
Abstract
Biodiversity–ecosystem functioning experiments have established generally positive species richness-productivity relationships in plots of single ecosystem types, typically grassland or forest. However, it remains unclear whether these findings apply in real-world landscapes that resemble a heterogeneous mosaic of different ecosystem and plant types that interact through biotic and abiotic processes. Here, we show that landscape-level diversity, measured as number of land-cover types (different ecosystems) per 250×250 m, is positively related to landscape-wide remotely-sensed primary production across all of North America, covering 16 of 18 ecoregions of Earth. At higher landscape diversity, productivity was temporally more stable, and 20-year greening trends were accelerated. These effects occurred independent of local species diversity, suggesting emergent mechanisms at hitherto neglected levels of biological organization. Specifically, mechanisms related to interactions among land-cover types unfold at the scale of entire landscapes, similar to, but not necessarily resulting from, interactions between species within single ecosystems.
Similar content being viewed by others

Introduction
Hundreds of biodiversity–ecosystem functioning (BEF) experiments across a vast range of experimental systems1 have provided broad evidence that plant communities containing more species on average have higher levels of ecosystem functioning, for example higher biomass production2,3,4. This finding has raised concerns that the ongoing global loss of biodiversity could impair ecosystem service delivery to humans5. Indeed, observational studies have shown that, after accounting for environmental drivers of productivity, ecosystem functions such as productivity are reduced when species diversity is low6,7,8.
An important feature of real-world landscapes is their mosaic of different, spatially integrated ecosystem types (e.g., grasslands, forests, aquatic ecosystems, urban areas). A consequence of this structural heterogeneity is that the diversity present in such landscapes can vary at levels of organization not found in small experimental plots that each represent only a single ecosystem type. First, species turnover occurs between ecosystems, giving rise to higher landscape-level (γ) species diversity than found within each component ecosystem (α-species diversity), and leading to complex dispersal and extinction dynamics in such interconnected communities9. Such meta-population10 and meta-community11 dynamics can promote local species richness, which in turn could promote local and, therefore, landscape-wide functioning. Second, there is evidence that ecosystems also interact via abiotic exchanges, e.g., energy and matter (carbon, water, and nutrients), which sometimes greatly modifies the functioning of ecosystems12,13,14,15.
The need to study diversity–productivity relationships in the real-world and at large scales is uncontested5,16,17,18. The few studies conducted at correspondingly large scales mostly focused on the effects of species diversity7,18. Here, we explored whether real-world landscape diversity—landscape functioning (LD-LF) relationships also emerged at levels of biological organization higher than species15. Specifically, we were interested in the effects of the diversity of land-cover (LC) types (a proxy for ecosystem-type diversity) found in a landscape. Early indications for such effects originate from a pilot study covering Switzerland19, but it is unclear whether such effects are general and important across continents and biomes. Analyzing a 20-year time series of satellite-sensed primary production covering North America, we found that landscape-level productivity increased with LCR and that productivity became temporally more stable. We further found landscape productivity increases over the observation period, and that this greening trend was accelerated in more LC-rich landscapes. Finally, species inventory data suggested that the landscape diversity effects we report here occurred in addition to the positive effects of local (α-)species richness.
Results and discussion
Decorrelation of environmental drivers from landscape diversity
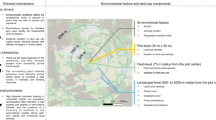
To test the hypothesis that LCR benefits landscape-wide productivity, we divided the entire continent of North America into 250 × 250 m entities (hereafter referred to as “landscapes”) and derived their LC-type composition and richness (LCR) from 30-m-resolution LC maps. To isolate LCR effects from effects of co-varying environmental conditions20,21, we systematically chose landscapes that formed a quasi-experimental set-up22. In brief, we first blocked North America into 3° latitude × 6° longitude tiles and further by climate (ecoregions sensu23) (Fig. 1), in analogy to randomized plot-scale BEF experiments that use blocks to statistically absorb spatial variation. We then constructed replicated LCR gradients within each block (starting from a richness of one, a single LC-type landscape, to a maximum of four LC-types in a landscape), using the LC types prevailing in that area. In addition to grassland, shrubland, forest, and wetland, we also considered urban and agricultural areas as LC types, given that real-world landscapes contain anthropogenic elements that may also interact with their more natural surroundings. Within each block, and for each pre-determined LCC, we then used a stochastic optimization process (Methods) to select a set of landscape plots for which LCR was decorrelated from abiotic factors known to be related to productivity (Fig. 2). In this way, we are isolating the effect of richness while keeping all else constant within each block. When we could not obtain a full richness gradient with all these conditions met, we removed individual LCs not replicated frequently enough in the mixtures that were part of the gradient. If this did not help, we dropped the entire block from the analysis.

North America was divided into blocks defined by the combination of climate (ecoregions) and space (3˚ latitude × 6˚ longitude grid). In total, the study design encompassed 298 blocks, with a total of 58,280 250×250 m landscape plots. These plots were composed of six different land-cover (LC) types (A: agriculture, F: forest, G: grassland, S: shrubland, U: urban, and W: wetland), resulting in 56 unique LC compositions. Thereby, gradients in land-cover type richness (LCR) were constructed within each block so that the different land covers were represented equally at all levels of LCR (e.g., if a block containing the LC types A, G, and U, the 2-LC mixtures AG, AU, GU were included, and the 3-LC mixture AGU; see Methods and Supplementary Table 1). Each LC composition was replicated 20 times per block.

To avoid a statistical confounding of landscape diversity, here measured as land-cover type richness (LCR), with other potential drivers of productivity, we used a stochastic subsampling technique that minimized correlations between these (Methods). The panels on the left show, in the full set of landscape plots, and for each block (Fig. 1), the correlations of LCR with the altitude, the north gradient of the slope, and the fraction of a landscape plot covered with a particular land-cover (only forest shown here as an example). With such a quasi-experimental study design and by probabilistic subsampling we obtained a dataset where the correlations of the landscape’s properties with LCR were minimized. Histograms at the right of the panels show the distribution of each variable.
Landscape productivity and stability increase with landscape diversity
As a metric of landscape functioning, we determined productivity proxies from 20-year time series (2000–2019) of a satellite-sensed vegetation index [MODIS Enhanced Vegetation Index (EVI)24, 250-m resolution, see Methods]. To account for productivity differences among ecoregions, we calculated a standardized vegetation index (EVI’) by dividing EVI values by the mean of all EVI data of the respective ecoregion. We then integrated EVI’ values over the growing seasons (see “Methods”), yielding EVIgs’. EVIgs’ statistically significantly increased with LCR (Fig. 3a, F1,53 = 9.4, P = 0.003). We then determined the net diversity effect (NE), i.e., the gain in EVIgs’ in the LC mixture relative to the average EVIgs’ of the corresponding single-LC landscapes25. Again, values of EVIgs’ in mixed landscapes significantly exceeded the ones of single-LC landscapes (Fig. 3c, t49 = 9.3, P < 0.001), and NE increased progressively with the number of LC types present in mixed landscapes (Fig. 3b, F1,51 = 8.7, P = 0.005). Similar effects were detected for growing-season peak and mean values of EVI’ (Table 1). We further found that landscape productivity was temporally more stable in mixed LC-type landscapes, evidenced in a larger inverse coefficient of inter-annual variation in mixed landscapes (Fig. 3d, F1,54 = 9.5, P = 0.003, Table 1).

Land-cover-type richness (LCR) effect on a normalized growing-season-integrated productivity (EVIgs’), b the net diversity effect (NE) calculated as the difference between observed and expected EVIgs’ in mixed landscapes, with (c) the distribution of the NE{EVIgs’} values of each LC type compositions shown as histogram, and (d) inverse coefficient of inter-annual variation of EVIgs’ (CV-1{EVIgs’}). Black lines and areas shaded in blue: model-predicted mean ± s.e.m; dots: averages for each land-cover composition. See Table 1 for corresponding statistical significance tests.
In our study, mixed LC landscapes had, on average, 4.2% higher EVIgs’ than single-LC landscapes. This effect may appear small compared to the increases in primary productivity in the range of a few ten percent that often are reported for traditional BEF experiments26. In these experiments, communities are usually established with an equal initial density of species. However, as communities develop, the relative abundance of species adjusts, with typically a few plant species that dominate cover and drive high community-level functioning after a few years27. A related phenomenon is species that fail: these are included in the calculation of the monoculture average, which serves as a reference when determining net biodiversity effects, but in mixtures, the space and resources they originally used become available to successful species that increase in abundance during the plot’s ‘internal’ community assembly28,29. In our study, such effects could not occur because the area covered by each LC type was held constant in the landscapes, i.e. no landscape-level EVI gain could be achieved by an expansion of the area covered by productive LC types. In this light, the net diversity effects in EVIgs’ of up to 5.4% (which we found at the highest LCR levels of 4, compared to single-LC landscapes) are highly conservative estimates and compare to yield gains deemed important in application contexts such as agriculture30,31,32.
Landscape diversity is associated with accelerated decadal-scale greening trends
Over the 20-year observation period, all our measures of productivity increased. For EVIgs’, the average gain across the entire study region amounted to 8.7% (an increase from 2000 to 2019 based on linear model predictions). This decadal trend varied among ecoregions and was generally larger in cooler and higher-latitude areas than in warmer and lower-latitude areas (Fig. 4a), in line with recent global analyses33,34,35. Importantly, we detected a statistically significant positive effect of LCR on these trends (F1,52 = 4.1, P = 0.048), i.e., the decadal increase in productivity was accelerated in mixed LC-type landscapes compared to single-LC-type landscapes (Fig. 4b). This association of landscape diversity with more pronounced signatures of global change on productivity resembles earlier finding that more species-rich landscapes—quantified as γ-diversity—show an accelerated growing-season lengthening8.

a 20-year trend (years 2000–2019) in growing-season-integrated EVI’ (EVIgs’) shown by ecoregion (see Fig. 1). b Net diversity effect on 20-year trend in EVIgs’. Data are shown as modeled linear trends relative to model-predicted values for the first year (2000). The gray areas represent the blocks that were dropped from the analysis.
Landscape diversity effects occur independent of local α-species diversity
The positive LC-type diversity–landscape functioning relationship we found resembles the species diversity–ecosystem functioning relationships reported from individual plot-scale experiments to a remarkable extent, suggesting that similarly-shaped diversity effects also occur at higher levels of biological organization. To test whether the LCR effects found were caused indirectly by higher local species richness, we used a separate dataset of landscapes that included tree-species inventory data from the Forest Inventory and Analysis Database of the United States of America (FIA)36. We adopted the same quasi-experimental design, but due to the lower number of available replicates, we blocked landscape plots at the level of ecoregions instead of the ecoregion × latitude/longitude tile combination. Modeling the net diversity effect, we detected independent positive effects of both local (α) tree-species richness (EVIgs’ F1,1.2 = 162.6, P = 0.035) and of LCR (EVIgs’ F1,0.1 = 170.6, P = 0.035). Yet, tree-species richness and LCR were not significantly correlated (Pearson’s product-moment correlation r = 0.04, t268 = 0.6, P = 0.5). This indicates that tree diversity promotes forest functioning, as has been demonstrated in observational studies6,37. More importantly, it further suggests that additional effects independent of local tree-species richness support landscape-level functioning. This demonstrates beyond species-level diversity effects on landscape-wide ecosystem functioning across an entire continent. Forests are the most productive LC type in our dataset and we therefore consider it unlikely that the local species diversity of other ecosystem types, which here were not available for analysis, would fundamentally change this conclusion.
Robustness of landscape diversity effects
Our study, while leveraging the benefits of systematic experimental designs, remains observational by nature. We, therefore, challenged our findings in several ways. First, to verify that our findings did not depend on the particular LC map we chose, we repeated all analyses with an independent dataset constructed using a different 30-m resolution global LC map with a different and coarser LC-type classification38. The results quantitatively and qualitatively confirmed the benefits of LCR for all measures of productivity, although effect sizes and significances varied to some extent (Supplementary Fig. 1; Supplementary Table 2). Second, we tested whether our findings depended on the presence of any particular LC type (Methods). To this end, we dropped each individual LC type from our dataset using a jack-knife approach and re-ran all analyses. Again, the results stayed consistent (Table 2), with net diversity effects that remained statistically significantly positive in all cases. Hence, this indicates that interactions among dissimilar LC types, whatever the nature of the specific underlying mechanisms, add up to higher functioning levels in landscapes with heterogeneous land cover, and that this effect does not depend on a single particular LC type but is spread across a wide set of LC combinations. To test whether specific LC combinations were particularly beneficial, we compared the net diversity effects of all 50 distinct LC mixtures present in our study. Indeed, these differed (NE EVIgs’: F1,49 = 6.6, P < 0.001), with responses ranging from neutral to positive, and none of the statistically significant effects being negative (Fig. 5).

Ranked net diversity effects on growing-season-integrated EVI (NE{EVIgs’}) in mixed LC-type landscapes. Black dots and error bars: mean ± s.e.m. for each LC composition. Red: NE is significantly positive; Gray: NE is not significantly different from zero; Blue: NE is significantly negative. A: agriculture, F: forest, G: grassland, S: shrub, U: urban, and W: wetland.https://datadryad.org/stash/dataset/doi:10.5061/dryad.v41ns1s3p.
Potential mechanisms underpinning landscape diversity effects
Which mechanisms, in addition to the diversity of primary producers, could promote the productivity of mixed LC landscapes? To date, this topic is not well investigated in the frame of diversity–functioning research, but evidence of beneficial interactions among different ecosystem types exists. First, landscape heterogeneity may modulate the effects of taxa other than plants on primary productivity. For example, agricultural studies have shown that habitat heterogeneity can promote pollinators and natural enemies of pests, increasing crop productivity16,39,40. Landscape heterogeneity may also reduce the long-range spread of enemies such as pathogens41,42. It is conceivable that these benefits extend to non-agricultural systems and could support primary productivity at the landscape scale. Second, different ecosystem types exchange abiotic resources such as carbon, and nutrients14,43,44. An example is the transfer of leaf litter and dung across the terrestrial-aquatic interface45 that can benefit the functioning of both ecosystems46,47. In this context, anthropogenically-managed LC types can have particularly large landscape-wide consequences48; for example, the deposition of nitrogen from anthropogenic sources enhances forest growth in many areas of the Western United States, although detrimental effects also occur49. Third, there might be effects driven by microclimate modifications. It is well established that vegetation around cities can moderate urban heat islands50,51, and that the surrounding vegetation can, in turn, benefit from a longer growing season52. The exchange of heat among different LC types is particularly complex because differences in the surface energy balance of LC types will create convection cells at LC boundaries53, where warm air in hotter patches rises and cooler moister air from the neighboring colder patch takes its place54,53. These flows not only re-distribute heat but can alter the average climate in the entire landscape55,56, with potential direct effects on plant growth and productivity.
The LCR effects we found document that phenomena operating beyond relatively small and uniform plots of BEF experiments can modify primary productivity at large scales relevant for global plant productivity and climate dynamics57. It is very likely that not all interactions among LC types are beneficial to productivity, and that such effects will depend on context such as edaphic and climatic conditions. However, our results indicate that these emergent landscape-level effects are—on average—positive. Two general phenomena may be of particular importance. First, “buffering” effects may occur at the landscape level; for example, wetlands can prevent flooding by absorbing excess water, and buffer droughts and fires by releasing moisture, thus providing environmental stability58,59. Similarly, patches of forest can reduce the impact of extreme weather events, providing moisture and cooling through evapotranspiration, as well as flood moderation at larger scales60,61. At the species level, landscape-level heterogeneity can foster ecosystem resilience by providing regional species pools that buffer detrimental local effects on species diversity62,63. Second, and related to the first phenomenon, a redistribution of resources from LC types in which these occur at high abundance to other LC types resembles a mixing process that spatially “averages out” differences in resource levels between LC types. For example, some LC types, like urban areas, heat up more strongly than others, like forests. In a mixed LC-type landscape, local temperature extremes will thereby be moderated, with adjacent forests leading to lower maximum temperatures in urban areas, and urban areas leading to higher minimum temperatures in forest. Similar reasoning applies to other mobile resources, such as nutrients. An interesting possibility, therefore, is that such a landscape-wide mixing could be beneficial per se for landscape functioning because it reduces potentially negative impacts of extreme variation64,65.
In our study, landscapes composed of multiple LC types not only were more productive, but productivity also was temporally more stable. Two mutually non-exclusive reasons may account for the increased stability in the more LC-diverse landscapes. First, the interactions among the different LC types discussed above may temporally stabilize the productivity of the individual LCs. One candidate mechanism for such stabilization is the buffering or environmental extremes. Second, landscape-wide stability in productivity may also arise from an asynchrony in the productivity of the individual LCs. These mechanisms have been discussed with respect to species richness64,66,67; for our study of LC type richness, the statistical patterns underlying increased temporal stability could not be studied in detail because the remote-sensing data we used here had too low a resolution to assess the productivity contributions of the individual LC types to total mixture productivity.
Conclusions
For over two decades, concerns about the applicability of BEF findings to real-world systems have been raised5,68. By adopting a quasi-experimental study design at large spatial scales across an entire continent, covering the majority of ecoregions present on Earth, we here show that variation in landscape-scale functioning can be explained by the real-world diversity of higher units of ecological organization than species, namely of entire LC types. The remarkable consistency of the pattern across biomes and including man-made ecosystems consolidates the idea that diversity at more than one level is essential to sustaining Earth’s productivity and stability.
Methods
Study area
North America (6˚N 139˚W to 62˚ N 12˚W) was divided into ≈520 million landscape plots that correspond to the 250-m resolution pixels of the MODIS satellite instrument24 that we used to quantify landscape functioning.
Landscape diversity
For each landscape plot, land-cover type richness (LCR) was determined by extracting 30-m spatial resolution LC information (1) from the Commission for Environmental Cooperation’s North American Land Monitoring System’s map (CEC map, based on Landsat-7 satellite imagery69), and (2) from the global GlobeLand30 map (GLC map, based on Landsat-5 and China Environmental Disaster Alleviation Satellite (HJ−1) imagery38). Comparable, evenly distributed LCs with reliable remotely sensed EVI readings were chosen: forest, grassland, shrubland, agriculture, wetland, and urban. To obtain comparable LC classes between maps, the different forest types distinguished in the CEC map were combined.
Quasi-experimental study design
The entire study area was divided into 3° latitude × 6° longitude tiles, and then further into ecoregions, based on a map of 16 out of 18 global environmental zones23. The zones “arctic” and “extremely cool and wet” were dropped due to a lack of productivity in the entire region. This spatial and environmental stratification defined the block structure and was used to separate the local LCR effects we were interested in from large-scale variation in environmental conditions, in the sets of LC types and particular vegetation types present in any region, and in regional species pool. Within each block, parallel sub-designs relating landscape functioning to LCR were established, following design principles from experimental BEF research. Specifically, our goals were to establish landscape plot sets that (1) reflected the dominant LC types present in each block, (2) for which LCR was orthogonal to (i.e. not correlated with) the average fraction of each particular LC type found at each LCR level, (3) for which LCR was orthogonal to potentially important environmental factors.
First, we calculated altitude, slope inclination, and the north-south component of the slope gradient for each landscape plot using the TanDEM-X 90-m resolution digital elevation model70. We then removed all landscape plots that did not fall within 0–2000 m altitude, all landscape plots with LC fractions that deviated more than 50% from perfect LC evenness, and all landscape plots for which less than 50% of the potential satellite observations were available (e.g. missing data due to cloud cover). Within each block, we then searched for a subset of landscape plots that minimized a cost function designed (1) to decorrelate LCR from the fractional contribution of each LC type, altitude, slope inclination, and the North-aspect of the slope gradient (Fig. 2), (2) to maximize LC evenness of the mixtures, and (3) to maximize the minimum distance between landscape plots of the same LC composition. This was achieved using simulated annealing, a probabilistic technique for approximating global optima in a large search space71. The algorithm is driven by a random walk taken during an imaginary cooling process in which the likelihood of uphill moves decreases as temperature declines. Per block, 100 million iterations were performed (>60 billion iterations for the entire study), with the algorithm typically converging after around 20 million steps.
This plot selection procedure yielded three distinct data sets. The first set of landscape plots is based on the CEC map (289 blocks with a total of 65,140 landscape plots representing 56 distinct LC compositions). The second set of landscape plots is based on the GLC map (299 blocks, 52,620 plots, 42 LC compositions). A third set of landscape plots was built using the CEC map, but only considering forest plots for which local species richness was available through forest inventories compiled by the FIA36. This smaller dataset comprised 270 landscape plots with 16 distinct LC compositions. Due to limited data, blocking for this third dataset was done by ecoregion (n = 4), ignoring tile. The FIA inventory plots varied in size, and we therefore down-sampled species richness (SR) by rarefying SR to the number of individuals expected in a standard area of 0.1 ha. Rarefied SR values averaged 4.68 across all plots.
Landscape functioning
Vegetation indices acquired by the MODIS instrument on the Terra satellite were used as a proxy of primary productivity. Specifically, we used the enhanced vegetation index (EVI, data product MOD13Q1), which is similar to the normalized difference vegetation index (NDVI) except that it uses blue-band data to adjust for atmospheric bias due to aerosols24. We fitted phenology models to EVI time series (years 2000–2019) using a modified Harmonious Analysis of Time Series (HANTS) algorithm based on a Fourier synthesis8,72. The HANTS model comprised of three harmonics and was fitted five times after each iteration replacing EVI values by predicted values when residuals exceeded 0.5, 0.2, 0.1, and 0.05 raw units. This robust fitting method effectively smooths the time series and eliminates spurious EVI values that occur, for example, because of undetected cloud interference.
Next, we determined the growing season (GS), which is an important aspect of land-surface phenology, using the NDVI-ratio method73. Specifically, the start of the GS (SOS) was determined as the first day at which EVI values exceeded the mean of their minimum and maximum value. Similarly, the end of the GS (EOS) was determined as the last day at which EVI fell below this threshold.
Next, we determined peak EVI (EVImax), GS-integrated EVI (EVIgs), and the average GS EVI (EVImean) for each pixel8. This was done using SOS and EOS values characterizing the potential GS in the respective ecoregion because the determination of SOS and EOS by the NDVI-ratio method is not very robust at the single-pixel level, in particular when pixels contain mosaics of different LC types that differ in phenology, for example, due to management in agriculture. The potential growing season of an ecoregion was identified by sampling 10,000 pixels from each block, and SOS (EOS) was determined as the 25th (75th) percentile of the distribution of SOS and EOS values obtained from the HANTS fits to 20-year EVI time series (see above). HANTS phenology models were then integrated from the ecoregion’s potential SOS to EOS to yield EVIGS and EVImean. Naturally, primary productivity (and therefore also EVI values) follows latitudinal and altitudinal trends. Since we were not interested in the variation of EVI among ecoregions, we normalized all EVI metrics by dividing them by the mean EVI value of the respective ecoregion (EVImax’, EVIgs’, and EVImean’). In this way, landscape diversity effects become relative and comparable across North America. The temporal stability of productivity was then calculated as the inverse coefficient of inter-annual variation (CV-1; years 2000–2019) of each response variable.
Net diversity effects (NE), also referred to as overyielding25, capture the difference in mixture productivity and the average productivity of the single LC-type landscapes (equivalent to monocultures in species-diversity experiments) contained in that mixture. NE was calculated within each block, weighing the single-LC values by the actual cover of the respective LC in the mixture.
Statistical methods
Effects of landscape diversity on landscape functioning were tested using general linear mixed models summarized by analysis of variance (ANOVA) with sequential (type I) tests. Landscape diversity was a fixed effect, and block and LC composition were included in the model as random effects74. Irrespective of landscape diversity, EVI values differ systematically between two broad groups of LC types: the first group, consisting of forest, agriculture, and wetland, had systematically larger EVI values than the second group, which consisted of grassland, shrubland, and urban. To remove this systematic effect from residuals, which should only contain random variation, a fixed model term was added that captures the fraction of the low-productivity LC types present in a landscape. For the analyses of the effects of local tree-species richness (SR) on the EVI of the landscape plot, we used the logarithm of rarefied species richness multiplied by the percentage of forest cover as a predictor. Temporal trends over the 20-year observation period were analyzed by first extracting the linear trend for each composition × block combination. These slopes were then averaged by composition, and the resulting slopes were analyzed using general linear mixed models summarized by analysis of variance (ANOVA).
Data availability
The data used in this study, and all data shown in figures are available on Dryad, under DOI 10.5061/dryad.v41ns1s3p.
Code availability
Code used for statistical analyses, together with results, are available on Zenode, at https://zenodo.org/records/13628018.
References
O’Connor, M. I. et al. A general biodiversity-function relationship is mediated by trophic level. Oikos 126, 18–31 (2017).
Roscher, C. et al. Overyielding in experimental grassland communities—irrespective of species pool or spatial scale. Ecol. Lett. 8, 419–429 (2005).
Reich, P. B. et al. Impacts of biodiversity loss escalate through time as redundancy fades. Science 336, 589–592 (2012).
Weisser, W. W. et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 23, 1–73 (2017).
Isbell, F. et al. Linking the influence and dependence of people on biodiversity across scales. Nature 546, 65–72 (2017).
Liang, J. et al. Positive biodiversity-productivity relationship predominant in global forests. Science 354, aaf8957 (2016).
Barnes, A. D. et al. Species richness and biomass explain spatial turnover in ecosystem functioning across tropical and temperate ecosystems. Philos. Trans. R. Soc. B: Biol. Sci. 371, 20150279 (2016).
Oehri, J., Schmid, B., Schaepman-Strub, G. & Niklaus, P. A. Biodiversity promotes primary productivity and growing season lengthening at the landscape scale. Proc. Natl Acad. Sci. USA 114, 10160–10165 (2017).
Liao, J. et al. Modelling plant population size and extinction thresholds from habitat loss and habitat fragmentation: effects of neighbouring competition and dispersal strategy. Ecol. Model. 268, 9–17 (2013).
Hanski, I. Metapopulation dynamics. Nature 396, 41–49 (1998).
Leibold, M. A. et al. The metacommunity concept: a framework for multi-scale community ecology. Ecol. Lett. 7, 601–613 (2004).
Shen, W., Lin, Y., Jenerette, G. D. & Wu, J. Blowing litter across a landscape: effects on ecosystem nutrient flux and implications for landscape management. Landsc. Ecol. 26, 629–644 (2011).
Didham, R. K. et al. Agricultural intensification exacerbates spillover effects on soil biogeochemistry in adjacent forest remnants. PLoS ONE 10, e0116474 (2015).
Gounand, I., Harvey, E., Little, C. J. & Altermatt, F. Meta-ecosystems 2.0: rooting the theory into the field. Trends Ecol. Evol. 33, 36–46 (2018).
Mayor, S. et al. Diversity–functioning relationships across hierarchies of biological organization. Oikos e10225 (2023).
Fahrig, L. et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 14, 101–112 (2011).
Tscharntke, T. et al. Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol. Rev. 87, 661–685 (2012).
Gonzalez, A. et al. Scaling-up biodiversity-ecosystem functioning research. Ecol. Lett. 23, 757–776 (2020).
Oehri, J., Schmid, B., Schaepman-Strub, G. & Niklaus, P. A. Terrestrial land-cover type richness is positively linked to landscape-level functioning. Nat. Commun. 11, 154 (2020).
Gough, L., Grace, J. B. & Taylor, K. L. The relationship between species richness and community biomass: the importance of environmental variables. Oikos 70, 271–279 (1994).
Grace, J. B. et al. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 529, 390–393 (2016).
Imbens, G. W. & Rubin, D. B. Causal Inference for Statistics, Social, and Biomedical Sciences: An Introduction. (Cambridge University Press, Cambridge, 2015).
Metzger, M. J. et al. A high-resolution bioclimate map of the world: a unifying framework for global biodiversity research and monitoring. Glob. Ecol. Biogeogr. 22, 630–638 (2013).
Huete, A., Didan, K., van Leeuwen, W., Miura, T. & Glenn, E. MODIS Vegetation Indices. Vol. 11 (Springer, Dordrecht, 2011).
Schmid, B., Hector, A., Saha, P. & Loreau, M. Biodiversity effects and transgressive overyielding. J. Plant Ecol. 1, 95–102 (2008).
Cardinale, B. J. et al. The functional role of producer diversity in ecosystems. Am. J. Bot. 98, 572–592 (2011).
Crawford, M. S. et al. The function-dominance correlation drives the direction and strength of biodiversity–ecosystem functioning relationships. Ecol. Lett. 24, 1762–1775 (2021).
Pillai, P. & Gouhier, T. C. Not even wrong: the spurious measurement of biodiversity’s effects on ecosystem functioning. Ecology 100, e02645 (2019).
Hagan, J. G., Vanschoenwinkel, B. & Gamfeldt, L. We should not necessarily expect positive relationships between biodiversity and ecosystem functioning in observational field data. Ecol. Lett. 24, 2537–2548 (2021).
Thirtle, C., Irz, X., Lin, L. & Wiggins, S. The relationship between changes in agricultural productivity and the incidence of poverty in developing countries. Extension to Report 7946 of the UK Department for International Development (DFID), London (2001).
Tao, F., Yokozawa, M., Liu, J. & Zhang, Z. Climate–crop yield relationships at provincial scales in China and the impacts of recent climate trends. Clim. Res. 38, 83–94 (2008).
Edgerton, M. D. et al. Transgenic insect resistance traits increase corn yield and yield stability. Nat. Biotechnol. 30, 493–496 (2012).
Zhu, W. et al. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 21, 260–271 (2012).
Kong, D., Zhang, Q., Singh, V. P. & Shi, P. Seasonal vegetation response to climate change in the Northern Hemisphere (1982–2013). Glob. Planet. Change 148, 1–8 (2017).
Campbell, J. E. et al. Large historical growth in global terrestrial gross primary production. Nature 544, 84–87 (2017).
Gray, A., Brandeis, T., Shaw, J., McWilliams, W. & Miles, P. Forest inventory and analysis database of the United States of America (FIA). Biodivers. Ecol. 4, 225–231 (2012).
Barrufol, M. et al. Biodiversity promotes tree growth during succession in subtropical forest. PLoS ONE 8, e81246 (2013).
Chen, J. & Chen, J. GlobeLand30: Operational global land cover mapping and big-data analysis. Sci. China Earth Sci. 61, 1533–1534 (2018).
Martin, E. A. et al. The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol. Lett. 22, 1083–1094 (2019).
Massaloux, D., Sarrazin, B., Roume, A., Tolon, V. & Wezel, A. Landscape diversity and field border density enhance carabid diversity in adjacent grasslands and cereal fields. Landsc. Ecol. 35, 1857–1873 (2020).
Real, L. A. & Biek, R. Spatial dynamics and genetics of infectious diseases on heterogeneous landscapes. J. R. Soc. Interface 4, 935–948 (2007).
Jones, E. O., Webb, S. D., Ruiz-Fons, F. J., Albon, S. & Gilbert, L. The effect of landscape heterogeneity and host movement on a tick-borne pathogen. Theor. Ecol. 4, 435–448 (2011).
Gounand, I., Little, C. J., Harvey, E. & Altermatt, F. Cross-ecosystem carbon flows connecting ecosystems worldwide. Nat. Commun. 9, 4825 (2018).
Scherer-Lorenzen, M. et al. Pathways for cross-boundary effects of biodiversity on ecosystem functioning. Trends Ecol. Evol. 37, 454–467 (2022).
Gratton, C., Donaldson, J. & Zanden, M. J. V. Ecosystem linkages between lakes and the surrounding terrestrial landscape in northeast Iceland. Ecosystems 11, 764–774 (2008).
Pusey, B. J. & Arthington, A. H. Importance of the riparian zone to the conservation and management of freshwater fish: a review. Mar. Freshw. Res. 54, 1–16 (2003).
Frainer, A., Polvi, L. E., Jansson, R. & McKie, B. G. Enhanced ecosystem functioning following stream restoration: the roles of habitat heterogeneity and invertebrate species traits. J. Appl. Ecol. 55, 377–385 (2018).
Kaye, J. P., Groffman, P. M., Grimm, N. B., Baker, L. A. & Pouyat, R. V. A distinct urban biogeochemistry? Trends Ecol. Evol. 21, 192–199 (2006).
Fenn, M. E. et al. Ecological effects of nitrogen deposition in the western United States. BioScience 53, 404–420 (2003).
Qiu, G. et al. Effects of evapotranspiration on mitigation of urban temperature by vegetation and urban agriculture. J. Integr. Agri. 12, 1307–1315 (2013).
Manoli, G. et al. Magnitude of urban heat islands largely explained by climate and population. Nature 573, 55–62 (2019).
Dallimer, M., Tang, Z., Gaston, K. J. & Davies, Z. G. The extent of shifts in vegetation phenology between rural and urban areas within a human-dominated region. Ecol. Evol. 6, 1942–1953 (2016).
Foley, J. A., Costa, M. H., Delire, C., Ramankutty, N. & Snyder, P. Green surprise? How terrestrial ecosystems could affect Earth’s climate. Front. Ecol. Environ. 1, 38–44 (2003).
Pielke, R. A. S. Influence of the spatial distribution of vegetation and soils on the prediction of cumulus convective rainfall. Rev. Geophys. 39, 151–177 (2001).
Segal, M., Avissar, R., Mccumber, M. & Pielke, R. Evaluation of vegetation effects on the generation and modification of mesoscale circulations. J. Atmos. Sci. 45, 2268–2292 (1988).
Mendes, C. B. & Prevedello, J. A. Does habitat fragmentation affect landscape-level temperatures? A global analysis. Landsc. Ecol. 35, 1743–1756 (2020).
de Jong, R., Schaepman, M. E., Furrer, R., de Bruin, S. & Verburg, P. H. Spatial relationship between climatologies and changes in global vegetation activity. Glob. Change Biol. 19, 1953–1964 (2013).
Brinson, M. M. & Eckles, S. D. U. S. Department of Agriculture conservation program and practice effects on wetland ecosystem services: a synthesis. Ecol. Appl. 21, S116–S127 (2011).
Narayan, S. et al. The value of coastal wetlands for flood damage reduction in the northeastern USA. Sci. Rep. 7, 9463 (2017).
Ellison, D. et al. Trees, forests and water: Cool insights for a hot world. Glob. Environ. Change 43, 51–61 (2017).
Gohr, C., Blumröder, J. S., Sheil, D. & Ibisch, P. L. Quantifying the mitigation of temperature extremes by forests and wetlands in a temperate landscape. Ecol. Inform. 66, 101442 (2021).
Oliver, T. H. et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 30, 673–684 (2015).
Isbell, F. et al. Quantifying effects of biodiversity on ecosystem functioning across times and places. Ecol. Lett. 21, 763–778 (2018).
Wang, S. et al. Biotic homogenization destabilizes ecosystem functioning by decreasing spatial asynchrony. Ecology 102, e03332 (2021).
Qiao, X. et al. Spatial asynchrony matters more than alpha stability in stabilizing ecosystem productivity in a large temperate forest region. Glob. Ecol. Biogeogr. 31, 1133–1146 (2022).
Sasaki, T., Lu, X., Hirota, M. & Bai, Y. Species asynchrony and response diversity determine multifunctional stability of natural grasslands. J. Ecol. 107, 1862–1875 (2019).
Schnabel, F. et al. Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment. Sci. Adv. 7, eabk1643 (2021).
Duffy, J. E. Why biodiversity is important to the functioning of real-world ecosystems. Front. Ecol. Environ. 7, 437–444 (2009).
CEC. Land Cover of 2010 (Landsat, 30m). http://www.cec.org/north-american-environmental-atlas/land-cover-2010-landsat-30m/ (Commission for Environmental Cooperation, 2017).
German Aerospace Center (DLR). TanDEM-X 90m DEM. https://tandemx-science.dlr.de/ (German Aerospace Center (DLR), 2018).
Kirkpatrick, S., Gelatt, C. & Vecchi, M. Optimization by simulated annealing. Science 220, 671–680 (1983).
Roerink, G. J., Menenti, M. & Verhoef, W. Reconstructing cloudfree NDVI composites using Fourier analysis of time series. Int. J. Remote Sens. 21, 1911–1917 (2000).
White, M. A., Thornton, P. E. & Running, S. W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 11, 217–234 (1997).
Schmid, B., Baruffol, M., Wang, Z. & Niklaus, P. A. A guide to analyzing biodiversity experiments. J. Plant Ecol. 10, 91–110 (2017).
Acknowledgements
This study was funded by the University of Zurich Research Priority Program Global Change and Biodiversity (URPP GCB) grant to P.A.N.
Author information
Authors and Affiliations
Contributions
P.A.N., B.S., F.A., and M.E.S. conceived the idea and acquired funding. P.A.N. and S.M. designed the study. S.M. implemented the study and analyzed the data, with support from P.A.N. T.W.C., and I.H. helped with species-level data and analysis. S.L. contributed to data curation and verified all final analyses. S.M. and P.A.N. wrote the first draft. F.M., T.W.C., I.H., S.L., J.O., M.R.C., M.E.S., and B.S. contributed to the data interpretation and the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Youhua Chen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Huai Chen and Joe Aslin. [A peer review file is available.]
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Mayor, S., Altermatt, F., Crowther, T.W. et al. Landscape diversity promotes landscape functioning in North America. Commun Earth Environ 6, 28 (2025). https://doi.org/10.1038/s43247-025-02000-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02000-1
