
Abstract
Seagrass meadows facilitate the capture and storage of organic carbon, but spatial variability in carbon has been observed among and within meadows. Here we combine sediment and seagrass data with aerial images collected near the Annisquam River, Massachusetts, USA to examine spatial variations in carbon retention across a patchy seagrass meadow. Tidal velocities were reduced within patches and elevated in bare regions, which was expected to promote carbon accumulation within the patches. However, organic carbon was not correlated with the spatial distributions of seagrass or velocity. Historical aerial images showed continual patch movement, with vegetation persistence of less than a decade throughout the meadow. The highest carbon stock occurred in the largest area of recent vegetation persistence. Though present for >45 years, the meadow accumulated negligible carbon, likely due to the migration of patches. Overall, we provide insight into a potential limitation on carbon accretion and storage in patchy meadows.
Similar content being viewed by others


Introduction
Seagrasses are marine flowering plants that can form dense meadows along every continent except Antarctica1. The ability of seagrass meadows to accumulate and retain sedimentary organic carbon has received growing interest for the potential of meadow restoration projects to compensate for anthropogenic carbon dioxide emissions2,3. However, substantial variability in sediment organic carbon content has been observed both among different seagrass meadows4,5 and within the same meadow6,7, which complicates carbon budget accounting. A wide range of environmental factors influences the spatial differences in carbon content and accretion rates within and among seagrass meadows, including seagrass species8, hydrodynamic exposure6,9, sediment grain size, which can be a function of hydrodynamic exposure8,10, climatic region5, proximity to carbon sources11, biogeochemistry4, and landscape configuration, i.e., patchy versus continuous meadows7,11,12.
When an array of individual shoots forms a meadow (also known as a canopy), meadow-scale drag significantly reduces the current velocity experienced by individual shoots in the meadow13,14, and also reduces wave energy propagating across the meadow15,16. In this way, seagrasses engineer the local ecosystem to promote their survival17. Further, by reducing currents and waves, a meadow creates conditions that favor deposition and accretion of sediment. Marine organic matter, derived from both seagrass biomass (known as autochthonous carbon) and non-seagrass sources (known as allochthonous carbon) can be preserved through adsorption on fine sediments (silts and clays)18,19, such that the promotion of deposition within a meadow also promotes carbon accretion. Whether an individual carbon-laden particle will deposit within a meadow depends on the relative timescales of deposition and advection through the meadow, which in turn depends on the size of the meadow. Further, seagrass roots and rhizomes can exude complex oxygen compounds and sugars, which may inhibit decomposition, such that the roots and rhizomes of some seagrass species may promote organic carbon preservation20.
Seagrass meadows are disappearing rapidly on a global scale due to human activities and climate change21. The loss of seagrass may result in enhanced fragmentation of once continuous meadows into patches of varying scales22,23,24. Patchiness may also result from natural disturbances25 and growth through seed dispersal26. Patchy configurations of seagrass can create complex mosaics of lower velocity regions within patches separated by higher velocity regions within sufficiently large and connected bare regions between patches27,28. This contrasting hydrodynamic exposure could promote spatial heterogeneity in sediment organic carbon content, with carbon accretion and storage enhanced within the patches but suppressed in bare regions. However, depending on the density of shoots in the meadow and the sediment grain size, it is possible that the presence of seagrass may provide little effective protection for organic carbon particles, and thus would not promote locally enhanced carbon accretion29.
Only a few studies have examined spatial patterns in sediment organic carbon across patchy seagrass meadows. For large (100 m) patches of Zostera muelleri Irmisch ex Asch., Ricart et al.7 observed that sediment organic carbon content in the top 10 cm of sediment was lower near the patch edge, which they attributed to higher hydrodynamic exposure and younger shoot ages near patch edges. A similar observation (lower sediment organic carbon at the meadow edge) was made by Oreska et al.30, who proposed that a large continuous meadow should be more effective at accumulating organic carbon than a patchy meadow of the same overall size, due to the lower proportion of meadow area near an edge. Consistent with this, Ricart et al.12 observed that carbon stock in the upper 2 cm of sediment was higher in continuous meadows than in nearby patchy meadows of Posidonia oceanica (L.) Delile, where patches were typically 2 m × 2 m in size. When considering the influence of landscape configuration on sediment carbon storage, the size of the patches, as well as the species of seagrass, is important (see the discussions of landscape fragmentation scales in Bunnell31; Wu32). In particular, a vegetated patch must be of sufficient size and shoot density to reduce current velocity and create a region of distinct hydrodynamic exposure13,28,33, which might lead to a region of enhanced sediment deposition. Similarly, a bare patch within a meadow must be of sufficient size to accelerate the flow and experience enhanced velocities relative to the nearby vegetated patches. However, Asplund et al.11 found that patchy landscape metrics, such as mean patch size and patch edge perimeter, did not consistently predict spatial patterns in carbon storage across different patchy meadows in Tsimipaika Bay, Madagascar. Therefore, patch size is not the only factor controlling spatial variability in carbon stocks in patchy meadows.
The present study sought to contribute additional insight into patchy meadow dynamics by combining measurements of hydrodynamic exposure and carbon stocks within a patchy seagrass (Zostera marina Linnaeus, also known as eelgrass) meadow. The field site was Norwood Heights Beach near the Annisquam River in Gloucester, Massachusetts, USA. The original hypothesis was that, at a current-dominated site, hydrodynamic intensity would be lower within patches, compared to adjected bare areas, which would result in higher sediment organic carbon content and carbon accretion rates within the patches. However, this hypothesis was proven to be wrong. Although current and resuspension were lower within patches, compared to the bare regions, this did not translate into enhanced organic carbon stock within patches. Instead, the spatial variation in carbon stock seemed to be related to the persistence of individual patches within a meadow of shifting patches. By revealing a dynamically shifting state of individual patches within a meadow, this study provides insight into the vulnerability of seagrass sediment carbon pools in patchy landscapes.
Results
Stable isotope measurements
Comparing the carbon stable isotope ratios δ13C between seagrass and sediment samples provides insight into the similarity of carbon sources. The sediment samples were more depleted in δ13C than the seagrass. Specifically, seagrass leaf δ13C ranged from −13.12 to −10.43‰, sediment trap δ13C ranged from −29.30‰ to −18.14‰, and sediment core δ13C ranged from −28.12‰ to −12.35‰ (Fig. 1). While there was a wide range in individual δ13C measurements in the sediment cores, there were no clear trends with sediment depth in the profiles of δ13C (see example for S1V in Fig. 2, and more details in Supplementary Fig. 1).

Box plots of stable carbon isotope ratio δ13C (parts per thousand relative to Vienna Pee Dee Belemnnite) from seagrass leaves, sediment trap, and sediment cores from vegetated (“V”) stations and from unvegetated bare (“B”) stations. The upper and lower boundaries of each box represent the 75th and 25th quartiles, respectively. The horizontal line within each box denotes the median. The vertical dashed lines extending from the upper and lower boundaries of each box are the whiskers and end at the 95th and 5th percentiles, respectively. Dots denote outliers.

a δ13C (parts per thousand relative to Vienna Pee Dee Belemnnite) and b organic carbon Corg density over cumulative mass depth for station S1V.
Seagrass meadow parameters
Shoot density and morphological measurements for the vegetated stations are given in Supplementary Table 1. The shoot density in July 2022 varied between the measurement stations, with averages ± standard errors of 166 ± 12 shoots m−2 at S1V, 291 ± 17 shoots m−2 at S2V, 300 ± 30 shoots m−2 at S3V, and 210 ± 20 shoots m−2 at S4V. However, the number of leaves per shoot, sheath length, longest leaf length, and shoot width were similar across the vegetated stations. The number of leaves per shoot ranged from 3.80 ± 0.13 at S2V to 4.10 ± 0.10 at S4V (averages ± standard errors). The sheath length ranged from 18.6 ± 1.8 cm at S2V to 19.8 ± 0.9 cm at S4V (averages ± standard errors). The length of the longest leaf (beginning at the top of the sheath) ranged from 9.5 ± 0.9 cm at S2V to 12.3 ± 0.8 cm at S1V (averages ± standard errors). Finally, the shoot width ranged from 0.35 ± 0.04 cm at S3V to 0.43 ± 0.02 cm at S1V (averages ± standard errors).
Hydrodynamic conditions
Moving averages with a 60-min window of velocities measured by the tilt current meters (TCMs), denoted as \({U}_{{{{\rm{c}}}},{{{\rm{TCM}}}}}\), are shown in Fig. 3a, with positive denoting the flood direction and negative denoting the ebb direction. Hourly-averaged wave velocities are shown in Fig. 3b. Because the TCM extended above the height of the meadow, especially during peak tidal velocity, the velocity within the patches, \({U}_{1}\), was only 30-50% of the velocity measured by the TCM, \({U}_{{{{\rm{c}}}},{{{\rm{TCM}}}}}\). In bare regions, the peak current velocities were typically twice as large as peak wave velocities (except on day 11), indicating that on most days of the deployment, tidal currents dominated hydrodynamic exposure and, therefore, sediment transport. This difference was smaller within vegetation patches, where current velocity experienced greater reduction relative to adjacent bare areas than wave velocity. A higher current velocity outside the patches can reinforce the patch edge by reducing the probability of colonization of bare areas34,35,36.

a Time series of 60-min moving average velocity \({U}_{{{{\rm{c}}}},{{{\rm{TCM}}}}}\). b Time series of hourly-averaged wave velocity \({U}_{{{{\rm{w}}}}}\), defined as \(\sqrt{2}\) multiplied by hourly-averaged root-mean-square velocity, \({U}_{{{{\rm{w}}}},{{{\rm{RMS}}}}}\).
Sediment sample analyses
The sediment trap mass deposition rates at bare stations (\(n\)=12) were greater than at vegetated stations (\(n=12\); compare left-hand brown and green bars for each site in Fig. 4a), where the 95% credible interval for the effect of vegetation status on sediment trap mass deposition rate did not include zero (Table 1). The sediment organic carbon content (%OC) in the sediment traps was higher at vegetated stations compared to bare stations (compare right-hand stippled brown and green bars for each site in Fig. 4a), where the 95% credible interval for the effect of vegetation status on %OC did not include zero (Table 1). The average sediment trap mass deposition rate increased with average peak tidal velocity, \(\overline{{U}_{1,{{{\rm{peak}}}}}}\) (Fig. 4b), and the bare stations, which experienced higher velocity, collected higher sediment trap mass compared to vegetated sites. Finally, note that the organic carbon content of the sediment traps was roughly ten times greater than that of the sediment cores, which could be due to the higher lability of the particulate organic matter present in the traps combined with shorter timescales of exposure to microbial decomposition processes.

a Sediment trap mass deposition rate (left bar in each site pair; left axis) and sediment trap percent organic carbon content (right stippled bar in each site pair; right axis). b Sediment trap mass deposition rate versus average peak tidal velocity \(\overline{{U}_{1,{{{\rm{peak}}}}}}\). Sediment organic carbon Corg stocks in the top 27 g cm−2 of sediment cores c at each station and d versus average peak tidal velocity \(\bar{{U}_{1,{{{\rm{peak}}}}}}\). Vertical error bars represent standard errors. Green and brown bars and symbols denote vegetated and bare sites, respectively.
The sediment organic carbon content at each station was characterized by the total carbon stock in the top 27 g cm−2 mass depth of each sediment core (see Supplementary Fig. 2 for sediment core carbon density profiles and Fig. 2b for an example profile; also see Supplementary Fig. 3 for sediment core carbon content profiles). Whether a core location was vegetated or bare at the time of core collection likely had no effect on sediment organic carbon stock (the 95% credible interval for this effect included zero, see Table 1). The vegetated site S3V had higher carbon stock compared to all other sites (Fig. 4c), even though S3V experienced similar velocity to other vegetated sites (green symbols in Fig. 4d). The excess 210Pb activity (beyond background levels of 210Pb) in the core samples were too low to extract a mass accumulation rate, which was likely due to a combination of coarse sediments and sediment mixing37. Further, there were no 137Cs peaks outside of the range of detector uncertainty. Based on the 137Cs and excess 210Pb activity results, we assumed that the long-term average organic carbon accumulation rate in the meadow was zero.
The surface grain size distributions are shown in Supplementary Fig. 4. S3V and S3B had slightly finer median grain sizes than the other stations, 0.13 mm compared to 0.18 mm, likely due to the closer proximity to the mouth of the Annisquam River. The percentage finer than \(\phi\)=3.5 on the Krumbein \(\phi\) scale, equivalent to a diameter of 0.09 mm (the finest threshold tested), was less than 0.01% across the stations.
Summarizing the conclusions drawn from the sediment samples, sediment core organic carbon stocks did not vary with exposure to different peak tidal velocity (Fig. 4d), and were higher at vegetated site S3V, which was not hydrodynamically distinct from the other vegetated sites (Fig. 4d). Therefore, the two-week snapshot of spatial differences in hydrodynamic exposure measured in the summer of 2022 (Fig. 4b, d) cannot explain the observed distribution of sediment organic carbon stocks. Next, we considered how past changes to hydrodynamic spatial variability associated with patch persistence and migration correlated with spatial variability in sediment organic carbon content.
Landscape configuration analysis
Historical aerial and satellite images show that the seagrass patches within the meadow have continuously shifted over time. In other words, the landscape distribution has not been static. Figure 5a shows a heat map of \({N}_{21}\), the frequency of vegetation presence in each pixel between 1972 to 2022 (21 images). Brighter colors indicate areas that have been vegetated more often. Black pixels were never vegetated in any of the images. Importantly, no regions were vegetated for all of the time periods captured by the curated images. The height of each white vertical bar represents the cumulative sediment organic carbon stock down to a 27 g cm−2 mass depth taken at the position of the bar. While S3V had the highest carbon stock, it did not occur in a particularly distinct hotspot of \({N}_{21}\) compared to other sites. Therefore, we narrowed the time frame to the most recent 10 years (one image per year from 2013 to 2022) in Fig. 5b. Within this time frame, the distinctly higher carbon stock at site S3V corresponded with the largest number of years in the last decade with vegetation (\({N}_{{{{\rm{last}}}}10}\), Fig. 6). Station S3V had the highest \({N}_{{{{\rm{last}}}}10}\) and \({N}_{{{{\rm{lastcons}}}}}\) among the stations. Further, S3V was located within a larger contiguous region of high \({N}_{{{{\rm{last}}}}10}\) compared to the other stations.

Heat maps representing the number of times a pixel was vegetated over a all 21 images (1972 to 2022), \({N}_{21}\), and b over the last 10 images, \({N}_{{{{\rm{last}}}}10}\), with one image per year 2013 to 2022. The vertical white bars indicate the cumulative carbon stocks down to a 27 g cm−2 mass depth of the sediment cores at the locations corresponding to the bases of the bars.

Sediment organic carbon stocks in the top 27 g cm−2 of sediment cores versus (a) \({N}_{{{{\rm{last}}}}10}\), the number of times the corresponding station was vegetated within the past 10 years (images), and (b) \({N}_{{{{\rm{lastcons}}}}}\), the number of years the corresponding station was vegetated in consecutive images stepping back from 2022 in year intervals, both calculated as averages over 3-m diameter circles centered at the station GPS locations. Standard errors in \({N}_{{{{\rm{last}}}}10}\) and \({N}_{{{{\rm{lastcons}}}}}\) are smaller than the symbol sizes.
Discussion
The original hypothesis motivating this study was that tidal velocities would be attenuated within the patches (of scale 10 to 20 m), which would preferentially promote the deposition and retention of organic carbon, producing differences in the carbon stock between vegetated and bare regions within the meadow. A two-week snapshot of hydrodynamic conditions confirmed that patches of this size did attenuate tidal velocity. However, the attenuated velocity within patches did not correlate with the observed spatial distribution of carbon stocks. For most bare and vegetated regions in the meadow, the carbon stock was the same, and the single patch with high carbon stock experienced similar hydrodynamic conditions as other patches in the meadow, i.e., the contemporary mosaic of hydrodynamic conditions within the meadow did not explain the observed spatial variation in carbon stock. Factors that might explain the observed spatial distribution in organic carbon stocks are explored next.
The depletion of δ13C in sediment samples compared to seagrass samples (Fig. 1) indicated a dominant contribution to sediment carbon from non-seagrass sources, i.e., allochthonous carbon. The likely sources that are more depleted in δ13C than seagrass are salt marshes along the Annisquam River and terrestrial vegetation within the Annisquam River watershed. For example, common salt marsh plants have δ13C values between −27 and −14‰38. The median δ13C among sediment cores at bare stations was slightly less depleted than that at vegetated stations, which could be related to the exclusion of larger seagrass detrital fragments by the sediment trap baffles as well as localized enhancement of the eelgrass contribution within certain sections of the sediment cores. The presence of predominantly allochthonous carbon suggested the importance of hydrodynamic conditions in determining carbon storage across the meadow. In this seagrass meadow, most organic matter would be expected to originate from the Annisquam River at the southern end of the meadow. A comparison of advection and settling timescales suggested that an even supply of suspended material over the meadow was likely. In saline conditions, clay particles and organic matter form irregularly shaped, porous flocs39,40. The settling velocity of a floc depends on the floc diameter and density, and for an exposed marine environment, such as our site, it has been measured to be roughly 0.5 mm s−1 41. Across the meadow, the average water depth was 1 m at low tide, indicating a lower limit of the floc settling time scale of 2000 s. In contrast, the peak tidal velocities were on the order of 0.1 m s−1, which indicates a transit time of 1500 s across the roughly 150 m meadow. Because the settling time scale is comparable to or larger than the advection time scale, flocs could be delivered to the entire meadow without supply limitation.
If supply is not limiting, differences in net deposition are attributed to differences in resuspension. Based on the near-surface median grain sizes (fine sand), the critical friction velocity \({u}_{* {{{\rm{c}}}}}\) for the initiation of sediment motion is 1.2 cm s−142. Many studies report ratios of depth-averaged current to friction velocity \({U}_{{{{\rm{c}}}}}/{u}_{* }\) = 1043,44, from which we inferred a critical depth-averaged current velocity of 10 cm s−1. The tidal velocity at bare sites exceeded this during ebb tide (Fig. 4d), suggesting the bed material was regularly resuspended at bare sites. In contrast, tidal velocity within the patches (Fig. 4d) was likely too low to regularly resuspend bed material. Higher resuspension rates near bare sites, which would locally enhance suspended sediment concentration, could explain the higher sediment trap mass at bare sites (Fig. 4a). We did not have suspended sediment concentration to support this conjecture. Similarly, lower resuspension within vegetated patches could explain the higher organic carbon content (percentage by weight) in the sediment traps at vegetated sites (Fig. 4a). Organic carbon particles passing through bare regions could mix with higher levels of sediment resuspended from the bed. Note that the small percentages of silt and clay particles at the site suggest that a substantial portion of organic carbon storage could be related to sorption to sand surfaces or deposition as unattached particles. However, it is difficult to determine the relative contributions of the different size fractions to organic carbon preservation.
Sediment can also be remobilized from the bed during high-energy storm events or by ship wakes, but the relative contributions of the tidal currents, storms, and ship wake to sediment resuspension could not be identified from this two-week study. Specifically, higher winds during the winter months (14 to 16 km hr−1 in January through April versus 10 to 11 km hr−1 in July and August, as reported by a local Windfinder weather station) could shift the hydrodynamic conditions from tidal-current-dominated to wave-dominated, which would be more efficient in resuspending material within vegetation, because wave velocity more readily penetrates through the vegetation layer45. In addition, a lower leaf density during winter months could expose the bed to higher currents and, thus, greater sediment mobilization.
Frequent remobilization of sediment could explain the absence of long-term sediment carbon storage. In particular, the organic carbon density in this patchy meadow (1.25 ± 0.05 mg cm−3 Corg averaged across stations ± standard error, but excluding S3V; see Supplementary Fig. 2) was lower than values measured in continuous Z. marina L. meadows in similar substrates (fine to medium sands) in New England reported in Novak et al.46, 5.3 to 15.6 mg cm−3 Corg, and in Lei et al.6, 2.7 to 8.0 mg cm−3 Corg, and comparable to values measured at bare sites adjacent to those meadows (1.0 to 10.4 mg cm−3 Corg in Novak et al.46 and 2.8 to 4.0 mg cm−3 Corg in Lei et al.6. That is, over most of its area, the patchy meadow had not enhanced the sediment carbon density. In contrast, the carbon density at S3V (with an average ± standard error of 4.2 \(\pm\) 0.6 mg cm−3 Corg) fell within the range of other continuous New England meadows. Recall that S3V was within a region of the meadow that maintained vegetation for most of the last 10 years (Fig. 5). It is likely that the other patches have not persisted over sufficient time to accumulate organic carbon. The higher levels of resuspension at the bare sites suggested that once a site becomes bare, the previously stored carbon is remobilized and removed so that a meadow of shifting patches will remain carbon-poor.
Let’s consider how this patchy meadow persists. Patches of Z. marina L. can maintain and expand their populations through clonal (horizontal rhizome extension) or sexual (seedling dispersal) reproduction25. Seed dispersal tends to be more important for large-scale recolonization of unvegetated areas47,48, but seedling burial in one year and growth in a future year can contribute to the spatiotemporal shifts in patch configuration within a meadow49. However, Z. marina L. meadows in shallow water depths, such as the site studied here, are typically more successful at clonal than sexual reproduction47,50,51,52, in which case the survival of the meadow requires the clonal growth rate to outpace the rate of mortality. The effect of clonal growth on patch dynamics has been modeled as a stochastic contact process53, in which at each time step empty parts of a patchy meadow can be colonized with a certain growth probability only if there is a shoot within the range of annual horizontal rhizome extension, and existing seagrass shoots each have a certain probability of mortality. The ratio of growth rate to decay rate has a critical threshold of 1.65, below which patches completely disappear over long periods of time (see Marro & Dickman54).
Marro & Dickman54 only considered biological processes, but the seagrass landscape is also sensitive to hydrodynamic drivers that produce feedback mechanisms that stabilize a patchy distribution. Within vegetated patches of sufficient size, velocity is reduced within the patch (e.g., Fig. 4b), which enhances sediment retention and shoot survival, providing positive feedback to patch stability28,36. Complementary to this, where flow is directed into bare areas between vegetated patches, the local velocity increases, which can reinforce the bare region (another positive feedback) by eroding the edges of patches and reducing the chances of successful seagrass colonization in bare areas55. It is important to note that the positive feedbacks that maintain a patchy meadow structure only exist if the flow can be redirected from vegetated to bare regions, which requires that the bare regions be sufficiently connected to facilitate a continuum of flow34. In other words, an isolated bare patch within a broader meadow will not experience elevated current, because continuity requires that the elevated current have a contiguous channel through which to travel.
Luhar et al.34 used percolation theory to identify the threshold of vegetation coverage at which bare regions of low flow resistance connect into contiguous channels. Consider a two-dimensional grid of squares, where each square has a probability (\(p\)) of being bare and probability (1-\(p\)) of being vegetated. According to percolation theory, at the critical probability \(p\) = 0.593, bare regions can connect and form percolation channels (Table 1 in Stauffer & Aharony56). This threshold is associated with a probability of vegetated area of (1-\(p\))= 0.407. Based on this, Luhar et al.34 hypothesized that if the vegetated area reached 0.407, channels would emerge, and once channels existed, they would persist because of the positive feedback described above. Consistent with this, Luhar et al.34 used observations of Fonseca & Bell57 to show that individual seagrass meadows clustered around two landscape conditions, a continuous meadow (100% area coverage) and a fragmented meadow at 40% area coverage. This demonstrated that a fragmented meadow near the percolation threshold of 40% vegetated area coverage is a stable configuration.
A similar threshold was observed at the field site in this study. The percentage of vegetated area within the meadow area, \({A}_{{{{\rm{v}}}}}/A,\) estimated from the historical images is shown in Fig. 7a. The average \({A}_{{{{\rm{v}}}}}/A\) = 40 ± 3% (mean ± standard error), which is consistent with the percolation theory and hydrodynamic feedback described above. Note that the vegetation coverage was variable between years, sometimes higher or lower than 40%, which could be attributed to the uncertainty in determining the irregularly shaped \(A\) and to the lack of pure randomness in seagrass growth. The persistence of vegetation coverage near 40% suggested that the meadow was stable in a patchy landscape configuration. The meadow has not exceeded the critical threshold that might lead to a continuous meadow. Or, if it has exceeded the threshold, sporadic disturbances reduced the percentage cover to below the threshold (see Duarte58). The meadow was not detected in the 1978 image (open symbol in Fig. 7a), which was likely due to the cyclone and/or notable winter storms in the area between 1975 and 1978. It is possible that there were small patches of surviving seagrass not distinguished by the aerial camera technology available in 1978. Further, the seeds and rhizome fragments buried within the sediment survived and drove the recolonization of the area in the following years.

a Area of vegetation cover \({A}_{{{{\rm{v}}}}}\) within the meadow area \(A\), versus year of the image. In 1978 (open symbol) no vegetation was visible, so the ratio \({A}_{{{{\rm{v}}}}}/A\) was not determinable. b Percentage of vegetation cover in an image of interest that was consecutively vegetated in chronologically earlier images versus the time interval between the images. The symbols outlined in red (connected by red lines) denote comparisons with the July 2022 drone image. More recent images are denoted by darker shades of gray.
Figure 7b shows the history of vegetation persistence marching back in time from different initial time points (different initial images). These data suggest that at each point within the meadow, vegetation persists for less than five years. When jointly considering Fig. 7a and b, the changes in patch distribution over time resemble the shifting mosaic, the steady-state ecological model proposed in Bormann & Likens59 for terrestrial forests. In their model, biomass patterns shift over time as trees propagate or fall, but on the scale of the entire forest, there is no long-term change in total biomass. At the seagrass meadow in this study, while individual patches have actively moved across the landscape, the total seagrass area fraction has remained near the mean value of 40 \(\pm\) 3% (mean ± standard error).
Even though S3V had significantly higher organic carbon compared to other sites in the summer of 2022 (Fig. 4), if it became bare in the future, it would likely lose its elevated sediment organic carbon stocks, returning to the average state represented throughout the rest of the meadow. Rapid changes in surface sediment organic carbon following a shift from vegetated to bare status could result from a combination of enhanced sediment erosion60 and enhanced rates of aerobic decomposition due to higher oxygen exposure at the unvegetated sediment-water surface61. For example, higher near-bed current velocities over bare sediment promote erosion and mixing, as well as enhance oxygen fluxes across the sediment-water interface62. Furthermore, the burrowing and pumping of bivalves and other bio-irrigating organisms both facilitate the diffusion of oxygen into the sediment and enhance sediment mixing63,64. Because these organisms cannot penetrate easily through the dense network of living seagrass roots, their impact is greater in bare sediments65,66,67.
The top meter of sediment is particularly vulnerable to remineralization and erosion when seagrass is lost2,68. For example, Moksnes et al.69 studied several sites off the Swedish coast and recorded >35 cm of erosion following the loss of Z. marina L. meadows 10 to 40 years ago. Macreadie et al.70 found that a disturbance-induced seagrass loss reduced organic carbon in the top meter of sediment in Australian P. oceanica seagrass meadows, but once aboveground biomass returned, half of the carbon lost was recovered within five to ten years. Serrano et al.71 compared seagrass meadows (Amphibolis antarctica [Labill.] and Posidonia australis Hook.f., Shark Bay, Australia) with nearby bare areas that had lost seagrass six years prior. They found significantly lower sediment organic carbon stocks in the previously vegetated bare areas compared to the continuously vegetated areas, indicating a significant loss in carbon over six years. Within the present study, S3V was bare approximately 3 out of the past 10 years prior to 2022 (Fig. 6a), but did not remain bare long enough for the accumulated sediment organic carbon to be lost. Similarly, the other sites vegetated in 2022, were vegetated for only two to four years in the preceding decade (Fig. 6a), and for only one to two years preceding 2022 (Fig. 6b), which was not enough time in a vegetated state to accumulate carbon stock. Considering the previous studies described above alongside the present study, the loss and subsequent recovery of buried sediment organic carbon in seagrass meadows occurs on timescales of five to ten years.
Understanding the effectiveness of seagrass in accumulating and retaining carbon is important for monitoring, reporting, and verifying carbon credits3,72. The present study demonstrated conditions in which a patchy meadow can persist for decades, but not accumulate any permanent carbon stock, which was attributed to the shifting mosaic of patches that comprise the meadow. An aggressive restoration project might lift the \({A}_{{{{\rm{v}}}}}/A\) ratio above the critical percolation threshold to facilitate the coalescence of patches and thereby the stabilization of carbon stores. However, it is possible that the hydrodynamic conditions at this site might drive the system back toward the current dynamic stability of migrating patches. When assessing potential locations for blue carbon and/or restoration, managers should consider not only the historic persistence of a meadow but also the shifting distribution of vegetation within the meadow. Aerial imagery may aid managers in understanding the primary causes of seagrass loss, which should be addressed before beginning a restoration. Furthermore, when choosing locations to measure sediment organic carbon within a seagrass meadow, scientists should use aerial imagery to evaluate the potential influence of landscape configuration on spatial heterogeneity in carbon stores.
Conclusions
The spatial variability in hydrodynamic intensity across a patchy seagrass meadow correlated with the spatial variability in sediment resuspension but did not correlate with the spatial variability in sediment organic carbon stocks. Our hypothesis that organic carbon stocks would be higher within vegetated patches, compared to bare patches, was based on the assumption of a static landscape configuration. However, analysis of historical aerial images revealed that the meadow was in a dynamic, stable state, with individual patches shifting position in time while maintaining an overall coverage fraction close to the percolation limit. The continuous shifting of patch location limited sediment carbon to levels comparable to other bare sites in the region. This sort of historical analysis could provide insight into the suitability of a seagrass meadow for restoration to a contiguous meadow and its potential for carbon storage. If a meadow is patchy and the patches migrate over time, it is likely that the long-term accumulation and retention of organic carbon will be limited.
Methods
Study site
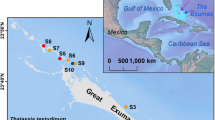
The field site was a patchy meadow of Z. marina L. near the northern mouth of the Annisquam River in Gloucester, Massachusetts, on the East Coast of the United States of America. The Annisquam River is a waterway that cuts through Gloucester. To capture the landscape configuration before field measurements, a drone photographic survey was conducted in July 2022 at 60-m altitude (Fig. 8). Four seagrass patches and four adjacent unvegetated bare areas were chosen as stations, numbered S1 through S4, with a “V” suffix denoting a vegetated patch and a “B” suffix denoting an unvegetated bare area. A boat survey in August 2022 recorded bathymetric contours using side-scan sonar. Mean lower low tide water depth ranged from about 0.5 to 1.5 m (Fig. 8), such that all stations were always submerged. The mean tidal range at the nearby Lobster Cove, Massachusetts tidal gauge station is 2.7 m.

Left-hand image shows the field site in Gloucester, Massachusetts, United States of America. Unvegetated bare (white circles) and vegetated (green circles) measurement stations are marked on a drone orthomosaic image. Bathymetric contours are at mean lower low tide. Map data outside of the drone orthoimage from Google. Bottom right image is a Google composite of images from Scripps Institute of Oceanography, National Oceanic and Atmospheric Administration, United States Navy, National Geospatial-Intelligence Agency, General Bathymetric Chart of the Oceans. The green rectangle in the bottom right image encompasses the left image. Image copyright United States Geological Survey. The top right image of the United States East Coast is copyright Landsat/Copernicus.
Sediment and seagrass sample collection
At each station, suspended sediment was collected in a sediment trap consisting of a horizontal frame holding three cylindrical polyvinyl chloride tubes of length 15 cm and inner diameter 5 cm, with the bottoms of the tubes resting on the bed, following common practice73. Sediment traps were placed at least 5 m into the vegetated or bare region, i.e., at least 5 m from the nearest vegetated-unvegetated edge. The sediment trap tubes were fitted with honeycomb baffles (Plascore, Zeeland, MI) to eliminate sediment resuspension out of the tubes. Sediment traps were deployed for 14 days between July and August 2022.
During the same time-period, one sediment core was taken at each station. To extract a core, a 50-cm long (7.0-cm outer diameter, 6.7-cm inner diameter) polycarbonate tube was manually driven with a T-handled core driver into the sediment by scientific divers to the greatest possible depth, which was limited by rocks. The sediment core lengths ranged from 20 to 30 cm. The sediment cores were capped underwater and kept vertical during transport out of the water to the beach. Core compression during the coring process was recorded by marking the tube at the level of the exterior sediment surface before the core was extracted. On the shore, the sediment cores were systematically extruded into 1-cm thick cylindrical slices between 0 and 20 cm and 2-cm thick slices for sediment deeper than 20 cm. Each slice was stored in a separate pre-weighed Whirl-Pak bag and placed into a cooler for transport to a laboratory. Core shortening during the coring and extrusion processes ranged from 12% to 27%, with an average of 20.4% ± 1.9% (± standard error). Core shortening was not measured for the S2V core. To compare carbon stocks between the cores, we used the equivalent mass approach previously used for seagrass meadows70, mangrove forests74, and discussed for terrestrial systems in Gifford and Roderick75. Specifically, we calculated the cumulative carbon stocks down to a 27 g cm−2 mass depth, which is the shallowest mass depth among the bottom-most slices of the sediment cores. This method avoids the inaccuracies introduced by assuming a uniform and linear core shortening correction factor to convert to sediment depth.
At each vegetated station, seagrass shoots were counted within five haphazardly placed 0.0625 m2 quadrats and used to estimate the density of shoots per bed area. Six to ten shoots were randomly removed from each station to measure the length of the longest leaf above the sheath, sheath length, width, and number of leaves per shoot on the shore. Seagrass samples were stored in plastic bags filled with seawater and kept in a cooler. The second leaf of fifteen of the collected shoots was retained for laboratory analysis.
Laboratory sample analysis
After collection, sediment core, sediment trap, and seagrass samples were transported in a cooler to a laboratory. In the laboratory, each sediment trap tube was uncapped and left alone to allow sediment to settle before the overlying water in the tubes was decanted. The sediment from each trap tube was stored in a separate pre-weighed Whirl-Pak bag. Shellfish, arthropods, shell fragments, as well as living and dead roots and rhizomes, were removed from each sample so that organic carbon measurements represented the long-term storage component46,76. Only a small fraction of seagrass detritus, consisting of microaggregates that can attach to finer sediment particles, is immobilized in the sediment77 and should be considered to be part of long-term carbon stocks. Sediment samples were dried in a drying oven at 60 °C to constant mass and homogenized into powder using a mortar and pestle. In addition, seagrass leaf samples were scraped clean of epiphytes, dried, and ground into powder.
Sediment core slices were also dried at 60 °C to constant mass. The dry mass of each sample was divided by the wet volume of a core slice to calculate the dry bulk density. Animals, shell fragments, and plant matter were removed from the samples before they were homogenized with a mortar and pestle. Sediment grain size distributions were measured for each station by combining subsamples from the top 5 cm of homogenized sediment core slices and dry sieving into the Krumbein \(\phi\) size fractions 3.5 to 0.5, in increments of 0.5.
Stable isotope ratios δ13C (relative to the Vienna Pee Dee Belemnite [V-PDB] international standard) and δ15N (relative to atmospheric air), as well as the dry weight percentage of carbon content (%C) and dry weight percentage of nitrogen content (%N), were measured in tinned subsamples of a sediment trap and sediment core samples and tinned seagrass samples at the Boston University Stable Isotope Laboratory using the Isoprime 100 isotope ratio mass spectrometer (IRMS) interfaced with a Micro Vario Elemental Analyzer (Elementar Americas, Mt. Laurel, New Jersey, USA, with a reported absolute accuracy of <0.1% for a homogeneous standard). Internal laboratory standards (peptone and glycine) were run every 10 to 20 samples. Two mg eelgrass sample masses and (due to low sediment carbon) 60 mg sediment sample masses were used. Based on replicate samples, the relative measurement precision of the instrument was reported to be within 0.2% (e.g., 0.2% relative precision on a measurement of 0.01 (or 1%) means the measurement is 0.01 ± 0.00002, or 1% ± 0.002%). Due to the mechanical removal of visible calcified structures, the latitude of the field site, the local geology, and the low inorganic carbon measured in sediments of similar Z. marina L. meadows in the northeastern USA46, sediment samples were not washed with acid. Additional sediment samples collected at the field site in August 2024 confirmed the assumption of a low inorganic carbon contribution. Specifically, after processing and homogenizing the samples in the same way as described above, we conducted two-stage loss-on-ignition measurements78 to estimate the organic and inorganic carbon fractions and found that the average inorganic carbon content was approximately 10% of the average organic carbon content (see Supplementary Note 1, Supplementary Fig. 5, Supplementary Fig. 6, and Supplementary Table 2 for more details). Therefore, we will refer to the carbon content (%C) measurements as organic carbon content (%OC). Note that the IRMS did not produce reliable nitrogen measurements with sediment core subsamples due to very low nitrogen content and instrument limitations.
To obtain estimates of sediment mass accumulation rates, one subsample from what remained of each sediment core slice was sent to the Virginia Institute of Marine Science Geochronology Lab for tracer analysis of 210Pb and 137Cs radioisotopes using gamma spectrometry (Canberra GL 2020 Low Energy Germanium detector). However, sediment mass accumulation rates could not be determined from the 210Pb and 137Cs activity profiles, potentially due to frequent mixing or coarse median sediment grain sizes. See the Supplementary Methods for details of the geochronology methods, with an example set of profiles in Supplementary Fig. 7.
Velocity data collection and processing
Tilt current meters (Lowell Instruments, Massachusetts) tethered to square concrete paving stones at six stations (S1V, S2V, S3V, S1B, S2B, S3B) measured the magnitude and direction of velocity at 16 Hz for 60 s bursts every 5 min. Velocity data were bisected based on the tilt direction into a flood (moving south toward the Annisquam River, denoted as positive) and ebb (moving north away from the Annisquam River, denoted as negative). The index of each tidal flow peak was found using the MATLAB findpeaks function. The velocities of the peaks during the flood and ebb were separately averaged and denoted as \(\overline{{U}_{{{{\rm{c}}}},{{{\rm{peak}}}}}}\). Both the seagrass shoots and the TCMs are deflected by currents, and at peak tidal velocity, the heights of the deflected shoots were lower than the heights of the deflected TCMs. In this condition, each TCM recorded an average of velocities within and above the meadow patch. As the velocities near the bed are most important for sediment transport, we used a two-layer velocity model to infer the velocity within the canopy, \({U}_{1}\), from measured velocity and meadow characteristics. The in-canopy velocity was used to estimate \(\overline{{U}_{1,{{{\rm{peak}}}}}}\) for the TCM sites located within meadow patches. The wave orbital velocity amplitude was calculated as \({U}_{w}=\sqrt{2}{U}_{{{{\rm{w}}}},{{{\rm{RMS}}}}}\), in which \({U}_{{{{\rm{w}}}},{{{\rm{RMS}}}}}\) is the average of the hourly peak root-mean-squared velocities within each tidal cycle. See the Supplementary Information of Lei et al.6 and Schaefer et al.79 for more details on estimating in-canopy current and wave orbital velocity with TCMs.
Image analysis
We used historical aerial and satellite images of the field site to estimate patch distribution at discrete points in time between 1972 and 2022. The image sources and dates are listed in Supplementary Table 3. Images were selected based on minimal cloud cover, optimal water depth and surface conditions, and spatial resolution. In addition, images were only drawn from months corresponding to the typical Z. marina L. growing season in Massachusetts, USA. Images, including the drone orthoimage, were geometrically transformed into alignment using the MATLAB Registration Estimator tool, cropped, and resampled to the same area of interest and pixel resolution. Vegetation patches were delineated using the MATLAB Image Segmenter tool. The MATLAB Image Processing Toolbox was used for further analysis of the delineated patches. Supplementary Fig. 8 shows a chronology of vegetated pixels across the images.
The images were used to estimate the total area of all vegetated patches within the meadow and to delineate the area of the whole meadow. In each image, the first and last pixels that were determined to be vegetated in each column were connected by lines to the first and last vegetated pixels of adjacent columns to create an outline of the entire meadow. The number of pixels contained within this outline defines the total meadow area \(A\). The number of pixels contained within each of the delineated patches was summed to define the area of vegetation, \({A}_{{{{\rm{v}}}}}\). See Supplementary Fig. 9 for an example of this method.
To quantify the persistence of vegetation through time, we counted the number of images across the collection of 21 images in which a given pixel was vegetated, denoted as \({N}_{21}\), and across the most recent ten images (2013-2022, with one image per year), denoted as \({N}_{{{{\rm{last}}}}10}\). The last 10 images were considered because the gaps in time between successive images were larger before 2013 (Supplementary Table 3). Finally, we considered the number of consecutive images in which a given pixel was vegetated stepping back from 2022, denoted as \({N}_{{{{\rm{lastcons}}}}}\). Each quantity was calculated at each pixel, and then an average of pixel values was made within a circle of 3-m diameter around the GPS waypoints, which conservatively accounted for the typical accuracy of the handheld GPS model that was used.
Statistical analyses
Statistical analyses on sediment trap and sediment core data were carried out using R packages80. Given the small sample sizes and spatial nature of the measurements, we used approximate Bayesian inference with integrated nested Laplace approximations (INLA81 version 24.2.9, www.r-inla.org). To account for spatial dependence among the measurement stations, we combined the INLA model with a stochastic partial differential equation approach (SPDE)82, with the default log-gamma priors for hyperparameters. For analyzing sediment core organic carbon stocks and sediment trap deposition rates, we tested the Gaussian, lognormal, and gamma likelihoods, selecting the model with the lowest Watanabe-Akaike Information Criterion. We used the beta likelihood for analyzing sediment trap percentage organic carbon content measurements (which were rescaled to range between 0 to 1). Point estimates and two-tailed 95% credible intervals were calculated for the predictor variables. Meshes based on longitude and latitude quantified the spatial distances between measurement stations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data for this study can be accessed at https://doi.org/10.6084/m9.figshare.28108040.v1.
Code availability
The open-source Domino software from Lowell Instruments was used to process raw tilt current meter data.
References
Unsworth, R. K. F. et al. Global challenges for seagrass conservation. Ambio 48, 801–815 (2019).
Fourqurean, J. W. et al. Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 5, 505–509 (2012).
Lafratta, A. et al. Challenges to select suitable habitats and demonstrate ‘additionality’ in Blue Carbon projects: a seagrass case study. Ocean Coast Manag. 197, 105295 (2020).
Lavery, P. S., Mateo, M. Á., Serrano, O. & Rozaimi, M. Variability in the carbon storage of seagrass habitats and its implications for global estimates of blue carbon ecosystem service. PLoS ONE 8, e73748 (2013).
Mazarrasa, I. et al. Factors determining seagrass blue carbon across bioregions and geomorphologies. Glob. Biogeochem. Cycles 35, 1–17 (2021).
Lei, J. et al. Spatial heterogeneity in sediment and carbon accretion rates within a seagrass meadow correlated with the hydrodynamic intensity. Sci. Total Environ. 854, 158685 (2023).
Ricart, A. M. et al. Variability of sedimentary organic carbon in patchy seagrass landscapes. Mar. Pollut. Bull. 100, 476–482 (2015).
Serrano, O. et al. Can mud (silt and clay) concentration be used to predict soil organic carbon content within seagrass ecosystems? Biogeosciences 13, 4915–4926 (2016).
Samper-Villarreal, J., Lovelock, C. E., Saunders, M. I., Roelfsema, C. & Mumby, P. J. Organic carbon in seagrass sediments is influenced by seagrass canopy complexity, turbidity, wave height, and water depth. Limnol. Oceanogr. 61, 938–952 (2016).
Kennedy, H. et al. Species traits and geomorphic setting as drivers of global soil carbon stocks in seagrass meadows. Glob. Biogeochem. Cycles 1–18 https://doi.org/10.1029/2022gb007481 (2022).
Asplund, M. E. et al. Dynamics and fate of blue carbon in a mangrove–seagrass seascape: influence of landscape configuration and land-use change. Landsc. Ecol. 36, 1489–1509 (2021).
Ricart, A. M., Pérez, M. & Romero, J. Landscape configuration modulates carbon storage in seagrass sediments. Estuar. Coast Shelf Sci. 185, 69–76 (2017).
Chen, Z., Jiang, C. & Nepf, H. Flow adjustment at the leading edge of a submerged aquatic canopy. Water Resour. Res. 49, 5537–5551 (2013).
Raupach, M. R., Finnigan, J. J. & Brunet, Y. Coherent eddies and turbulence in vegetation canopies: the mixing-layer analogy. Bound-Layer. Meteorol. 78, 351–382 (1996).
Infantes, E. et al. Effect of a seagrass (Posidonia oceanica) meadow on wave propagation. Mar. Ecol. Prog. Ser. 456, 63–72 (2012).
Kobayashi, N., Raichle, A. W. & Asano, T. Wave attenuation by vegetation. J. Water. Port. Coast Ocean Eng. 119, 30–48 (1993).
Jones, C. G., Lawton, J. H., Shachak, M. & Organisms, M. Organisms as ecosystem engineers. Oikos 69, 373–386 (1994).
Keil, R. G. & Hedges, J. I. Sorption of organic matter to mineral surfaces and the preservation of organic matter in coastal marine sediments. Chem. Geol. 107, 385–388 (1993).
Hemingway, J. D. et al. Mineral protection regulates long-term global preservation of natural organic carbon. Nature 570, 228–231 (2019).
Sogin, E. M. et al. Sugars dominate the seagrass rhizosphere. Nat. Ecol. Evol. 6, 866–877 (2022).
Dunic, J. C., Brown, C. J., Connolly, R. M., Turschwell, M. P. & Côté, I. M. Long‐term declines and recovery of meadow area across the world’s seagrass bioregions. Glob. Change Biol. 27, 4096–4109 (2021).
Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu Rev. Ecol. Evol. Syst. 34, 487–515 (2003).
Lindenmayer, D. B. & Fischer, J. Tackling the habitat fragmentation panchreston. Trends Ecol. Evol. 22, 127–132 (2007).
Warry, F. Y., Hindell, J. S., Macreadie, P. I., Jenkins, G. P. & Connolly, R. M. Integrating edge effects into studies of habitat fragmentation: a test using meiofauna in seagrass. Oecologia 159, 883–892 (2009).
Duarte, C. M., Fourqurean, J. W. & Olesen, B. In Seagrasses: Biology, Ecology and Conservation. 271–294 (Springer Dordrecht, 2006).
Kendrick, G. A. et al. The central role of dispersal in the maintenance and persistence of seagrass populations. BioScience 62, 56–65 (2012).
Fonseca, M. S., Zieman, J. C., Thayer, G. W. & Fisher, J. S. The role of current velocity in structuring eelgrass (Zostera marina L.) meadows. Estuar. Coast Shelf Sci. 17, 367–380 (1983).
Licci, S. et al. The role of patch size in ecosystem engineering capacity: a case study of aquatic vegetation. Aquat. Sci. 81, 1–11 (2019).
Dahl, M. et al. Increased current flow enhances the risk of organic carbon loss from Zostera marina sediments: Insights from a flume experiment. Limnol. Oceanogr. 63, 2793–2805 (2018).
Oreska, M. P. J., McGlathery, K. J. & Porter, J. H. Seagrass blue carbon spatial patterns at the meadow-scale. PLoS ONE 12, 1–18 (2017).
Bunnell, F. L. Let’s kill a panchreston: giving fragmentation meaning. In Forest Fragmentation – Wildlife and Management Implications (eds. Rochelle, J.A., Lehmann, L, & Wisniewski, J.) vii–xiii (Brill, Leiden, 1999).
Wu, J. Effects of changing scale on landscape pattern analysis: scaling relations. Landsc. Ecol. 19, 125–138 (2004).
Fonseca, M. S., Fisher, J. S., Zieman, J. C. & Thayer, G. W. Influence of the seagrass, Zostera marina L., on current flow. Estuar. Coast Shelf Sci. 15, 351–364 (1982).
Luhar, M., Rominger, J. & Nepf, H. Interaction between flow, transport and vegetation spatial structure. Environ. Fluid Mech. 8, 423–439 (2008).
Meysick, L., Infantes, E. & Boström, C. The influence of hydrodynamics and ecosystem engineers on eelgrass seed trapping. PLoS ONE 14, e0222020 (2019).
Van Wesenbeeck, B. K., Van De Koppel, J., Herman, P. M. J. & Bouma, T. J. Does scale-dependent feedback explain spatial complexity in salt-marsh ecosystems? Oikos 117, 152–159 (2008).
Arias-Ortiz, A. et al. Reviews and syntheses: 210Pb-derived sediment and carbon accumulation rates in vegetated coastal ecosystems—setting the record straight. Biogeosciences 15, 6791–6818 (2018).
Bouillon, S. & Boschker, H. T. S. Bacterial carbon sources in coastal sediments: a review based on stable isotope data of biomarkers. Biogeosciences 3, 175–185 (2006).
Abolfazli, E. & Strom, K. Salinity impacts on floc size and growth rate with and without natural organic matter. JGR Oceans 128, e2022JC019255 (2023).
Sutherland, B. R., Barrett, K. J. & Gingras, M. K. Clay settling in fresh and salt water. Environ. Fluid Mech. 15, 147–160 (2015).
Droppo, I. G. Structural controls on floc strength and transport. Can. J. Civ. Eng. 31, 569–578 (2004).
Julien, P. Y. Erosion and Sedimentation (Cambridge University Press, 2010).
Amos, C. L., Grant, J., Daborn, G. R. & Black, K. Sea Carousel—a benthic, annular flume. Estuar. Coast Shelf Sci. 34, 557–577 (1992).
McPhee, M. G. Turbulent stress at the ice/ocean interface and bottom surface hydraulic roughness during the SHEBA drift. J. Geophys. Res. 107, 8037 (2002).
Lowe, R. J., Koseff, J. R. & Monismith, S. G. Oscillatory flow through submerged canopies: 1. Velocity structure. J. Geophys. Res. Oceans 110, C10016 (2005).
Novak, A. B. et al. Factors influencing carbon stocks and accumulation rates in eelgrass meadows across New England, USA. Estuaries Coasts 43, 2076–2091 (2020).
Olesen, B., Krause-Jensen, D. & Christensen, P. B. Depth-related changes in reproductive strategy of a cold-temperate Zostera marina meadow. Estuaries Coasts 40, 553–563 (2017).
Orth, R. J. et al. Restoration of seagrass habitat leads to rapid recovery of coastal ecosystem services. Sci. Adv. 6, 1–9 (2020).
Johnson, A. J., Shields, E. C., Kendrick, G. A. & Orth, R. J. Recovery dynamics of the seagrass Zostera marina following mass mortalities from two extreme climatic events. Estuaries Coasts 44, 535–544 (2021).
Cunha, B. A. H., Duarte, C. M. & Krause-jensen, D. in European Seagrasses: an Introduction to Monitoring and Management. 72–76 (Monitoring and Managing of European Seagrasses Project, 2004).
Greve, T. M., Krause-Jensen, D., Rasmussen, M. B. & Christensen, P. B. Means of rapid eelgrass (Zostera marina L.) recolonisation in former dieback areas. Aquat. Bot. 82, 143–156 (2005).
Plus, M., Deslous-Paoli, J. M. & Dagault, F. Seagrass (Zostera marina L.) bed recolonisation after anoxia-induced full mortality. Aquat. Bot. 77, 121–134 (2003).
Oborny, B., Szabó, G. & Meszéna, G. in Scaling Biodiversity (eds. Storch, D., Marquet, P. & Brown, J.) 409–440 (Cambridge University Press, 2007).
Marro, J. & Dickman, R. Nonequilibrium Phase Transitions in Lattice Models (Cambridge University Press, 1999).
Fonseca, M. S. & Fisher, J. S. A comparison of canopy friction and sediment movement between four species of seagrass with reference to their ecology and restoration. Mar. Ecol. Prog. Ser. 29, 15–22 (1986).
Stauffer, D. & Aharony, A. Introduction to Percolation Theory (Taylor & Francis Inc, London, 1985).
Fonseca, M. S. & Bell, S. S. Influence of physical setting on seagrass landscapes. Mar. Ecol. Prog. Ser. 171, 109–121 (1998).
Duarte, C. M. Variance and the description of nature. In Comparative Ecology of Ecosystems: Patterns, Mechanisms and Theones. (eds. Cole, J., Lovett, G. & Findlay, S.) 301–318 (Springer-Verlag, New York, 1991).
Bormann, F., Herbert & Likens, Gene, E. Catastrophic disturbance and the steady state in northern hardwood forests: a new look at the role of disturbance in the development of forest ecosystems suggests important implications for land-use policies. Am. Scientist 67, 660–669 (1979).
Marbà, N. et al. Impact of seagrass loss and subsequent revegetation on carbon sequestration and stocks. J. Ecol. 103, 296–302 (2015).
Scheef, L. P. & Marcus, N. H. Mechanisms for copepod resting egg accumulation in seagrass sediments. Limnol. Oceanogr. 56, 363–370 (2011).
Berg, P. et al. Eddy correlation measurements of oxygen fluxes in permeable sediments exposed to varying current flow and light. Limnol. Oceanogr. 58, 1329–1343 (2013).
Norkko, J. & Shumway, S. E. in Shellfish Aquaculture and the Environment (ed. Shumway, S. E.) 297–317 (Wiley, 2011).
Volkenborn, N. et al. Intermittent bioirrigation and oxygen dynamics in permeable sediments: an experimental and modeling study of three tellinid bivalves. J. Mar. Res. 70, 794–823 (2012).
Brenchley, G. A. Mechanisms of spatial competition in marine soft-bottom communities. J. Exp. Mar. Biol. Ecol. 60, 17–33 (1982).
González‐Ortiz, V. et al. Submerged vegetation complexity modifies benthic infauna communities: the hidden role of the belowground system. Mar. Ecol. 37, 543–552 (2016).
Goshima, S. & Peterson, C. Both below- and aboveground shoalgrass structure influence whelk predation on hard clams. Mar. Ecol. Prog. Ser. 451, 75–92 (2012).
Pendleton, L. et al. Estimating global ‘blue carbon’ emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 7, e43542 (2012).
Moksnes, P. O. et al. Major impacts and societal costs of seagrass loss on sediment carbon and nitrogen stocks. Ecosphere 12, e03658 (2021).
Macreadie, P. I. et al. Losses and recovery of organic carbon from a seagrass ecosystem following disturbance. Proc. R. Soc. B Biol. Sci. 282 (2015).
Serrano, O., Arias-Ortiz, A., Duarte, C. M., Kendrick, G. A. & Lavery, P. S. Impact of marine heatwaves on seagrass ecosystems. 345–364 https://doi.org/10.1007/978-3-030-71330-0_13 (2021).
Howard, J. et al. Blue carbon pathways for climate mitigation: Known, emerging and unlikely. Mar. Policy 156, 105788 (2023).
Blomqvist, S. Sediment trapping—a subaquatic in situ experiment. Limnol. Oceanogr. 26, 585–590 (1981).
Arias-Ortiz, A. et al. Losses of soil organic carbon with deforestation in nangroves of Madagascar. Ecosystems 24, 1–19 (2021).
Gifford, R. M. & Roderick, M. L. Soil carbon stocks and bulk density: spatial or cumulative mass coordinates as a basis of expression? Glob. Change Biol. 9, 1507–1514 (2003).
Greiner, J. T., McGlathery, K. J., Gunnell, J. & McKee, B. A. Seagrass restoration enhances ‘blue carbon’ sequestration in coastal waters. PLoS ONE 8, 1–8 (2013).
Hassink, J. Decomposition rate constants of size and density fractions of soil organic matter. Soil Sci. Soc. Am. J. 59, 1631–1635 (1995).
Wang, Q., Li, Y. & Wang, Y. Optimizing the weight loss-on-ignition methodology to quantify organic and carbonate carbon of sediments from diverse sources. Environ. Monit. Assess. 174, 241–257 (2011).
Schaefer, R., Colarusso, P., Simpson, J. C., Novak, A. & Nepf, H. Proximity to inlet channel drives spatial variation in sediment carbon across a lagoonal seagrass meadow. Sci. Total Environ. 955, 177022 (2024).
R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing (2024).
Rue, H., Martino, S. & Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. Soc. Ser. B: Stat. Methodol. 71, 319–392 (2009).
Lindgren, F., Rue, H. & Lindström, J. An explicit link between Gaussian fields and Gaussian Markov random fields: the stochastic partial differential equation approach. J. R. Stat. Soc. Ser. B Stat. Methodol. 73, 423–498 (2011).
Acknowledgements
This study was supported by Shell International Exploration and Production through the MIT Energy Initiative. R. Schaefer was supported by the National Science Foundation Graduate Research Fellowship under Grant No. 2141064 and the Massachusetts Institute of Technology Department of Civil and Environmental Engineering. We thank the U.S. Environmental Protection Agency dive team for their assistance in the field: Eric Nelson, Jean Brochi, Danielle Gaito, Chuck Protzmann, and Brent England. The bathymetry and meadow mapping were collected by Mike Sacarny under award NA18OAR4170105 from the National Sea Grant College Program of the U.S. Department of Commerce’s National Oceanic and Atmospheric Administration. Mike Sacarny also provided boat support for divers. Drone aerial images were collected in July 2022 by a team led by pilot Sue Bickford of New England UAV, LLC. Gary Lei, Ian Koe, Chuyan Zhao, and Ernie Lee assisted with field data collection. Kimberly Candelario assisted with field data collection and laboratory sample processing. Advice on spatial statistics was provided by data science specialist Steven Worthington, at the Institute for Quantitative Social Science, Harvard University.
Author information
Authors and Affiliations
Contributions
Rachel Schaefer contributed to the conceptualization of the study, the data collection, data curation, formal analysis, as well as data visualization, prepared the original draft, and prepared revised drafts. Phil Colarusso contributed to the conceptualization of the study, the data collection, and data curation, as well as the writing and revision process. Juliet C. Simpson contributed to the conceptualization of the study, the data collection, data curation, as well as the writing and revision process. Alyssa Novak contributed to the conceptualization of the study, the data collection, and data curation, as well as the writing and revision process. Heidi Nepf contributed to the conceptualization of the study, the data collection, data curation, formal analysis, supervision, project administration, funding acquisition, as well as the writing and revision process.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Johannes Krause, Dorte Krause-Jensen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Nadine Schubert and Martina Grecequet. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Schaefer, R., Colarusso, P., Simpson, J.C. et al. Continual migration of patches within a Massachusetts seagrass meadow limits carbon accretion and storage. Commun Earth Environ 6, 129 (2025). https://doi.org/10.1038/s43247-025-02049-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02049-y
