
Abstract
The dawn of dinosaurs is marked by the appearance of the saurischian lineages in the Late Triassic fossil record, around 230 million years ago. This early burst of diversification of the group is majoritarily represented by sauropodomorphs and herrerasaurids in late Carnian to early Norian of Brazil, Argentina, India, and Zimbabwe. However, “silesaurids”, an older and enigmatic group of quadrupedal dinosauromorphs, were recently found, in some works, as stem ornithischians. In this scenario, dinosaurs would have originated far earlier than the end of the Ladinian, a time in which “silesaurids” are already spread through Gondwana. Despite being also recorded in more recent dinosaur-bearing beds in Brazil, “silesaurids” are absent in strata from the early Carnian, an important time frame for dinosaur evolution. Here we present a new “silesaurid”, Itaguyra occulta gen. et sp. nov., that fills up the remaining gap of occurrence of these dinosauromorphs and provides new clues to the success of these putative early ornithischians.
Similar content being viewed by others

Introduction
The Middle-Upper Triassic of South America yields an exceptional record of early dinosaurs and their akins1–5. South American Triassic avemetatarsalians are referred to three main lineages: Aphanosauria, represented by Spondylosoma absconditum6,7; Pterosauromorpha, represented by lagerpetids8–10; and Dinosauromorpha, represented by dinosaurs and their close relatives8,11–20. At least seven dinosauromorphs from South America have already been considered “silesaurids”1,9,19,21–28, a diverse grade of Triassic faunivorous to herbivorous dinosauromorphs, also found in several units of Europe29,30, North America31,32, and Africa33–36. Typical “silesaurids” were found as a monophyletic group and sister-clade to Dinosauria in several studies3,10,32,35,35,36,37,38,39,40,41,42, with some exceptions1,9,18,21. More recently, they were recovered within the Dinosauria, as early-divergent members of Ornithischia19,20,24–27,43. Throughout the text we will treat them as “silesaurids” to indicate its instability as a clade25.
In South America, “silesaurids” and other disputed early ornithischians are recorded in the upper Carnian to lower Norian Ischigualasto Formation (Ignotosaurus fragilis17 and Pisanosaurus mertii12, at least the first from the bottom layers of this unit17) of Argentina, and in in the late Carnian to early Norian Candelária Sequence in Brazil (from the base of this Sequence: Amanasaurus nesbitti20 and two unnamed “silesaurids”, here referred as Alemoa44 and Faixa Nova45 “silesaurids”; and from the upper layers: Sacisaurus agudoensis). The oldest members of the group are found in the Pinheiros-Chiniquá Sequence (Ladinian-?earliest Carnian) of Brazil, represented by Gamatavus antiquus19 and Gondwanax paraisensis28; and the record of Lewisuchus admixtus from the Chañares Formation of Argentina22,23,46, though its affinities with silesaurids are disputed by some authors5. We report in this study the first dinosauromorph remains from the lower Carnian Santa Cruz Sequence (Santacruzodon Assemblage Zone) of Brazil and assign them to a new “silesaurid” (Fig. 1). The Santacruzodon Assemblage Zone (AZ) represents an intermediate interval between the oldest South American “silesaurids” and the more recent ones, filling this gap regarding this moment of the evolutionary history of the group.

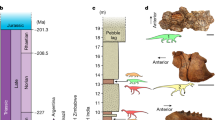
The new “silesaurid” Itaguyra occulta gen. et sp. nov. (A) Holotype UFRGS-PV-1365-T. (B) Placement of Rio Grande do Sul State (RS) in Brazil. (C) Exposure of the Santa Maria Supersequence in RS. (D) Locality of the type material (red star) and other correlated localities (pink stars). (E) Restoration of the skeleton (artwork by MG) with known elements in pink.
Geological settings
Triassic deposits of the Paraná Basin in Brazil are found only in the Rio Grande do Sul State (Fig. 1) and are recognized as two second-order sequences, the Sanga do Cabral Supersequence (Induan-Olenekian) and the Santa Maria Supersequence (Ladinian-Rhaetian)47–49. The Middle-Upper Triassic Santa Maria Supersequence is subdivided into four third-order sequences, the first three being characterized by a distinctive faunal association4,47–49. The Ladinian-?lower Carnian Pinheiros-Chiniquá Sequence is characterized by the Dinodontosaurus AZ, in which the tusked-dicynodont Dinodontosaurus abound, associated with traversodontid cynodonts and large carnivorous pseudosuchians4; the lower Carnian Santa Cruz Sequence has a faunal content dominated by a great abundance of traversodontid cynodonts defining the Santacruzodon AZ4,50; the upper Carnian to Norian Candelária Sequence, by its turn, has a faunal content characterized by two different associations, the lower Hyperodapedon AZ (late Carnian), characterized by the abundance of hyperodapedontine rhynchosaurs, associated with herrerasaur and sauropodormorph dinosaurs4,51; and the Riograndia AZ in the uppermost portion (early Norian), which reveal a faunal content of larger sauropodomorph dinosaurs, small non-mammaliaform prozostrodontian cynodonts and sphenodonts4,50. The youngest Mata Sequence lacks vertebrate fossils and is probably Norian to Rhaetian in age4,48,49.
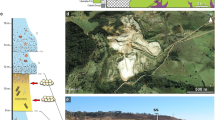
The Santa Cruz Sequence (SCS) presents a general trend from ephemeral river to overlaying loess deposits47,52,53, deposited in semi-arid to arid climate conditions in an ephemeral fluvial environment, with predominance of aeolian deposition and occasional seasonal climatic variations52,54,55. Based on LA-MC-ICPMS U-Pb detrital zircon data, the SCS maximum depositional age is 236 ± 1.5 Ma56. This sequence, and the faunal content that distinguishes it, is geographically restricted to a few kilometers, between the municipalities of Venâncio Aires, Vera Cruz and Santa Cruz do Sul47,56,57 (Fig. 1). The faunal content is represented by dicynodonts, traversodontid and probainognathian cynodonts50,57,58,59,60,61, paracrocodylomorphs62, proterochampsids63,64,65, and recently by the occurrence of rhynchosaurs66 and procolophonoids67. Based on taphonomic evidence and stratigraphic distribution of the traversodontid remains from the Schoenstatt locality, a possible sub-zonation within the Santacruzodon AZ was proposed67,68, but further studies are required to properly characterize it.
The Schoenstatt Sanctuary, an outcrop with a thickness of about 20 m consisting mainly of massive mudstones57, is the richest in terms of diversity57,58,59,60,61,62,63,64,65,66,67,68,69. The specimens from this outcrop are relatively well preserved exhibiting external carbonate concretions and an internal silica enrichment71, with some also showing a degree of deformation, mainly due to the overlying sedimentary load61,63. The SCS represents an important episode of the Santa Maria Supersequence as it does not present a proper correlate with the Ischigualasto-Villa Unión Basin in Argentina, being deposited probably around the same time of the Los Rastros Formation, which lack tetrapod body fossils (fossils from this units include plants, bivalves, crustaceans, insects, fishes, and tetrapod footprints; e.g.71,72). However, recent findings of the traversodontid Menadon remains from the upper most portion of the Argentinean Chañares Formation would extend the temporal correlation between the part of the Santa Cruz Sequence and the Argentine deposits67,73.
Institutional abbreviations: DMNH, Denver Museum of Nature and Science, Denver, USA; GR, Ghost Ranch Ruth Hall Museum of Paleontology, Abiquiu, USA; UFRGS-PV-T, Triassic vertebrate collection of the Museu de Paleontologia da UFRGS Irajá Damiani Pinto, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil; UFSM, Universidade Federal de Santa Maria, Santa Maria, Brazil; ZPAL AbIII, Institute of Paleobiology of the Polish Academy of Sciences, Warsaw, Poland.
Systematic paleontology
ARCHOSAURIA Cope, 1869 (sensu Gauthier and Padian, 1985).
AVEMETATARSALIA Gauthier, 1986 (sensu Sereno, 1991).
DINOSAUROMORPHA Benton, 1985 (sensu Ezcurra et al. 2020).
Itaguyra occulta gen. nov. et sp. nov.
[urn: lsid: zoobank.org: act: 7 A382537-0 ADC-47 FB-8344-4D02E25306E6 (genus)]
[urn: lsid: zoobank.org: act: AD1 FCA98-574B-4317–889 C-E92EED1 A7 A4 C (species)]
Etymology
The genus name combines the native Tupi words Ita- (= stone) and -guyra (= bird), referring to the avemetatarsalian nature of the specimen. The specific epithet, derived from Latin, means hidden and refers to the fact that the remains were initially identified as being “mixed up” with other indeterminate cynodont fragments with which they had been collected.
Holotype
UFRGS-PV-1365(a)-T, a left ilium (Fig. 1A).
Paratype
UFRGS-PV-1365(b)-T, associated ischium (Fig. 1A).
Type and referred locality
The type-materials were collected in the Schoenstatt Sanctuary fossil site (52°27′0.5″ W; 29°44′26.2″ S), in the Santa Cruz do Sul municipality (Fig. 1D), Rio Grande do Sul State, Brazil.
Diagnosis
Itaguyra occulta differs from all other known “silesaurids” with comparable material in (*local autapomorphy): having a faint crest connecting the preacetabular process and the supracetabular crest of the ilium, unlike the prominent ridge of most “silesaurids”*; height of the postacetabular process sub-equal to the height of the acetabulum (shared with Silesaurus opolensis and Kwanasaurus williamparkeri), differing from Lutungutali sitwensis and Gamatavus antiquus (see Discussion); supracetabular crest rounded (shared with Asilisaurus kongwe, Lewisuchus admixtus, Lutungutali sitwensis, and Gamatavus antiquus), unlike the straight outline of Ignotosaurus fragilis, and Kwanasaurus williamparkeri; brevis fossa almost entire visible in lateral view (shared with Silesaurus opolensis Kwanasaurus williamparkeri), being different from Lewisuchus admixtus and Asilisaurus kongwe which are more ventrally oriented; presence of a triangular process continuous to the posterior margin of the brevis fossa (absent in Gamatavus antiquus); triangular ventral margin of the medial wall of the iliac acetabulum (shared with Asilisaurus kongwe, Lutungutali sitwensis, Ignotosaurus fragilis and probably Gamatavus antiquus, but not with Silesaurus opolensis and Kwanasaurus williamparkeri); and, tall shaft of the ischium* (see Discussion).
Description
Ilium
The left ilium is partially preserved (Fig. 2), missing the preacetabular process and the central dorsal portion of the iliac blade. In addition, there are signs of erosion over the lateral surface of the posterior end. It resembles the morphology of the saddle-shaped ilia of most “silesaurids” (Fig. 3A), like in Silesaurus opolensis (Fig. 3H), Lutungutali sitwensis (Fig. 3F), Ignotosaurus fragilis (Fig. 3G), and Gamatavus antiquus (Fig. 3B)17,19,29,34. The preserved portion of the iliac blade is strongly inclined medially, resembling the condition seen in some specimens of Silesaurus opolensis as well as in Kwanasaurus williamparkeri, Ignotosaurus fragilis, and Gamatavus antiquus17,19,29,32. The morphology of the iliac blade can be, however, subject to some intraspecific variation, as observed in Silesaurus opolensis74, and its shape may be strongly controlled by taphonomic compression, as seen in other dinosauromorphs75, and in other specimens from the same locality65. Although the full dorsal extension of the iliac blade is inaccessible, the posterior portion preserves remnants of the central portion, suggesting that the dorsal limit is lower than the dorsal margin of the posterior end, as in other “silesaurids”. This would mean that the iliac blade represents approximately 0.94 of the dorsoventral height of the iliac portion of the acetabulum, a condition compatible with other “silesaurids”, but not with the 1.43 of Lutungutali sitwensis, which is considered as an autapomorphy of this species34.

Ilium of Itaguyra occulta gen. et sp. nov. Left ilium in lateral (A), posterior (B), anterior (D) and medial (E). Schematic drawings of the lateral (C) and medial (F) views. Abbreviations: ac, acetabulum; bs, brevis shelf; bf, brevis fossa; cst, crest; isp, ischiadic peduncle; pra, preacetabular process; poa, postacetabular process; pup, pubic peduncle; sac, supracetabular crest; sra, sacral rib scars.

Comparative schematic drawings of ilia in lateral view of “silesaur” and target taxa with highlighted shared main features with Itaguyra occulta: concave supracetabular crest (yellow circle), preacetabular crest (purple circle), laterally oriented brevis fossa (blue circle) and triangular process of the brevis fossa (red circle). (A) Itaguyra occulta; (B) Gamatavus antiquus; (C) Sacisaurus agudoensis; (D) Asilisaurus kongwe; (E) Lewisuchus admixtus; (F) Lutungutali sitwensis; (G) Ignotosaurus fragilis; (H) Silesaurus opolensis; (I) Kwanasaurus williamparkeri; (J) Indeterminate silesaur GR 225; (K) Teleocrater rhadinus; (L) Mambachiton fiandohana. Elements are not to scale.
The basal portion of preacetabular process of the iliac blade is thick and bears a faint crest (Fig. 1C), implanted near the second third of the acetabulum length. This crest seems to be homologous to the prominent one of most “silesaurids”1,32,35,44 and the aphanosaur Teleocrater rhadinus, being absent in Mambachiton fiandohana and lagerpetids (Fig. 3)74,76. The supracetabular crest is laterally projected as in most “silesaurids”1,17,29, not lateroventrally as in Lutungutali sitwensis34. The supraacetabular crest is curved anteriorly (Fig. 3A), similar to Lewisuchus admixtus3 (Fig. 3E) and Gamatavus antiquus (Fig. 3B)19, but contrasts with the more anterodorsally straight crest of Lutungutali sitwensis (Fig. 3F), Ignotosaurus fragilis (Fig. 3G) and Kwanasaurus williamparkeri (Fig. 3I)17,32,34. It extends without interruption toward the pubic peduncle and its posterior portion is straight, fading posteriorly at the level of the ischiadic peduncle without connecting to it. The iliac portion of the acetabulum is sub-elliptical, in lateral view, being antero-posteriorly longer than high. Similar to Lutungutali sitwensis and Kwanasaurus williamparkeri32,34 there is no trace of an antitrochanter, which is observed in Silesaurus29.
As in most “silesaurids” the ventral portion of the acetabulum is triangular in lateral view (Fig. 2), being distinct from the concave form of Kwanasaurus williamparkeri32. The ventral margin of the posterior half of the medial wall lacks a notch, whereas this feature occurs in Asilisaurus kognwe, Gamatavus antiquus, Gondwanax paraisensis, and Ignotosaurus fragilis17,19,28,35. The pubic peduncle is thicker than the ischiadic peduncle, being long and stout, with a subtriangular shape in anterior view (Fig. 2D). In lateral view, its articular facet is continuous with the anteroventral margin of the acetabulum, giving it a more ventral orientation to the pubic peduncle, like in Asilisaurus kongwe and GR 225, unlike the flexed ones of most “silesaurids”. It contributes less than half the length of the contact with the pubis, as in Gamatavus antiquus and Sacisaurus agudoensis, unlike the prominent condition observed in Lutungutali sitwensis and Ignotosaurus fragilis. The ischiadic peduncle is drop-shaped in ventral view, being slightly lateroventrally projected (Fig. 2), but lacks any “lip-like” posterior projection. The absence of a posterior projection results in a more straight margin, differing the specimen from the more concave margin observed in Lutungutali sitwensis and Gamatavus antiquus. Interestingly, the contact with the pubis represents 0.81 of the contact with the ischium as in Ignotosaurus fragilis and Gamatavus antiquus, unlike Asilisaurus kongwe (0.70) and Lutungutali sitwensis (0.75).
The postacetabular portion of the iliac blade is posterodorsally projected extending well beyond the ischiadic peduncle (Fig. 2). In lateral view, a mid-crest develops toward the posterior end in the postacetabular process, increasing the thickness of the iliac blade, although its lateral morphology is somewhat eroded posteriorly, precluding us from determining the morphology of the area of attachment of the flexor tibialis musculature, widely seen in dinosauromorphs17,29. This crest, which we interpret as the anteriormost portion of the brevis shelf, does not contact the supracetabular crest anteriorly (unlike the Alemoa silesaur, Ignotosaurus fragilis, and Kwanasaurus williamparkeri). Posteriorly, the crest runs till the preserved end of the iliac blade resulting in a sub-triangular end in posterior view (Fig. 2B). The brevis shelf limits a shallow fossa present at the posteroventral region of the postacetabular process. This represents the brevis fossa, which is hypothesized as the attachment site for the caudofemoralis brevis muscle in avemetatarsalians (e.g.77). The brevis shelf is not as pronounced as in Kwanasaurus williamparkeri(e.g.32, DMNH EPV.48506), Gamatavus antiquus19 or in Lewisuchus admixtus41, being similar to that of Silesaurus opolensis (e.g.29, ZPAL AbIII 907/8 and 2517). Nevertheless, since the lateral surface of the shelf is not fully preserved in the new material, this condition should be considered with caution.
The brevis fossa in Itaguyra occulta is as laterally oriented (Fig. 2C) as in Silesaurus opolensis29 (ZPAL AbIII 907/8 and 2517, Fig. 3H), Sacisaurus agudoensis (Fig. 3C), and Kwanasaurus williamparkeri32 (DMNH EPV.48506, Fig. 3I), being different from Lewisuchus admixtus (Fig. 3E)41 and Asilisaurus kongwe35 (Fig. 3D), which have more ventrally oriented fossae. Either way, the brevis shelf and fossa of Itaguyra occulta is more evident than in Teleocrater rhadinus or in Mambachiton fiandohana. The brevis fossa of Itaguyra occulta shows a well-developed medial wall with a distinct triangular process, differing from lagerpetids, Asilisaurus kongwe (Fig. 3D) and Gamatavus antiquus (Fig. 3B)19,35,41. The presence of this triangular process is shared with Lewisuchus admixtus (Fig. 3E), Ignotosaurus fragilis (Fig. 3G), Kwanasaurus williamparkeri (Fig. 3I) and Silesaurus opolensis (Fig. 3H)17,29,32,46.
On the medial surface of the ilium (Fig. 2E and F), two main sacral rib attachment sites for the primordial sacrals can be discerned as distinct fossae and ventral rugosities (Fig. 2). The anteriormost is smaller and more circular, almost reaching the anterior end of the ilium. A dorsoventral ridge separates both sacral attachment sites, being the second more ellipsoid, continuing and expanding ventrally till the posterior ventral margin projection of the postacetabular process. The morphology of the sacral rib attachment resembles that of Asilisaurus kongwe35, suggesting the presence of two sacral ribs as in most “silesaurids”19,34,35,41, but contrasting with the three or four estimated for Silesaurus opolensis77, Gondwanax paraisensis28, Kwanasaurus williamparkeri32, and Pisanosaurus mertii23. The medial ridge that bounds dorsally the attachment of the second sacral is very faint in Itaguyra occulta. This poor medial development of the ridge contributes to the brevis fossa having almost no ventral orientation, contrasting to other “silesaurids”19,29,35,41.
Ischium
The left ischium preserves the entire ischial shaft (Fig. 4), but misses part of the proximal region, including most of the obturator plate and the region that contacted the pubis. It is slightly longer than the preserved iliac length, and would probably be longer than the entire iliac blade if it was complete. The articular surface that received the ilium is somewhat eroded, but it is the lateromedially thickest portion of the ischium. The acetabular rim is damaged in the specimen and its morphology cannot be securely assessed. A faint depression anterior to the iliac articular facet may represent the fossa (Fig. 4C) that, in other dinosauromorphs, separates the iliac and pubic articular facets of the ischium34,35.

Ischium of Itaguyra occulta gen. et sp. nov. Left ischium in lateral (A), ventral (B), dorsal (D), medial (E) and posterior (G) views. Schematic drawings of the lateral (C) and medial (F) views. Abbreviations: dmr, dorsal ridge; ex, expansion; fil, facet for the illium; fos, fossa; rug, rugosities.
The ischial shaft is oriented posteroventraly, with the shaft reducing the height relative to the proximal portion in its initial third, and maintaining subparallel dorsal and ventral margins for most of its length, except for a slight distal expansion. The dorsal groove is shallow in Itaguyra occulta, differing from Lutungutali sitwensis. The groove is elongated, reaching the posterior half of the shaft, andmedially bounded by an elongated ridge. The medial surface of the shaft lacks marked rugosities. The shaft is tall, similar to Lewisuchus admixtus46, but differing from Gondwanax paraisensis, Silesaurus opolensis, Lutungutali sitwensis, and Sacisaurus agudoensis21,28,29,34. Distally, the shaft expands dorsoventrally and slightly laterally (Fig. 3), being ellipsoidal in distal view (Fig. 4G). In lateral view, the distal end is almost squared, resembling the condition of Silesaurus opolensis29; ZPAL AbIII 4004/7). Differing from Gondwanax paraisensis, Silesaurus opolensis, and Lewisuchus admixtus, the ischium of Itaguyra occulta is remarkable for its short proximodistal length. This, combined with the aforementioned tall shaft, results in a unique morphology for the specimen.
Phylogenetic analysis
The phylogenetic analysis recovered 144 MPTs of 1082 steps (CI = 0.296; RI = 0.690). The general topology of the strict consensus tree follows the same pattern as that recovered by Müller28 regarding the position of the main groups (Fig. 5), where Dinosauria is composed of a traditional Saurischia (Theropoda plus Sauropodomorpha) and Ornithischia. The latter includes “silesaurids” as early diverging members in a paraphyletic array, following previous iterations of this data matrix19,24,25and other independent investigations26,27. Itaguyra occulta nests as one of the earliest members of Ornithischia, within traditional “silesaurids”. Whereas the new species nests in a basal position (Fig. 5), the basalmost member of the clade is a node supporting Lewisuchus admixtus plus Saltopus elginensis. This node is the sister taxa to the clade that includes Itaguyra occulta and the other ornithischians. The position of Itaguyra occulta is supported by the angle of the pubic peduncle of the ilium (character 186: 1→2), whereas it is excluded from the more derived nodes by the presence of a short postacetabular ala of the ilium (state 0 of character 183). This subsequent clade includes Asilisaurus kongwe plus Sulcimentisauria. As in the former analysis, Gamatavus antiquus nests as the early divergent member of Sulcimentisauria according to the absence of a femoral trochanteric shelf (character 214: 1→0). Regarding the position of Mambachiton fiandohana, the taxon is nested within Aphanosauria. This alternative position compared to the original study78 likely reflects the nature of the data matrix, which is designed to investigate affinities within Dinosauromorpha rather than among more early divergent groups of Avemetatarsalians.

Time-calibrated strict consensus tree depicting the phylogenetic position of Itaguyra occulta gen. et sp. nov. Silhouettes based on the artwork of Matheus Fernandes Gadelha and MG.
Discussion
Avemetatarsalians are found in several Middle and lower Upper Triassic units of South America3,9,13,17,19,20,41, Europe29,30, North America31,32 and Africa34–36,42,78,79. Most of these records are representatives of “silesaurids”, a group known in South America by the Brazilian Gamatavus antiquus and Gondwanax paraisensis (Pinheiros-Chiniquá Sequence, Ladinian to ?early Carnian)19,28, Amanasaurus nesbitti (lower Candelária Sequence, late Carnian)20, and Sacisaurus agudoensis (upper Candelária Sequence, Norian)1; and the Argentinean Lewisuchus admixtus (Chañares Formation, early Carnian)8, Ignotosaurus fragilis (Ischigualasto Formation, late Carnian to early Norian)17, and possibly Pisanosaurus mertii (Ischigualasto Formation, late Carnian to early Norian)12.
The early to middle Carnian “silesaurid” Itaguyra occulta described here from the Santa Cruz Sequence of Brazil fulfills a time gap for the record of the group worldwide (Fig. 6). It is putatively younger than the Middle Triassic forms (e.g., Asilisaurus kongwe, Gamatavus antiquus, Gondwanax paraisensis, Lewisuchus admixtus, and Lutungutali sitwensis) and older than all late Carnian/Norian “silesaurid” records (e.g., Amanasaurus nesbitti, Eocoelophysis baldwini, Kwanasaurus williamparkeri, Sacisaurus agudoensis, Silesaurus opolensis, and Technosaurus smalli). The record of Itaguyra occulta in the Santa Cruz Sequence demonstrates an uninterrupted presence of “silesaurids” in the region that corresponds to present-day southern Brazil (Fig. 7), within the available Triassic units of South America (Fig. 6), spanning a time interval of approximately 11 million years, from the Ladinian to the early Norian.

Worldwide record of “silesaurids” through the Triassic. Paleogeographic maps of the world during the Anisan to Norian, showing the occurrences of ‘silesaurids’. Silhouettes not to scale, redrawn from different sources. Silhouettes based on the artwork of Matheus Fernandes Gadelha, MG and VDPN.

Faunal temporal distribution of the Santa Maria Supersequence Assemblage zones51,56,80. Silhouettes not to scale, redrawn from different sources. The upper limits of the temporal range of traversodontids and rhynchosaurs are disputed by some authors81. Abbreviatons: AZ, Assemblage Zone; CS, Candelária Sequence; SCS, Santa Cruz Sequence; PCS, Pinheiro-Chiniquá Sequence.
Therefore, “silesaurids” persisted in this region even during drastic faunal changes: (i) the group lived in therapsid-dominated assemblages (i.e., Dinodontosaurus AZ, Santacruzodon AZ), side-by-side with giant top-tier pseudosuchian predators63,82 and a wide array of cynodonts61,69,82,83 (Fig. 7). Dinosauromorphs were rare components of these assemblages according to the fossil record, restricted to “silesaurids”19,28 (this study) and indeterminate remains 84; (ii) during the late Carnian, a drastic faunal change is documented in the fossil record of the Santa Maria Supersequence, with tusked dicynodonts disappearing and rhynchosaurs becoming the dominant terrestrial tetrapods4,85. These late Carnian assemblages are part of the Hyperodapedon AZ and represent the dawn of the dinosaur age5. The fossil record of dinosauromorphs from the Hyperodapedon AZ is much more diverse than the record from the older AZs. In addition to “silesaurids”20,45 these beds yielded abundant records of early sauropodomorph9,15,86–88 and herrerasaurian dinosaurs13,18,89,90. Despite the faunistic turnover, “silesaurids”, either being a clade or early-ornithischians, persisted within these assemblages (Fig. 7), something not common to all groups (like the disrupted distribution of dicynodonts and procolophonids). The continuous presence of these enigmatic dinosauromorphs over this relatively long time interval indicates a high capacity for adaptation to different ecosystems. Indeed, “silesaurids” have been recovered from the Upper Triassic beds of Brazil1,91, which correspond to the upper portion of the Candelária Sequence48, a moment associated with another documented faunistic turnover4. The drastic faunal changes of the Santa Maria Supersequence are reflections of climatic and floristic changes which have affected specialized herbivorous animals like dicynodonts and rhynchosaurs. The presence of “silesaurids” in the four biozones may suggest that these changes did not affect the geographical distribution of the group in this region.
A number of studies have supported the hypothesis that “silesaurids” may have been the first ornithischian dinosaurs9,18,21,24–27. Based on this scenario, the earliest dinosaurs lived as discrete components in ecosystems dominated by other tetrapods for several million years, with their first marked expansion in diversity occurring during the early Carnian, a time when saurischian dinosaurs branched into three main lineages (e.g., Herrerasauria, Theropoda, and Sauropodomorpha)5. On the other hand, considering the scenario that places “silesaurids” as the dinosaurian sister-group, it is remarkable that dinosaur precursors persisted millions of years before and after the first dinosaurian radiation32,33,92. Despite the phylogenetic scenario, the new record from the SCS provides compelling evidence of the adaptive capabilities of “silesaurids”, as they were components of the ecosystems immediately preceding the faunistic turnover that witnessed the emergence of “typical” dinosaurs.
Methods
Material
The specimen UFRGS-PV-1365-T consists of two associated elements of compatible size (Fig. 1), an ilium (a) and an ischium (b), collected at the Schoenstatt outcrop but lacking any stratigraphic information, being probably already exhumed when collected. It was previously cataloged as “indeterminate” and has been recently attributed to Archosauriformes (“new indet. 2”)68. The precise collection point and stratigraphic level of the UFRGS-PV-1365-T specimen is unfortunately unknown, however, based on the presence of root ‘hair’ petrifactions93on the bone surfaces, UFRGS-PV-1365-T is tentatively placed in the central to upper portion of the biostratigraphic scheme for the site68, thus belonging to Menadon or Santacruzodon subzones. Due to their similar size, shared features with avemetatarsalians, and their state of preservation, the elements here are treated as belonging to the same taxon. The elements were described following the archosaur literature with main comparisons with other avemetatarsalians.
Phylogenetic analysis
The phylogenetic affinities of UFRGS-PV-1365-T were investigated using an updated version of the data matrix published by Müller28. We inserted the early avemetatarsalian Mambachiton fiandohana79 and removed Soumyasaurus aenigmaticus94 from the data matrix. The exclusion of the latter operational taxonomic unit (OTU) is based on the fragmentary nature of the specimen, which hampers a proper assignment to “Silesauridae” or related groups, especially because the presence of ankylothecodonty appears more widely spread among putative coeval groups, such as allokotosaurians95. The final data matrix includes 282 morphological characters and 74 OTUs. The specimen UFRGS-PV-1365-T was scored for 7.1% of the characters in the matrix. It was processed in the software TNT v. 1.596 in order to recover the most parsimonious trees. Following the former study, all characters received the same weight and characters 4, 13, 18, 25, 63, 82, 84, 87, 89, 109, 142, 166, 174, 175, 184, 186, 190, 201, 203, 205, 209, 212, 225, 235, 236, 239, 250 and 256 were treated as additive (ordered). Euparkeria was used to root the most parsimonious trees (MPTs), which were recovered employing New Technology Search Algorithm, searching for a minimum length 100 times with the default Ratcheting, Drift, and Tree-fusing parameters. Topologies retained as replicates were branch-swapped for MPTs using TBR. The strict consensus tree was constructed using all trees recovered in the analysis and included all OTUs.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the International Code of Zoological Nomenclature. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix ‘http://zoobank.org/’. The LSID for this publication is: AD1 FCA98-574B-4317–889 C-E92EED1 A7 A4 C.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Ferigolo, J. & Langer, M. C. A late triassic dinosauriform from South Brazil and the origin of the ornithischian predentary bone. Hist. Biol. 19(1), 23–33 (2007).
Garcia, M. S., Fonseca, A. O., Doering, M., da Rosa, Á. A. & Müller, R. T. A new sympatric occurrence of lagerpetids (Pan-Aves, Pterosauromorpha) in the upper triassic of Southern Brazil. J. South. Am. Earth Sci. 140, 104897 (2024).
Ezcurra, M. D., Nesbitt, S. J., Fiorelli, L. E. & Desojo, J. B. New specimen sheds light on the anatomy and taxonomy of the early late triassic dinosauriforms from the Chañares formation, NW Argentina. Anat. Rec. 303(5), 1393–1438 (2020).
Schultz, C. L. et al. Triassic faunal successions of the Paraná basin, Southern Brazil. J. South. Am. Earth Sci. 104, 102846 (2020).
Novas, F. E. et al. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. J. South. Am. Earth Sci. 110, 103341 (2021).
von Huene, F. Die Fossilen Reptilien Des Südamerikanischen Gondwanalandes 342 (C.H. Beck, Munich, 1942).
Nesbitt, S. J., Butler, R. J., Ezcurra, M. D., Charig, A. J. & Barrett, P. M. The anatomy of Teleocrater rhadinus, an early avemetatarsalian from the lower portion of the Lifua Member of the Manda Beds ( Middle Triassic) in Vertebrate and climatic evolution in the Triassic rift basins of Tanzania and Zambia (eds. Sidor, C. A. & Nesbitt, S. J.). J. Vertebr. Paleontol. 37(sup1), 142–177 (Society of Vertebrate Paleontology, Memoir 17, 2018).
Romer, A. S. The Chañares (Argentina) triassic reptile fauna. X. Two new but incompletely known long–limbed pseudosuchians. Breviora 378, 1–10 (1971).
Cabreira, S. F. et al. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Curr. Biol. 26, 3090–3095 (2016).
Müller, R. T. et al. New reptile shows dinosaurs and pterosaurs evolved among diverse precursors. Nature 620, 589–594 (2023).
Reig, O. A. La presencia de dinosaurios saurisquios en los estratos de Ischigualasto (Mesotriasico Superior) de Las provincias de San Juan y La Rioja (Argentina). Ameghiniana 3(1), 3–20 (1963).
Casamiquela, R. M. Un nuevo dinosaurio ornitisquio triásico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto, Argentina. Ameghiniana 5(2), 47–64 (1967).
Colbert, E. H. A saurischian dinosaur from the Triassic of Brazil. Am. Mus. Novit 2405, 1–60 (1970).
Sereno, P. C., Forster, C. A., Rogers, R. R. & Monetta, A. M. Primitive dinosaur skeleton from Argentina and the early evolution of dinosauria. Nature 361, 64–66 (1993).
Langer, M. C., Abdala, F., Richter, M. & Benton, M. J. A sauropodomorph dinosaur from the Upper Triassic (Carnian) of Southern Brazil. C R Acad. Sci. Ser. IIa: Sci. Terre Planet. 329(7), 511–517 (1999).
Ezcurra, M. D. A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: A reassessment of dinosaur origin and phylogeny. J. Syst. Palaeontol. 8(3), 371–425 (2010).
Martínez, R. N. et al. Vertebrate succession in the Ischigualasto Formation. J. Vertebr Paleontol. 32, 10–30 (2013).
Pacheco, C. et al. Gnathovorax cabreirai: a new early dinosaur and the origin and initial radiation of predatory dinosaurs. PeerJ 7, e7963 (2019).
Pretto, F. A. et al. The oldest South American silesaurid: new remains from the Middle Triassic (Pinheiros-Chiniquá sequence, Dinodontosaurus Assemblage Zone) increase the time range of silesaurid fossil record in Southern Brazil. J. South. Am. Earth Sci. 120, 104039 (2022).
Müller, R. T. & Garcia, M. S. A new silesaurid from Carnian beds of Brazil fills a gap in the radiation of avian line archosaurs. Sci. Rep. 13(1), 4981 (2023).
Langer, M. C. & Ferigolo, J. The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities in Anatomy, Phylogeny, and Palaeobiology of Early Archosaurs and their Kin (eds. Nesbitt, S. J., Desojo, J.B. & Irmis, R. B.) 353–392 (Geological Society, London, Special Volume 379, (2013).
Bittencourt, J. S., Arcucci, A. B., Marsicano, C. A. & Langer, M. C. Osteology of the middle triassic archosaur Lewisuchus admixtus Romer (Chañares formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs. J. Syst. Palaeontol. 13,189 – 219 (2014).
Agnolín, F. L. & Rozadilla, S. Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina. J. Syst. Palaeontol. 16(10), 853–879 (2018).
Müller, R. T. & Garcia, M. S. A paraphyletic ‘silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biol. Lett. 16(8), 20200417 (2020).
Norman, D. B., Baron, M. G., Garcia, M. S. & Müller, R. T. Taxonomic, Palaeobiological and evolutionary implications of a phylogenetic hypothesis for ornithischia (Archosauria: Dinosauria). Zool. J. Linn. Soc. 196(4), 1273–1309 (2022).
Cau, A. A unified framework for predatory dinosaur macroevolution. Boll Soc. Paleontol. Ital. 63(1), 1–19 (2024).
Fonseca, A. O. et al. R. T. A comprehensive phylogenetic analysis on early ornithischian evolution. J. Syst. Palaeontol. 22(1), 2346577 (2024).
Müller, R. T. A new silesaurid from the oldest dinosauromorph-bearing beds of South America provides insights into the early evolution of bird-line archosaurs. Gondwana Res. 137, 13–28 (2025).
Dzik, J. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. J. Vertebr Paleontol. 23, 556–574 (2003).
Benton, M. J. & Walker, A. D. Saltopus, a dinosauriform from the upper triassic of Scotland. Earth Environ. Sci. Trans. 101(3–4), 285–299 (2011).
Ezcurra, M. D. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan and Lucas, 1999 from the upper triassic of new Mexico, USA. Geodiversitas 28, 649–684 (2006).
Martz, J. W. & Small, B. J. Non-dinosaurian dinosauromorphs from the Chinle formation (Upper Triassic) of the Eagle Basin, Northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7, e7551 (2019).
Nesbitt, S. J. et al. Ecologically distinct dinosaurian sister-group shows early diversification of ornithodira. Nature 464, 95–98 (2010).
Peecook, B. R. et al. D. A new silesaurid from the upper Ntawere formation of Zambia (Middle Triassic) is among the oldest bird-line archosaur (Avemetatarsalia). J. Vertebr Paleontol. 33, 1127–1137 (2013).
Nesbitt, S. J., Langer, M. C. & Ezcurra, M. D. The anatomy of Asilisaurus kongwe, a dinosauriform from the Lifua member of the Manda beds (middle Triassic) of Africa. Anat. Rec. 303(4), 813–873 (2020).
Kammerer, C. F., Nesbitt, S. J. & Shubin, N. H. The first basal dinosauriform (Silesauridae) from the Late Triassic of Morocco. Acta Palaeontol. Pol. 57, 277–284 (2012).
Langer, M. C., Ezcurra, M. D., Bittencourt, J. S. & Novas, F. E. The origin and early evolution of dinosaurs. Biol. Rev. 85, 55–110 (2010).
Nesbitt, S. J. The early evolution of archosauria: relationships and the origin of major clades. Bull. Am. Mus. Nat. Hist. 352, 1–292 (2011).
Martínez, R. N., Apaldetti, C., Correa, G. A. & Abelín, D. A Norian lagerpetid dinosauromorph from the Quebrada del Barro formation, Northwestern Argentina. Ameghiniana 53(1), 1–13 (2016).
Ezcurra, M. D. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4, e1778 (2016).
Ezcurra, M. D. et al. Enigmatic dinosaur precursors Bridge the gap to the origin of pterosauria. Nature 588(7838), 445–449 (2020).
Kammerer, C. F., Nesbitt, S. J., Flynn, J. J., Ranivoharimanana, L. & Wyss, A. R. A tiny ornithodiran archosaur from the triassic of Madagascar and the role of miniaturization in dinosaur and pterosaur ancestry. Proc. Natl. Acad. Sci. 117(30), 17932–17936 (2020).
Norman et al. The origin and early evolution of the ornithischian Bauplan: evaluation and implications for the dinosauria. Rev. Paléobiol 43, 61–832024 (2024).
Mestriner, G., Marsola, J. C., Nesbitt, S. J., Da-Rosa, Á. A. S. & Langer, M. Anatomy and phylogenetic affinities of a new silesaurid assemblage from the carnian beds of South Brazil. J. Vertebr Paleontol. 43(1), e2232426 (2023).
Campo, M. L., da Silva, F. O., Paes-Neto, V. D., Ferigolo, J. & Ribeiro, A. M. Overview on the tetrapods from Faixa Nova-Cerrito I site (Hyperodapedon assemblage Zone), Upper Triassic of southernmost Brazil. Hist. Biol. 1–19. https://doi.org/10.1080/08912963.2024.2344791 (2024).
Agnolín, F., Brissón Egli, F., Ezcurra, M. D., Langer, M. C. & Novas, F. New specimens provide insights into the anatomy of the dinosauriform Lewisuchus admixtus Romer, 1972 from the upper triassic levels of the Chañares formation, NW Argentina. Anat. Rec. 305(5), 1119–1146 (2022).
Horn, B. L. D. et al. A new third-order sequence stratigraphic framework applied to the triassic of the Paraná basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. J. South. Am. Earth Sci. 55, 123–132 (2014).
Zerfass, H. et al. Sequence stratigraphy of continental triassic strata of southernmost Brazil: A contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sediment. Geol. 161(1–2), 85–105 (2003).
Zerfass, H., Chemale, F., Schultz, C. L. & Lavina, E. Tectonics and sedimentation in Southern South America during triassic. Sediment. Geol. 166(3–4), 265–292 (2004).
Soares, M. B., Abdala, F. & Bertoni-Machado, C. A sectorial toothed cynodont (Therapsida) from the triassic Santa Cruz do Sul fauna, Santa Maria formation, Southern Brazil. Geodiversitas 33(2), 265–278 (2011).
Langer, M. C., Ramezani & J. Da Rosa, Á. A. U-Pb age constraints on dinosaur rise from South Brazil. Gondwana Res. 57, 133–140 (2018).
Horn, B. L. D., Goldberg, K. & Schultz, C. L. A loess deposit in the late triassic of Southern Gondwana, and its significance to global paleoclimate. J. South. Am. Earth Sci. 81, 189–203 (2018).
Horn, B. L. D., Goldberg, K. & Schultz, C. L. Interpretation of massive sandstones in ephemeral fluvial settings: A case study from the upper Candelária sequence (Upper triassic, Paraná basin, Brazil). J. South. Am. Earth Sci. 81, 108–121 (2018).
Corecco, L., Pereira, V. P., Soares, M. B. & Schultz, C. L. Geochemical study of the vertebrate assemblage zones of the Santa Maria supersequence (Middle to late Triassic), Paraná Basin, Brazil. Braz J. Geol. 50(4), e20200014 (2020).
Mancuso, A. C., Horn, B. L. D., Benavente, C. A., Schultz, C. L. & Irmis, R. B. The paleoclimatic context for South American triassic vertebrate evolution. J. South. Am. Earth Sci. 110, 103321 (2021).
Philipp, R. P. et al. Middle Triassic SW Gondwana paleogeography and sedimentary dispersal revealed by integration of stratigraphy and U-Pb Zircon analysis: the Santa Cruz sequence, Paraná basin, Brazil. J. South. Am. Earth Sci. 88, 216–237 (2018).
Abdala, F., Ribeiro, A. M. & Schultz, C. L. A rich cynodont fauna of Santa Cruz do Sul, Santa Maria formation (Middle-Late Triassic), southern Brazil. N Jb Geol. Palaeont Mh 11, 669–687 (2001).
Abdala, F. & Ribeiro, A. M. A new traversodontid cynodont from the Santa Maria formation (Ladinian-Carnian) of Southern Brazil, with a phylogenetic analysis of Gondwanan traversodontids. Zool. J. Linn. Soc. 139(4), 529–545 (2003).
Martinelli, A. G., Soares, M. B. & Schwanke, C. Two new cynodonts (Therapsida) from the middle-early late triassic of Brazil and comments on South American probainognathians. PLoS ONE 11(10), e0162945 (2016).
Melo, T. P., Abdala, F. & Soares, M. B. The Malagasy cynodont Menadon besairiei (Cynodontia; Traversodontidae) in the Middle-Upper triassic of Brazil. J. Vertebr Paleontol. 35(6), e1002562. (2015).
Schmitt, M. R., Martinelli, A. G., Melo, T. P. & Soares, M. B. On the occurrence of the traversodontid Massetognathus ochagaviae (Synapsida, Cynodontia) in the early late triassic Santacruzodon assemblage zone (Santa Maria supersequence, Southern Brazil): Taxonomic and biostratigraphic implications. J. South. Am. Earth Sci. 93, 36–50 (2019).
Lacerda, M. B., Schultz, C. L. & Bertoni-Machado, C. First ‘rauisuchian’ archosaur (Pseudosuchia, Loricata) for the Middle Triassic Santacruzodon Assemblage Zone (Santa Maria Supersequence), Rio Grande do Sul State, Brazil. PLoS ONE 10(2), e0118563 (2015).
Raugust, T., Lacerda, M. & Schultz, C. L. The first occurrence of Chanaresuchus bonapartei Romer 1971 (Archosauriformes, Proterochampsia) of the Middle Triassic of Brazil from the Santacruzodon Assemblage Zone, Santa Maria Formation (Paraná Basin). in Anatomy, Phylogeny, and Palaeobiology of Early Archosaurs and their Kin (eds. Nesbitt, S. J., Desojo, J.B. &, Irmis, R. B.), 303–318 (Geological Society, London, Special Volume 379, 2013).
Ezcurra, M. D., Desojo, J. B. & Rauhut, O. W. Redescription and phylogenetic relationships of the proterochampsid Rhadinosuchus gracilis (Diapsida: Archosauriformes) from the early late triassic of Southern Brazil. Ameghiniana 52(4), 391–417 (2015).
Paes-Neto, V. D. et al. New Rhadinosuchine proterochampsids from the late Middle‐early late triassic of Southern Brazil enhance the diversity of archosauriforms. Anat. Rec. 307(4), 851–889 (2024).
Battista, F., Martinelli, A. G., Ribeiro, A. M., Andrade, M. B. & Schultz, C. L. First record of rhynchosaurs (Archosauromorpha: Rhynchosauria: Hyperodapedontinae) from the early Late Triassic Santacruzodon Assemblage Zone of the Santa Maria Supersequence, Brazil. Anat. Rec. (2024), in press.
Battista, F., Ribeiro, A. M., Melo, T. P. & Schultz, C. L. A reinterpretation of taphonomy and palaeoecology of the early carnian Santacruzodon assemblage zone type-locality (Santa Maria Supersequence), Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 639, 112080 (2024).
Melo, T. P., Martinelli, A. G. & Soares, M. B. New occurrences of massetognathine traversodontids and chiniquodontids (Synapsida, Cynodontia) from the early Late Triassic Santacruzodon assemblage zone (Santa Maria supersequence, Southern Brazil): Geographic and biostratigraphic implications. J. South. Am. Earth Sci. 115, 103757 (2022).
Battista, F., Ribeiro, A. M., Erthal, F. & Schultz, C. L. Post-collection taphonomy, sampling effects and the role of the collector in palaeontological collections: A case study from an early late triassic bone accumulation from southernmost Brazil. Acta Palaeontol. Pol. 68, 359–372 (2023).
Reichel, M., Schultz, C. L. & Pereira, V. P. Diagenetic pattern of vertebrate fossils from the traversodontidae biozone, Santa Maria formation (Triassic), Southern Brazil. Rev. Bras. Paleontol. 8(3), 173–180 (2005).
Melchor, R. N., Bellosi, E. S. & Genise, G. S. Invertebrate and vertebrate trace fossils from a triassic lacustrine delta: The Los Rastros formation, Ischigualasto provincial park, San Juan, Argentina. PE-APA 9 (1), 17–33 (2003).
López-Arbarello, A., Rogers, R. & Puerta, P. Freshwater actinopterygians of the Los Rastros formation (Triassic), Bermejo basin, Argentina. Foss Rec. 9, 238–258 (2006).
Martinelli, A. G. et al. First record of Menadon (Cynodontia: Traversodontidae) in the early Late Triassic Chañares Formation of the Ischigualasto-Villa Unión Basin, NW Argentina: Gondwanan biostratigraphic implications. in XII Simpósio Brasileiro de Paleontologia de Vertebrados – Boletim de Resumos (eds. Machado, A. F., Machado, E. F. & Bulsing, K. P.) 123 (2023).
Piechowski, R., Tałanda, M. & Dzik, J. Skeletal variation and ontogeny of the late triassic dinosauriform Silesaurus opolensis. J. Vertebr Paleontol. 34, 1383–1393 (2014).
Müller et al. Under pressure: effect of sedimentary compression on the Iliac morphology of early sauropodomorphs. J. South. Am. Earth Sci. 88, 345–351 (2018).
Bertoni-Machado, C. & Holz, M. Biogenic fossil concentration in fluvial settings: An example of a cynodont taphocoenosis from the middle triassic of Southern Brazil. Rev. Bras. Paleontol. 9(3), 273–282 (2006).
Nesbitt, S. J. et al. The earliest bird-line archosaurs and assembly of the dinosaur body plan. Nature 544, 484–487 (2017).
Nesbitt, S. J. et al. The earliest-diverging Avemetatarsalian: a new osteoderm-bearing taxon from the triassic (Earliest late triassic) of Madagascar and the composition of Avemetatarsalian assemblages prior to the radiation of dinosaurs. Zool. J. Linn. Soc. 199(2), 327–353 (2023).
Griffin, C. T. et al. Africa’s oldest dinosaurs reveal early suppression of dinosaur distribution. Nature 609, 313–319 (2022).
Roberto-Da-Silva, L., Müller, R. T., França, M. A. G. D., Cabreira, S. F. & Dias-Da-Silva An impressive skeleton of the giant top predator Prestosuchus chiniquensis (Pseudosuchia: Loricata) from the triassic of Southern Brazil, with phylogenetic remarks. Hist. Biol. 32(7), 976–995 (2020).
Philipp, R. P. et al. U-Pb Zircon geochronology of detrital and Ash fall deposits of the Southern Paraná basin: A contribution for provenance, tectonic evolution, and the paleogeography of the SW Gondwana. Geosciences 13(8), 225 (2023).
Reichel, M., Schultz, C. L. & Soares, M. B. A new traversodontid cynodont (Therapsida, Eucynodontia) from the middle triassic Santa Maria formation of Rio Grande do Sul, Brazil. Palaeontology 52 (1), 229–250 (2009).
Martinelli, A. G. et al. The African cynodont Aleodon (Cynodontia, Probainognathia) in the triassic of Southern Brazil and its biostratigraphic significance. PLoS ONE 12(6), e0177948 (2017).
Müller, R. T. & Garcia, M. S. Oldest dinosauromorph from South America and the early radiation of dinosaur precursors in Gondwana. Gondwana Res. 107, 42–48 (2022).
Langer, M. C., Ribeiro, A. M., Schultz, C. L. & Ferigolo, J. The continental tetrapod-bearing triassic of South Brazil. Bull. New. Mexico Mus. Nat. Hist. Sci. 41, 201–218 (2007).
Cabreira, S. F. et al. New stem-sauropodomorph (Dinosauria, Saurischia) from the triassic of Brazil. Naturwissenschaften 98, 1035–1040 (2011).
Pretto, F. A., Langer, M. C. & Schultz, C. L. A new dinosaur (Saurischia: Sauropodomorpha) from the late triassic of Brazil provides insights on the evolution of sauropodomorph body plan. Zool. J. Linn. Soc. 185(2), 388–416 (2019).
Langer, M. C. et al. The early radiation of sauropodomorphs in the carnian (Late Triassic) of South America. In South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution (eds Otero, A., Carballido, J. L., Pol, D. et al.) 1–49 (Springer, 2022).
Garcia, M. S., Müller, R. T., Pretto, F. A., Da-Rosa, Á. A. & Dias-Da-Silva, S. Taxonomic and phylogenetic reassessment of a large-bodied dinosaur from the earliest dinosaur-bearing beds (Carnian, upper Triassic) from Southern Brazil. J. Syst. Palaeontol. 19(1), 1–37 (2021).
Garcia, M. S., Cabreira, S. F., da Silva, L. R., Pretto, F. A. & Müller, R. T. A saurischian (Archosauria, Dinosauria) ilium from the upper triassic of Southern Brazil and the rise of herrerasauria. Anat. Rec. 307(4), 1011–1024 (2024).
Doering, M., Ezcurra, M. D., Schiefelbein, J. H., Garcia, M. S. & Müller, R. T. New archosauromorph remains provide data on the age of a unique late triassic assemblage from Southern Brazil. J. South. Am. Earth Sci. 145, 105046 (2024).
Irmis, R. B. et al. A late triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317, 358–361 (2007).
Klappa, C. F. Rhizoliths in terrestrial carbonates: classification, recognition, genesis and significance. Sedimentology 27, 613–629 (1980).
Sarigul, V., Agnolin, F. & Chatterjee, S. Description of a multitaxic bone assemblage from the upper triassic post quarry of Texas (Dockum Group), including a new small basal dinosauriform taxon. Historia Nat. 8(1), 5–24 (2018).
Marsh, A. D. et al. Puercosuchus traverorum n. gen. n. sp.: a new malerisaurine azendohsaurid (Archosauromorpha: Allokotosauria) from two monodominant bonebeds in the Chinle formation (Upper triassic, Norian) of Arizona. J. Paleontol. 96(S90), 1–39 (2022).
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
Acknowledgements
VDPN was supported by a postdoctoral grant Programa Pós-Doutorado Nota 10 (E-26/200.121/2024) provided by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro - FAPERJ. AGM was supported by PICT 2020-SERIEA-01498, CONICET PIBAA 1137, and MILENIO NCN2023-025. FB was supported by a postdoctoral grant Programa Institutos Nacionais de Ciência, Tecnologia e Inovação / Conselho Nacional de Desenvolvimento Científico e Tecnológico (INCT-PALEOVERT CNPq 153528/2024-9). We thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico for its support (CNPq 409318/2023-0 to FAP; CNPq 404095/2021-6 and 303034/2022-0 to RTM; CNPq #16811/2021-1 to FLP; CNPq #309463/2021-1 to HF; CNPq #406565/2021-0 for CLS; CNPq #406779/2021-0 and #308707/2023-0 to AWK; #308515/2023-4 to MBS). MG thanks the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES #88887.826787/2023–0). HF was supported by Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul - FAPERGS (#24/2551-0001306-7). AWAK and MBS acknowledge founding by Fundação de Desenvolvimento Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro - FAPERJ (#E-26/201.095/2022 to AWAK, and #E-26/204.181/2024 to MBS). We thank Benjamin T. Breeden III by sharing photographs of the indeterminate Ghost Ranch silesaur. Access to the free version of TNT 1.5 was possible owing to the Willi Hennig Society. This publication was made possible by the INCT-PALEOVERT CNPq project #406902/2022-4. We thank Federico Agnolín and an anonymous reviewer for the comments and suggestions that improved this manuscript.
Author information
Authors and Affiliations
Contributions
V.D.P.N data curation, formal analysis, investigation, writing and preparation of figures. F.A.P. formal analysis, investigation, writing and preparation of figures. A.G.M. data curation, formal analysis, investigation, writing. F.B. data curation, formal analysis, investigation, writing. M.G. formal analysis, investigation, writing and preparation of figures. R.T.M. formal analysis, investigation, writing and preparation of figures. M.R.S. investigation, writing and preparation of figures. T.P.M. investigation. H.F. data curation, writing. C.L.S. data curation, writing. M.B.S. investigation, writing. F.L.P. investigation, writing. A.W.K. funding acquisition, investigation, writing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Paes Neto, V.D., Pretto, F.A., Martinelli, A.G. et al. Continuous presence of dinosauromorphs in South America throughout the Middle to the Late Triassic. Sci Rep 15, 18498 (2025). https://doi.org/10.1038/s41598-025-99362-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-99362-5
