
Abstract
Understanding the occurrence of vagrant individuals away from their usual distribution range can shed light on animal navigation systems. In autumn, migratory Nearctic landbirds often occur as far as Western Europe, and while the link between these occurrences and wind conditions over the Atlantic Ocean is largely established, the drivers and natal origins of these transatlantic vagrants remain elusive. We conducted feather hydrogen isotopic (δ2Hf) analyses from 72 Nearctic individuals representing 26 landbird species captured in the Azores (Portugal), Iceland and France over the last century to infer their likely origins. While potential origins of several individuals came from northeastern Nearctic populations (e.g., Blackpoll Warbler, Northern Parula), we also found that some individuals likely originated from northwestern (e.g., American Redstart, Gray-cheeked Thrush) or southern (Yellow-billed Cuckoo) populations. Our results confirm that northeastern populations migrating along the coast or undertaking over-water flights are more likely to be displaced. However, they also show that even northwestern populations migrating southeast towards the eastern coast of North America have the potential to be displaced across the Atlantic Ocean. Further research is needed to fully understand the drivers of these transatlantic vagrancy events and to determine whether wind is the only factor that can push migratory individuals off their intended course. Overall, this study sheds light on the broad geographic origins of transoceanic vagrants and the potential implications for the colonization of new regions.
Similar content being viewed by others
Introduction
Due to the broadscale geographic movements and the distances they can travel to and from their breeding and non-breeding grounds1,2, migratory birds have fascinated mankind since ancient times3. Among these movements, the appearance of individuals far from their usual distributional ranges, a phenomenon called vagrancy4, still amazes both scientists and amateur ornithologists5,6,7,8,9. Because instances of vagrancy are rare, the study of this phenomenon remains complex and relatively little investigated. Yet, evidence is mounting that vagrancy of migratory birds, like other rare events10,11, can have important ecological and evolutionary consequences and that their study could contribute to our understanding of the neuro- physiological bases and heritability of animal navigation4,12.
Transoceanic movements represent among the most spectacular cases of vagrancy13,14,15 and can even, in rare cases, lead to the colonization of new territories16, and ultimately to the evolution of new species17. Several factors can contribute to vagrancy and while both external and internal drivers can explain why individuals move beyond the typical distribution of their species (see5), weather conditions and especially strong winds associated with rapid movements of warm and moist air masses seem to be the key to such successful transoceanic crossings18. Birds choose optimal wind conditions to initiate their migratory flights19,20,21 but how migrants react when encountering unfavorable wind conditions (e.g., opposite or crosswinds) remains a complex question22,23,24. When travelling along coasts, migrating birds can be drifted offshore by strong crosswinds. In such circumstances, they can (1) try to head back towards land via an energy-intensive struggle against the wind; (2) attempt to compensate for the wind drift and maintain the desired heading; (3) continue to fly towards the desired heading without compensating for drift; or (4) fly in the direction the wind is blowing (i.e., downwind)7. The time spent flying above water, the strength of the wind, energy reserves, and the fact that certain species are more inclined to accept wind drift will all have a combined influence on the probability that an individual will follow the wind direction if confronted with adverse winds22,25. But beyond changing their heading, the strength and direction of the winds can also influence the success of long-distance crossings. Strong winds can push individuals into higher flight speeds, enabling them to travel distances that would likely be impossible under normal circumstances26.
In North America, hundreds of millions of birds of numerous migratory species move southwards in autumn after the breeding period along the eastern Atlantic seaboard27. Among these, some species reach non-breeding grounds in the Caribbean, Central or South America by completing a long uninterrupted flight over the Atlantic Ocean (up to 2500 km for the Blackpoll Warbler Setophaga striata28, see29), while others follow the coast in a series of southward movements30,31,32. Such individuals hence face prevailing westerly winds that commonly flow at mid-latitude regions during the autumn season33. Every autumn, several individuals of different species of North American landbirds are observed on islands in the Atlantic Ocean and in continental Europe, thousands of kilometers away from their usual breeding range, and their occurrence there has been attributed to favorable wind conditions (e.g.,34,35,36,37,38). For these transoceanic movements to occur, several climatic elements must occur simultaneously including the departure of migrants heading south and/or east (associated with changes in atmospheric pressure, tailwinds, and clear skies; see31) and the occurrence of a storm including hurricanes, which commonly move north along the coastline from the Caribbean36. We might expect that a massive departure of migrants would lead to a greater likelihood of entrainment across the ocean, but even a smaller number of individuals encountering the right set of conditions could still result in these transoceanic movements. In both circumstances, birds that have been travelling for several hours above the water, near or along the coast may be transported northeast across the Atlantic Ocean by strong westerly winds (see18 for a comprehensive analysis of the unprecedented arrival of American vagrants to the British Isles in September 2023 and see also7 about ship-assistance).
Transoceanic wind-driven vagrancy is therefore expected to be more common in populations which routinely perform over-water flights or closely follow coastlines during migration and hence intersect with prevailing offshore winds38,39. In this study, we analyzed hydrogen stable isotopes of feathers (δ2Hf) sampled from 72 individuals of 26 North American landbird species from live captures and museum specimens between 1936 and 2023 in the Azores (Portugal), Iceland and France (see details in Fig. 1) and estimated the probabilistic origins of feather growth40. All the species sampled have broad breeding ranges in North America including populations on the East Coast, therefore we expected to find that most, if not all, individuals would originate from northeastern populations. Eastern populations breeding in areas north of 45 degrees of latitude are indeed closer to the transoceanic sites where the birds were sampled, and the species mostly take long flights over water during part of their autumn migration. This means they are more likely to encounter adverse wind conditions, without any possibility to land on the continent, thus being displaced across the Atlantic with a shorter distance to cover to reach the Azores, Iceland or Western Europe. Finally, because it has been suggested that misoriented individuals (i.e., those heading in a certain direction but with inaccurate orientation or directional perception, see4) are more likely to come from areas of higher species density41, we have also investigated for each species whether those individuals sampled in Europe originated from Nearctic breeding areas of relatively higher abundance.

Origin of the feather samples analyzed in this study. 72 samples from 26 North American landbird species were collected between 1936 and 2023 in the Azores (Portugal), Iceland and France. Pictures: Yellow-billed Cuckoo and Red-eyed Vireo by Vincent Legrand; Yellow-rumped Warbler and Northern Parula by David Monticelli.
Material and methods
Samples
Feather samples were taken from birds that were captured in the wild and from museum specimens (some of the birds were captured alive to integrate public museum collections or found dead and then deposited in museum collections). The proportion of samples collected from birds captured compared to samples collected from museum specimens varied according to locality and species. In the Azores (Portugal), of the 17 samples used, four came from museum specimens while the others were collected during standard ringing activities. In Iceland, the 48 samples came from specimens of the Icelandic Institute of Nature Research (formerly known as the Icelandic Institute of Natural History) collection. In France, of the seven samples used, one came from ringing activity and the other six from museum specimens. During ringing activities, birds were captured with mist nets and song playback (see34 for details). They were marked with a metal ring (mandatory), measured, sampled and released after ca. 5 min of handling (in accordance with ARRIVE guidelines, https://arriveguidelines.org). Body coverts were collected from ringed individuals, and undertail and/or uppertail coverts were collected from museum specimens. After sampling feathers were stored as dry material in envelopes. All sampled individuals were juvenile (first calendar year) individuals, meaning that the feathers collected were grown at natal sites. We do not expect these individuals to molt any feathers during migration, except in the case of accidental molt due to predation, which could be noticeable if it occurs on flight feathers. Birds were captured and handled with permission from the respective national ringing authorities of France (MNHN-CRBPO) and Portugal (CEMPA- ICNF, references 247/2018 & 260/2019). Mist-netting operations and feather collection in the Azores (Portugal) were conducted under permits delivered by the Regional Secretariat of Environment and Climate Change (permits no. 102/2018/DRA, 84/2019/DRA, 109/2020/DRA, 94/2022/DRAAC) and internationally recognized compliance certificates issued by the Regional Direction of Science and Technology (IRCC no. 56/2018/DRCT, 47/2019/DRCT, 10/2020/DRCT, 56/2021/DRCT, 60/2022/DRCT).
In total, we obtained feather samples of 72 individuals of 26 species including one species of Caprimulgiformes (Common Nighthawk Chordeiles minor), two species of Cuculiformes (Yellow-billed Cuckoo Coccyzus americanus and Black-billed Cuckoo Coccyzus erythropthalmus) and 23 species of Passeriformes (songbirds; see Fig. 1). On average, we sampled 2.5 individuals per species, with many having only a single individual. The species with the most samples were Yellow-billed Cuckoo (n = 10), Red-eyed Vireo (Vireo olivaceus; n = 6), Blackpoll Warbler (n = 6), Gray-cheeked Thrush (Catharus minimus; n = 5), Northern Parula (Setophaga americana; n = 5) and Yellow-rumped Warbler (Setophaga coronata; n = 5; see Fig. 1).
Stable isotope analyses
Feathers were cleaned in a 2:1 chloroform:methanol solvent, dried in a fume hood, and subsamples were weighed out for δ2H analysis at the Environment & Climate Change Canada Stable Isotope Hydrology and Ecology Laboratory, Saskatoon, Saskatchewan, Canada. The δ2H of the non-exchangeable hydrogen of feathers was determined using the method described by42 based on two calibrated keratin δ2H reference materials (CBS: − 197 ‰, KHS: − 54.1 ‰). We performed hydrogen isotopic measurements on H2 gas derived from high-temperature (1350 °C) flash pyrolysis on glassy carbon (Eurovector 3000; Milan, Italy) of 350 ± 10 ug feather subsamples and keratin standards loaded into silver capsules. Resultant separated H2 was analyzed on an interfaced Thermo Delta V Plus continuous-flow isotope-ratio mass spectrometer (Bremen, Germany). Measurement of the two keratin laboratory reference materials corrected for linear instrumental drift were both accurate and precise with typical within-run measurement error < 2 ‰. All results are reported for non-exchangeable H expressed in the typical delta notation, in units of per mil (‰), and normalized on the Vienna Standard Mean Ocean Water–Standard Light Antarctic Precipitation (VSMOW-SLAP) standard scale. Note that three samples (see details in Supplementary Information) were analyzed at the « Ecologie Isotopique » platform of the LEHNA (Université Claude Bernard Lyon 1, France), using high temperature (1450 °C) pyrolysis on glassy carbon (VarioPyrocube, Elementar GmbH, Langenselbold, Germany) coupled in continuous flow to an Isoprime (Elementar UK Ltd, Cheadle Hulme, UK) isotope-ratio mass spectrometer.
Probabilistic assignments to origin
We performed Bayesian geographic assignments to estimate the origin of American landbirds caught in the Western Palearctic based on feather δ2H (δ2Hf) values using the assignR package43. To calibrate our precipitation-feather isoscape, we used growing season precipitation isoscape rasters downloaded from waterisotopes.org at 5 arc-minute resolution44,45 and the isotopic composition of feathers collected on breeding birds from known locations and analyzed in previous studies46,47,48. The package assignR includes a database from known origin samples that we first filtered for landbird species breeding in North America before separating them into two categories: one for long-distance migratory canopy- and one for long-distance migratory ground-foraging species (according to46 and Birds of the World; lists can be found in Supplementary Information). This categorization is based on the methods of previous studies, which expected that trophic position could influence δ2H values46,47. Specifically, canopy-foraging species are likely to have higher δ2H values due to the direct incorporation of enriched leaf water, while ground-foraging species would have δ2H values closer to the local precipitation or groundwater signature. After filtering, the reference samples used in this study came from three publications on the use of isotope measurements for tracking avian movements in North America46,47,48. We produced two calibrated, group-specific isoscapes to perform the assignments analyses following the assignR package guidelines (the calibration equations used for the assignments can be found in Supplementary Information). Then, for each species, we used the breeding distribution from49 to filter the geographic assignments with the raster package50. For each species, we thus estimated the likely origin of each individual by extracting the geographic cell coordinates with different levels (10%, 25%, 50%) of posterior probability.
Statistical analyses
Our first aim was to investigate whether transatlantic vagrants originate from a specific area within the respective species’ breeding distribution, expecting to find that most individuals derive from their northeastern populations. To achieve this, we first compared, for each individual, the latitudinal and longitudinal distribution of posterior probability values of Bayesian geographic assignments against a null model, which included a random selection of geographic grid cells from within the species’ range. We tested different posterior probability thresholds to assess how much of the study area is represented in the assignment region and to evaluate the uncertainty of our assignments. We used and compared three different threshold values (10%, 25%, 50%), meaning that the assignment maps displayed the top 10%, 25%, or 50% of grid cells with the highest posterior probability. For the null model, we randomly sampled the same number of grid cells as identified by the probabilistic assignment method. We then compared the median latitude and longitude origin of each individual with the null model.
Since a species’ abundance is not uniform across the breeding range51, and given that a linear relationship between the number of individuals and the number of potential vagrants is logically expected (even if it has never been formally proved), we then repeated the same analysis by correcting the distribution of each species for breeding abundance. To do so, we first downloaded mean eBird abundance maps52 using the seasonal raster estimates in the ebirdst package53 at a 27 × 27 km resolution for the breeding distribution, rescaling abundance values between 0 and 1. Then, we created a null model, which included a random selection of geographic grid cells from the species’ range with a probability of sampling based on abundance values (extracting the same number of cells). We compared the same threshold values of posterior probability origin (10%, 25%, 50%) and compared the median latitude and longitude origin of each individual with this null, abundance-weighted, model. Because our sampling included both recent and historic samples, we tested if the estimated origins varied as a function of the age of the sample. To do so, we used the results with the 10% highest posterior probability origin and fitted four different linear mixed-effects models (for latitude, longitude, distance between the probable latitudinal and longitudinal origin and the current area of highest abundance) with species as a random effect using the lme4 package54.
Our second aim was to test whether transatlantic vagrants sampled in Europe come from breeding areas of relatively higher abundance. Using the same eBird abundance maps rescaled between 0 and 1, we extracted abundance values in cells that were ranked in the same threshold values of probability of origin (10%, 25%, 50%). We then compared these values with abundance estimates of randomly selected values within the breeding range of the species (extracting the same number of cells). We compared the difference between the median of the distribution of abundance values from the top probability cells based on the assignment using isotope values and the median of the distribution of randomly selected values based on the species’ breeding range. We summarized the information for each species by averaging the differences (of the medians) of all individuals to estimate the proportion of species which individuals come from areas of greater or lower abundances. As abundance data were not available for years corresponding to our earliest samples, we used seasonal raster estimates for the year 2022. We acknowledge that these abundance values may have changed considerably, given that some of the individuals analyzed were caught more than 50 years ago. However, we believe that this bias may be limited, as we mainly analyzed several individuals of the same species caught several years apart, we considered average species patterns, and we assumed that relative abundance remained generally static over broad geographic areas of the breeding ranges over that time period.
Results
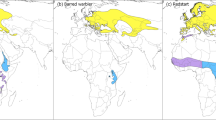
The geographic origin of the Nearctic landbirds captured and sampled at three European sites were more variable than expected (Fig. 2). By comparing with the median distribution of the species, corrected or not for abundance, we indeed found individuals coming from populations located in the northeast, in the south or in the northwest of the species’ distribution (Figs. 2, 3).

Inter- and intraspecific variability in the probabilistic assignments to origin of some transatlantic vagrants. Each map represents the breeding distribution of one species (depicted in light yellow) and the likely origin of all individuals (colored dots) of the species. Each dot represents the median latitude and longitude of the cells appearing in the top 10% of grid cells with the highest posterior probability of origin, the bars indicate the standard-deviation (see Figure S2 for other values). The probability map of origin of all individuals can be found in Figure S1. Note that × 03 (Hermit Thrush Catharus guttatus) was not displayed as cells with the 10% highest posterior probability were outside of the range of the species. Pictures: American Redstart by Adrian Jordi; Yellow-billed Cuckoo and Red-eyed Vireo by Vincent Legrand; Gray-cheeked Thrush by Matthieu Bally; Blackpoll Warbler and Hermit Thrush by David Monticelli.

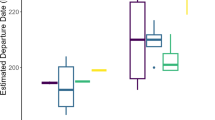
Latitudinal and longitudinal origin of each individual based on probabilistic assignments. The origin of each individual is compared to two null models corresponding to a random selection of grid cells within the breeding range of the species (using the same number of cells), either corrected (plain triangle) or not corrected (empty triangle) for species abundance values. The intensity of color of the points represents the deviation between the origin of an individual and what would be expected by random chance for each species (random but abundance-weighted). The top 10% of grid cells with the highest posterior probability of origin was used in this figure (see Figure S3 for other values).
We found that 17 individuals (24%) of 9 different species likely came from the eastern part of the species distribution, considering that their origin were estimated as at least 10 degrees east of the median distribution of the species (abundance-weighted; Fig. 3). This was particularly true for Northern Parula, Hermit Thrush Catharus guttatus and Grey-cheeked Thrush (Fig. 2 and 3) and was also the case for several species for which we had a smaller number of samples (e.g., Fox Sparrow Passerella iliaca, Black-and-white Warbler Mniotilta varia, Canada Warbler Cardellina canadensis, American Robin Turdus migratorius, Black-billed Cuckoo in Fig. 3 and S1). Conversely, following the same arbitrary criterion, we found that 14 individuals (19%) of 9 different species likely came from the western part of the species distribution (Fig. 3). Hence, we found that one Blackpoll Warbler, two American Redstarts Setophaga ruticilla, two Gray-cheeked Thrushes, one Hermit Thrush had a potential origin in the west (Figs. 2 and 3). All remaining individuals likely originated from areas close to the median of their respective distributions, which does not rule out the possibility that birds could still come from eastern populations, especially if these populations have high abundances (see Blackpoll Warbler and Red-eyed Vireo in Fig. 2 and 3). These results remained the same when using different threshold values of top cells’ probability to estimate likely origins (Figure S2 and S3). Regarding latitude, we found similarly balanced results, with individuals originating from both the northern and southern parts of the species’ distribution (Fig. 3). Importantly, the slight southward tendency in origin may be related to the species’ distribution, which extends much farther north in the western part of the North American continent (e.g., see the distribution of the Blackpoll Warbler in Fig. 2). Nonetheless, we found that 9 out of the 10 Yellow-billed Cuckoos and both Baltimore Orioles Icterus galbula likely originated from the southern part of their breeding distributions and that a Common Nighthawk likely originated from the northern part of the species’ distribution (Fig. 3 and S1). Importantly, since the distributions of isotopic values can be similar over large geographical areas, we found that the probable origin of certain species and individuals could extend over large geographical areas (e.g., see Gray-cheeked Thrush × 09 or × 10 in Fig. 2; Yellow-rumped Warbler, Common Yellowthroat Geothlypis trichas or American Pipit Anthus rubescens in Figure S1). Note that we did not find any significant effect of the age of samples on probabilistic assignments to origin (see Table S1).
By comparing the origin of the samples to the median distribution of species (in latitude and longitude), corrected or not for abundance, we found that vagrants, on average, come from areas closer to the abundance-corrected distribution than to the non-corrected zones (Fig. 3). For latitude, we found that 64 individuals (64%) were closer to the abundance-corrected distribution (in average 4.7 ± 3.4 latitudinal degrees against 5.3 ± 3.8, here for the 10% top values) and 40 individuals (56%) for longitude (12.3 ± 13.2 longitudinal degrees against 14.4 ± 15.7, here for the 10% top values). At the species-level, our results showed that out of the 26 species considered in our study, 18 species (67%) likely originated in average from areas of high relative abundances (Fig. 4). This pattern remained when using different threshold values of top cells’ probability (67% with 25%; 85% with 50%; Figure S4). Apart from a few species with highly variable results between individuals (e.g. Northern Parula, Blackpoll Warbler), we found that almost all individuals of a species generally came from areas of high abundance (e.g. Red-eyed Vireo) or from areas of low abundance (e.g. Yellow-rumped Warbler in Fig. 4).

Link between the likely origin of transatlantic vagrants and areas of relative abundance. For each individual, abundance values were extracted from the eBird abundance maps in cells appearing in the top 10% of grid cells with the highest posterior probability of origin (see Figure S4 for other values) and compared with abundance values extracted from randomly selected values within the breeding range of the species (using the same number of cells). The difference between the median of these two distributions was compared and averaged for each species. Each grey dot represents an individual and colored dots (blue / red) represent the average for the species. Blue colored dots hence represent species originating from areas of lower abundances while red dots represent species originating from areas of higher abundances.
Discussion
Rather unexpectedly, our results showed that migratory individuals from broad geographic breeding origins across North America can be displaced during autumn migration to end up as far as Western Europe. First, we found that a large proportion of vagrant individuals sampled in the Western Palearctic originated from populations located in northeastern regions of North America. This is consistent with our expectations as these populations mostly migrate along the coast or take long migratory flights over the ocean to reach their non-breeding grounds particularly Neotropical migrants (e.g., in Blackpoll Warbler28; Red-eyed Vireo55; Gray-cheeked Thrush56; Canada Warbler32), which makes them more vulnerable to displacement by strong winds during their flight bouts18,38.
We also found that 9 out of 10 Yellow-billed Cuckoos, and two Baltimore Orioles, likely came from the southern part of their breeding range corresponding to the southeastern regions of the US. The literature mentions large flocks of Yellow-billed Cuckoos observed in the northeast US following hurricanes (between mid-September and mid-October) considered to be “reverse migrants from the south” rather than locally breeding birds57. Such birds are, most likely, not actively flying in the reverse direction (i.e., not misoriented individuals4) but are being pushed by predominantly southerly winds, as suggested by the studies of Nisbet58, Richardson59, McLaren et al.38,60 and well-illustrated with the case of the massive displacement of Chimney Swifts Chaetura pelagica as far as Atlantic Canada (and western Europe) by the Hurricane Wilma in 200561. Such birds departed offshore following cold fronts in the southeast of North America and were rapidly transported north-eastwards and pushed ashore in Nova Scotia by strong easterly winds ahead of a low-pressure system that was deepening at the north-eastern end of the front. In this context, it is conceivable to imagine that birds such as Yellow-billed Cuckoos, which are already farther south, could be moved northwards and then across the Atlantic following the track of tropical storms58. Alternatively, such movements towards the northeast could also represent an anticipated retreat from an approaching weather system. To our knowledge, no such movement has been firmly documented (but see62,63), though we can hope that the increasing use of GPS will make it possible.
Finally, we also found that several individuals likely originated from populations in the northwest of the species’ breeding range (and mostly north for the Common Nighthawk). For Blackpoll Warbler, our results are consistent with geolocator tracking and an isotope study showing that birds breeding as far west as Alaska migrate eastwards before making a long flight across the Atlantic to their non-breeding grounds in South America64,65. Therefore, individuals occurring in Europe might have been swept offshore or diverted from their regular migration route when flying southwards over the Atlantic. For individuals of Hermit Thrush (n = 1) and Common Nighthawk (n = 1), geolocator tracking studies (66,67) suggest that at least part of northwestern populations of these species adopt an initial southeast migratory orientation before heading south to reach their non-breeding grounds in Latin America. Even if there is no tracking data available for American Redstart (n = 2) or Gray-cheeked Thrush (n = 2), tracking studies of similar species from northwestern populations also suggest a similar southeast orientation at the start of their migration (see for Veery Catharus fuscescens68, Swainson’s Thrush Catharus ustulatus69, and Yellow-rumped Warbler70). It is plausible that individuals of these species, especially juveniles, which often exhibit a more scattered orientation at the start of their migration71,72, could reach areas of changeable conditions, i.e., the East Coast of North America, and become equally vulnerable to storms during their autumn migration.
Interestingly, previous records of landbird taxa of western origin already exist in Western Europe. They are limited to a few records, e.g. a Western Kingbird Tyrannus verticalis (in Flores, Azores, 201873), Gambel’s White-crowned Sparrows Zonotrichia leucophrys gambelii (in Corvo, Azores, 201374, in Cheshire, UK, 201675) and possible Common Nighthawks from the northern great Plains or unidentified interior subspecies (sennetti, in Corvo, Azores, 200876 and in the UK77), which are all not known to migrate in the eastern half of the American continent. These records raise the possibility that transatlantic vagrants could also include misoriented individuals that were following an erroneous orientation at the start of their migration (i.e. intentionally flying east, towards the Atlantic ocean37,38,78). In such cases, the wind could be a secondary factor as it could facilitate the crossing of individuals that were already heading in a wrong direction. The idea that vagrants can follow an erroneous orientation has been discussed in the literature, both in Nearctic79,80 and Palearctic contexts81. But while much remains to be discovered about the proximal mechanisms of such misorientations5,82,83, these previous records and the several individuals with northwestern origins that we found in this study, suggest that this mechanism could also be a driver of some instances of transatlantic vagrancy. Further research should be conducted such as carrying out orientation cage tests84,85 on Nearctic landbirds captured in Europe or tagging birds (as in81) to track their migratory orientation and ultimately examine their misorientation.
Our findings also suggest that transatlantic vagrants mostly came from areas of higher abundances and align with previous studies showing that the number of vagrants can be linked to the size of their population41. In the case of transatlantic vagrancy, we can consider that many offspring being produced provides a greater probability of having individuals displaced by tropical storms. However, we can also expect that populations that are migrating when tropical storms are passing are the most likely to be displaced, so that a geographical and temporal bias may exist. It is perhaps because of this bias that some species tend to come from areas of lower abundance (e.g. Yellow-rumped Warbler) but this would require further research, certainly on finer geographical scales, and using tracking techniques more precise than isotope tracing.
Our study, using stable isotope techniques, also comes with some limitations. Firstly, there is an inherent spatial ambiguity in isotopic landscapes, meaning that it is generally not possible to obtain precise origins, and distant geographical areas can also exhibit similar isotope signatures. Secondly, even if we did not find any significant influence of the age of our samples, possible changes in precipitation isoscapes since the 1930s86 and the use of calibration equations developed in some cases for adults on juveniles can have an impact on probabilistic assignments. Nevertheless, we believe that using multiple individuals per species and different values of assignment probabilities made our conclusions not overly biased by these limitations. Finally, our sampling was not appropriate for studying proximal mechanisms, particularly the communities of species prone to displacement and the conditions that favor such transoceanic movements. The sampling was scattered over time, and several birds were captured long after their initial discovery, which could have occurred long after their oceanic crossing, making it difficult to trace the wind conditions likely influencing their long-distance vagrancy. It was also not representative of landbird species most regularly observed in autumn in Western Europe. For example, the Red-eyed Vireo, which dominates the list of species regularly occurring on the eastern side of the Atlantic87, can be challenging to capture as most individuals remain in tree canopies.
Finally, we cannot rule out the possibility that some individuals in this study completed part of their journey on a ship (ship-assisted vagrancy). Since all the species considered here undertake long-distance migrations, we considered them as naturally capable of transoceanic movements. However, while no quantified data exist on the frequency or avoidance of overwater flights, some species (e.g., American Robin, Fox Sparrow) may cross water less regularly than others, making them more likely candidates for ship assistance. That said, birds must still reach a ship on their own, and ships capable of carrying them are widespread across the Atlantic. Therefore, we believe our results should not biased by this mechanism.
To summarize, our study suggests that transatlantic vagrants can come from diverse origins with most individuals originating from the northeast of the species’ distribution while several individuals originated from southern or northwestern populations. Little is known about the number of birds that perish without making such transatlantic crossing, but our result show that individual survival is possible even from birds of various geographic origins. Our study calls for a detailed understanding of the context in which these crossings became possible which could improve our understanding about the biogeographic processes that enabled the colonization of isolated archipelagos17. Indeed, in the insular context, there is a tendency to consider that it is the nearby populations that are most likely to colonize a new territory. This is probably often the case, but not exclusively.
Data availability
All data generated and analyzed during this study are included in this published article in a supplementary data file.
References
Egevang, C. et al. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proc. Natl. Acad. Sci. 107, 2078–2081 (2010).
Gill, R. E. et al. Extreme endurance flights by landbirds crossing the Pacific Ocean: Ecological corridor rather than barrier?. Proc. R. Soc. B Biol. Sci. 276, 447–457 (2009).
Aristotle & Balme, D. M. History of Animals. Books VII-X / Aristotle ; Edited and Translated by D.M. Balme. (Harvard University Press, Cambridge, Massachussets, 1991).
Dufour, P., Lees, A. C., Gilroy, J. & Crochet, P.-A. The overlooked importance of vagrancy in ecology and evolution. Trends Ecol. Evol. 39, 19–22 (2024).
Tonelli, B. A., Youngflesh, C. & Tingley, M. W. Geomagnetic disturbance associated with increased vagrancy in migratory landbirds. Sci. Rep. 13, 414 (2023).
Booth, J. E., Gaston, K. J., Evans, K. L. & Armsworth, P. R. The value of species rarity in biodiversity recreation: A birdwatching example. Biol. Conserv. 144, 2728–2732 (2011).
Lees, A. C. & Gilroy, J. J. Vagrancy in Birds (Christopher Helm, 2021).
Jiguet, F. & Barbet-Massin, M. Climate change and rates of vagrancy of Siberian bird species to Europe. Ibis 155, 194–198 (2013).
Pease, B. S., Gilbert, N. A., Casola, W. R. & Akamani, K. The Steller’s Sea-Eagle in North America: An economic assessment of birdwatchers travelling to see a vagrant raptor. People Nature 5, 1937–1947 (2023).
Anderson, S. C., Branch, T. A., Cooper, A. B. & Dulvy, N. K. Black-swan events in animal populations. Proc. Natl. Acad. Sci. 114, 3252–3257 (2017).
Grant, P. R. et al. Evolution caused by extreme events. Philos. Trans. R. Soc. B Biol. Sci. 372, 20160146 (2017).
Grinnell, J. The role of the ‘Accidental’. Auk 39, 373–380 (1922).
Arkhipov, V. Y. & Ławicki, Ł. Nearctic passerines in Russia. Dutch Bird 38, 201–216 (2016).
Howell, S. N. G., Lewington, I. & Russell, W. Rare Birds of North America. (2014).
Suchan, T. et al. A trans-oceanic flight of over 4,200 km by painted lady butterflies. Nat. Commun. 15, 5205 (2024).
Dufour, P. et al. Multilocus barcoding confirms the occurrence of Elegant Terns in Western Europe. J. Ornithol. 158, 351–361 (2017).
Dufour, P. et al. The importance of migratory drop-off for island colonization in birds. Proc. R. Soc. B Biol. Sci. 291, (2024).
Lees, A. C., Farnsworth, A., Curley, S. R. & Gilroy, J. J. Unprecedented arrival of Nearctic landbirds in Britain and Ireland. Br. Birds 117, 292–307 (2024).
Liechti, F. Birds: blowin’ by the wind?. J. Ornithol. 147, 202–211 (2006).
Richardson, W. J. Timing of bird migration in relation to weather: Updated review. In Bird Migration 78–101 (Springer Berlin Heidelberg, Berlin, Heidelberg, 1990). https://doi.org/10.1007/978-3-642-74542-3_6.
Åkesson, S. & Hedenström, A. Wind selectivity of migratory flight departures in birds. Behav. Ecol. Sociobiol. 47, 140–144 (2000).
Horton, K. G. et al. Nocturnally migrating songbirds drift when they can and compensate when they must. Sci. Rep. 6, 21249 (2016).
McLaren, J. D., Shamoun-Baranes, J., Dokter, A. M., Klaassen, R. H. G. & Bouten, W. Optimal orientation in flows: Providing a benchmark for animal movement strategies. J. R Soc. Interface 11, 20140588 (2014).
Gill, R. E. et al. Hemispheric-scale wind selection facilitates bar-tailed godwit circum-migration of the Pacific. Anim. Behav. 90, 117–130 (2014).
Horton, K. G. et al. Navigating north: how body mass and winds shape avian flight behaviours across a North American migratory flyway. Ecol. Lett. 21, 1055–1064 (2018).
Hameed, S., Norwood, H. H., Flanagan, M., Feldstein, S. & Yang, C. The Influence of El Niño on the Spring Fallout of Asian Bird Species at Attu Island. Earth Interact. 13, 1–22 (2009).
Dokter, A. M. et al. Seasonal abundance and survival of North America’s migratory avifauna determined by weather radar. Nat. Ecol. Evol. 2, 1603–1609 (2018).
DeLuca, W. V. et al. Transoceanic migration by a 12 g songbird. Biol. Lett. 11, 20141045 (2015).
McKinnon, E. A., Artuso, C. & Love, O. P. The mystery of the missing warbler. Ecology 98, 1970–1972 (2017).
La Sorte, F. A., Fink, D., Hochachka, W. M. & Kelling, S. Convergence of broad-scale migration strategies in terrestrial birds. Proc. R. Soc. B Biol. Sci. 283, 20152588 (2016).
Cooper, N. W. et al. Atmospheric pressure predicts probability of departure for migratory songbirds. Mov. Ecol. 11, (2023).
Roberto-Charron, A. et al. Widely distributed breeding populations of Canada warbler (Cardellina canadensis) converge on migration through Central America. BMC Zool. 5, 10 (2020).
La Sorte, F. A. et al. The role of atmospheric conditions in the seasonal dynamics of North American migration flyways. J. Biogeogr. 41, 1685–1696 (2014).
Monticelli, D. et al. Ringing American passerines in the Azores: preliminary results with special mention of the first record of Connecticut Warbler for the Western Palearctic. Br. Birds 115, 72–87 (2022).
Lees, A. C. & Gilroy, J. J. Vagrancy mechanisms in passerines and near-passerines. Rare Birds Where When 2, (2009).
Elkins, N. Nearctic landbirds in Britain and Ireland: A meterological analysis. Br. Birds 72, 417–433 (1979).
Elkins, N. Nearctic landbirds in Britain and Ireland. Br. Birds 92, 83–95 (1999).
McLaren, I. A., Lees, A. C., Field, C. & Collins, K. J. Origins and characteristics of Nearctic landbirds in Britain and Ireland in autumn: a statistical analysis. Ibis 148, 707–726 (2006).
Robbins, C. S. Predictions of future Nearctic landbird vagrants to Europe. Br. Birds 73, 448–457 (1980).
Hobson, K. A. & Wasseaar, L. I. Tracking Animal Migration with Stable Isotopes (Academic Press, 2018).
Zawadzki, L. C., Veit, R. R. & Manne, L. L. The influence of population growth and wind on vagrancy in a North American passerine. Ardea 107, 131 (2019).
Wassenaar, L. I. & Hobson, K. A. Comparative equilibration and online technique for determination of non-exchangeable hydrogen of keratins for use in animal migration studies. Isotopes Environ. Health Stud. 39, 211–217 (2003).
Ma, C., Vander Zanden, H. B., Wunder, M. B. & Bowen, G. J. assignR : An R package for isotope-based geographic assignment. Methods Ecol. Evol. 11, 996–1001 (2020).
Bowen, G. J., Wassenaar, L. I. & Hobson, K. A. Global application of stable hydrogen and oxygen isotopes to wildlife forensics. Oecologia 143, 337–348 (2005).
Bowen, G. J. & Revenaugh, J. Interpolating the isotopic composition of modern meteoric precipitation. Water Resour Res. 39, (2003).
Hobson, K. A., Van Wilgenburg, S. L., Wassenaar, L. I. & Larson, K. Linking Hydrogen (δ2H) Isotopes in Feathers and Precipitation: Sources of Variance and Consequences for Assignment to Isoscapes. PLoS ONE 7, e35137 (2012).
Hobson, K. A. & Koehler, G. On the use of stable oxygen isotope (δ18O) measurements for tracking avian movements in North America. Ecol. Evol. 5, 799–806 (2015).
Hobson, K. A. & Wassenaar, L. I. Linking breeding and wintering grounds of neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers. Oecologia 109, 142–148 (1997).
BirdLife International. IUCN Red List for Birds. (2020).
Hijmans, R. J. & van Etten, J. raster: Geographic analysis and modeling with raster data. Preprint at (2012).
Brown, J. H. On the relationship between abundance and distribution of species. Am. Nat. 124, 255–279 (1984).
Fink, D. et al. eBird Status and Trends, Data Version: 2022; Released: 2023. . Preprint at (2023).
Strimas-Mackey, M., Ligocki, S., Auer, T. & Fink, D. ebirdst: Access and Analyze eBird Status and Trends Data Products. R package. Preprint at (2023).
Bates D, Maechler M, Bolker B, W. S. lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1–7. (2014).
Callo, P. A., Morton, E. S. & Stutchbury, B. J. M. Prolonged spring migration in the Red-eyed Vireo (Vireo olivaceus). Auk 130, 240–246 (2013).
Whitaker, D. M., Warkentin, I. G., Hobson, K. A., Thomas, P. & Boardman, R. Fall and Winter Movements of Newfoundland Graycheeked Thrushes (Catharus minimus minimus). Anim. Migr. 5, 42–48 (2018).
Veit, R. R. & Petersen, W. R. The Birds of Massachusetts. (Massachussetts Audubon Society, 1993).
Nisbet, I. C. T. American passerines in western Europe, 1951–62. Br. Birds 55, 139–158 (1963).
Richardson, W. J. Northeastward Reverse Migration of Birds over Nova Scotia, Canada, in Autumn: A Radar Study. Behav. Ecol. Sociobiol. 10, 193–206 (1982).
McLaren, I. A. A notable autumn arrival of reverse migrants in southern Nova Scotia. North American Birds 54, 4–10 (2000).
Dionne, M., Maurice, C., Gauthier, J. & Shaffer, F. Impact of Hurricane Wilma on migrating birds: The case of the Chimney Swift. Wilson J. Ornithol. 120, 784–792 (2008).
Lisovski, S. et al. Inherent limits of light-level geolocation may lead to over-interpretation. Curr. Biol. 28, R99–R100 (2018).
Streby, H. M. et al. Tornadic storm avoidance behavior in breeding songbirds. Curr. Biol. 25, 98–102 (2015).
DeLuca, W. V. et al. A boreal songbird’s 20,000 km migration across North America and the Atlantic Ocean. Ecology 100, (2019).
Dunn, E. et al. Feather isotopes ( δ 2 H f ) and morphometrics reveal population-specific migration patterns of the Blackpoll Warbler (Setophaga striata). Avian Conserv. Ecol. 18, art16 (2023).
Ng, J. W. et al. First full annual cycle tracking of a declining aerial insectivorous bird, the Common Nighthawk (Chordeiles minor), identifies migration routes, nonbreeding habitat, and breeding site fidelity. Can. J. Zool. 96, 869–875 (2018).
Alvarado, A. H., Fuller, T. L. & Smith, T. B. Integrative tracking methods elucidate the evolutionary dynamics of a migratory divide. Ecol. Evol. 4, 3456–3469 (2014).
Hobson, K. A. & Kardynal, K. J. Western Veeries use an eastern shortest-distance pathway: New insights to migration routes and phenology using light-level geolocators. Auk 132, 540–550 (2015).
Justen, H. C., Easton, W. E. & Delmore, K. E. Mapping seasonal migration in a songbird hybrid zone—heritability, genetic correlations, and genomic patterns linked to speciation. Proc. Natl. Acad. Sci. 121, (2024).
Szarmach, S. J. et al. Migration strategies of a high-latitude breeding songbird (Setophaga coronata coronata) revealed using multi-sensor geolocators and stable isotopes. Preprint at https://doi.org/10.1101/2024.10.09.617415 (2024).
Ralph, C. J. Disorientation and possible fate of young passerine coastal migrants. Bird-Banding 49, 237 (1978).
Åkesson, S., Bakam, H., Martinez Hernandez, E., Ilieva, M. & Bianco, G. Migratory orientation in inexperienced and experienced avian migrants. Ethol. Ecol. Evol. 33, 206–229 (2021).
Tipper, R. et al. Aves de ocorrência rara ou acidental em Portugal. Relatório do Comité Português de Raridades referentes aos anos de 2018 e 2019. Anuário Ornitológico 13, 3–101 (2021).
Tipper, R. et al. Aves de ocorrência rara ou acidental em Portugal. Relatório do Comité Português de Raridades referentes aos anos de 2013 e 2014. Anuário Ornitológico 11, 3–55 (2019).
BOURC. British Ornithologists’ Union Records Committee: 50th Report (October 2019). Ibis 162, 263–268 (2020).
Jara, J. et al. Aves de ocorrência rara ou acidental em Portugal. Relatório do Comité Português de Raridades referente aos anos de 2008 e 2009. Anuário Ornitológico 3–71 (2010).
Cleere, N. A review of common Nighthawks in Britain. Br. Birds 117, 10–25 (2024).
Alerstam, T. Bird Migration. (Cambridge., 1990).
DeSante, D. F. An analysis of the fall occurrences and nocturnal orientations of vagrant wood warblers (Parulidae) in California. (Stanford University, 1973).
Diamond, J. M. Mirror-image navigational errors in migrating birds. Nature 295, 277–278 (1982).
Thorup, K. et al. Orientation of vagrant birds on the Faroe Islands in the Atlantic Ocean. J. Ornithol. 153, 1261–1265 (2012).
Dufour, P. et al. The Yellow-browed Warbler (Phylloscopus inornatus) as a model to understand vagrancy and its potential for the evolution of new migration routes. Mov. Ecol. 10, 59 (2022).
Bensch, S., Duc, M. & Valkiūnas, G. Brain parasites and misorientation of migratory birds. Trends Parasitol. https://doi.org/10.1016/j.pt.2024.02.008 (2024).
Bianco, G. et al. Emlen funnel experiments revisited: methods update for studying compass orientation in songbirds. Ecol. Evol. 6, 6930–6942 (2016).
Emlen, S. T. & Emlen, J. T. A technique for recording migratory orientation of captive birds. Auk 83, 361–367 (1966).
Vander Zanden, H. B. et al. Contrasting assignment of migratory organisms to geographic origins using long‐term versus year‐specific precipitation isotope maps. Methods Ecol. Evol. 5, 891–900 (2014).
Hobbs, J. List of Nearctic Passerines in the Western Palearctic. version 4.0. https://www.dutchbirding.nl/references/1678/joe_hobbs_list_of_birding_references (2024).
Acknowledgements
It is not usual to include one of the co-authors within the acknowledgements section, but we consider imperative to dedicate this work to Dr. Keith Hobson, who passed away during the publication process of this paper. We are deeply grateful for his pioneering and major contributions in the domain of ornithology and bird migration, and of course the valuable help he provided in the conceptualization and execution of this work. We thank Laurent Simon from UMR LEHNA for his help for the analyses of three samples; Alison Johnston from the University of St Andrews for first guidance into the eBird abundance predictive data; Pierre-André Crochet for fruitful discussions during various stages of the project development; curators of the Paris and Le Havre Museum of Natural History; the Icelandic Institute of Nature Research (formerly known as the Icelandic Institute of Natural History) for granting us access to their collection; persons who found these vagrants and who transported birds found dead to collections; Mathieu Bally, Adrian Jordi, Vincent Legrand for their photos, all taken in Europe, which were used to illustrate some figures; K. Courtney and G. Baudrin for preparing samples for stable isotope analysis and to G. Koehler for conducting the stable isotope analysis.
Author information
Authors and Affiliations
Contributions
PD performed all statistical analyses and wrote the initial version of the manuscript; FJ and DM conceptualized the study, and has been assembling feather samples since a decade; YK organized the feather sampling for Icelandic specimen; DM, PA, DK, TV organized or participated to feather collection in the Azores; KH, KK, and LS performed the stable isotope analyses and KK performed preliminary isotopic assignments; all co-authors revised and commented the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Dufour, P., Kardynal, K.J., Hobson, K.A. et al. Origins of Nearctic migratory landbird vagrants recorded in Europe revealed by feather isotopic analysis. Sci Rep 15, 15456 (2025). https://doi.org/10.1038/s41598-025-99765-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-99765-4
