
Abstract
The expansion and interconnection of protected areas are central to achieving the Kunming-Montreal Global Biodiversity Framework’s ambitious goals, yet their synergistic relationship remains underexplored. Here, we propose a framework integrating wildlife dispersal-based connectivity to address two key objectives in China: (1) constructing a cost-effective nature conservation network by combining connectivity and biodiversity prioritization, and (2) evaluating climate and anthropogenic risks while addressing habitat representation gaps. The framework aims to designate 30% of land as protected areas and informally allocate additional 30% of land as conservation priority corridors. Results show this strategy connects 57% of existing protected areas, protects 74% of priority zones, and achieves 89% of habitat representation targets. While current protected areas mitigate climate and anthropogenic threats, future expansion faces challenges due to geographic variations in these threats and the necessity for adequate representation. Our approach identifies and prioritizes these challenges, offering a data-driven pathway to achieve Kunming-Montreal targets.
Similar content being viewed by others

Introduction
The global biodiversity crisis, exacerbated by human activities and climate change, has triggered the implementation of integrated conservation strategies in recent decades1. While the establishment of protected areas (PAs) has emerged as an effective approach to mitigate and potentially reverse biodiversity loss2, the efficacy of existing PAs is often compromised by insufficient size, high fragmentation, and inadequate connectivity3. These limitations underscore the urgent need for a more holistic strategy in conservation planning and management. Expanding the boundaries of PAs is essential to safeguarding core habitats and ensuring long-term protection under strict legal and policy frameworks4. While PAs are formally established to conserve these core habitats, conservation priority corridors (CPCs) play a complementary role by enhancing connectivity across fragmented landscapes, facilitating species movement, and linking isolated PAs5. Unlike PAs, CPCs are not formally designated but serve as flexible, informal mechanisms that strengthen the overall functionality of the nature conservation networks. Both PA expansion and CPC designation require an integrated approach, considering factors such as functional connectivity, biodiversity importance, and cost-effectiveness. Together, these formal and informal protection strategies create a cohesive network that enhances biodiversity protection while balancing ecological, social, and economic priorities. This diversified approach ensures a more resilient and adaptive framework for nature conservation6.
International conservation efforts have set ambitious targets expanding global PA networks6. Currently, there are more than 200,000 PAs worldwide, covering about 16.6% of the global terrestrial land surface and 7.7% of the total marine area6. The Kunming-Montreal Global Biodiversity Framework (GBF) aims to increase PA coverage to 30% by 20307. Some researchers even advocate for a “Half-Earth” coverage target to effectively conserve biodiversity8. However, achieving these area-based conservation targets is challenging, particularly in regions like East Asia9, where expanding PAs may conflict with human welfare and ecosystem services10. Addressing this challenge, especially in light of the unfulfilled Aichi Target 1111, highlights the pressing need to establish and address spatial conservation priorities12. This approach, establishing and addressing spatial conservation priorities, aims to identify priority areas as flexible, informal pathways that balance biodiversity conservation with economic development, imposing fewer restrictions to human activities compared to formally PAs. However, research on the integration of priority areas as complementary extensions to legally defined PAs remains limited5. This gap underscores the urgent need for integrative approaches that reconcile competing land-use demands while optimizing biodiversity protection.
One of the biggest challenges in expanding PAs and designing CPCs is identifying suitable candidate areas13. This task is further complicated by the spatial mismatch between current PA distributions and biodiversity patterns14. However, a recurring limitation in related research is the frequent oversight of connectivity15, a vital component for maintaining ecological processes and species movement. This gap significantly hampers the effectiveness of existing PA networks in mitigating biodiversity loss16. Currently, two-thirds of critical global connectivity areas17 and key biodiversity areas (KBAs)18 remain unprotected, revealing significant spatial mismatches between these ecologically crucial zones. Therefore, it is imperative to expanding PA networks and designing CPCs that incorporate critical connectivity regions overlapping with other global conservation priorities, thereby achieving potential conservation synergies4,17. Such nature conservation networks would serve multiple functions: reconnecting habitat fragments, preventing population isolation, and maintaining ecological processes like migration under future climate scenarios19. By establishing CPCs as stepping stones beyond the static boundaries of PAs20, this strategy facilitates wildlife migration and gene flow. These CPCs play a critical role in mitigating the impacts of anthropogenic pressures, climate change, geographic isolation, and habitat loss21, thereby enhancing ecological resilience and promoting long-term biodiversity conservation.
Climate change and shifts in anthropogenic activity represent one of the most pressing challenges to contemporary conservation science16. These factors pose significant threats to species survival and undermine the effectiveness of PAs as well as broader conservation efforts. For example, climate change triggers changes in ecosystem dynamics, which can reduce habitat quality and increase species vulnerability22, thereby compromising the ability of PAs to fulfill their conservation objectives23,24,25. Moreover, while threats within PA boundaries may shift over time, unprotected areas face increasing pressures from human expansion. This leads to greater habitat fragmentation and loss of critical connectivity corridors1,21, impeding the movement of even highly mobile wildlife and limiting opportunities for interbreeding among populations in different PAs26,27. A recent study indicates that a 50% reduction in the human footprint could decrease national PA isolation by 28%17, benefiting the future migration of populations along climate gradients22. Traditional PA effectiveness evaluations, which often compare PAs with unprotected areas, can introduce statistical confounding due to site background biases28. To address this limitation, recent research has employed counterfactual approaches, comparing changes within PAs to unprotected areas to explore hypothetical scenarios without PA establishment16,29. However, the application of this method in robust evaluations for future PA network planning remains limited, a concerning gap given the incomplete fulfillment of previous Aichi targets11. The significant spatial variation further complicates this issue, as conservation interventions effective in one region may be inappropriate for others, even those in close proximity30. Consequently, there is an urgent need for robust assessments to inform future PA network expansions under the Kunming-Montreal GBF and the United Nations Sustainable Development Goals31.
To address these multifaceted challenges, this study introduces the Connectivity & Biodiversity Conservation (CBC) framework (Fig. 1), a systematic approach designed to optimize biodiversity conservation by integrating connectivity, biodiversity preservation, and cost-effectiveness. The CBC framework aims to enhance and expand PA networks through a scientifically grounded and practical methodology. Applied across China, this framework seeks to achieve two key objectives. First, it identifies a future nature conservation network by formally designating 30% of the national land as PAs and informally designating an additional 30% as CPCs. This is accomplished through a graph-based connectivity analysis combined with biodiversity prioritization overlays, ensuring a balanced strategy that maximizes ecological effectiveness while maintaining cost-efficiency. Second, the framework assesses the risks posed by climate change and anthropogenic activities in relation to habitat representation shortfalls. By evaluating the spatial distribution and overlap patterns of these risks and deficiencies, the analysis enables the prioritization of future PAs. This integrative approach not only advances the scientific understanding of complex ecosystem dynamics but also provides actionable tools for policymakers and conservation practitioners. By bridging the gap between theoretical conservation biology and applied landscape management, the CBC framework offers a pathway towards more effective, resilient, and sustainable biodiversity protection in the face of global environmental change.

a Framework Innovation: This framework focuses on creating habitat networks informed by mammal movement patterns, enabling the design of natural wildlife corridors. These corridors enhance ecological connectivity and resilience, fostering a more integrated and sustainable conservation approach. b Techniques and Methods: Advanced techniques, such as connectivity-based systematic conservation planning, form the foundation of this framework. It integrates risk and deficiency assessments to prioritize and evaluate conservation areas, ensuring adaptability and effectiveness under diverse ecological and human-driven pressures. c Aim of the Framework: The framework aims to establish a conservation network that combines static protected area boundaries with dynamic priority corridors. This integrated approach enhances biodiversity conservation by accommodating environmental changes and species movement, promoting flexibility and long-term ecological sustainability.
Methods
Data collection
This study utilized a comprehensive dataset of Chinese PA boundaries, compiled from governmental documents, published materials, scientific research reports. Marine PAs, as well as those located in Hong Kong, Macau, and Taiwan, which are all part of China, were excluded due to data limitations. Only national parks, nature reserves, and forest/wetland parks were classified as PAs, as data availability precluded the inclusion of secondary national parks. The final dataset comprised 4406 PAs, including 1030 with polygon boundary data and 3376 with point location data. For point-only PAs, a 5 km buffer zone was created to represent the PA boundary. PAs representing multiple protection types or geographically separated zones were merged, and overlapping areas were removed. Patches smaller than 10 km² were excluded. The resulting dataset included 2892 PA patches, covering 132,183 km² (14% of mainland China’s land area), projected using the GCS_Krasovsky_1940 coordinate system. It is worth noting that there is a discrepancy between our coverage and the 18% coverage published by the government (https://www.gov.cn/xinwen/2021-10/03/content_5640816.htm). This is partly a limitation of data availability. However, it is caused more by our selection of specific PA categories suitable for this connectivity assessments.
Identification of cost-effective connectivity corridors
Cost-effective connectivity corridors (CCCs) were identified using Graphab 2.632, a graph-based connectivity analysis software. The model of connectivity requires two critical inputs: the dispersal distance of wildlife and the resistance to wildlife moving across different regional surfaces.
Dispersal distance play a critical role in determining the corridors that connect adjacent PA patches, enabling wildlife movement within a predefined range. However, defining dispersal distance is inherently challenging due to the variability in movement abilities across species. To address this, we adopted a coarse-filter approach by establishing dispersal distance gradients designed to encompass the movement abilities of a wide range of terrestrial species. Based on previous studies33,34, we set typical dispersal distances at 10, 30, and 100 km, threshold that are known to cover the movement ranges of most terrestrial species. To ensure these gradients are applicability to China, we followed the methodology of Sun et al3., focusing specifically on terrestrial mammals. Using a list of important habitats for protected terrestrial wildlife species in China (https://www.forestry.gov.cn/search/538542). We identified 126 priority-protected mammals and estimated their movement abilities based on body weight, diet, and other ecological niche parameters through allometric relationships35,36. By aggregating these dispersal distances, we calculated the median, mean, and 90th percentile of movement ability across regions. The results indicate that the calculated values closely align with the predefined dispersal thresholds of 10 km, 30 km, and 100 km, confirming the applicability of these thresholds for studies conducted in China33,34,37.
Connectivity “resistance” was modeled using China’s human footprint dataset38, weighted by slope derived from a digital elevation model (DEM) dataset39 to account for the combined impact of human activities and steep topography on mammal movement. This resistance surface was used to identify least-cost paths (LCPs) between adjacent PAs by minimizing cumulative resistance. The “cost” associated with LCPs encompasses two dimensions: ecological cost and economic cost. Ecological cost reflects the hypothetical resistance species encounter when moving between PAs, serving as a proxy of CCCs in maintaining landscape connectivity. Economic cost represents the resources required to conserve CCCs, which escalate in areas with higher human activity. By prioritizing CCCs based on these dual dimensions, we enhance connectivity while balancing ecological and economic benefits, ensuring a cost-effectiveness approach to conservation planning.
To assess the priority of these connectivity corridors (referred as “corridor importance” hereafter, see Fig. 2), we defined by the number of overlapping CCCs within a given area. Unlike linear pathways, CCCs span broader regions characterized by a cost gradient (like a spreading watershed). The spatial overlap of CCCs across their coverage areas underscores their significance in enhancing connectivity. Specifically, we delineated CCCs using LCP analysis, which identifies corridors by minimizing cumulative resistance across the landscape. This approach enabled us to quantify corridor importance based on the spatial overlap and distribution of these CCCs within each region, providing a robust framework for prioritizing connectivity conservation efforts.

a–c Existing protected area (PA) networks and cost-effective connectivity corridors (CCCs) based on 10 km, 30 km, and 100 km wildlife dispersal distances. The darkness of green color gradient in the corridor priority area map indicates the number of potential corridors, with values normalized to a range of 0–1 to indicate the corridor importance. d The entire nature conservation networks in China. Dark green represents the 30% coverage target achieved by the current and candidate PAs. Light green represents the informally conservation priority corridors (CPCs). Together with the future PA network, they achieve the 50% coverage target.
The connectivity model in this study was designated to enhance the efficiency of the nature conservation network in conserving biodiversity, rather than focusing on linking similar habitat types. Specifically, the model aims to maintain biodiversity and ecological processes by improving connectivity across all suitable habitats, thereby ensuring a more robust and functional conservation network. Some species may require connectivity corridors spanning multiple habitat types to fulfill their lifecycle needs. Additionally, as climate change alters habitat types and species distributions, connecting different habitat types provides broader adaptation and migration spaces. Therefore, the model focuses on larger-scale connectivity planning. We acknowledge the limitations of the current model and suggest that future research should refine and improve the model by incorporating more detailed habitat data. This will better serve the design and planning of PA networks.
Identification of connectivity & biodiversity corridors
Connectivity & biodiversity corridors (CBCs) were derived by overlaying CCCs with conservation priority zones (CPZs). To identify CPZs, we employed the methodology of Yang et al9., which involved spatially integrating seven global biodiversity prioritization templates (BH, CE, CPD, EBA, G200, IFL, and KBA) and extracting the CPZs specific to the China region (Fig. S1). The biodiversity importance of each CPZ (referred to as “biodiversity importance” hereafter, see Fig. 2) was quantified based on the number of overlapping templates, ranging from 1 to 7. By overlaying these CPZs with CCCs, we obtained CBCs that integrate low cost, high corridor importance, and high biodiversity value, thereby optimizing conservation efforts across ecological and economic dimensions.
It is important to acknowledge that our method of overlaying conservation prioritization layers has limitations, particularly due to the varying resolutions and hierarchical differences among these layers (e.g., G200 Ecoregions vs. Key Biodiversity Areas [KBAs]). A more precise alternative would involve utilizing fine-scale prioritizations, such as KBAs, within a systematic conservation planning40 driven by habitat representation. However, this approach was not feasible in our study due to the challenges in integrating KBAs with our geographically widespread CCCs. Despite these constraints, the integration of multiple prioritization layers in our study provides a broader spatial perspective, which can guide actionable conservation planning.
Planning of candidate protected areas and conservation priority corridors
Candidate PAs were defined as CBCs identified under a 30 km dispersal distance scenario. These areas were selected for their high connectivity and biodiversity importance, aligning with the mean dispersal distance of mammals (see “Identification of cost-effective connectivity corridors”). In contrast, CBCs identified under a 100 km dispersal distance were designated as CPCs, which facilitate the dispersal of the majority (90th percentile) of mammals. While CPCs are not formally included in the PA network, they are recognized as priority areas that enhance overall connectivity. The distinction between candidate PAs and CPCs primarily lies in their planning objectives: candidate PAs are designated for inclusion in the PA network to meet the targets set by the Kunming-Montreal Global Biodiversity Framework (GBF), whereas CPCs are intended to support connectivity beyond the network, thereby contributing to landscape-scale connectivity.
Climate and anthropogenic activity changes
To measure human impact on PAs and beyond, we used a human footprint dataset with a spatial resolution of 1 km × 1 km for the period 2000–201838. Changes in human pressure (ΔHFP) were quantified for each region inside and outside PAs to measure long-term changes in anthropogenic activity (“anthropogenic activity change” hereafter). Historical monthly weather data for 1960–2018 from CRU-TS-4.0641 were downscaled to a spatial resolution of 2.5 min (~21 km² at the equator) to quantify past (1960–1969) and present (2010-2018) climate conditions42. Six climate indicators were calculated for each period following Wang et al30,43., using the biovars function in the R package ‘dismo’. Principal component analysis (PCA) reduced the dimensionality and collinearity of the climate indicators following Wang et al30. and Carroll et al44., with the first two PCA axes explaining 98% of the variance. To quantify climate change, we calculated the Euclidean distance between the past and present positions of each PA within a two-dimensional climate space, defined by PCA1 and PCA2 axes. This distance serves as a measure of the degree of climate shift, with larger distances indicating greater deviations from historical climate conditions and, consequently, higher levels of climate change. All datasets were normalized to a range of 0-1 using min-max normalization, with values close to one indicating high levels of change and values close to zero indicating low levels of change (Fig. S2).
Statistical analysis
Two metrics are used to measure the conservation coverage target: coverage based on the national terrestrial land surface (“area-based” hereafter) and coverage based on CPZs (“CPZs-based” hereafter). Area-based coverage means the percentage of the national terrestrial land surface covered by PAs or CPCs, and CPZs-based coverage represents the percentage of the total CPZs covered by PAs and CPCs.
To quantify the connectivity performance over PAs and beyond, we use two indicators, Probability of Connectivity (PC)45 and Protected Connected (ProtConn)46. Connectivity within networks was measured using the probability of connectivity (PC) index45. This index provides the probability that two points randomly placed in the study area are connected. It is the most widely used index to quantify the functional connectivity efficiency of ecological networks. The Protected Connected (ProtConn) indicator was used to define the percentage of terrestrial lands in China that are protected and connected46. PC and ProtConn are given by the following equation:
where ai and aj are the areas of the habitat patches i and j, and AL is the total studied area (both habitat and non-habitat patches). p∗ij is defined as the maximum product probability of all possible paths between patches i and j47.
To evaluate the habitat representation of the PAs and beyond, we used Mean Target Achievement (MTA)48. MTA measures the extent to which conservation targets for habitats are achieved. These metrics were calculated based on biomes and natural vegetation types (“ecoregions” hereafter) from the vegetation distribution map of China (http://westdc.westgis.ac.cn), offering a systematic assessment of the network’s habitat representation. Additionally, the representation values were determined through a two-step process. First, we created a representation map based on ecoregions (see Fig. S2). The representation value for each ecoregion was calculated using the combined coverage of existing and candidate PAs. Each point in the figure represents a candidate PA patch, formed by merging spatially adjacent and overlapping areas. Second, we used this ecoregion representation map to perform zonal statistics in ArcGIS, assigning a representation value to each candidate PA patch. This approach ensured that patches spanning multiple ecoregions received appropriate representation values.
To assess and compare climate and anthropogenic changes across current PAs and candidate areas for the 30% conservation target, we employed a statistically matched method. This approach minimizes non-random effects caused by location bias and accounted for contextual biases. For example, PAs are often established in steeper, more remote areas that are less affected by human activities, which can confound observed shifts in climate and human impacts. Using the R package “MatchIt,” we employed propensity score matching (PSM) to pair each pixel in current PAs with a similar pixel in candidate PAs or unprotected areas49. The matching process considered five covariates: (1) elevation; (2) slope; (3) topographic conditions (characterized by the topographic position index, TPI)39; (4) initial climate, human footprint condition; and (5) consensus land cover type (CLC)50 (Fig. S3-S5). For the first three covariates, matching was conducted without replacement using the nearest-neighbor method with a caliper of 0.25 standard deviations of the propensity score, while the last two covariates were matched exactly. To avoid spillover versus leakage effects from PAs29, neighboring areas outside each PA boundary were excluded from the matching analysis using QGIS software. All data were standardized to a 1 km resolution for consistency.
Results
Identification of 30% and 50% target connectivity & biodiversity corridors
Three CBCs scenarios identified 1549, 3356, and 6389 corridors, respectively. These corridors collectively covered 7.6%, 20.0%, and 47.8% of China’s terrestrial land surface (Table S1). Notably, as dispersal distance increased, the spatial extent of the CBCs expanded, with corridors identified under smaller dispersal distance scenarios nested within those of larger dispersal distance scenarios. Efforts to meet the coverage targets of the Kunming-Montreal Framework simultaneously, must consider the performance of CBCs in different scenarios, particularly in terms of coverage, representation, cost-effectiveness, and connectivity (Fig. 3; Table S2–S4). Integrating CBCs with a 30 km dispersal distance into PA networks (30% target candidate PAs) effectively achieved the 30% coverage targets, with an actual coverage of 34%. Similarly, CBCs with a 100 km dispersal distance were included as CPCs, serving as informal priority areas beyond the PA networks. When combined, future PA networks and CPCs exceeded the 50% coverage targets, achieving 61.8 % coverage. These formal and informal networks were collectively defined as nature conservation networks (Fig. 2d). The integration of these networks significantly reduced network isolation, decreasing from 914 dispersed habitat components under the current PA networks to just 108 under nature conservation networks (Fig. S6). This reduction is critical because wildlife within isolated components cannot successfully traverse or communicate with regions outside their immediate habitat. Regional disparities in the causes of isolation were evident. In the western part of the Hu-line, a socioeconomic divide (Fig. S7)51, isolation was primarily driven by the absence of high biodiversity importance habitats, such as CPZs, which act as essential stepping stones. In contrast, isolation in the eastern of the Hu-line was mainly due to the human footprint impacts.

a Bivariate plot of corridor importance and biodiversity importance for candidate protected areas. b The proportion of corridor and biodiversity importance regions for each biome in China. Abbreviations: Trop./Sub. Moist Br. For., Tropical and Subtropical Moist Broadleaf Forests; Temp. Br./Mix. For., Temperate Broadleaf and Mixed Forests; Temp. Con. For, Temperate Coniferous Forests; Temp. G./Sav/Sh., Temperate Grasslands, Savannas, and Shrublands; Flooded G./Sav., Flooded Grasslands and Savannas; Montane G./Sh., Montane Grasslands and Shrublands; Deserts/X. Sh., Deserts and Xeric Shrublands.
A bivariate choropleth map revealed spatial relationships between corridor importance and biodiversity importance for candidate PAs (Fig. 3a). Candidate PAs located in tropical and subtropical moist broadleaf forests and temperate coniferous forests exhibited the highest biodiversity importance. Conversely, candidate PAs in montane grasslands and shrublands, deserts and xeric shrublands, and northern temperate broadleaf and mixed forests were most valuable for enhancing connectivity. Notably, temperate broadleaf and mixed forests, particularly in the North China Plain, showed both low levels of biodiversity and corridor importance (Fig. 3b).
Contribution of nature conservation networks
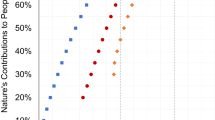
Our proposed nature conservation network performs well in achieving area-based conservation coverage targets. By integrating formal PAs and informal CPCs, the network significantly expanded the spatial coverage compared to the current PA networks, increasing it from 14.0% to 34.0%. Additionally, it designated an extra 27.8% of the landscape as informal CPCs pathways (Fig. 4; Table S1). This integration also enhanced the coverage of CPZs, with PA-based CPZ coverage rising from 14.8% to 39.6%. When combined with informal CPCs, CPZ coverage reached 73.8% (Fig. 4; Table S2). However, notable gaps persisted, particularly in deserts and xeric shrublands, where only 23.3% of existing PAs overlapped with CPZs, significantly lower than the average of 85.9% observed in other biomes (Tables S2). Even with the inclusion of informal CPCs, CPZs coverage in these biomes reaches only 83.5%, which is 13.4% below the national average. While the top ten largest PAs for the desert and grassland-type biomes account for 52% of the total PA coverage (Fig. S8), most are in wilderness regions with minimal CPZs overlap.

Dark green represents the 30% coverage target achieved by the current and candidate protected areas (PAs). Light green represents the informally conservation priority corridors. Together with the future PA network, they achieve the 50% coverage target. For the PC metric (*): To maintain consistency in percentage representation, the PC values are multiplied by 100. For the Mean Target Achievement (MTA) metric (**): The MTA metric is calculated based on distinct target representation levels. The separately developed target representation levels are displayed within a single bar chart to ensure consistency in visualization.
The integration of new PAs and informal CPCs dramatically improved connectivity within PA networks. Connectivity within PA networks increased five-fold with addition of new PAs, while the connectivity of nature conservation network was 53 times higher than that of the current PA networks due to the synergies provided by informal CPCs. The proportion of effectively connected habitats increased from 7.3% to 16.2% within PA networks and further to 57.1% within the nature conservation network (Fig. 4; Table S1). Well-connected patterns were observed in regions such as the Sanjiang Plain, Qinghai-Tibet Plateau, and Yangtze River Basin, whereas connectivity remained limited in the North China Plain and Yunnan-Guizhou Plateau (Fig. S6). Finally, the network contributed significantly to habitat representation across biomes and ecoregions. At the biome scale, the contribution to biome representation increased from 38.9% to 92.6% based on the 30% coverage target for PA networks. It further increased to 95.4% when incorporating informal CPCs based on the 50% coverage target (Fig. 4; Table S3). Similarly, at the ecoregion scale, the contribution to ecoregions rose from 47.7% to 89% based on the 30% coverage target, and reached 94.9% when informal CPCs are included based on the 50% coverage target (Fig. 4; Table S4).
Risks and habitat representation in future protected areas
A ternary map was employed to visualize the spatial distribution of climate changes, anthropogenic activity changes, and habitat representation in China (Fig. 5a). The analysis revealed distinct regional patterns: areas around the Hu Line and in northeast China were predominantly characterized by high anthropogenic activity change, coupled with limited climate change and relatively high habitat representation. In contrast, Southeastern China, primarily in tropical and subtropical moist broadleaf forests, exhibited high climate change and low habitat representation, despite experiencing relatively low levels of anthropogenic activity change. Candidate PAs in northwest China, particularly those in the deserts and xeric shrublands, faced the dual pressures of high climate and low habitat representation, alongside anthropogenic activity changes. Notably, only near Northeast China Tiger and Leopard National Park and parts of the Tibetan Plateau were less exposed to both climate and anthropogenic activity changes, while also maintaining high habitat representation. To assess the differential impacts of climate and anthropogenic activity changes, we applied a counterfactual matching approach, creating site-level matched pairs to compare current PAs with candidate PAs and other unprotected areas (see Methods). The results showed that established PAs experienced lower levels of climate change and anthropogenic activities compared to unprotected areas (Fig. 6b, d). This revealed that candidate PAs were subject to greater changes in climate anthropogenic activities than current PAs (Fig. 6a, c).

a Ternary plot showing the intersection of climate, anthropogenic activity risks and habitat deficiencies regions. Each point in the figure represents a candidate protected area patch formed by merging spatially adjacent and overlapping regions. In total, there are 689 points included in the representation. The plus sign (+) in the ternary plot legend shows the intersection of the means of the potential risk and deficiency types. b Distribution of the potential risks and deficiencies faced by candidate protected areas for each biome in China. Abbreviations: Trop./Sub. Moist Br. For., Tropical and Subtropical Moist Broadleaf Forests; Temp. Br./Mix. For., Temperate Broadleaf and Mixed Forests; Temp. Con. For, Temperate Coniferous Forests; Temp. G./Sav/Sh., Temperate Grasslands, Savannas, and Shrublands; Flooded G./Sav., Flooded Grasslands and Savannas; Montane G./Sh., Montane Grasslands and Shrublands; Deserts/X. Sh., Deserts and Xeric Shrublands.

Violin plots showing (a) climate and (c) anthropogenic activity changes for matched established protected areas (PAs) and unprotected areas (excluding a 1-fold PA buffer), as well as (b) climate and (d) anthropogenic activity changes for matched established and candidate PAs. Points and lines show means and standard deviations, respectively. P-values were calculated using Mann-Whitney U tests. \({\bar{x}}\) represents the percent difference in mean changes.
Discussion
Establishing nature conservation networks is crucial for advancing the post-2020 biodiversity agenda
Effective conservation strategies in the 21st century must transcend traditional area-based approaches to PA establishment. Although integrating multiple conservation priorities presents challenges in meeting the GBF targets52. It is imperative to prioritize biodiversity important zones, prioritize well-connected, expansive habitat networks, and balance the trade-offs between conservation and economic development. It is also necessary to consider habitat representation and exposure to climate and anthropogenic activity changes. Our study indicates that existing terrestrial PAs in China are hindered by limited connectivity and insufficient protection of CPZs. There is a significant spatial mismatch between corridor importance areas and biodiversity importance areas, emphasizing the need for region-specific conservation strategies. In northwestern China, characterized by high altitudes and arid climates53, intact habitats exhibit high connectivity54 but relatively low biodiversity importance, particularly in terms of species richness and endemism4. This suggests that conservation efforts in this region should prioritize maintaining and preventing the degradation of existing corridors55, which are crucial for sustaining ecological processes in these fragile ecosystems. In contrast, southeastern and southwestern China, with their warm climate, diverse habitats, and exceptional biodiversity significance in terms of high species richness and the presence of threatened species4,56, face escalating pressures from land-use intensification and urbanization56. Here conservation strategies must focus on protecting biodiversity important areas while simultaneously enhancing corridor importance to improve ecosystem function and resilience.
Our proposed CBC framework addresses this need by integrating PA networks and informal CPCs. This framework considers the balance between human development and biodiversity conservation, adding a critical connectivity perspective to determine cost-effective priorities57. Unlike previous approaches that have primarily focused on carbon sequestration or ecological restoration, our framework explicitly emphasizes the importance of habitat connectivity for the long-term persistence of biodiversity13,58. Our findings reveal that nature conservation networks strongly align with areas designated for ecological restoration, particularly in southeast China58. However, there remains a significant mismatch with high-priority areas for carbon sequestration59, suggesting that focusing solely on carbon sequestration could potentially reduce global habitat connectivity, thereby limiting species dispersal under climate change. Additionally, practical considerations such as corridor width are crucial, as long but narrow corridors may not facilitate adequate dispersal among PAs. Wider corridors are advantageous in mitigating excessive edge effects5.
The potential risks and deficiencies of future PA networks
Our proposed CBC framework underscores the importance of considering the climate and anthropogenic activity change risks and habitat representation deficiencies inherent in PA networks, introducing a crucial element for future PA design. Regions may be exposed to singular or multiple threats, such as climate change, deforestation, and land-use change. The CBC framework offers significant advantages for mitigating these risks and deficiencies by mapping and ranking them, thereby identifying regions that require the most protection and the most pressing issues to address. Our analysis indicates that candidate PAs in southeastern China (mainly in tropical and subtropical moist broadleaf forests) exhibit the highest climate changes, suggesting an increased risk of extinction in these areas. Conversely, the regions around the Hu Line and in the northeastern China shows the highest anthropogenic activity changes, indicating habitats are at a greater risk of quality degradation and fragmentation, which could result in corridors connecting “empty” habitats. In addition, almost all tropical and subtropical moist broadleaf forests and nearly half of deserts and xeric shrublands face habitat representation deficits.
Our study also contributes to recent advancements in employing robust counterfactual approaches for evaluating climate and anthropogenic activity changes across existing PAs, candidate PAs, and unprotected areas. These counterfactual approaches should be implemented in the design of new PAs15,30. Our findings further demonstrate that existing PAs are exposed to anthropogenic disturbances, driven by unpredictable fluctuations in human activity intensity over time. However, these disturbances are significantly lower in PAs compared to candidate PAs and unprotected areas, underscoring their effectiveness in mitigating human impacts. By analyzing the 2000–2018 human footprint dataset, our study provides a comprehensive assessment of conservation outcomes in the 21st century, highlighting the critical role of current PAs in managing anthropogenic pressures. These results not only emphasize the success of existing PAs in reducing disturbances but also underscore the urgent need to address anthropogenic impacts as a central challenge in conservation planning. As the global network of PAs expands, effectively managing these impacts will be essential to ensuring the long-term resilience and functionality of protected ecosystems. Additionally, our analysis shows that candidate PAs have greater climate changes than existing PAs. This pattern may reflect a selection bias in the historical designation of existing PAs. These areas were often established in regions with high biodiversity, ecosystem integrity, and long-term sustainability, which are more likely to occur in climatically stable environments. However, this climate change risk should be considered in context23. Integrating these regions into PA networks could mitigate the impacts of climate change by enhancing connectivity, thereby facilitating species migration and adaptation19.
Conservation implications
Our framework emphasizes prioritizing zones that offer low cost, high biodiversity importance, and strong connectivity, revealing distribution patterns that align with climate and anthropogenic activity change versus habitat representation. This approach provides several significant advantages and management implications for enhancing PAs and habitat networks in China. First, it addresses the deficiencies of current PA networks and identifies future PAs. For example, there is an urgent need to improve PA distribution in biodiversity importance regions, such as those in southeastern China, which are currently unprotected (8.1% coverage compared to the 14.8% national average) and often consist of numerous small PAs. These relatively small but crucial regions are frequently overlooked in conservation planning60. Furthermore, our analysis indicates that temperate broadleaf and mixed forests are subject to high levels of human disturbance. In these regions, PAs are severely geographically isolated, and few candidate PAs were identified due to extensive anthropogenic modifications. Additionally, the percentage coverage gains are low for PAs in desert and grassland biomes because existing PAs are large and well-isolated from each other. Therefore, focusing solely on large wilderness areas to achieve high PA coverage may not enhance biodiversity conservation effectiveness at the national level. A more effective strategy would involve utilizing small-sized PAs or other conservation-friendly land uses to improve connectivity within the PA network.
Second, our framework enables the evaluation of potential risks and deficiencies within nature conservation networks, which is crucial for allocating conservation priorities. For instance, regions exposure to climate change largely overlap with critical priority zones in tropical and subtropical moist broadleaf forests. Facilitating species dispersal through corridors and migration in response to climate change is essential for protecting wildlife populations. Our study also shows that the future nature conservation networks identified by our framework achieves better coverage of representative habitats. Most biomes and ecoregions perform well in meeting 30% of the representative target. Almost all biomes and ecoregions can achieve 50% coverage protection objectives through the synergy of CPCs.
Third, given the observed variation among and within regions, it is important to consider certain social and environmental issues at finer scales to effectively address policy trade-offs13,61. For example, regions with high biodiversity importance often experience significant human-land conflicts, making PA network expansion potentially combative with human development. Previous studies have shown that implementing stricter and more efficient management in these regions with limited coverage can offset the constraints of smaller PA sizes62,63. Additionally, the United Nations Decade on Ecosystem Restoration aims to safeguard and restore one million hectares of degraded land by 203064. This goal will benefit regions like grasslands, which experience slower climate change rates, are less impacted by human activity, and provide important ecosystem services65,66. Furthermore, the Northeastern China Plain and Northern China Plain are critical regions for intensified agricultural land aimed at safeguarding food security66,67. However, the limitations of smaller PAs, such as reduced habitat connectivity and limited protection of threatened species, should be acknowledged68. Formal protection may be contested due to livelihood concerns or food supply needs in these areas. Nonetheless, there remains an opportunity to establish informal conservation priority corridors, even in the most constrained scenarios outlined in our framework. In particular, habitat connectivity can be maintained and enhanced through trade-offs and synergistic conservation-friendly land uses, such as agroforestry, livestock grazing in forests, and other agroecological management practices69.
Data availability
All data used for our analyses are publicly available. Digital elevation model (DEM) data are available from https://doi.org/10.1594/PANGAEA.867115. Terrestrial Human Footprint data are available from https://doi.org/10.6084/m9.figshare.16571064.v7. Historical monthly weather data for 1960–2018 from CRU-TS-4.06 (https://crudata.uea.ac.uk/cru/data/hrg). The full consensus land-cover (CLC) product is available at http://www.earthenv.org. The data sources for seven global biodiversity prioritizations (BH, CE, CPD, EBA, G200, IFL, and KBA) can be found in the references provided in the supplementary materials. The main data analyzed are the connectivity model outputs, for the data used to plot the main graphs used in this study can be accessed at: https://doi.org/10.6084/m9.figshare.25932532.v2.
Code availability
All statistical analyses were conducted using Graphab (version 2.6), ArcMap (version 10.2), ArcGIS pro (version 2.6), QGIS (version 3.20), and R (version 4.2.2). For our analyses, we used the following R packages: dismo (version 1.3–16), ConsTarget (developed by Jantke et al., 2018), and MatchIt (version 4.7.1). The parameters for building the connectivity model in Graphab were guided by the user manual. Our data analyses in R primarily relied on standard package functions, with all specific parameters thoroughly detailed in the Methods section. Additionally, custom codes used in this study have been uploaded along with source data for transparency and reproducibility.
References
Jones, K. R. et al. One-third of global protected land is under intense human pressure. Science 360, 788–791 (2018).
Terraube, J., Van Doninck, J., Helle, P. & Cabeza, M. Assessing the effectiveness of a national protected area network for carnivore conservation. Nat. Commun. 11, 2957 (2020).
Sun, W. C., Zhao, Y. J., Chen, W.H. & Bai, Y.F. Current national nature reserves are insufficient to safeguard the long-term survival of birds and mammals in China. Commun. Earth Environ. 5, 304 (2024).
Xu, W. et al. Strengthening protected areas for biodiversity and ecosystem services in China. PNAS 114, 1601–1606 (2017).
Feeley, K. J. & Rehm, E.M. Priorities for conservation corridors. Nat. Clim. Change 4, 405–406 (2014).
UNEP-WCMC, I., NGS. Protected Planet Live Report 2020.
Convention on Biological Diversity. Kunming-Montreal Global Biodiversity Framework. (Montreal, Montreal, 2022).
Wilson, E. O. Half-Earth. (Liveright, 2017).
Yang, R., et al. Cost-effective priorities for the expansion of global terrestrial protected areas: Setting post-2020 global and national targets. Sci. Adv.6, eabc3436 (2020).
Hoang, N. T., et al. Mapping potential conflicts between global agriculture and terrestrial conservation. PNAS 120, e2208376120 (2023).
Convention on Biological Diversity. Strategic plan for biodiversity 2011-2020, including Aichi biodiversity targets. (Montreal, 2010).
Brooks, TM. et al. Global biodiversity conservation priorities. Science 313, 58–61 (2006).
Shen, X. et al. Countries’ differentiated responsibilities to fulfill area-based conservation targets of the Kunming-Montreal Global Biodiversity Framework. One Earth 6, 548–559 (2023).
Wilson, K. A. et al. Prioritizing conservation investments for mammal species globally. Philos. T R. Soc. B 366, 2670–2680 (2011).
Rodrigues, A. S. L. & Cazalis, V. The multifaceted challenge of evaluating protected area effectiveness. Nat. Commun. 11, 5147 (2020).
Santangeli, A., et al. Mixed effects of a national protected area network on terrestrial and freshwater biodiversity. Nat. Commun. 14, 5426 (2023).
Brennan, A. et al. Functional connectivity of the world’s protected areas. Science 376, 1101–1104 (2022).
KBA Partnership. KBA Programme Annual Report 2021.
McGuire, JL., Lawler, JJ., McRae, BH., Nunez, TA. & Theobald, DM. Achieving climate connectivity in a fragmented landscape. PNAS 113, 7195–7200 (2016).
Parks, S.A., Holsinger, L.M., Abatzoglou, J.T., Littlefield, C.E. & Zeller, K.A. Protected areas not likely to serve as steppingstones for species undergoing climate-induced range shifts. Glob. Chang Biol. 29, 2681–2696 (2023).
Haddad, N. M., et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, e1500052 (2015).
Gottfried, M. et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Change 2, 111–115 (2012).
Asamoah, E, Beaumont, L & Maina, J Climate and land-use changes reduce the benefits of terrestrial protected areas. Nat. Clim. Change 11, 1105–1110 (2021).
Dobrowski, S. Z. et al. Protected-area targets could be undermined by climate change-driven shifts in ecoregions and biomes. Commun. Earth Environ. 2, 198 (2021).
Williams, D. R., Rondinini, C. & Tilman, D. Global protected areas seem insufficient to safeguard half of the world’s mammals from human-induced extinction. PNAS 119, e2200118119 (2022).
Pacifici, M., Di Marco, M. & Watson, J. E. M. Protected areas are now the last strongholds for many imperiled mammal species. Conserv. Lett. 13, e12748 (2020).
Anderson, E. & Mammides, C. The role of protected areas in mitigating human impact in the world’s last wilderness areas. Ambio 49, 434–441 (2020).
Meng, J., et al. Anthropogenic vulnerability assessment of global terrestrial protected areas with a new framework. Biol. Conserv.283, 110064 (2023).
Brodie, J. F. et al. Landscape-scale benefits of protected areas for tropical biodiversity. Nature 620, 807–812 (2023).
Shrestha, N., Xu, XT., Meng, JH. & Wang, Z. H. Vulnerabilities of protected lands in the face of climate and human footprint changes. Nat. Commun.12, 1632 (2021)
Zeng, Y., Koh, L. P. & Wilcove, D. S. Gains in biodiversity conservation and ecosystem services from the expansion of the planet’s protected areas. Sci. Adv. 8, eabl9885 (2022).
Savary, P. et al. graph4lg: A package for constructing and analysing graphs for landscape genetics in R. Methods Ecol. Evol. 12, 539–547 (2020).
Saura, S., Bastin, L., Battistella, L., Mandrici, A. & Dubois, G. Protected areas in the world’s ecoregions: How well connected are they?. Ecol. Indic. 76, 144–158 (2017).
Palfrey, R., Oldekop, J. A. & Holmes, G. Privately protected areas increase global protected area coverage and connectivity. Nat. Ecol. Evol. 6, 730–737 (2022).
Bowman, J. Is dispersal distance of birds proportional to territory size?. Can. J. Zool. 81, 195–202 (2003).
Santini, L. et al. Ecological correlates of dispersal distance in terrestrial mammals. Hystrix 24, 181–186 (2013).
Tucker, M. A. et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 359, 466–469 (2018).
Mu, H., et al. A global record of annual terrestrial Human Footprint dataset from 2000 to 2018. Sci. Data 9, 176 (2022).
Amatulli, G., et al. A suite of global, cross-scale topographic variables for environmental and biodiversity modeling. Sci. Data 5, 180040 (2018).
Margules, C. R. & Pressey, R. L. Systematic conservation planning. Nature 405, 243–253 (2000).
Harris, I., Osborn, TJ., Jones, P. & Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 7, 109 (2020).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J. Climatol. 37, 4302–4315 (2017).
Robert J. Hijmans, S. P., Leathwick, J. and Elith, J.. dismo: Species Distribution Modeling. (2022).
Carroll, C., Parks, S.A., Dobrowski, S.Z. & Roberts, D.R. Climatic, topographic, and anthropogenic factors determine connectivity between current and future climate analogs in North America. Glob. Chang Biol. 24, 5318–5331 (2018).
Saura, S. & Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan 83, 91–103 (2007).
Saura, S. et al. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 219, 53–67 (2018).
Foltête, J. C., Clauzel, C. & Vuidel, G. A software tool dedicated to the modelling of landscape networks. Environ. Model Softw. 38, 316–327 (2012).
Jantke, K., Kuempel, C. D., McGowan, J., Chauvenet, A.L.M. & Possingham, HP. Metrics for evaluating representation target achievement in protected area networks. Divers Distrib. 25, 170–175 (2019).
Meng, Z. et al. Post-2020 biodiversity framework challenged by cropland expansion in protected areas. Nat. Sustain. 6, 758–768 (2023).
Tuanmu, M. N. & Jetz, W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Glob. Ecol. Biogeogr. 23, 1031–1045 (2014).
Li, J., Lu, D., Xu, C., Li, Y. & Chen, M. Spatial heterogeneity and its changes of population on the two sides of Hu Line. Acta Geogr. Sin. 72, 148–160 (2017).
Gray, C. L., et al. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 7, 12306 (2016).
Wei, K. & Wang, L. Reexamination of the Aridity Conditions in Arid Northwestern China for the Last Decade. J. Clim. 26, 9594–9602 (2013).
Zhao, G., Liu, J., Kuang, W., Ouyang, Z. & Xie, Z. Disturbance impacts of land use change on biodiversity conservation priority areas across China: 1990-2010. J. Geographical Sci. 25, 515–529 (2015).
Li, B. V., Wu, S., Hua, F. & Mi, X. The past and future of ecosystem restoration in China. Curr. Biol. 34, R379–R387 (2024).
Dong, X., et al. Importance of including Key Biodiversity Areas in China’s conservation area-based network. Biol. Conserv. 296, 110676 (2024).
Pouzols, F. M. et al. Global protected area expansion is compromised by projected land-use and parochialism. Nature 516, 383–386 (2014).
Strassburg, B. B. N. et al. Global priority areas for ecosystem restoration. Nature 586, 724–729 (2020).
Ward, M., et al. Just ten percent of the global terrestrial protected area network is structurally connected via intact land. Nat. Commun. 11, 4563 (2020).
Pimm, S. L., Jenkins, CN. & Li, BV. How to protect half of Earth to ensure it protects sufficient biodiversity. Sci. Adv. 4, eaat2616 (2018).
Wu B. F., Qian J. K. & Y, Z. Land cover atlas of the people’s republic of china (1:1,000,000). (Beijing: SinoMaps Press, 2017).
Wauchope, H. S. et al. Protected areas have a mixed impact on waterbirds, but management helps. Nature 605, 103–107 (2022).
Françoso, R. D. et al. Habitat loss and the effectiveness of protected areas in the Cerrado Biodiversity Hotspot. Nat. Conservacao. 13, 35–40 (2015).
UNEP, F. The UN Decade on Ecosystem Restoration 2021-2030. (The United Nations Environment Programme (UNEP), The Food and Agriculture Organization (FAO), 2020).
Watson, J. E. M. et al. Persistent disparities between recent rates of habitat conversion and protection and implications for future global conservation targets. Conserv. Lett. 9, 413–421 (2016).
Bardgett, R. D. et al. Combatting global grassland degradation. Nat. Rev. Earth Env. 2, 720–735 (2021).
Duan, J. et al. Consolidation of agricultural land can contribute to agricultural sustainability in China. Nat. Food 3, 179 (2022).
Ricciardi, V., Mehrabi, Z., Wittman, H., James, D. & Ramankutty, N. Higher yields and more biodiversity on smaller farms. Nat. Sustain. 4, 651–657 (2021).
Sietz, D., Klimek, S. & Dauber, J. Tailored pathways toward revived farmland biodiversity can inspire agroecological action and policy to transform agriculture. Commun. Earth Environ. 3, 211 (2022).
Acknowledgements
This study was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (Grant Nos. XDA23080000).
Author information
Authors and Affiliations
Contributions
Y.B. conceived the study. W.S., E.Z., Y.Z., Z.W., W.C., Y.W., and Y.B. conducted data analysis and drafted the manuscript. All authors contributed to the revision of the manuscript, and Y.B. edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Diogo Alagador and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Heike Langenberg and Aliénor Lavergne. A peer review file is available
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sun, W., Zhang, E., Zhao, Y. et al. Conservation priority corridors enhance the effectiveness of protected area networks in China. Commun Earth Environ 6, 275 (2025). https://doi.org/10.1038/s43247-025-02227-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-025-02227-y
