used for amplification were as follows: GJB5 F: GAGGTTCAGGAGAAGAGGCA, GJB5 R: TTTGGGGTAGAATGAGTGGAA.
EdU incorporation assay
For EdU analysis, the protocol provided with the BeyoClick™ EdU-647 Kit (Catalog Number: C0081S) was followed. Briefly, the treated cells were incubated with 50 µmol/L 5-ethynyl-2′-deoxyuridine (EdU) for 3 h at 37 °C. To assess Hoechst-positive cells, the cells were fixed in 4% paraformaldehyde (PFA), permeabilized with 0.3% Triton X-100, and then stained for 30 min using 1× Click Reaction Solution. Afterward, the cells were counterstained with Hoechst 33,342 for an additional 30 min. Hoechst- and EdU-positive cells were immediately analyzed using an inverted fluorescence microscope (OLYMPUS IX81).
Clinical samples
A total of 18 samples were obtained from colorectal cancer (CRC) patients who had undergone curative colorectal tumor resection at West China Hospital (WCH, Chengdu, China). Among these, 9 CRC tissues were paired with corresponding adjacent normal tissues. These tissues were used for quantitative PCR validation.
Statement
The study was conducted in compliance with the International Conference on Harmonization guidelines for Good Clinical Practice (E6) and the 2013 Declaration of Helsinki. The protocols followed in this study were approved by the Ethical Review Committees of West China Hospital [the ethical approval number: 2023 Review (1501)], and all patients provided written informed consent.
Statistical assessments
GraphPad Prism 5, R 4.1.1, and SPSS 26.0 were employed for all analyses, and the data are given as mean ± SEM. p-values were measured via the Students’ t-test. where, *p < 0.05, **p < 0.01, ***p < 0.001, #FDR ≤ 0.05.
Results
Expression analyses of GJB5 gene in pan-cancers
It has been observed that tumors have heterogeneity, and various molecules have different expressions in different types of tumors. Therefore, this research first verified the overall GJB5 expression in different cancers by assessing its transcriptional profiles in 33 different cancers via TIMER 2.0. Furthermore, its mRNA expression was identified in pan-cancer, including esophageal carcinoma (ESCA), colon adenocarcinoma (COAD), lung squamous cell carcinoma (LUSC), pancreatic adenocarcinoma (PAAD), and cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC) (Fig. S1A). It was observed that GJB5 levels were reduced in prostate adenocarcinoma (PRAD) and breast invasive carcinoma (BRCA). Moreover, these data were further assessed via the GEPIA2. The results revealed that GJB5 had variable expression in different cancers and their corresponding normal tissues, including PAAD, CESC, thymoma (THYM), skin cutaneous melanoma (SKCM), LUSC, and PRAD (all p < 0.05 in Fig. S1B–G; Fig. S2A–L, S3A–L). Most cancer types (14/21 in Fig. S1A; 19/30 in Fig. S1B–G and Figs. S2–S3) indicated increased GJB5 expression compared with para-cancer tissues such as stomach adenocarcinoma (STAD), head and neck squamous cell carcinoma (HNSC), and uterine corpus endometrial carcinoma (UCEC). Notably, we re-analyzed the data directly from the TCGA datasets and used the statistical software R for plotting. Our analysis revealed that the expression of GJB5 in the tumor types analyzed was fully consistent with the data in the database (Fig. 1). Furthermore, we found that GJB5 expression was significantly higher in tumor tissues of kidney renal papillary cell carcinoma (KIRP) and kidney renal clear cell carcinoma (KIRC) compared to normal tissues.

GJB5 expression levels in pan-cancer. ***p < 0.001, **p < 0.01, *p < 0.05.
GJB5 levels are linked with clinicopathological features and prognosis in pan-cancer
The relationships between GJB5 levels and various clinicopathological features of pan-cancer patients were examined using the TCGA dataset, with analysis and visualizations performed with R. The data indicated that GJB5 mRNA expression was not consistently associated with AJCC/UICC tumor node metastasis stages in patients with KIRP, lung adenocarcinoma (LUAD), esophageal carcinoma (ESCA), kidney chromophobe (KICH), pancreatic adenocarcinoma (PAAD), skin cutaneous melanoma (SKCM), and stomach adenocarcinoma (STAD), although some stages in these cancers did show a statistically significant relationship with GJB5 expression (p < 0.05 in some cases, Fig. 2A–G). Specifically, GJB5 levels were positively correlated with cancer stages in SKCM (stages I–III), KICH (stages I–II), PAAD (stages I–II), LUAD (stages I and III), and STAD (stages I, II, and IV), but not in SKCM stage IV, or STAD stage III (Fig. 2B, D–G). A similar trend was observed in patients with testicular germ cell tumor (TGCT), adrenocortical carcinoma (ACC), rectal adenocarcinoma (READ), head and neck squamous cell carcinoma (HNSC), KIRC, cholangiocarcinoma (CHOL), liver hepatocellular carcinoma (LIHC), colon adenocarcinoma (COAD), and bladder urothelial carcinoma (BLCA) (Fig. S4A–I).
Moreover, increased GJB5 expression was associated with lower stages in ESCA (stages II and III) and KIRP (stages I, II, and IV), but not in KIRP stage III patients (Fig. 2A, C). A similar trend, though not statistically significant, was observed in lung squamous cell carcinoma (LUSC), mesothelioma (MESO), breast cancer (BRCA), thyroid carcinoma (THCA), and uveal melanoma (UCM) (Fig. S4J–N). Overall, the pan-cancer assessment revealed variability in GJB5 expression across different pathological stages of various cancers.
Moreover, univariate Cox regression results indicated that GJB5 upregulation was associated with shorter PFI in stomach and esophageal carcinoma (STES), pan-kidney cohort (KIRP + KIRC + KICH and KIPAN), and PAAD, shorter DSS in LUAD, PAAD, SKCM, KIPAN, and MESO, shorter DFI in PAAD, KIPAN, and STES, and shorter OS in LUAD, SKCM, PAAD, and KIPAN (Fig. 2H–K). Overall, TCGA data assessment indicated that GJB5’s prognostic significance varies across pan-cancer.

Analysis of GJB5 clinicopathologic expression in pan-cancer. (A–G) Association of GJB5 expression with predominant pathological stages, including stage I to IV in different cancers, including ESCA, KICH, KIRP, LUAD, PAAD, SKCM, and STAD in TCGA data. (H–K) Association of GJB5 expression with PFI, DSS, DFI, and OS in cancer patients. Forest plots indicate the data of the univariate Cox regression test. ESCA esophageal carcinoma, KICH kidney chromophobe, KIRP kidney renal papillary cell carcinoma, LUAD lung adenocarcinoma, PAAD pancreatic adenocarcinoma, SKCM skin cutaneous melanoma, STAD stomach adenocarcinoma.
GJB5 levels are associated with tumor molecular subtypes in pan-cancer
GJB5 levels in molecular subtypes of pan-cancers was assessed using the TISIDB database (Fig. 3A–P). The data indicated that GJB5 had specific expression trends in 16 different cancer molecular subtypes, including ACC, HNSC, Pheochromocytoma, Paraganglioma (PCPG), COAD, ESCA, STAD, KIRP, Brain Lower Grade Glioma (LGG), LUSC, BRCA, PRAD, Glioblastoma multiforme (GBM), READ, SKCM, Ovarian serous cystadenocarcinoma (OV), and UCEC.

Correlation of GJB5 gene expression with molecular subtype and tumor stage. (A–P) Association between GJB5 expression and the molecular subtype in different cancers, including BRCA, STAD, LUSC, COAD, GBM, LGG, OV, KIRP, PCPG, HNSC, PAAD, ESCA, READ, ACC, SKCM, and UCEC. BRCA breast cancer, STAD stomach adenocarcinoma, LUSC lung squamous cell carcinoma, COAD colon adenocarcinoma, GBM glioblastoma multiforme, LGG brain lower grade glioma, OV ovarian serous cystadenocarcinoma, KIRP kidney renal papillary cell carcinoma, PCPG pheochromocytoma and paraganglioma, HNSC head and neck squamous cell carcinoma, PAAD pancreatic adenocarcinoma, ESCA esophageal carcinoma, READ rectum adenocarcinoma, ACC adrenocorticalcarcinoma, SKCM skin cutaneous melanoma, UCEC uterine corpus endometrial carcinoma.
GJB5 mutations in pan-cancer
The GJB5 gene alterations in pan-cancer were evaluated using the TCGA dataset from the cBioPortal database. We also reanalyzed the data directly from the cBioPortal and used R for visualization (Fig. 4A). The prevalence of GJB5 gene mutations in ovarian serous cystadenocarcinoma was > 4%, and the majority of mutations occurred as “Amplification” (Fig. 4A). However, most mutations of the GJB5 gene were identified in PCPG, including “Deep Deletion” (Fig. 4A). Moreover, it was observed that GJB5 consists of two structural domains: the Connexin domain and Connexin_CCC, both of which are susceptible to missense mutations (Fig. 4B).

GJB5 gene mutations in pan-cancer. (A,B) Analysis of GJB5 gene mutations in various types of tumor tissues.
Single-cell analysis of GJB5 expression
A comprehensive single-cell analysis of GJB5 across 11 cancer types revealed its predominant expression in malignant cells (Fig. 5). Figure 5A presents a heatmap illustrating consistent GJB5 enrichment in malignant cells across most datasets, with particularly high expression observed in monocytes/macrophages (Mono/Macro) within the nasopharyngeal carcinoma (NPC) dataset (GSE150430). Figure 5B–F display cell type annotations across five cancer types, where malignant cells consistently show elevated GJB5 expression. These cancer types include HNSC, oral squamous cell carcinoma (OSCC), NPC, basal cell carcinoma (BCC), and squamous cell carcinoma (SCC).

GJB5 single-cell analysis. (A) A heatmap of GJB5 expression across all cell types in the single-cell datasets, with darker colors indicating higher expression levels. This visualization underscores the enrichment of GJB5 in both malignant cells, such as basal cell carcinoma (BCC) and head and neck squamous carcinoma (HNSC), as well as specific immune subsets, including monocytes/macrophages (Mono/Macro). (B–F) Validation of GJB5’s high expression in malignant cells within each dataset. Malignant clusters are distinctly labeled with unique colors to facilitate clear identification and comparison. BCC basal cell carcinoma, HNSC head and neck squamous carcinoma, NPC nasopharyngeal carcinoma, OSCC oral squamous cell carcinoma, SCC squamous cell carcinoma.
Gene set enrichment analysis
This study also performed GSEA of signature patterns to elucidate the biological significance of GJB5 across various cancers. The results revealed associations between GJB5 expression levels and specific hallmark terms, such as alternative miRNA splicing via the spliceosome and immune response, in multiple cancers, including KIRP (Fig. 6A), LGG (Fig. 6B), MESO (Fig. 6C), PCPG (Fig. 6D), PRAD (Fig. 6E), READ (Fig. 6F), THCA (Fig. 6G), and Uveal Melanoma (UVM) (Fig. 6H). The enriched pathway details from Fig. 6 are systematically tabulated in supplementary material 3. This tabulated format not only enhances readability but also facilitates direct comparison of pathway statistics across different cancer types.

GSEA of different cancers. The association of GJB5 and gene sets in KIRP (A), LGG (B), MESO (C), PCPG (D), PRAD (E), READ (F), THCA (G), and UVM (H). KIRP kidney renal papillary cell carcinoma, LGG brain lower grade glioma, MESO mesothelioma, PCPG pheochromocytoma and paraganglioma, PRAD prostate adenocarcinoma, READ rectum adenocarcinoma, THCA thyroid carcinoma, UVM uveal melanoma.
GJB5 levels are related to immune infiltration
To investigate the relationship between GJB5 expression and the immune landscape of tumors, we analyzed its association with established immune subtypes (C1–C6) across a diverse panel of cancers (Fig. 7A–L, Fig. S6). Our analysis revealed a complex, cancer-type-specific pattern of associations. Notably, GJB5 expression exhibited a significant positive correlation with the C2 (IFN-γ dominant) immune subtype in several cancers, including ACC, BLCA, ESCA, HNSC, CESC, and LUSC. The C2 subtype is characterized by high M1/M2 macrophage polarization, strong CD8+ T cell signaling, and elevated TCR diversity, suggesting that GJB5 may be associated with a pro-inflammatory, anti-tumor immune response in these cancers.
Conversely, a significant negative correlation was observed between GJB5 expression and the C5 (immunologically quiet) subtype in ACC and LGG. The C5 subtype is marked by a paucity of lymphocytes and a strong macrophage response, predominantly driven by M2 macrophages. This suggests a potential role for GJB5 in pathways counteracting immune silencing or its association with the absence of an immune response, depending on the cancer context.
In other cancers, GJB5 expression was positively associated with the C3 (inflammatory) subtype in BLCA, HNSC, and LUSC. The C3 subtype is characterized by increased Th17 and Th1 gene expression, indicating a potential role for GJB5 in inflammatory responses within the tumor microenvironment. A positive correlation between GJB5 expression and the C4 (lymphocyte-depleted) subtype was also observed in BLCA, ESCA, HNSC, CESC, and LUSC. The C4 subtype is marked by prominent macrophage features, Th1 suppression, and an elevated M2 response.
We further assessed the relationship between GJB5 expression and the TME using the ESTIMATE method (Fig. 8A–H). A negative correlation between GJB5 expression and immune scores was observed in HNSC, KICH, LGG, and LUSC. However, GJB5 expression was positively correlated with immune scores in COAD, BRCA, CESC, and PRAD. Specifically, regarding immune cell infiltration (ICI), GJB5 expression showed significant positive correlations in BRCA, CESC, COAD, and PRAD, suggesting enhanced immune cell presence. Conversely, negative correlations were observed in ESCA and THCA, potentially indicating reduced immune cell infiltration or an association with an immune-cold microenvironment. The data also revealed relationships between GJB5 levels and different immune cells in various cancers, indicating that GJB5 can modulate ICI in the TME (Figs. S6–S8).
Altogether, GJB5 expression is either positively or negatively associated with ICI, depending on the cancer type.

The association of GJB5 expression with TISIDB dataset tumor immune subtypes. The association of GJB5 expression with TISIDB dataset tumor immune subtypes in ACC (A), BLCA (B), BRCA (C), ESCA (D), GBM (E), HNSC (F), CESC (G), CHOL (H), COAD (I), LGG (J), LUAD (K) and LUSC (L). The immunological subtypes included C1: wound healing, C2: IFN-gamma dominant, C3: inflammatory, C4: lymphocyte depleted, C5: immunologically quiet, and C6: TGF-b dominant. ACC adrenocorticalcarcinoma, BLCA bladder urothelial carcinoma, BRCA breast cancer, ESCA esophageal carcinoma, GBM glioblastoma multiforme, HNSC head and neck squamous cell carcinoma, CESC cervical squamous cell carcinoma and endocervical adenocarcinoma, CHOL cholangiocarcinoma, COAD colon adenocarcinoma, LGG brain lower grade glioma, LUAD lung adenocarcinoma, LUSC lung squamous cell carcinoma.

The association of GJB5 expression with immune score in different cancers. (A–H) GJB5 expression was related to the immune score in different TIMER cancers.
The association of immune-related genes with GJB5 levels in pan-cancer
To further elucidate the role of GJB5 in the tumor immune microenvironment, we analyzed its co-expression patterns with a panel of established immune-related genes across a diverse range of cancers (Fig. 9). Our analysis revealed predominantly positive, cancer-type-specific associations between GJB5 and these genes. In the majority of cancers analyzed, including BRCA, CESC, COAD, and PRAD, GJB5 expression exhibited significant positive correlations with multiple immune-related genes. Specifically, GJB5 expression was positively correlated with PD-L1 (CD274), CTLA4, and TNFSF9 in several cancer types. These findings suggest a potential role for GJB5 in enhancing or co-occurring with the expression of genes involved in immune activation, checkpoint regulation, and other immune response pathways.
However, in a subset of cancers, notably ESCA and LUSC, GJB5 expression demonstrated significant negative correlations with certain immune-related genes. For example, in ESCA and LUSC, GJB5 expression was negatively correlated with CD28, CD40LG, and CD160. This contrasting pattern suggests that the role of GJB5 in immune regulation may be context-dependent, potentially reflecting differences in the tumor microenvironment, dominant immune evasion mechanisms, or other cancer-specific factors.

The association of immune-associated genes with GJB5 gene expression in pan-cancer. ***p < 0.001, **p < 0.01, *p < 0.05.
GJB5 expression association with MSI and TMB
The damaged microsatellite’s mismatch repair causes MSI, which impacts tumor prognosis by causing abnormalities in gene replication and promoting tumor proliferation. It was observed that GJB5 levels were positively related to MSI in LGG, STAD, THYM, BRCA, and COAD while negatively associated in SKCM, KIRP, PRAD, LUAD, THCA, and BLCA (all p < 0.05, Fig. 10A). TMB is a novel predictor of tumor immunotherapy effectiveness since it promotes the synthesis of tumor-specific and highly immunogenic antibodies. The data revealed a positive link between GJB5 expression and TMB in ACC, GBM, LUAD, PRAD, LGG, SARC, UCEC, STAD, COAD, READ, HNSC, PAAD, LUSC, BLCA, SKCM, and UCS (all p < 0.05, Fig. 10B), whereas the GJB5 level was negatively linked to TMB in ESCA and THYM (all p < 0.05, Fig. 10B).

GJB5 expression is related to immunological markers. The association of GJB5 expression (A) with MSI and (B) TMB.
The co-expression of GJB5 with key MSI-related proteins, such as PMS2, MSH6, MLH1, and other molecules promoting carcinogenesis, was also assessed. A positive association was observed in most cancer types (Fig. 11), whereas in some cancer types, GJB5 was positively related to oncogenes, such as NRAS, KRAS, ERBB2, and EGFR, suggesting a cancer-promoting function of GJB5 and possible association with prognosis prediction.

Analysis of key proteins related to MSI, such as MSH6, PMS2, MLH1, and other molecules promoting carcinogenesis and the pan-cancer gene (HOXB7) expression. ***p < 0.001, **p < 0.01, *p < 0.05.
Moreover, GJB5’s possible roles in tumor cells were evaluated, which revealed that GJB5 expression significantly impacts tumor cell processes, including cell cycle, DNA damage, apoptosis, hormone AR, RAS-MAPK, PI3K-AKT, EMT, RTK, and TSC-mTOR (Fig. S9). This inferred that differential GJB5 expression influences carcinogenesis and development based on cancer type.
Given that GJB5 was significantly upregulated in the bioinformatics analysis (Fig. 1A) compared to normal tissues, we first validated this finding in clinical colorectal cancer samples. GJB5 expression was assessed in colorectal cancer tissues and their corresponding adjacent normal tissues using reverse transcription quantitative polymerase chain reaction (RT-qPCR). The results revealed that GJB5 was upregulated in colorectal cancer tissues compared to normal tissues (Fig. 12A), consistent with the bioinformatics analysis shown in Fig. 1A.
We also measured GJB5 expression in various pancreatic cell lines, including HPDE, HPNE, AsPC-1, Capan-1, Capan-2, CFPAC-1, MIA PaCa-2, and PANC-1, by RT-qPCR (Fig. 12B). Compared to normal pancreatic epithelial cells (HPDE and HPNE), GJB5 was highly expressed in pancreatic cancer cells.
To investigate the effect of GJB5 on pancreatic cell proliferation, we knocked down its expression in PANC-1 cells using three independent siRNAs. GJB5 knockdown efficiently reduced its expression in PANC-1 cells (Fig. 12C). Furthermore, EdU incorporation assays revealed a significant reduction in PANC-1 cell proliferation following GJB5 knockdown (Fig. 12D, E).

The role of GJB5 in colorectal cancer and pancreatic cancer. (A) GJB5 expression was detected in colorectal cancer tissues and their corresponding adjacent normal tissues from clinical patients using RT-qPCR, (B) GJB5 expression was detected in HPDE, HPNE, AsPC-1, Capan-1, Capan-2, CFPAC-1, MIA PaCa-2 and PANC-1 cells by RT-qPCR. (C) GJB5 expression detected in PANC-1 cells with GJB5 knockdown or control by RT-qPCR. (D,E) Rate of cell proliferation in PANC-1 cells determined using EdU incorporation assays. Scale bar = 100 μm, 10. *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, ****p < 0.00001.
Discussion
The objective of pan-cancer studies is an accurate evaluation of transmission genes and recurrent genomic events or abnormalities in various cancers. For pan-cancer analysis, data should be standardized across various platforms to acquire standards for data processing and results presentation15. Several studies have indicated a predictable and causal link between gene expression and genomic alterations in various tumors. The association of transcriptome quantitative data and pan-cancer genomic status is significant16. Pan-cancer studies identify the variabilities and similarities among genes, investigate cellular alterations in all cancer types, and perform specific tests on single cells, functional elements, related sequences, and clinical features to predict related modulatory pathways, patient prognosis, and immune relevance17,18,19.
TCGA pan-cancer studies provide an in-depth understanding of variabilities and similarities between tumor types20. This research investigated the expression profile of GJB5 and its association with prognostic factors in 33 different cancers, including CESC, ESCA, LUAD, HNSC, LUSC, COAD, PAAD, CHOL, READ, BLCA, STAD and UCEC, which showed increased GJB5 levels. Moreover, univariate Cox analysis showed that in the PAAD and KIPAN patients, shorter PFI, DSS, DFI, and PFS were related to higher GJB5 levels. In clinical samples, up-regulated GJB5 levels were linked with patients with worse OS, especially at early stages5. These data indicated that GJB5 may be a potential predictive index for PAAD and KIPAN patients.
The TME is the environment surrounding cancer cells and mediates their survival21. It comprises a highly complex ecosystem containing peripheral immune and inflammatory cells, tumor cells, tumor vascular system, and other cellular components. Stromal and immune cells in TME can affect cancer progression and patient survival rates22,23. Cancer cells’ interaction with TME essentially mediates tumor prognosis, development, and response to therapy24. Here, single-cell analyses were employed to evaluate GJB5’s function in the TME, which suggested that GJB5 was primarily expressed in malignant cells, which modulates various cellular functions. Moreover, enrichment analysis demonstrated that GJB5 levels were linked with immune response in some cancers (LGG, THCA, MESO, KIRP, PRAD, READ, PCPG, and UVM). The literature has observed that epidermal cell development and differentiation are frequently enriched in most cancer types, including CESC, KIRC, SARC, LUSC, SKCM, LIHC, STAD, ESCA, TGCT, BRCA, and THYM, which was consistent with previous studies in malignant melanoma and laryngeal cancer5,25,26.
It has been observed that ICI markedly affects TME, and ICI and tumor growth are related to prognosis27,28. TME immune cells include M1 and M2 macrophages, activated natural killer cells, and regulatory T cells. These cells have been reported to affect EMT, modulate tumor behavior, and exhibit anticancer activities29,30,31,32. Several studies have associated tumor ICI with cancer prognosis33,34. Here, a statistically essential association was observed between GJB5 levels and different immune-infiltrating cells in CESC, KIRP, HNSC, COAD, LUSC, BRCA, and PRAD. Moreover, GJB5 expression significantly positively modulated immune function in KIRP and PRAD. Certain immune-infiltrating cells, like CD8+ T cells, recognize and destroy cancer cells; however, tumors can modulate the immune TME to aid immune escape. Furthermore, tumors can disrupt antigen presentation, cell function, and CD8+ T cell-cell infiltration via immunosuppressive cells and soluble cell surface mediators (for instance, PD-L1). Therefore, assessing the interaction of tumor-immune cells can predict immunotherapeutic responses and aid novel immunotherapy target development. GJB5 levels can be assessed to evaluate ICI, which can benefit patients and identify new targeted drugs for immunotherapies against cancers. This is the first research study to investigate the association between immune functions and GJB5 expression.
TMB is a dependable bio-index for predicting immunotherapy response against various cancers35. This investigation revealed that TMB was related to GJB5 expression in 33 different cancers. Furthermore, increased GJB5 levels were linked with high TMB in most cancers except ESCA and THYM. Additionally, GJB5 level was observed to be positively and strongly associated with MSI in STAD, THYM, COAD, LGG, and BRCA. Therefore, it was inferred that GJB5 may influence immunotherapy effectiveness in some cancers, such as STAD, LGG, and COAD, where GJB5 expression was negatively related to MSI and TMB. These results imply that GJB5 has the potential as a valuable marker for predicting the efficiency of immunotherapy in patients.
Furthermore, the bioinformatics analysis indicated that pan-oncogene GJB5 is variably expressed in normal and tumor tissues and is associated with TME ICI. GJB5 can serve as an immunotherapeutic target and a potential index for different cancers. Evaluation of novel bio-indices can improve outcomes in patients. However, there are certain limitations in this research. This research is primarily data-driven, with further cell studies needed to confirm the broader applicability of the results. Animal studies are also required to validate these findings. More importantly, these cell and animal models are essential to explore potential mechanisms.
Conclusions
In summary, the findings demonstrated that GJB5 expression varies across different tumors, with higher expression observed in PAAD, CRC, and others, and lower expression in PRAD, BRCA, and others, where it was found to modulate various tumor-related processes, such as cell proliferation, cell cycle and immune response. This suggests that GJB5 could serve as a therapeutic target as well as a prognostic biomarker. Further experimental studies and longitudinal clinical trials are needed to fully evaluate its role in cancer development.
Data availability
All data generated or analysed during this study are included in this published article [and its supplementary information files]. The datasets generated and/or analysed during the current study are available in the TCGA repository, [https://portal.gdc.cancer.gov/], TCGA raw data is shown in ZIP 2–7.
References
Sung, H. et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71 (3), 209–249. https://doi.org/10.3322/caac.21660 (2021).
Siegel, R. L., Giaquinto, A. N. & Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 74 (1), 12–49. https://doi.org/10.3322/caac.21820 (2024).
Haefliger, J. A. et al. Four novel members of the connexin family of gap junction proteins. Molecular cloning, expression, and chromosome mapping. J. Biol. Chem. 267 (3), 2057–2064. https://pubmed.ncbi.nlm.nih.gov/1370487 (1992).
Goliger, J. A. & Paul, D. L. Expression of gap junction proteins Cx26, Cx31.1, Cx37, and Cx43 in developing and mature rat epidermis. Dev. Dyn. Off Publ Am. Assoc. Anat. 200 (1). https://pubmed.ncbi.nlm.nih.gov/8081010 (1994).
Scatolini, M. et al. GJB5 association with BRAF mutation and survival in cutaneous malignant melanoma. Br. J. Dermatol. 186 (1), 117–128. https://doi.org/10.1111/bjd.20629 (2022).
Zhang, D. et al. Cx31.1 acts as a tumour suppressor in non-small cell lung cancer (NSCLC) cell lines through Inhibition of cell proliferation and metastasis. J. Cell. Mol. Med. 16 (5), 1047–1059. https://doi.org/10.1111/j.1582-4934.2011.01389.x (2012).
Li, T. et al. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 48 (W1), W509–W514. https://doi.org/10.1093/nar/gkaa407 (2020).
Tang, Z. et al. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 45 (W1). https://doi.org/10.1093/nar/gkx247 (2017).
Hu, W. et al. Coupled immune stratification and identification of therapeutic candidates in patients with lung adenocarcinoma. Aging 12 (16), 16514–16538. https://doi.org/10.18632/aging.103775 (2020).
Blum, A., Wang, P. & Zenklusen, J. C. SnapShot: TCGA-Analyzed tumors. Cell 173 (2), 530. https://doi.org/10.1016/j.cell.2018.03.059 (2018).
Navarro Gonzalez, J. et al. The UCSC genome browser database: 2021 update. Nucleic Acids Res. 49 (D1), D1046–D1057. https://doi.org/10.1093/nar/gkaa1070 (2021).
Yang, X. J., Xu, Y. F. & Zhu, Q. SPOP expression is associated with tumor-infiltrating lymphocytes in pancreatic cancer. PLoS One. 19 (7), e0306994. https://doi.org/10.1371/journal.pone.0306994 (2024).
Yang, X., Xian, X., Wang, Y. & Qiu, M. Assessing prognostic value of early tumor shrinkage and depth of response in first-line therapy for patients with advanced unresectable pancreatic cancer. BMC Gastroenterol. 21 (1), 294. https://doi.org/10.1186/s12876-021-01870-x (2021).
Cerami, E. et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2 (5), 401–404. https://doi.org/10.1158/2159-8290.CD-12-0095 (2012).
Li, S. et al. Multi-omics integration analysis of GPCRs in pan-cancer to uncover inter-omics relationships and potential driver genes. Comput. Biol. Med. 161, 106988. https://doi.org/10.1016/j.compbiomed.2023.106988 (2023).
Zhang, Y. et al. Pan-cancer analyses reveal the stratification of patient prognosis by viral composition in tumor tissues. Comput. Biol. Med. 167, 107586. https://doi.org/10.1016/j.compbiomed.2023.107586 (2023).
Jiang, X. et al. Comprehensive pan-cancer analysis of the prognostic and immunological roles of the METTL3/lncRNA-SNHG1/miRNA-140-3p/UBE2C axis. Front. Cell. Dev. Biol. 9, 765772. https://doi.org/10.3389/fcell.2021.765772 (2021).
Zhang, M. et al. In-depth characterization and identification of translatable LncRNAs. Comput. Biol. Med. 164, 107243. https://doi.org/10.1016/j.compbiomed.2023.107243 (2023).
Chen, W. S. et al. Novel RB1-loss transcriptomic signature is associated with poor clinical outcomes across cancer types. Clin. Cancer Res. Off J. Am. Assoc. Cancer Res. 25 (14), 4290–4299. https://doi.org/10.1158/1078-0432.CCR-19-0404 (2019).
Weinstein, J. N. et al. The cancer genome atlas Pan-Cancer analysis project. Nat. Genet. 45 (10), 1113–1120. https://doi.org/10.1038/ng.2764 (2013).
Jin, M., Cao, W., Chen, B., Xiong, M. & Cao, G. Tumor-derived lactate creates a favorable niche for tumor via supplying energy source for tumor and modulating the tumor microenvironment. Front. Cell. Dev. Biol. 10, 808859. https://doi.org/10.3389/fcell.2022.808859 (2022).
Xiao, Y. & Yu, D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol. Ther. 221, 107753. https://doi.org/10.1016/j.pharmthera.2020.107753 (2021).
Ren, B. et al. Tumor microenvironment participates in metastasis of pancreatic cancer. Mol. Cancer. 17 (1), 108. https://doi.org/10.1186/s12943-018-0858-1 (2018).
Matarrese, P. et al. The sex-related interplay between TME and cancer: On the critical role of Estrogen, MicroRNAs and autophagy. Cancers 13 (13). https://doi.org/10.3390/cancers13133287 (2021).
Fromme, J. E. & Zigrino, P. Melanoma metastasis, BRAF mutation and GJB5 connexin expression: A new prognostic factor. Br. J. Dermatol. 186 (1), 13–14. https://doi.org/10.1111/bjd.20756 (2022).
Broghammer, M. et al. Reduced expression of connexin 31.1 in larynx cancer is not caused by GJB5 mutations. Cancer Lett. 214 (2), 225–229 (2004). https://pubmed.ncbi.nlm.nih.gov/15363549
Cheng, Y. et al. Targeting CXCR2 inhibits the progression of lung cancer and promotes therapeutic effect of cisplatin. Mol. Cancer. 20 (1), 62. https://doi.org/10.1186/s12943-021-01355-1 (2021).
van der Leun, A. M., Thommen, D. S. & Schumacher, T. N. CD8+ T cell States in human cancer: Insights from single-cell analysis. Nat. Rev. Cancer. 20 (4), 218–232. https://doi.org/10.1038/s41568-019-0235-4 (2020).
Galluzzi, L., Humeau, J., Buqué, A., Zitvogel, L. & Kroemer, G. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat. Rev. Clin. Oncol. 17 (12), 725–741. https://doi.org/10.1038/s41571-020-0413-z (2020).
Ferrari, S. M. et al. Immune and inflammatory cells in thyroid cancer microenvironment. Int. J. Mol. Sci. 20 (18). https://doi.org/10.3390/ijms20184413 (2019).
Kießler, M. et al. Tumor-infiltrating plasmacytoid dendritic cells are associated with survival in human colon cancer. J. Immunother. Cancer. 9 (3). https://doi.org/10.1136/jitc-2020-001813 (2021).
Ren, F. et al. Immune infiltration profiling in gastric cancer and their clinical implications. Cancer Sci. 112 (9), 3569–3584. https://doi.org/10.1111/cas.15057 (2021).
Chen, L. & Musa, A. E. Boosting immune system against cancer by Resveratrol. Phytother Res. PTR 35 (10), 5514–5526. https://doi.org/10.1002/ptr.7189 (2021).
Cherry, A. B. C., Gherardin, N. A. & Sikder, H. I. Intracellular radar: Understanding Γδ T cell immune surveillance and implications for clinical strategies in oncology. Front. Oncol. 12, 1011081. https://doi.org/10.3389/fonc.2022.1011081 (2022).
Goodman, A. M. et al. Tumor mutational burden as an independent predictor of response to immunotherapy in diverse cancers. Mol. Cancer Ther. 16 (11), 2598–2608. https://doi.org/10.1158/1535-7163.MCT-17-0386 (2017).
Funding
This research was funded by Sichuan Provincial Natural Science Foundations Youth Funds (2025ZNSFSC1901 and 2025ZNSFSC1900).
Author information
Authors and Affiliations
Contributions
Xiaojuan Yang: Writing—review and editing, Writing—original draft. Xunjie Cao: Visualization, Software, Methodology, Formal analysis, Data curation. Qing Zhu: Writing—review and editing, Supervision. Hong Wu: Writing—review and editing, Supervision, Resources, Funding acquisition, Conceptualization.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang, X., Cao, X., Zhu, Q. et al. Pan-cancer analysis of GJB5 as a novel prognostic and immunological biomarker. Sci Rep 15, 14879 (2025). https://doi.org/10.1038/s41598-025-96389-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-96389-6
Keywords
This article is cited by
-
Gap junction protein beta 5 interacts with Gαi3 to promote Akt activation and cervical cancer cell growth
Cell Death & Disease (2025)
