
Abstract
Infectious diseases can drive populations and species to extinction. Beak and feather disease virus (BFDV) is a circovirus of global conservation concern that can infect all Psittaciformes and some other species. Yet some parrot species, such as Crimson rosellas (Platycercus elegans), can live successfully with high BFDV prevalence (>40%) with no clinical signs reported in infected individuals. We assessed BFDV load in 10-12 tissues per bird, from n = 66 P. elegans, to reveal tissue tropism and BFDV persistence in tissues. Here we show that in 94% of individuals, BFDV was detected in one or more tissues. While BFDV replicated to high levels in subadults, in adults (some confirmed seropositive) the virus persisted in various tissues at much lower levels. Our findings reveal that BFDV is much more common in wild P. elegans than previously thought and suggest that current screening practices (mostly on blood) may substantially underestimate BFDV infection estimates, with implications for biosecurity and conservation programs globally.
Similar content being viewed by others
Introduction
Infectious diseases are considered one of the drivers of local population declines in wildlife1,2. Some diseases have been linked to species extinctions, such as chytrid fungus which has caused a catastrophic loss of amphibian biodiversity worldwide3, or avian malaria and pox virus which have caused the decline and extinction of Hawaiian forest bird species4,5. The fade-out of diseases in a declining population can result in reduced herd immunity, which then may have catastrophic consequences as a result of a spill-over event6. Around a third of the 398 species that form the order Psittaciformes globally are classified as threatened according to the IUCN Red List7, which makes it one of the bird orders of greatest conservation concern. Beak and feather disease virus (BFDV) is an avian viral disease that mostly affects this group6,8,9 and is considered to be the dominant viral pathogen in wild psittacine birds in Australia10. Accordingly, BFDV has the potential to cause population declines and hamper conservation programmes especially through spill-over from abundant reservoir hosts11,12,13. Examples have been reported in Orange-bellied parrots (Neophema chrysogaster) in Australia14,15, Red-crowned parakeets (Cyanoramphus novaezelandiae) in New Zealand12,16, Echo parakeets (Alexandrinus eques) in Mauritius17 and Cape parrots (Poicephalus robustus) in South Africa18. Thus, research to reveal the epidemiology and infection biology of BFDV in the wild is urgently needed.
BFDV is a small ssDNA circular virus from the Family Circoviridae that can cause Psittacine beak and feather disease (PBFD)19,20. All psittacine birds (parrots, cacatuids and lorikeets) are considered susceptible to infection with BFDV, although intensity and disease outcomes may vary among species20,21. Clinical disease often develops in cacatuids and some parrot species, and infected individuals may eventually die due to starvation and/or secondary infections12,14,22,23,24,25,26. Common clinical signs include symmetric feather loss with growing of dystrophic feathers, beak and claw deformity and immunosuppression19,26. However, there are some clinically healthy psittacine bird species in which BFDV has been detected, both in captivity and in the wild25,27,28,29,30,31,32. BFDV infection has also been detected in non-psittacine bird species33,34,35,36, as well as in captive and wild psittacine birds outside Australia9,32,37,38,39. The widespread occurrence of BFDV raises concerns about its emergence and transmission pathways that have contributed to its global spread in spite of the biosecurity measures in place9.
The infection biology and epidemiology of BFDV remains poorly understood, especially in wild free-ranging birds. Crucially it is unknown how some species live with high levels of infection while others develop clinical signs and succumb to disease. To investigate this, we used the Crimson rosella (Platycercus elegans), a common abundant endemic parrot of eastern and southeast Australia, which does not show clinical signs despite having high BFDV prevalence and virus load in blood27,28,29. Previous studies from our group have found that: subadult P. elegans had higher detection rates and virus loads in blood compared to adults27,40,41; they could produce high antibody titre against BFDV42; and at least some adults (using samples of recaptured birds) had been able to clear the virus from blood43. It was hypothesized that P. elegans had evolved mechanisms to live with BFDV infection in the wild, but how they do this is unknown. For example, it was not known if the virus has particular tissue tropism, or whether BFDV persists in tissues of infected birds after clearing from blood (similarly to what occurs with other circoviruses in vertebrates44,45). Understanding these features of infection will provide important insights into the infection biology and epidemiology of BFDV. The aim of this study was to quantify BFDV genome load (hereafter referred to as virus load) in multiple tissues (including blood) from wild birds, and to obtain seroprevalence data from wild P. elegans.
Similar studies quantifying virus load and investigating virus distribution in tissues exist for some similar pathogens and include the porcine circovirus-2 (PCV-2) in pigs46, chicken anaemia virus (CAV) in poultry47, and Aleutian mink disease parvovirus in mink48. However, to the best of the authors’ knowledge our study is the first to investigate this for BFDV in any wild or captive birds. Our findings indicate that BFDV infects a higher proportion of wild P. elegans individuals than previously thought and suggest that current screening approaches would significantly underestimate BFDV prevalence in this host species, and likely others, with implications for Psittaciformes conservation and welfare globally.
Results
BFDV detection and load
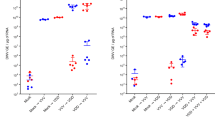
We detected BFDV DNA in all tissue types analysed by qPCR. The detection rate of BFDV was significantly higher in tissues from subadult P. elegans (F = 193.457, p < 0.001; Fig. 1, Supplementary Fig. 1a), while it was not significantly affected by the sex of the bird (F = 1.497, p = 0.120; Supplementary Fig. 1b). While bursa of Fabricius was only tested in subadults, the conclusions remained the same after the exclusion of this tissue in this pooled analysis (F = 199.691, p < 0.001 for age effect; F = 1.124, p = 0.339 for sex effect). Similarly, the mean BFDV load of all positive tissues pooled together was significantly higher in subadult birds compared to adults (F = 5.122, p < 0.001; Supplementary Fig. 1a) and no sex effect was detected in virus load among tissues (F = 0.810, p = 0.640; Fig. S1b) nor within each age group (p > 0.5; Supplementary Fig. 1c). These results did not change either when excluding bursa from the analysis (F = 5.088, p < 0.001 for age effect; F = 0.808, p = 0.632 for sex effect). Nonetheless, when considering just subadults we found a significant interaction in which the post-hoc test revealed that males had higher virus load in spleen than females even after applying sequential Bonferroni correction for multiple comparisons (t = 2.493, p = 0.013); this was not the case for any other tissue types in subadults nor any of the tissue types in adults. Thus, from hereafter we report our results divided by age class.

plot shows the distribution of BFDV virus load for each tissue (each dot represents a BFDV-positive data point) by age class, from all P. elegans (n = 66). Virus load is expressed in log10 form, with the median (black line) and the 25–75 percentiles (blue lines) indicated for each tissue type. The detection limit was set at log10 = 0.5 for illustration purposes only, indicating that we considered negative or “undetected” any value < 3 BFDV copies/106 cells. To highlight the differences between positive tissues, negative values were not plotted. Fractions on top of each plot indicates the total number of positive tissues out of the total analysed. Bursa was only identifiable in subadults.
Comparison of BFDV detection and load between age group
In subadult P. elegans (n = 32), BFDV was detected in 74% (95% CI 57.9, 86.8) of blood samples, in 48% (95% CI 30.7, 66.0) of frozen faeces, and in 76–87% of all other tissues (Fig. 1, results for faeces not shown). Out of the subadults analysed, 66% of individuals had BFDV detectable in all tested tissue types, 28% had BFDV in only some tissues (excluding blood and faeces) and only 6% were negative in all tissues tested (Supplementary Fig. 2). Furthermore, the virus load for each tissue type was significantly higher when compared to adults (p < 0.001), except for blood feather (p = 0.587) (Fig. 1). The highest virus loads were detected in bursa, spleen and skin of subadults (Fig. 1).
In adult P. elegans (n = 34), BFDV was not detected in blood nor faeces and detection in other tissue types varied from 6% in liver to 67% in lung and 68% in spleen (Fig. 1, results for faeces not shown). Despite not detecting the virus in blood, BFDV was still detected at a lower level than subadults (Fig. 1) in between 10 to 50% of tissues in approximately 77% of the adult birds. Only 6% of the adults had no BFDV detectable in any of the tissues analysed (Supplementary Fig. 2). Although still at low levels compared to the subadult loads, the highest virus loads in adults were detected in spleen, lung, skin and blood feathers (Fig. 1). We saw that the few positive blood feathers from adults (n = 5) had significantly higher virus load than five other tissues in adults (crop, heart, kidney, liver and pectoral muscle; p < 0.05), and that the virus load was similar to the blood feathers (n = 20) from subadults (t = 0.544, p = 0.587) (Fig. 1).
Overall, we found that 94% (95% CI 79.9, 98.3) of the subadults tested have at least one tissue with detectable BFDV. It further appears that the infection carries through to adulthood, with 94.1% (95% CI 80.9, 98.4) of adults having detectable BFDV in at least one tissue. BFDV was detected in all the tissues tested from all the subadults with BFDV-positive blood except for two birds (Supplementary Fig. 3). Each of these two subadults (N017 and N029) had one tissue testing negative to BFDV, which were lung and blood feather respectively. There was a positive and significant correlation between virus load in blood and virus load in each different tissue type (Fig. 2). The most common BFDV-positive tissues in blood-negative birds were spleen (62%), lung (59%), and skin (58%) (Fig. 2). In line with previous studies in this species27,28,29, the birds sampled for this study did not have external signs indicative of PBFD. Internally, no inclusion bodies typical of PBFD were detected in the histopathology exam of a subset of infected birds, composed of three subadults and three adults.

Pearson’s correlation coefficient (r) and significance (p-value) are shown in grey boxes for each correlation. Faeces were not included as their virus load values were not obtained.
BFDV blood-negative birds
Of the birds that were negative in blood samples (n = 42), 38 had BFDV detectable in at least one tissue (adults 32/34, subadults 6/8). Had we relied on blood sampling alone, the apparent prevalence of BFDV in these P. elegans would be 35.3% (23 out of 65 individuals positive or a sensitivity of 37%). Tissues that were more frequently positive in blood negative birds were lung (n = 24), spleen (n = 23), skin (n = 22), kidney (n = 18) and heart (n = 17). Conversely, liver (n = 4), bone marrow (n = 4), pectoral muscle (n = 5), ovary (n = 1) and testis (n = 4) were the least frequently positive tissues in blood-negative birds. This suggested that screening blood in adults would be particularly insensitive to detect infection in tissues, while the most sensitive sample types would be the spleen (apparent prevalence of 75% and a sensitivity of 80%) followed by the skin (apparent prevalence of 72% and specificity of 77%) (Fig. 1).
Serology
Only the few positive serology results on haemagglutination inhibition (HI) could be trusted as positive, which are shown in the Supplementary Information (Supplementary Table 1). This was due to our recent findings that extended storage duration of dried blood spots on filter paper at room temperature reduces the ability to detect antibodies to BFDV by HI42 and that most of our samples on filter paper had been stored at room temperature for a long period before being tested. Despite this, and that we probably had several false negatives, we still saw that antibodies were more commonly detected in adults (6/36) compared to subadults (2/32). We found that even with a positive HI titre, birds were still BFDV positive in some tissues and in one case even in blood (Supplementary Table 1). Adult bird (N009) with 1:320 antibody titre had been initially tested after only 6 weeks of storage, which explains the higher titre in comparison to the other birds.
Discussion
Our findings have major implications for the understanding and management of this globally distributed circovirus, and potentially other circoviruses in wild vertebrates. As 94% of individuals were infected in one or more tissue types, our findings clearly show that BFDV infection is widely present in this species. This rate of infection is much higher than expected, and much higher than has been reported for BFDV in any wild parrot species to date25,29,30,39,49,50. We also reveal that BFDV persists and is detectable in multiple tissues of adult P. elegans, and even when the virus is not detectable in blood. This and the low detection rates in blood from adults compared to subadults has major implications for future screening of BFDV status in parrots. It shows that screening adults using blood samples alone is likely to severely underreport the true abundance and perhaps distribution of BFDV in wild populations. This has wide implications, encompassing errors in surveillance of wild birds, assessing risks of BFDV globally, implementing conservation programmes and captive breeding programmes (for example, in the Orange-bellied parrot captive breeding programme where infection with BFDV has threatened the survival of this species51) and for biosecurity screening of pets and the bird trade.
In contrast to previous studies that have only reported BFDV load in the blood of wild parrots24,27,29, we show that virus load varies greatly among tissue types. The higher virus loads we found in spleen, skin, bursa (in subadults) and lung (in adults) strongly suggest that BFDV possesses tropism for these tissues or that the cell types it infects most commonly (e.g. lymphocytes or macrophages19) predominate in such tissue types. Studies using histopathology in combination with immunohistochemistry or in situ hybridization52,53 for BFDV and determination of positive cell types would be needed to test these hypotheses. BFDV was not detected in blood of any of the adults tested for this study, yet most of them still had a few tissues with detectable BFDV, although at much lower load than in subadults.
While our results indicate that BFDV replicated to high levels in apparently healthy subadult P. elegans, the most parsimonious interpretation was that adults had been able to reduce the virus to undetectable levels in blood and at least partially from other tissues. It appeared that these same adults were, however, not able to clear the virus completely, as the detection rate, but not the actual virus level, in tissues was practically the same between subadults and adults. If adults were able to clear the virus completely, or if subadults died from infection and so could not be sampled as adults, then a much lower detection rate of positive tissues would be expected in adults, but this was not the case. Thus our findings seem to indicate that BFDV resembles other circoviruses in that it persists in tissues after infection has cleared from blood44,45. How long this particular circovirus stays in tissues, and whether it may later reactivate under certain circumstances (e.g. stress, breeding) remains to be determined.
Our findings present a challenge for BFDV screening in birds, as obtaining a tissue sample such as spleen or lung are virtually impracticable in live birds, and collecting a skin biopsy is still a much more invasive procedure than would be obtaining blood, a feather, or an oral/cloacal swab. We showed that the sensitivity of testing blood alone was very low (37%), while testing skin had a much better sensitivity (72%). A skin biopsy can be collected from a living bird with minimal risk, although sedation/anaesthesia would be recommended54. To improve sensitivity further, a blood feather could be collected along with a skin biopsy, which would bring the apparent prevalence up to 87% (41 individual birds had both of these samples collected and 36 were positive). The caveat here is that blood feathers might only be present if the individual is currently moulting. Therefore, in situations where it may be important to know if an individual is positive for BFDV, and necropsy is not an option, testing skin has twice the sensitivity of testing blood alone and testing skin with a blood feather brings the sensitivity up to 92%.
We observed a strong age effect on BFDV detection and load in multiple tissues in P. elegans, with subadults having a much higher load than adults. This is in line with previous work on blood from wild P. elegans27,40,41,55. This finding on P. elegans could be a species-specific characteristic of BFDV or, more likely, could be simply due to the difficulty of ageing other species, in combination with a lack of sampling of other species in the wild.
The lack of successful cell-culturing techniques for BFDV to date56 creates a challenge in separating the detection of low virus loads in tissues from active infection that is shed and contagious. While detection of virus DNA does not necessarily indicate active infection or replication, high copies of a virus in a tissue is suggestive of an active replication taking place, and usually associated with a higher probability of transmission than the presence of very low loads. We report high levels of BFDV DNA in several tissues including bursa, skin, spleen, lung, liver, kidney, bone marrow and crop. The highest virus loads were found in subadults, particularly in skin, spleen and bursa of Fabricius, which is an organ involved in the development of the immune system in birds, but that involutes in adults, and is known to be targeted by BFDV57. Our findings are in line with previous studies using in situ hybridization (ISH) which showed high virus load of other circoviruses (pigeon and geese circoviruses) in similar tissues58,59 and other studies showing BFDV inclusions in epithelial cells and also frequently in macrophages in bursa, skin or feathers19,60. Studies using ISH on infected tissues would provide further insights into the pathogenesis of BFDV.
Despite finding high amounts of BFDV DNA in tissues from most subadults, we did not detect PBFD-associated macroscopic signs of disease during the necropsies of subadults and adults. This supports the idea that P. elegans is unusual in being able to cope with BFDV, and is in line with much previous work by our group on P. elegans where, in two decades of field work, we have not found obvious clinical signs of disease caused by BFDV in wild-caught individuals27,28,29. Consistent with this, BFDV infection in clinically normal birds has also been described in closely related Eastern rosellas (Platycercus eximius) that were introduced to New Zealand25 and in some other captive psittacine species in Australia61 and Europe31,32. This suggests that subclinical infection, or chronic carrier state, may also exist in some host species other than P. elegans. While disease has been reported in some of these species, such as P. eximius on occasions and African grey parrots (Psittacus erithacus)12,22, it has not for P. elegans, to our knowledge, and also not to the knowledge of at least 25 veterinarians who we have consulted, who frequently see wild birds in their private practice in Australia (pers. comm. till 2024). In some other circoviruses, such as PCV-2, moderate to high amounts of virus has been associated with disease development while low levels usually lead to subclinical infections62. However, in our study of P. elegans even high amounts of virus were not associated with clinical signs of PBFD.
Detection of BFDV in faeces was much lower than in any other tissue tested, and it was only detected in individuals that also had BFDV in all or most of their tissues. BFDV in faeces is thought to originate in the bile from circovirus in the liver23, or from virus damage and persistence in the gastrointestinal tract and bursa of Fabricius63. Nevertheless, we found ten birds that were positive in the liver but negative in faeces, which, together with the general low detection rate in our study, could be explained by an intermittent excretion of virus in faeces, as reported earlier64, or by a lack of shedding in our birds. Despite some studies still using fresh faeces to screen wild birds for BFDV61, faecal testing alone is not recommended if other samples can be collected due to its low sensitivity65.
Given our recent finding about the rapid decline in antibody titres for BFDV in filter paper that is stored long-term at room temperature42, and that our samples for this study were stored 12–82 weeks at room temperature, it was likely that our HI testing underestimated antibody titres and so potentially had false negative results. We hypothesised that birds get infected while young and they develop antibodies, as they get older. Our small dataset suggested that a higher proportion of adults had seroconverted for BFDV, compared to subadults. This would be in line with our hypothesis, suggesting that the production of antibodies may help the birds to clear the virus from blood43, but not completely from other tissues.
In conclusion, we reveal that wild P. elegans have a very high detection rate (94%) of infection with BFDV in young and adult birds, and with no evidence of clinical disease. Adult birds can clear the virus completely from blood and reduce the load of virus in tissues likely due to being able to generate antibodies against BFDV. This supports the idea that either P. elegans has evolved mechanisms to successfully respond to and live with BFDV27, or that the BFDV variants in P. elegans have adapted to this host species, in comparison to many other Australian parrot species in which individuals get sick and eventually die from the disease. Further studies into the immune and inflammatory responses to infection with BFDV in P. elegans may help understand how the avian immune system could effectively respond to initial infection with BFDV and tolerate ongoing persistence of the virus in tissues. This may have implications for future control efforts in other psittacine species.
Methods
Sample collection
Work was approved by the Deakin University Animal Ethics Committee (permits B32-2018 and B02-2019) and conducted under the Australian Bird and Bat Banding Scheme authority number 2319 in compliance with the Wildlife Act 1975 research permit approved by the Victorian Department of Environment, Land, Water and Planning permits (numbers 10008963 and 10009114).
Samples were collected from wild P. elegans in Victoria, Australia, during 2019 (n = 51) and 2020 (n = 15). The majority of birds were collected between May and June of each year, and most (n = 61) were collected from licensed shooters holding an Authority to Control Wildlife permit to cull P. elegans as orchard pests. The culled birds came from sites in central Victoria within 200 km of our other field sites. The remaining birds were acquired as casualties (n = 6) collected recently after death from field sites in Bellbrae (S38° 19′ E144° 1′) and Meredith (S37° 51′ E144° 06′).
Carcasses were collected early in the morning in winter when temperatures did not exceed the 8–10 °C, and kept at 4 °C shortly after death and necropsied within 35 h. The age of the individuals was recorded as subadult (if mostly green contour plumage, first year post-fledgling) or adult (if red contour plumage, second year or older)40,55. For each individual (n = 66), we collected blood samples from the heart, as well as samples of the 12 different tissues listed in Table 1, up to a total of n = 813 samples. In some individuals, the tissues had been damaged and they could not all be identified during necropsy; therefore, the total number of samples collected from each individual varied (Table 1). Bursa of Fabricius (bursa) was only collected from subadults as it was involuted in adults and therefore not identifiable. Blood and tissue samples were stored in RNAlater® (Sigma Aldrich, USA) at −20 °C. Additional blood samples were stored in 90% ethanol and as dried blood spots on filter paper (Whattman®; Sigma Aldrich, USA), both at room temperature in the laboratory. Faeces collected directly from the cloaca and blood feathers were collected in Eppendorf tubes and stored frozen at −80 °C. During necropsy individuals were examined for gross lesions that could be consistent with PBFD (e.g. loss of or abnormally growing feathers, change in feather coloration, or abnormalities in beak and claws6). Additional tissue samples were collected in formalin and submitted for histopathology staining with haematoxylin and eosin (H/E) at the Australian Centre for Disease Preparedness (ACDP) in Geelong, Australia.
To minimize cross contamination between individuals and samples, all sampling materials were single-use when possible (e.g. scalpel, plates to cut tissues, gloves). These were swapped between bird necropsies, and new clean scalpels were used to cut internal organs for collection. F10 SC Veterinary Disinfectant (Health and Hygiene Pty Ltd, South Africa) followed by 2% bleach solution bath were used to sanitize all reusable utensils (e.g. forceps, scissors, cutting board) between necropsies43,66.
Creation of a standard for qPCR quantitation
We used a standard curve with known BFDV-positive quantities and molecular grade water as positive and negative controls, respectively, which allowed us to quantify the number of virus copies present in our samples. For this, we used a pectoral muscle sample from a Sulfur-crested cockatoo (Cacatua galerita) that was diagnosed with PBFD based on advanced clinical signs and subsequently confirmed by qPCR as in Martens et al.41. After extraction, we ran a conventional PCR by using the same protocol as above for the qPCR, but with ultrapure water instead of the probe. Then we ran the DNA on agar gel and pooled together the band obtained at approximately 100 base pair (bp) containing the BFDV DNA. We used a DNA high-sensitivity chip in the Bioanalyzer 2100 (Agilent Technologies, USA) to obtain the number of virus copies present in the BFDV DNA sample using three different concentrations (neat, 1:10 and 1:100). We followed the formula based on the assumption that the average weight of a bp is 650 Daltons:
We obtained the average “number of copies/µL” from the concentration of the three positive samples. The purified PCR product was used to create the standard by making aliquots and creating a fresh dilution series containing a known number of copies for each qPCR. Then we did tenfold serial dilutions of the pooled samples and ran them on the qPCR to assess which concentration would give the best detection sensitivity for our samples. We determined that the final standard curve would be between dilutions of 10−4 and 10−10. This standard curve was included in each qPCR run, which allowed us to determine the absolute virus genome load (referred to as virus load for the purpose of this study) for each sample.
DNA extraction and quantification by qPCR
We analysed a total of 813 tissue samples from n = 66 wild P. elegans (Table 1). For blood samples stored in ethanol, DNA was extracted using the Mechanical Lysis Module of the MagMAX™ CORE Nucleic Acid Purification Kit (Applied Biosystems, TX, USA). We followed the low-input workflow from the manufacturer’s protocol for the Mechanical Lysis Module, with a slight modification as described in Blanch-Lázaro et al.42. For other tissue samples stored in RNAlater, DNA was extracted using the MagMAX™ 96 Viral RNA Isolation Kit (Applied Biosystems, USA). A small piece of tissue (approx. 0.1 × 0.1 mm) was added into a tube with 250 µL of phosphate-buffered saline (PBS) and a Stainless Steel Bead (Ø5 mm; QIAGEN, Germany) and lysed using the TissueLyser II (QIAGEN, Germany) at 25 Hz for 2 min. After centrifuging for 1 min at 5000 rpm, we used the supernatant for the DNA extraction. DNA extraction was done in the KingFisher™ Flex (ThermoScientific, USA) extraction robot, following the manufacturer’s guidelines for the “MagMAX Pathogen high volume” script. To each extraction plate we added a known BFDV-positive sample from a PBFD-affected C. galerita and PBS, as positive and negative extraction controls respectively. To ensure the correct DNA extraction from each sample, we quantified the purified nucleic acid using the QIAxpert system (Qiagen, Germany). This sample was then either run on real-time quantitative PCR (qPCR) or stored frozen at −80 °C until analysis. Even though ribosomal RNA and mitochondrial DNA may have also been extracted using this kit, its contribution to the total nucleic acid readout would be minimal compared to the magnitude differences reported between tissues. Furthermore, we tend to compare the relative amount of virus in adults and subadults for the same tissues, so any such effects would not affect our results.
To determine the presence of BFDV DNA, we followed the probe-based qPCR method detailed in Eastwood et al.28, which uses a forward (5′-GAC GCG GAT AAC GAG AAG TAT TG-3′) and a reverse (5′-GCA ACA GCT CCA TCG AAA GC-3′) primer to target a 100 nucleotide region in the open reading frame of the replication-associated protein. In brief, the final reaction contained 2 µL extracted DNA, 1 µL of each primer (9 µM stock forward and 3 µM stock reverse; Applied Biosystems, USA), 5 µL of Brilliant Multiplex qPCR master mix (Agilent Technologies, USA) and 1 µL of probe (1 µM stock; Integrated DNA Technologies, USA). We used the QuantStudio™ 6 and 7 Flex real-time PCR system (Applied biosystems, Thermo Fisher Scientific, USA). The thermal cycling conditions included an initial denaturation of 10 min at 95 °C, followed by 40 cycles of 30 s at 95 °C, 1 min at 60 °C and 30 s at 72 °C, and a final extension step of 5 min at 72 °C. We ran each sample in duplicate and repeated them if the quantification cycle (Cq) value differed more than one cycle between duplicates28,42,43. We used a more conservative detection limit for BFDV (of Cq = 36, where greater values were considered negative) following previous published studies42,43,67.
We used the mean quantity values of each sample obtained on QuantStudio™ Real-Time PCR Software (v1.3; Applied Biosystems, Thermo Fisher Scientific, USA), together with the DNA concentration value of each sample obtained on the QIAxpert system (Qiagen, Germany) prior to the qPCR. The genome mean DNA content of Psittaciformes is 2.93 pg/nucleus68. Assuming a parrot cell contains about 3 pg DNA per cell and knowing the volume of sample per each qPCR reaction, we extrapolated the amount of BFDV copies per 106 cells present in our BFDV-positive samples (on qPCR, Cq < 36). We transformed the resulting virus load (log10) to achieve normality for statistical analysis as done in previous BFDV studies27,29.
Separately, three near-full length BFDV genomes were obtained by NGS of multiple overlapping PCR amplicons from P. elegans bursa samples included in our study (accession numbers OR122675, OR122676 and OR122677 published in GenBank; BioProject PRJNA979100 in the NCBI Short Read Archive (SRA) and as described in the Supplementary Methods and Supplementary Tables 2–6). This was achieved using two different primer pools, each with primers different from the ones used for the real-time PCR. To understand if qPCR efficiency could be influenced significantly by genetic diversity of BFDV at the primer and probe sites (Eastwood et al.28 had demonstrated the reliability and reproducibility of this qPCR in samples from P. elegans) we blasted the primers and probe used in our study against our three BFDV genomes as well as other available BFDV sequences from P. elegans available in GenBank using the NCBI Nucleotide Basic Local Alignment Search Tool (BLASTn) online algorithm. The alignments are shown in Supplementary Fig. 4. While some minor mismatches were found (e.g. 1–3 nucleotide mismatches at the 5′ end of primers, and 1–2 nucleotide mismatches at the 3′ end of the probe), these findings indicate that it is highly unlikely that the large differences in BFDV load observed in this study would be caused by sequence variation in the primer and probe region (the minimum nucleotide identity between BFDV sequences from P. elegans for the entire qPCR region was 93.5%). The more significant mismatches were mostly detected when blasting against BFDV sequences isolated from families of psittacine birds different to P. elegans, which is in line with previously published phylogenetic analyses of BFDV27,69.
To determine if the sequence differences in the qPCR target could potentially cause the huge differences (3 log10 difference or more) in virus load seen between adult and subadult birds in our study, we designed a sequencing PCR to amplify the entire qPCR amplicon and flanking regions (see Supplementary Methods) and used serial dilutions of purified sequencing PCR amplicons from multiple birds to calculate PCR efficiency. Samples used for this section included tissues of four subadults (high virus load) and four adults (low virus loads) (see Supplementary Methods). Within the forward primer, all circovirus sequences shared two common nucleotide differences to the forward primer, identical to those seen in the whole genome sequencing described above (Supplementary Fig. 4), and two adult and two subadult sequences had three nucleotide differences (Supplementary Table 7). There was no significant difference in the amplification rate per PCR cycle between sequences with two or three nucleotide differences (two-tailed t-test, p = 0.7, df = 15). At the probe site, circoviruses from two adult and four subadult tissues were a perfect match to the probe with no nucleotide differences. All other subadult and adult circoviruses and the three circovirus genomes described above shared a common single nucleotide difference (Supplementary Fig. 4, Supplementary Table 8). There was no significant difference between the rate of amplification between circoviruses with one or no nucleotide differences at the probe site (two-tailed t-test, p = 0.57, df = 15). At the reverse primer binding site, all sequences shared a common nucleotide difference identical to that seen in the whole genome sequencing (Supplementary Fig. 4), while two adult and two subadult circoviruses shared a second nucleotide difference (Supplementary Table 9). There was no statistically significant difference between the amplification rates of sequences with one or two nucleotide differences (two-tailed t-test, p = 0.06, df = 15). The amplification rate per PCR cycle varied between 1.55 and 1.99 between the P. elegans circovirus samples (Supplementary Table 10). While there was some variation in the amplification rate of the different circovirus sequences in the samples, there was no significant difference (two-tailed t-test, p = 0.498, df = 15) in the amplification rates between tissues from subadult versus adult P. elegans (Supplementary Fig. 5).
Haemagglutinin inhibition (HI) assay
We sent dried blood spots on filter paper (n = 65) to the Veterinary Diagnostic Laboratory (VDL), Charles Sturt University in Wagga Wagga, New South Wales, Australia, to determine the presence of antibodies to BFDV. Filter papers were transported at room temperature by express post, from Deakin University in Geelong, Victoria, Australia. Haemagglutination inhibition (HI) assays were carried out commercially by staff at VDL as described in Riddoch et al.65 for dried blood spots on filter paper42. HI titres were obtained from twofold serial dilutions starting at 1:20. Results were reported as the highest dilution of antibodies causing complete inhibition of the hemagglutination, with HI titres < 1:20 being considered negative or undetectable.
Statistics and reproducibility
Statistical analyses were performed in SPSS Statistics (v 26.0, IBM Corp, Armonk NY, USA). To test the effects of age and sex on the detection rate of BFDV in tissue samples we ran a generalised linear mixed model (GLMM). The response variable was the BFDV status (non-infected or infected) of each sample and categorical predictors were age class (adult, subadult), sex, tissue type and the two-way interactions between age class and tissue type, as well as sex and tissue type. We used binomial distribution (link Logit) and a robust estimation. To test the effects of age class and sex on virus load we ran a GLMM with virus load (log10) as the response with the same predictors as above including the addition of the three-way interaction between age class, sex, and tissue type. This model used a normal distribution (link identity). For this model we included bird ID as a random intercept to account for multiple samples from each individual and adjusted for multiple comparisons using the sequential Bonferroni procedure. Sample sizes and number of replicates are reported in sections “Sample collection” and “DNA extraction and quantification by qPCR”, and Table 1.
Figures were created using GraphPad Prism (v8.0.0 for Windows, GraphPad Software, San Diego, CA, USA, www.graphpad.com), Microsoft Excel (version 2108) and SPSS Statistics (v 26.0, IBM Corp, Armonk NY, USA).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All source data that support the findings of this study are available in the Supplementary Data 1 and Supplementary Information files. All sequence data have been deposited in GenBank with accession code numbers OR122675, OR122676, OR122677 and numbers PP496486 to PP496501, and in the NCBI Short Read Archive (SRA) as BioProject PRJNA979100. The raw PCR data generated during this study are available from the corresponding author on reasonable request.
References
Smith, K. F., Acevedo-Whitehouse, K. & Pedersen, A. B. The role of infectious diseases in biological conservation. Anim. Conserv. 12, 1–12 (2009).
de Castro, F. & Bolker, B. Mechanisms of disease-induced extinction. Ecol. Lett. 8, 117–126 (2005).
Scheele, B. C. et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 363, 1459–1463 (2019).
Atkinson, C. T. & LaPointe, D. A. Introduced avian diseases, climate change, and the future of Hawaiian honeycreepers. J. Avian Med. Surg. 23, 53–63 (2009).
Samuel, M. D., Woodworth, B. L., Atkinson, C. T., Hart, P. J. & LaPointe, D. A. The epidemiology of avian pox and interaction with avian malaria in Hawaiian forest birds. Ecol. Monogr. 88, 621–637 (2018).
Raidal, S. R. & Peters, A. Psittacine beak and feather disease: ecology and implications for conservation. Emu Austral Ornithol 118, 80–93 (2018).
Olah, G. et al. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 25, 205–223 (2016).
Australian Department of the Environment and Heritage. Threat Abatement Plan for Beak and Feather Disease Virus Affecting Endangered Psittacine Species. https://www.dcceew.gov.au/sites/default/files/documents/beak-feather-tap.pdf (Canberra, Commonwealth of Australia, 2005).
Fogell, D. J. et al. Trade and conservation implications of new Beak and feather disease virus detection in native and introduced parrots. Conserv. Biol. 32, 1325–1335 (2018).
Raidal, S. R., Sarker, S. & Peters, A. Review of psittacine beak and feather disease and its effect on Australian endangered species. Aust. Vet. J. 93, 466–470 (2015).
Department of the Environment. Psittacine Beak and Feather Disease and other identified Threats to Australian threatened Parrots. Australian Government. http://www.environment.gov.au/system/files/resources/9b85a93b-00c1-43d2-9e4f-bb6279e956e6/files/beak-feather-disease-and-other-threats-australian-threatened-parrots.pdf (2015).
Jackson, B. et al. Clinical beak and feather disease virus infection in wild juvenile eastern rosellas of New Zealand; biosecurity implications for wildlife care facilities. N. Z. Vet. J. 62, 297–301 (2014).
Massaro, M. et al. Molecular characterisation of beak and feather disease virus (BFDV) in New Zealand and its implications for managing an infectious disease. Arch. Virol. 157, 1651–1663 (2012).
Peters, A. et al. Evidence of Psittacine Beak and Feather Disease Virus spillover into wild critically endangered orange-bellied parrots (Neophema chrysogaster). J. Wildl. Dis. 50, 288–296 (2014).
Das, S. et al. Repeat spillover of beak and feather disease virus into an endangered parrot highlights the risk associated with endemic pathogen loss in endangered species. J. Wildl. Dis. 56, 896–906 (2020).
Jackson, B. et al. Emerging infectious disease or evidence of endemicity? A multi-season study of beak and feather disease virus in wild red-crowned parakeets (Cyanoramphus novaezelandiae). Arch. Virol. 160, 2283–2292 (2015).
Kundu, S. et al. Tracking viral evolution during a disease outbreak: the rapid and complete selective sweep of a circovirus in the endangered echo parakeet. J. Virol. 86, 5221–5229 (2012).
Regnard, G. L., Boyes, R. S., Martin, R. O., Hitzeroth, I. I. & Rybicki, E. P. Beak and feather disease viruses circulating in Cape parrots (Poicepahlus robustus) in South Africa. Arch. Virol. 160, 47–54 (2015).
Todd, D. Circoviruses: Immunosuppressive threats to avian species: a review. Avian Pathol. 29, 373–394 (2000).
Fogell, D. J., Martin, R. O. & Groombridge, J. J. Beak and feather disease virus in wild and captive parrots: an analysis of geographic and taxonomic distribution and methodological trends. Arch. Virol. 161, 2059–2074 (2016).
Sarker, S. et al. Mutability dynamics of an emergent single stranded DNA virus in a naïve host. PLoS ONE 9, e85370 (2014).
Schoemaker, A. N. J. et al. Severe leukopenia and liver necrosis in young african grey parrots (Psittacus erithacus erithacus) infected with psittacine circovirus. Avian Dis. 44, 470–478 (2000).
Raidal, S. R. & Cross, G. M. The haemagglutination spectrum of psittacine beak and feather disease virus. Avian Pathol. 23, 621–630 (1994).
Regnard, G. L., Boyes, R. S., Martin, R. O., Hitzeroth, I. I. & Rybicki, E. P. Beak and feather disease virus: correlation between viral load and clinical signs in wild Cape parrots (Poicepahlus robustus) in South Africa. Arch. Virol. 160, 47–54 (2015).
Ha, H. J., Anderson, I. L., Alley, M. R., Springett, B. P. & Gartrell, B. D. The prevalence of beak and feather disease virus infection in wild populations of parrots and cockatoos in New Zealand. N. Z. Vet. J. 55, 235–238 (2007).
Pass, D. A. & Perry, R. A. The pathology of psittacine beak and feather disease. Aust. Vet. J. 61, 69–74 (1984).
Eastwood, J. R. et al. Phylogenetic analysis of beak and feather disease virus across a host ring-species complex. Proc. Natl. Acad. Sci. 111, 14153–14158 (2014).
Eastwood, J. R. et al. Prevalence of beak and feather disease virus in wild Platycercus elegans: Comparison of three tissue types using a probe-based real-time qPCR test. Aust. J. Zool. 63, 1–8 (2015).
Martens, J. M. et al. Beak and feather disease virus (BFDV) prevalence, load and excretion in seven species of wild caught common Australian parrots. PLoS ONE 15, e0235406 (2020).
Ha, H. J., Alley, M. R., Cahill, J. I., Howe, L. & Gartrell, B. D. The prevalence of psittacine beak and feather disease virus infection in native parrots in New Zealand. N. Z. Vet. J. 57, 50–52 (2009).
Rahaus, M. & Wolff, M. H. Psittacine beak and feather disease: a first survey of the distribution of beak and feather disease virus inside the population of captive Psittacine birds in Germany. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 50, 368–371 (2003).
Valastanova, M., Petrikova, M., Kulikova, L. & Knotek, Z. Psittacine beak and feather disease virus and avian polyomavirus detection rate in clinically healthy captive birds in the Czech Republic. Vet. Med. 66, 72–75 (2021).
Circella, E. et al. Psittacine beak and feather disease – like illness in Gouldian Finches (Chloebia gouldiae). Avian Dis. 58, 482–487 (2014).
Sarker, S. et al. Evidence of a deep viral host switch event with beak and feather disease virus infection in rainbow bee-eaters (Merops ornatus). Sci. Rep. 5, 14511 (2015).
Sarker, S., Lloyd, C., Forwood, J. & Raidal, S. R. Forensic genetic evidence of beak and feather disease virus infection in a Powerful Owl, Ninox strenua. Emu Austral Ornithol. 116, 71–74 (2016).
Amery-Gale, J. et al. A high prevalence of beak and feather disease virus in non-psittacine Australian birds. J. Med. Microbiol. 66, 1005–1013 (2017).
Downs, C. T., Brown, M., Hart, L. & Symes, C. T. Review of documented beak and feather disease virus cases in wild Cape parrots in South Africa during the last 20 years. J. Ornithol. 156, 867–875 (2015).
Araújo, A. et al. Molecular diagnosis of beak and feather disease in native Brazilian Psittacines. Rev. Bras. Ciência Avícola 17, 451–458 (2015).
Morinha, F., Carrete, M., Tella, J. L. & Blanco, G. High prevalence of novel beak and feather disease virus in sympatric invasive parakeets introduced to spain from Asia and South America. Diversity 12, 192 (2020).
Eastwood, J. R. et al. Prevalence of BFDV in wild breeding Platycercus elegans. J. Ornithol. 1, 3 (2019).
Martens, J. M., Stokes, H. S., Berg, M. L., Walder, K. & Bennett, A. T. D. Seasonal fluctuation of beak and feather disease virus (BFDV) infection in wild Crimson Rosellas (Platycercus elegans). Sci. Rep. 10, 7894 (2020).
Blanch-Lázaro, B., Ribot, R. F. H., Berg, M. L., Alexandersen, S. & Bennett, A. T. D. Ability to detect antibodies to beak and feather disease virus in blood on filter paper decreases with duration of storage. PeerJ 8, e12642 (2021).
Martens, J. M. et al. Persistence of beak and feather disease virus (BFDV) infection in wild Crimson Rosellas (Platycercus elegans). Emu Austral Ornithol 119, 402–406 (2019).
Segalés, J., Allan, G. M. & Domingo, M. Porcine circovirus diseases. Anim. Health Res. Rev. 6, 119–142 (2005).
Schat, K. A. Chicken anemia virus. Curr. Top. Microbiol. Immunol. 331, 151–183 (2009).
Segalés, J. et al. Quantification of porcine circovirus type 2 (PCV2) DNA in serum and tonsillar, nasal, tracheo-bronchial, urinary and faecal swabs of pigs with and without postweaning multisystemic wasting syndrome (PMWS). Vet. Microbiol. 111, 223–229 (2005).
Yaqoob Wani, M., Dhama, K. & Singh Malik, Y. Impact of virus load on immunocytological and histopathological parameters during clinical chicken anemia virus (CAV) infection in poultry. Microb. Pathog. 96, 42–51 (2016).
Alexandersen, S. Pathogenesis of disease caused by Aleutian mink disease parvovirus. APMIS Suppl. 14, 1–32 (1990).
Fogell, D. J. et al. Evolution of beak and feather disease virus across three decades of conservation intervention for population recovery of the mauritius parakeet. Diversity 13, 584 (2021).
Sutherland, M. et al. Disease surveillance in wild Victorian cacatuids reveals co-infection with multiple agents and detection of novel avian viruses. Vet. Microbiol. 235, 257–264 (2019).
Das, S. et al. Psittacine beak and feather disease virus in wild Orange-bellied Parrots. Assoc. Avian Vet. Australas. Comm. Ltd. Annu. Confer. 23, 18–27 (2015).
Uttenthal, A. et al. Analysis of experimental mink enteritis virus infection in mink: in situ hybridization, serology, and histopathology. J. Virol. 64, 2768–2779 (1990).
Hansen, M. S. et al. Detection of porcine circovirus Type 2 and viral replication by in situ hybridization in primary lymphoid organs from naturally and experimentally infected pigs. Vet. Pathol. 50, 980–988 (2013).
Paterson, S. Structure and function of bird skin. In Skin Diseases of Exotic Pets 151–172 (Blackwell Publishing, 2006). https://doi.org/10.1002/9780470752432.ch13.
Eastwood, J. R. et al. Host heterozygosity and genotype rarity affect viral dynamics in an avian subspecies complex. Sci. Rep. 7, 1–11 (2017).
Raidal, S. R., Bonne, N. J., Stewart, M. & Shearer, P. Standardised Diagnostic Tests for Beak and Feather Disease Virus. https://www.dcceew.gov.au/sites/default/files/documents/49540.pdf (2008).
Latimer, K. S. et al. An updated review of psittacine beak and feather disease. J. Assoc. Avian Vet. 5, 211 (1991).
Smyth, J. A., Weston, J., Moffett, D. A. & Todd, D. Detection of circovirus infection in pigeons by in situ hybridization using cloned DNA probes. J. Vet. Diagnostic Investig. 13, 475–482 (2001).
Smyth, J., Soike, D., Moffett, D., Weston, J. H. & Todd, D. Circovirus-infected geese studied by in situ hybridization. Avian Pathol. 34, 227–232 (2005).
Pyne, M. Psittacine beak and feather disease. In Currumbin Wildlife Sanctuary, Gold Coast. National Wildlife Rehabilitation Conference 2005. 1–4. https://doi.org/10.1136/vr.120.12.287-a (2005).
Hulbert, C. L. et al. Survey of captive parrot populations around Port Phillip Bay, Victoria, Australia, for psittacine beak and feather disease virus, avian polyomavirus and psittacine adenovirus. Aust. Vet. J. 93, 287–292 (2015).
Darwich, L., Segalés, J. & Mateu, E. Pathogenesis of postweaning multisystemic wasting syndrome caused by Porcine circovirus 2: An immune riddle. Arch. Virol. 149, 857–874 (2004).
Latimer, K. S. et al. Extracutaneous viral inclusions in psittacine beak and feather disease. J. Vet. Diagnostic Investig. 2, 204–207 (1990).
Raidal, S. R., Sabine, M. & Cross, G. Laboratory diagnosis of psittacine beak and feather disease by haemagglutination and haemagglutination inhibition. Aust. Vet. J. 70, 133–137 (1993).
Riddoch, P. A., Raidal, S. R. & Cross, G. M. Psittacine circovirus antibody detection and an update on the methods for diagnosis of psittacine beak and feather disease. Aust. Vet. Pract. 26, 134–139 (1996).
Stanford, M. Control of circovirus infection in psittacine birds using F10SC disinfectant and avian gamma interferon. The Facts 2, 1–4 (2006).
Martens, J. M. et al. A non-invasive method to assess environmental contamination with avian pathogens: beak and feather disease virus (BFDV) detection in nest boxes. PeerJ 8, e9211 (2020).
Costantini, D., Racheli, L., Cavallo, D. & Dell’omo, G. Genome size variation in parrots: longevity and flying ability. J. Avian Biol. 39, 453–459 (2008).
Sarker, S. et al. Phylogeny of beak and feather disease virus in cockatoos demonstrates host generalism and multiple-variant infections within Psittaciformes. Virology 460–461, 72–82 (2014).
Acknowledgements
The authors thank the landowners and managers for their cooperation during sample collection. We also thank Helena Stokes, Johanne Martens, Justin Eastwood, Candice Lachenicht, Kate Buchanan and Scott Edwards for their advice on BFDV research, and four anonymous reviewers for their comments on the paper. This work was supported by an Australian Research Council (Discovery grant DP180103494 to Andrew T.D. Bennett, Soren Alexandersen, Scott Edwards and Mathew L. Berg), a Deakin University PhD stipend to Berta Blanch Lázaro, the Holsworth Wildlife Research Endowment, the Wildlife Disease Association-Australasia, Birdlife Australia and the University of Melbourne. The various funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
B.B.L., M.L.B., S.A., A.T.D.B conceived and designed the study. B.B.L., R.F.H.R., M.L.B., A.T.D.B. conducted field work and sample collection. B.B.L., A.C., T.R.B, M.L.B., S.A., A.T.D.B. conceived and performed the laboratory work. B.B.L., M.L.B., A.C., S.A. performed data analyses. B.B.L. wrote the paper. B.B.L., A.C., R.F.H.R., M.L.B., S.A., A.T.D.B. reviewed drafts and approved final draft.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Martina Jelocnik, Deborah Fogell and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Tobias Goris and Johannes Stortz.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Blanch-Lázaro, B., Chamings, A., Ribot, R.F.H. et al. Beak and feather disease virus (BFDV) persists in tissues of asymptomatic wild Crimson Rosellas. Commun Biol 7, 1017 (2024). https://doi.org/10.1038/s42003-024-06652-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-024-06652-1
