
Abstract
Ensuring habitat representativeness is fundamental to area-based conservation, yet the effectiveness and efficiency of individual protected areas in meeting this goal remain underexamined, potentially impeding targeted strategies. Here we assessed habitat representativeness, representativeness contribution and representativeness density across China’s protected areas to evaluate their performance in meeting species conservation needs, using refined species distribution data. The overall network’s habitat representativeness was 62.39%, fully meeting conservation needs for 28.33% of species. Among them, 175 protected areas contributed over half of existing achievement for 751 species, including 148 fully dependent on one site, revealing uneven conservation efforts and species-site dependencies. While protected area size correlated with habitat representativeness and contribution, species richness also correlated with contribution and density, showing size alone may not determine efficiency. We then classified protected areas into four efficiency-based categories and proposed tailored strategies. Our metric-driven assessment provides a systematic approach to identifying gaps and optimizing conservation priorities at site level across the country.
Similar content being viewed by others

Introduction
To prevent ongoing global biodiversity loss, the Kunming-Montreal Global Biodiversity Framework (KMGBF) emphasizes the importance of establishing ecologically representative protected areas (PAs) for effective conservation, alongside ensuring efficient resource use1. Central to the KMGBF is the 30 × 30 target, aiming to protect at least 30% of Earth’s representative areas by 2030. Other concepts such as half earth2 and cost-effective zones3 also advocate for protecting more global surfaces. To further support conservation implementation, KMGBF also aims to mobilize at least $200 billion annually by 2030, highlighting the need for effective, efficient, and transparent resource allocation. Achieving these outcomes, therefore, requires every PA to not only effectively support ecological representativeness but also to optimize resource use and contribute to conservation effects efficiently.
Effectiveness and efficiency serve distinct but complementary roles. Effectiveness refers to doing the right things4, measuring the absolute level of outcome5,6, and reflecting the ability to achieve conservation targets7,8. A key component of effectiveness mentioned in KMGBF is the representativeness of PAs, defined as the extent to which PAs capture the full variety of biodiversity9 and support the long-term persistence of species8. Early assessment of representativeness often considered a species protected if any portion of its habitat intersected with a PA. Later studies, grounded in theories such as minimum viable population10 and minimum area requirements11, developed various approaches to evaluate how effectively PAs meet species-specific conservation needs8,9,12,13. Such conservation need-based approaches have been applied at global14,15,16,17, continental18, and national scales (e.g., in Italy19,20, Costa Rica21, and Australia22).
Efficiency, on the other hand, refers to doing things right4 and generally measures the output-input ratio5,6, reflecting the ability to achieve the existing conservation effects7. In biodiversity conservation, efficiency has frequently been measured using area or cost as proxies for input, allowing for evaluations of cost-effectiveness and cost-efficiency of both current and potential PAs17,23,24. However, efficiency in conservation involves not only minimizing input but achieving conservation goals within existing constraints9. It requires considering maximizing the relative contribution value of each conservation unit within the broader conservation network, thereby enabling better resource allocation to improve the performance of underperforming areas while maintaining well-performing areas25,26,27. Therefore, efficiency in conservation extends beyond evaluating output per unit, to include maximizing the relative contribution of conservation units (i.e., individual PAs) to the achievement of the entire network, highlighting the unique importance and irreplaceability of each unit within the system28.
Despite the progress in evaluating effectiveness and efficiency in habitat representativeness, most studies considered PAs as a unified network, overlooking the unique achievements and contributions of individual PAs. Individual PAs are foundational for achieving conservation outcomes at the local level, serving as the primary units for implementing conservation actions. However, research has revealed that PAs are often established in areas with low conservation costs rather than in areas of high biodiversity importance17. Abellande et al.29. found that European nationally designated PAs even cover fewer amphibians and reptiles than randomly selected areas. Furthermore, species within PAs do not always meet their conservation requirement, according to Li et al.30. found declines in populations of non-target carnivores in areas designated primarily for giant panda conservation. This mismatch between PA location and conservation needs underscores the importance of evaluating individual PAs’ effectiveness and efficiency within the network to ensure alignment with broader conservation objectives31. Existing studies have explored the cost-effectiveness24, representativeness31, and irreplaceability32 by evaluating the effects of individual PAs and finding differences in the performance of different PAs. However, they have largely overlooked the individual contributions or spatial efficiency within the broader network, as contributions are not simply the sum of the effects33. Current assessments may fail to comprehensively and accurately reflect these unique contributions, resulting in suboptimal evaluations that inadequately support PA-specific policy-making and future PA designation. On the other hand, efficiency assessments that focus solely on cost-effectiveness—often using financial cost or area as proxies—can lead to resource misallocations. Prioritizing areas that meet only minimal conservation requirements risks neglecting irreplaceable areas, potentially compromising the long-term resilience of the network. Spatial prioritization tools have highlighted this limitation, indicating that insufficient heterogeneity in feature and cost can lead to unreasonable conservation solutions34.
China recently released its National Biodiversity Conservation Strategy and Action Plan (2023–2030)35, committing to improve the representativeness of PAs and optimize resource efficiency to effectively conserve at least 30% of the national terrestrial area by 2030. However, existing studies in China have overlooked both the comprehensive consideration of habitat conservation needs in assessments and the unique achievements and contributions of individual PAs. Research has primarily focused on species coverage and richness36,37,38,39 or the achievement of a singular target40. Despite Li et al.41. considered species-specific needs, their analysis was limited to a subset of PAs and species. This highlights the urgent need to evaluate the current effects and importance of individual PAs, to inform the establishment of strategically located and resource-efficient PAs that align with these ambitious goals.
Taking habitat representativeness of China’s terrestrial vertebrates, this study aimed to provide an initial comprehensive assessment of individual PAs’ effectiveness and efficiency. We calculated habitat representativeness for both PA network and individual PAs effectiveness, and calculated representativeness contribution and representativeness density for individual PAs efficiency. In this study, habitat representativeness was defined as the extent to which a PA network or an individual PA covers the habitat conservation needs of species for adequate conservation. Representativeness contribution of an individual PA was defined as the extent to which an individual PA contributes to the network’s overall habitat representativeness, reflecting efficiency in contribution; representativeness density of an individual PA referred to the extent of habitat representativeness achieved per unit area within an individual PA, reflecting the efficiency of spatial use (Table 1).
First, we evaluated overall habitat representativeness by examining both the PA network and individual PAs to establish a foundational understanding. Then we assessed the representativeness contribution and representativeness density of individual PAs. Lastly, we classified PA types based on efficiency metrics to inform targeted conservation strategies and support data-driven resource allocation within the network for the KMGBF global conservation target.
Result
Overall habitat representativeness and the achievement of species habitat conservation needs
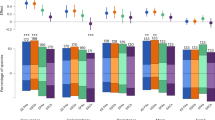
Habitat representativeness of current China’s PA network achieved 62.39%, indicating the average achievement of the species habitat conservation needs. Higher such proportional values were found in aves and mammals, but lower in amphibians and reptiles (Fig. 1 and Supplementary Table 1). There were 28.33% of species well-represented—conservation needs have been fully achieved—especially aves (37.07%) and mammals (33.79%). While 11.90% were severely under-represented with the achievement of conservation needs <10%, especially amphibians (22.59%) and reptiles (19.26%). Variation was also observed among different analysis lists (Fig. 2a). The habitat representativeness of the PA network of China’s key protected species (CNlist), at-risk species, and least concern species were 61.67%, 52.58%, and 66.57%, respectively (Supplementary Table 1). There were much less well-represented but much more under-represented species for at-risk species. Overall, we found the achievement of habitat conservation needs for every single species positively related (r = 0.38, p < 0.001) to their habitat areas, with severely under-represented species averaging only 7589.73 km2 (Fig. 2b).

a All species. b CNlist species. c At-risk species. d Least concern species. Taxa included amphibians (pink), aves (green), mammals (cerulean), and reptiles (purple).

a Habitat conservation needs (black circles), which is the representation target area, and conservation achievement (colored hexagons), which is the suitable habitat area covered by the PA network for species in different analysis lists. x-axis is log habitat area; y-axis is the percentage of the suitable habitat areas covered by the PA network. Hexagons above the black circles indicated the conservation needs of species’ habitat were fully achieved, and vice versa. b Relationship between habitat area and the conservation achievement of habitat conservation needs. x-axis presents the ratio of habitat areas covered by PA to the habitat conservation needs of each species; while (c) presents only the ratio of habitat areas covered by PA to the entire suitable habitat areas, namely, coverage.
Habitat representativeness of individual PAs ranged from 0.0004% to 55.51%, with an average of 0.35%. PAs with higher habitat representativeness exceeding the average were mostly located in western and northeastern China, including Qiangtang National Nature Reserve, Three-river-source National Park, Giant Panda National Park, Liangheyuantou Nature Reserve, and Hainan Tropical Rainforest National Park (Fig. 3a and Supplementary Table 2). However, the distribution of PAs with higher habitat representativeness varied across taxa. For instance, PAs in the Three-river source region showed higher habitat representativeness for aves and mammals, while PAs along the edge of Qinghai-Tibet Plateau (e.g., western Sichuan, southern Tibet, Qilian mountains) and Hainan demonstrated higher representativeness for amphibians (Supplementary Fig. 1). There were 56 PAs contained high-achievement species, whose habitat conservation needs were over 50% met within a single PA (Supplementary Fig. 2a and Supplementary Table 2), concentrating in western China, with few scattered in southern, northeastern China and Hainan, ranging in area from 40 to 321,376 km2. For instance, 34 species with more than 50% of their conservation needs have been achieved by Hainan Tropical Rainforest National Park. Hainan music frog (Nidirana hainanensis), Hainan odd-scaled snake (Achalinus hainanus), and Yellow-billed nuthatch (Sitta solangiae) reached 100%, 100%, and 99.42%, respectively.

a Habitat representativeness reflecting effectiveness. b Representativeness contribution and c representativeness density reflecting efficiency. Color legends are based on natural breaks classification in a and b, and quantile classification in c, with darker colors indicating higher values for each metric.
Efficiency of individual PAs
Efficiency of individual PAs was evaluated through representativeness contribution and representativeness density. Representativeness contribution ranged from 0.00051% to 20.74%, with an average of 0.36% (median = 0.07%). A total of 379 PAs exceeded the mean value, mostly located in western and southern China, and Hainan (Fig. 3b). Particularly, Hainan Tropical Rainforest National Park and Yarlung Zangbo Grand Canyon National Nature Reserve achieved 43.27% and 26.22% of habitat representativeness for at-risk species on average, respectively. The uneven contribution was discovered, as 175 PAs supported 751 high-dependent species (Supplementary Fig. 2b), with over 50% of their current conservation achievement contributed by a single PA, including 148 species fully dependent on one site. However, only 118 (16%) of these species had their conservation needs fully met (Supplementary Fig. 3). This also revealed that many of them remained under-represented and heavily reliant on specific PAs. For instance, the severely under-represented Spineless Stream Toad (Bufo aspinius), with only 4.65% of its conservation needs achieved, relied entirely on the Cangshan-Erhai National Nature Reserve for existing conservation.
Substantial difference was observed between representativeness density and contribution. The representativeness density reflecting spatial efficiency ranged from 0.000027% to 0.028% per km2, with an average of 0.00081% (median = 0.00059%) per km2. PAs with above-average concentration in southeastern China and along the eastern edges of Qinghai-Tibet Plateau, with others in northwestern and northeastern China (Fig. 3c).
To better inform conservation policy, we classified PAs into distinct categories based on the average value of representativeness contribution and density (Fig. 4). We identified 177 high-efficiency PAs, 202 contribution-efficient PAs, 401 spatially efficient PAs, and 1331 low-efficiency PAs. High-efficiency PAs were concentrated in southern and northwestern China, while contribution-efficient PAs were mainly distributed in western and northeastern China. Spatially efficient PAs, on the other hand, were concentrated in western Sichuan Province, eastern Jilin Province, and southern China. Significant relationships were found between current representativeness contribution and both species richness (r = 0.11, p < 0.001) and PA area (r = 0.60, p < 0.001) (Supplementary Table 3 and Supplementary Fig. 4). However, no significant relationship was found between current PA area and the number of high-dependence species (p = 0.26), suggesting that PA size alone would not determine their relative contribution. Similarly, no significant relationship was observed between representativeness density and PA area (p = 0.131), indicating that smaller or larger PAs were not inherently more spatial efficient in achieving conservation outcomes.

Protected areas across China were classified into four categories based on representativeness contribution and representativeness density: high-efficiency PAs (green), contribution-efficient PAs (purple), spatially-efficient PAs (blue), and low-efficiency PAs (gray).
Discussion
While conservation efforts continue to expand globally, the extent to which PAs achieve habitat representativeness varies across regions. Our findings on the unified PA network suggest that China is comparable to or exceeds some recent global estimates based on studies applying the principle from Rodrigues et al.8. For instance, global assessments have estimated that 21.7% of species met conservation needs16, whereas our results indicate 28.33% in China. Similarly, studies have suggested that 85% of globally threatened species remain underrepresented17, while in this study, the proportion was lower, with 78% of at-risk species and 70% of CNlist species lacking sufficient conservation. Another analysis has reported higher representativeness14, but they incorporated additional conservation designations beyond protected areas, including key biodiversity areas and ecologically intact landscapes.
This underscores that while China’s existing PA network provides a foundation for habitat representativeness, further efforts remain necessary to address remaining gaps. However, network-level assessments alone may overlook critical disparities in conservation efforts among individual PAs. By shifting the focus to individual PA performance, this study identified uneven conservation efforts, species-site dependencies, and efficiency trade-offs that inform more targeted and effective site-level conservation strategies. The following sections explore discovered gaps and their implications for optimizing conservation planning.
Disparities in conservation outcomes across regions and taxa
Current habitat representativeness revealed disparities across both regions and taxa, emphasizing gaps in China’s PA network42. These disparities could be shaped by a combination of regional ecological characteristics, taxonomic traits, and historical PA design, each contributing to the uneven conservation outcomes.
Regional disparities were reflected in the contrasting patterns of the efficiency of contribution and spatial use. Western China, characterized by large, intact PAs and environment, achieved higher representativeness contributions, and aligned well with previous studies emphasizing its crucial conservation outcomes42,43. However, the extensive size of western PAs often decreased their spatial efficiency, limiting per-unit effectiveness (although some were still small-areas PAs). Southern China, by contrast, demonstrated higher spatial efficiency may be due to smaller, spatially focused PAs addressing localized habitat needs, particularly for range-restricted taxa like reptiles. Yet, the intense human activity and fragmented habitats in the east of Hu line have constrained their performances38, especially the North China Plain, which faced additional challenges of low species richness and severe habitat fragmentation. This may be due to species richness was found to be related to efficiency metrics, with regions positively correlated with representativeness contribution and density. The lack of a consistent relationship between spatial efficiency and PA size further suggests that PA size alone does not determine efficiency; instead, the distribution of biodiversity also plays an important role. These patterns highlight the importance of regional ecological characteristics, including habitat fragmentation, species richness, and anthropogenic factors, in determining individual PAs effectiveness and efficiency.
Taxonomic disparities further emphasized the gaps in the existing PA network. Amphibians and reptiles exhibited the lowest conservation achievements, which may link to their fragmented ranges and specific environmental dependencies, making them harder to protect within the existing network44,45,46,47. Mammals and birds benefited from larger PAs such as Three-River-Source National Park and Giant Panda National Park, achieving higher conservation outcomes due to their wider and more contiguous ranges38,42. This uneven representation may also be due to historical PA designation, which prioritized wide-ranging, flagship species, leaving range-restricted and habitat-specific taxa under-represented30,38,46.
Addressing these disparities requires tailored and context-specific strategies. It is suggested to enhance connectivity among fragmented landscapes in southern China to improve both representativeness contribution and density. For example, expanding Laoshan Local Nature Reserve in Yunnan Province, which currently serves as the sole protector of the Maoson Shrub Frog (Philautus maosonensis) and Ludovic’s Glass Lizard (Dopasia ludovici), to include nearby suitable habitats could enhance the conservation for range-restricted species. These targeted approaches could only be identified by considering the individual PAs’ effectiveness, efficiency, and limitations.
Evaluating and balancing the roles of individual PAs
The PA network operates as a unified system composed of individual PAs, each contributing differently to conservation goals33. However, existing frameworks for analyzing PA conservation achievements often focus on generalized assessments, overlooking the need for PA-specific strategies that address individual PAs contributions and spatial efficiencies, which are critical for tailoring conservation efforts to specific regional or species needs48. To address this gap, we employed a dual-scale framework that integrates top-down and bottom-up perspectives to evaluate individual PAs effectiveness and efficiency within the context of habitat representativeness.
From a top-down perspective, network-level assessments offer a macro understanding of conservation outcomes, identifying systemic gaps such as underrepresented regions or taxa. However, this approach may overlook variability in individual PAs performance, which is essential for translating conservation goals into actionable strategies. The bottom-up perspective complements this by examining individual PAs’ effectiveness and efficiency, uncovering critical insights not apparent in network-level evaluations. Habitat representativeness could reveal how well individual PAs meet species-specific habitat conservation needs, revealing spatial disparities in effectiveness. Representativeness contribution assesses the relative contribution of individual PAs to network achievements, uncovering contribution-efficient PAs that disproportionately support conservation outcomes. This metric also highlights species heavily reliant on specific PAs under the existing conservation pattern and achievement, regardless of whether they are well- or under-represented. Representativeness density assesses spatial efficiency, providing insights into resource optimization. Our findings revealed spatial inefficiencies across both large and small PAs. Large PAs often underperformed due to diluted per-unit outcomes, while small PAs faced representativeness gaps or suboptimal site selection, challenging traditional assumptions that cost or size alone dictates efficiency.
This analysis also revealed a trade-off between the efficiency of contribution and spatial use, reflecting inherent complexities in balancing the support to conservation achievement and resource optimization. For instance, western China’s PAs excelled in contribution efficiency by protecting wide-ranging species but struggled with spatial efficiency due to their huge size. Conversely, southern China's PAs achieved higher spatial efficiency, but some contributed less, reflecting localized habitat focus. Spatially efficient PAs may indicate overprotection or a need for expansion. For overprotected PAs, they may protect species already effectively conserved by larger or more contribution-efficient PAs elsewhere—some in Sichuan Province, aligning with previous studies that found overlapping management42. In contrast, those that need expansion may contribute little due to their limited area, highlighting the necessity to include more important areas to enhance their contribution.
To optimize individual PAs, tailored strategies should integrate insights from efficiency metrics, classifications (Fig. 4), rankings (Supplementary Table 2), and network-level conservation gaps. Spatially efficient PAs, those potentially redundant should maintain their existing contributions and increase focus on delivering ecosystem services44, landscapes, or cultural45 values, while those requiring expansion should explore opportunities for increasing connectivity with surrounding priority areas to strengthen representativeness. Contribution-efficient PAs, given their irreplaceable roles in conservation outcomes, should focus on maintaining their impact while improving spatial efficiency through optimized management strategies. High-efficiency PAs, excelling in both contribution and spatial efficiency, could serve as benchmarks for replicating best practices. In contrast, low-efficiency PAs, those with suboptimal site selection, may prioritize addressing underrepresented taxa such as amphibians and reptiles or nearby gap species by expanding these PAs to increase connectivity and habitat representativeness.
By integrating effectiveness and efficiency metrics in habitat representativeness assessment on individual PAs, our framework bridges the gap between network-level discoveries and localized strategies, not only enhances resource allocation but also enables adaptive, context-specific conservation interventions to strengthen efforts across scales33.
Comparing conservation needs-based and traditional metrics
Traditional metrics used in China often assumed full habitat coverage or fixed targets (e.g., 30%) in assessments, neglecting differentiated conservation needs across species. We addressed this limitation by adopting a conservation needs-based metric that evaluates how well species’ habitat conservation requirements are met, leading to substantial differences in outcomes.
Metric incorporated species-specific conservation needs produced a more evenly distributed data structure, contrasting with the right-skewed distribution of traditional metrics that overlook the needs39 (Fig. 1 and Supplementary Fig. 5). This also led to needs-based metrics generally yielding higher values of habitat representativeness. Regarding conservation achievement for each species, a clear inverse correlation with habitat areas emerged, with well-represented species tending to have larger habitat, while under-represented species were predominantly small-range under needs-based metric, aligning with previous discoveries49. However, traditional metrics showed the opposite pattern (Fig. 2c), despite some studies showing that small-range species can be both well-represented and under-represented15,21,22.
These differences stem primarily from the habitat size. The traditional method, which relied on aggregate habitat coverage, inherently favors small-ranged species, such as amphibians and reptiles, as their smaller ranges were less challenged to meet fixed targets compared to wide-range species. This bias inflated the achievement for small-range species while masking unmet conservation needs. Conversely, wide-range species, such as birds and mammals, were often under-represented in traditional metrics because achieving full coverage across their extensive ranges is challenging and impractical. Needs-based metrics addressed this bias by linking representativeness directly to conservation needs, resulting in higher values for wide-range species and revealing gaps for small-range species. For example, birds were generally more under-represented in traditional metric but better represented in needs-based metric, whereas amphibians show the opposite trend (Supplementary Fig. 6). Overall, the need-based method better captured the feature of habitat size variation, not only allowed for better distinction between species with different conservation statuses but avoided misidentification of under-represented species due to wide distribution. Our results align with previous studies calculating such indices on global14,15 and national21,22,37 scale.
Limitations and future research
Habitat representativeness measures how well PAs meet species' habitat needs based on current distributions but does not account for historical habitat loss due to deforestation or urbanization. This requires caution in cross-national comparisons, as indices may not reflect baseline conditions. Also, representativeness alone may not fully capture conservation effectiveness, which depends on population dynamics and PA management quality.
Our calculations followed established metrics50,51 by averaging representativeness, contribution, and density, preventing PAs with higher species richness from automatically receiving higher values. This approach emphasized the relative responsibility of a PA for species within its boundaries rather than its absolute accumulation. However, averaging may have downscaled the absolute effect of species-rich PAs, potentially underestimating their broader conservation role. Future assessments should consider complementary metrics to balance site-level performance with overall conservation impact.
Using suitable habitat as a proxy may also overestimate representativeness if PA management is ineffective. Future research should include factors such as population viability, species interactions, and social traits to refine. Moreover, while this study assessed the current status of PAs, future research should also consider long-term dynamics, such as network expansion or climate-driven habitat shifts, to ensure adaptive conservation strategies. Additionally, randomization-based approaches could offer further insights into driven factors by exploring whether observed patterns are shaped by species distribution or driven by unique ecological or management factors. Such methods could broaden the applicability of this framework, offering refined perspectives on PA effectiveness and efficiency in diverse contexts.
Beyond assessing current conditions, our framework also offered a reference for conservation prioritization by incorporating multiple efficiency dimensions. Efficiency of contribution, which captures the relative importance and irreplaceability of individual PAs to network-level goals, should complement the efficiency of resource use to provide a more holistic assessment. Incorporating these insights into conservation decision-making could improve strategies and support the inclusion of efficiency metrics into broader and tailored conservation planning.
Methods
Data and materials
Multiple categories and data sources were selected in PAs data, including national parks52, national nature reserve53, local nature reserve54, scenic park55, national forest park55, and other PAs56. Except for scenic parks and national forest parks are point data, the others are polygon data. After removing duplicates, a total of 2149 PAs were included. Our data is consistent with the published research from Wang et al.57. To optimize calculation with 1 km × 1 km raster habitat data, a total of 2111 PAs (terrestrial part only) were retained in the final PA data set. This adjustment excluded 38 PAs smaller than 1 km², as these areas could not be effectively processed by the analytical tools. The retained PAs covered an area of 1.61 million km2, accounting for 16.75% of country’s terrestrial GIS area. The difference of area between the original 2149 PAs and the filtered 2111 terrestrial PAs was only 486.73 km2, accounting for 0.03% of original PAs. Compared to the officially designated PAs that cover approximately 18% of China’s terrestrial area, our final dataset accounts for about 93% of the total extent, focusing on spatially available and analyzable units, primarily composed of officially designated nature reserves and national parks.
Species distribution data for aves, mammals, reptiles, and amphibians were downloaded from IUCN Red List58. Using the minimum convex polygon method from Hou et al.45, the passage information, representing the migratory ranges from the breeding and wintering ranges for aves was also included. These original data were the extent of occurrence of each species, indicating the possible range of species occurrence59. To optimize and enhance the accuracy of data analysis, we then narrowed the extent of occurrence to the area of habitat that represented a more accurate range of species suitable habitat by taking into account species’ preference for different elevation and land cover categories45,60. There was a total of 2672 species in the suitable habitat dataset, including 1222 aves, 589 mammals, 405 reptiles, and 456 amphibians. Among them, 360 were threatened species (i.e., vulnerable, endangered, critically endangered), 157 were near threatened species, and 2024 were least concern species. In this research, we considered both threatened and near threatened species as at-risk species (n = 517) due to the latter being expected to face a high likelihood of qualifying for a threatened category in the near future58 and with special management concern61. Of the species analyzed, there were 564 belong to China’s key protected species lists (CNlist)62. Aves were the majority of all species, CNlist species, and least concern species, while at-risk species were mainly composed of amphibians. In our analysis, we included four species lists mentioned above—all, CNlist, at-risk, and least concern species. Given the urgent need for conservation, we would place greater emphasis on the first three lists. In addition, to facilitate calculations, all species habitat data were processed into 1 km × 1 km raster format.
Indicators for assessing effectiveness and efficiency in habitat representativeness
We calculated habitat representativeness for both PA network and individual PAs effectiveness; and calculated representativeness contribution and representativeness density for individual PAs efficiency (Table 1). For effectiveness metrics, we first followed the metric of mean target achievement50 and species protection index51 to calculate habitat representativeness, but equal weight was applied in this research to reflect the conservation status for the species within the context of China. The logic of equal weight is consistent with the logic of CNlist, which is to consider the status of both global and national key species. In other words, conservation actions should also be taken for species that are widely distributed globally but narrowly distributed and at risk in China. For example, the Large Indian civet (Viverra zibetha), Sable (Martes zibellina), and Wolverine (Gulo gulo) are LC species in IUCN Red List but first-class protected species in CNlist.
Habitat representativeness of a PA network was calculated as the average ratio of habitat areas covered by the PA network to the conservation needs of each species habitat (Eq. 1).
\(N\) is the total number of PAs. \({P}_{i}\) is the suitable habitat area covered by the PA network of species habitat \(i\). \({{RTA}}_{i}\) is the representation target area of species habitat \(i\), expressed as a percentage, indicating the proportion of suitable habitat area that need to be conserved of each species habitat for adequate conservation. We calculated representation target areas for each species habitat (Fig. 2a) using a piecewise function from Rodrigues et al.8, with thresholds adjusted according to Maiorano et al.18. to focus on the national scale. The lower limit of species habitat areas that should be protected is at least 10%, with its threshold adjusted from 250,000 km2 to 100,000 km2. The upper limit of species habitat areas that should be protected is at least 100%, with its threshold maintained at 1000 km2. Targets for species with habitat areas between these thresholds were interpolated proportionally.
Habitat representativeness of an individual PA \(j\) was calculated as the average ratio of habitat areas covered by an individual PA to the conservation needs of each species habitat (Eq. 2).
\({P}_{{ij}}\) is the suitable habitat area covered by PA \(j\) of species habitat \(i\). \(M\) is the total number of species within the corresponding PA \(j\).
For efficiency metrics, the efficiency of individual PAs was evaluated through representativeness contribution and representativeness density. Considering that species habitats that have achieved conservation needs may be contributed by one or more PAs, if 1 is still used as the maximum value, it is not able to reflect the true efficiency. Hence, in the calculation of the efficiency of individual PAs, we did not set the maximum value for the ratio of habitat area covered by PAs to the conservation needs of each species habitat.
Representativeness contribution of an individual PA \(j\) was calculated as each species' habitat conservation needs achieved by an individual PA to the network’s (Eq. 3).
Representativeness density of an individual PA \(j\) was calculated as the average ratio of each species habitat conservation needs achieved by an individual PA to its area (Eq. 4). \({{PA\; Area}}_{j}\) is the area of PA \(j\).
We then considered the average value of representativeness contribution and representativeness density as the threshold of the PA classification.
For species richness of an individual PA, we overlayed species suitable habitats with the polygon of individual PAs and counted the number of species occurring within that range.
Comparing results from different habitat representativeness calculation methods
We discussed and compared our conservation needs-based habitat representativeness to previous commonly used indices in China that did not account for the species' conservation needs. The habitat representativeness without representation target of a PA network calculated as Eq. (5):
\({{Habitat\; Area}}_{i}\) is the species suitable habitat areas of species i.
We compared the habitat representativeness value, data distribution, and the relationship between habitat representativeness and the area of species suitable habitat ofthe two methods.
Spatial analysis and statistics were performed by ArcGIS Pro and R 4.1.363.
Data availability
All datasets used in this study are described in the Methods section, with corresponding sources and access links listed in the references. Data used to plot the main graphs used in this study can be accessed at https://doi.org/10.6084/m9.figshare.28680446.v1
Code availability
Codes are available on request from the authors.
References
Convention on Biological Diversity. Kunming-Montreal Global Biodiversity Framework. (Convention on Biological Diversity, 2022).
Wilson, E. O. Half-Earth: Our Planets Fight for Life. (WW Norton & Company., 2016).
Yang, R. et al. Cost-effective priorities for the expansion of global terrestrial protected areas: Setting post-2020 global and national targets. Sci. Adv. 6, eabc3436 (2020).
Sundqvist, E., Backlund, F. & Chronéer, D. What is project efficiency and effectiveness?. Procedia Soc. Behav. Sci. 119, 278–287 (2014).
Goodman, P. S. & Pennings, J. M. New Perspectives on Organizational Effectiveness. (Jossey-Bass San Francisco, 1977).
Ostroff, C. & Schmitt, N. Configurations of organizational effectiveness and efficiency. Acad. Manag. J. 36, 1345–1361 (1993).
Abellán, M. D., Martínez, J. E., Palazón, J. A., Esteve, M. Á & Calvo, J. F. Efficiency of a protected-area network in a Mediterranean region: a multispecies assessment with raptors. Environ. Manag. 47, 983–991 (2011).
Rodrigues, A. S. L. et al. Global gap analysis: Priority regions for expanding the global protected-area network. Bioscience 54, 1092–1100 (2004).
Margules, C. R. & Pressey, R. L. Systematic conservation planning. Nature 405, 243–253 (2000).
Shaffer, M. L. Minimum population sizes for species conservation. Bioscience 31, 131–134 (1981).
Pe’er, G. et al. Toward better application of minimum area requirements in conservation planning. Biol. Conserv. 170, 92–102 (2014).
Cowling, R., Pressey, R., Rouget, M. & Lombard, A. A conservation plan for a global biodiversity hotspot—the Cape Floristic Region, South Africa. Biol. Conserv. 112, 191–216 (2003).
Pressey, R. L., Cowling, R. M. & Rouget, M. Formulating conservation targets for biodiversity pattern and process in the Cape Floristic Region, South Africa. Biol. Conserv. 112, 99–127 (2003).
Allan, J. R. et al. The minimum land area requiring conservation attention to safeguard biodiversity. Science 376, 1094 (2022).
Cantu-Salazar, L., Orme, C. D. L., Rasmussen, P. C., Blackburn, T. M. & Gaston, K. J. The performance of the global protected area system in capturing vertebrate geographic ranges. Biodivers. Conserv. 22, 1033–1047 (2013).
Maxwell, S. L. et al. Area-based conservation in the twenty-first century. Nature 586, 217–227 (2020).
Venter, O. et al. Targeting global protected area expansion for imperiled biodiversity. PloS Biol. 12, https://doi.org/10.1371/journal.pbio.1001891 (2014).
Maiorano, L. et al. On how much biodiversity is covered in Europe by national protected areas and by the Natura 2000 network: insights from terrestrial vertebrates. Conserv. Biol. 29, 986–995 (2015).
D’Amen, M. et al. Protected areas and insect conservation: questioning the effectiveness of N atura 2000 network for saproxylic beetles in I taly. Anim. Conserv. 16, 370–378 (2013).
Maiorano, L., Falcucci, A. & Boitani, L. Gap analysis of terrestrial vertebrates in Italy: Priorities for conservation planning in a human dominated landscape. Biol. Conserv. 133, 455–473 (2006).
Gonzalez-Maya, J. F., Vquez-R, L. R., Belant, J. L. & Ceballos, G. Effectiveness of Protected areas for representing species and populations of terrestrial mammals in Costa Rica. PLoS ONE 10, e0124480 (2015).
Watson, J. E. M. et al. The capacity of Australia’s protected-area system to represent threatened species. Conserv. Biol. 25, 324–332 (2011).
Burkhalter, J., Lockwood, J., Maslo, B., Fenn, K. & Leu, K. Effects of cost metric on cost-effectiveness of protected-area network design in urban landscapes. Conserv. Biol. 30, 403–412 (2016).
Fuller, R. A. et al. Replacing underperforming protected areas achieves better conservation outcomes. Nature 466, 365–367 (2010).
Aagaard, K., Lyons, J. E. & Thogmartin, W. E. Quantifying the relative contribution of an ecological reserve to conservation objectives. Glob. Ecol. Conserv. 9, 142–147 (2017).
Carwardine, J. et al. Conservation planning with irreplaceability: does the method matter?. Biodivers. Conserv. 16, 245–258 (2007).
Smith, P. G. & Theberge, J. B. Evaluating natural areas using multiple criteria: theory and practice. Environ. Manag. 11, 447–460 (1987).
Pressey, R., Johnson, I. & Wilson, P. Shades of irreplaceability: towards a measure of the contribution of sites to a reservation goal. Biodivers. Conserv. 3, 242–262 (1994).
Abellán, P. & Sánchez-Fernández, D. A gap analysis comparing the effectiveness of Natura 2000 and national protected area networks in representing European amphibians and reptiles. Biodivers. Conserv. 24, 1377–1390 (2015).
Li, S. et al. Retreat of large carnivores across the giant panda distribution range. Nat. Ecol. Evol. 4, 1327–1331 (2020).
Wiersma, Y. F. & Nudds, T. D. Efficiency and effectiveness in representative reserve design in Canada: the contribution of existing protected areas. Biol. Conserv. 142, 1639–1646 (2009).
Macedo, L., Monjeau, A. & Neves, A. Assessing the most irreplaceable protected areas for the conservation of mammals in the Atlantic forest: lessons for the governance of mosaics. Sustainability 11, 3029 (2019).
Rodrigues, A. S. L. & Cazalis, V. The multifaceted challenge of evaluating protected area effectiveness. Nat. Commun. 11, https://doi.org/10.1038/s41467-020-18989-2 (2020).
Hanson, J. O. & Schuster, R. Prioritizr Workshop Manual. https://prioritizr.github.io/PacMara_workshop/prioritizr-workshop-manual.pdf (2019). Access date: 24 November 2024.
Environment, M. O. E. A. China National Biodiversity Conservation Strategy and Action Plan (2023–2030). https://www.cbd.int/doc/world/cn/cn-nbsap-v3-zh.pdf (2024).
Hu, R. et al. A bird’s view of new conservation hotspots in China. Biol. Conserv. 211, 47–55 (2017).
Li, B. V. & Pimm, S. L. How China expanded its protected areas to conserve biodiversity. Curr. Biol. 30, R1334–R1340 (2020).
Xu, W. et al. Strengthening protected areas for biodiversity and ecosystem services in China. Proc. Natl. Acad. Sci. USA 114, 1601–1606 (2017).
Zhang, S.-Y. et al. Representativeness of threatened terrestrial vertebrates in nature reserves in China. Biol. Conserv. 246, https://doi.org/10.1016/j.biocon.2020.108599 (2020).
Fan, X., Xu, W., Zang, Z. & Ouyang, Z. Representativeness of China’s protected areas in conserving its diverse terrestrial ecosystems. Ecosyst. Health Sustain. 9, 0029 (2023).
Li, B. V. & Pimm, S. L. China’s endemic vertebrates sheltering under the protective umbrella of the giant panda. Conserv. Biol. 30, 329–339 (2016).
Xu, W. et al. Transforming protected area management in China. Trends Ecol. Evol. 34, 762–766 (2019).
Zhao, Z. et al. A protected area connectivity evaluation and strategy development framework for post-2020 biodiversity conservation. Land 11, 1670 (2022).
Cao, Y. et al. Identifying ecosystem service value and potential loss of wilderness areas in China to support post-2020 global biodiversity conservation. Sci. Total Environ. 846, 157348 (2022).
Hou, S. et al. A framework for identifying bird conservation priority areas in croplands at national level. J. Environ. Manag. 324, https://doi.org/10.1016/j.jenvman.2022.116330 (2022).
Wu, R. et al. Effectiveness of China’s nature reserves in representing ecological diversity. Front. Ecol. Environ. 9, 383–389 (2011).
Yang, Y. M., Tian, K., Hao, J. M., Pei, S. J. & Yang, Y. X. Biodiversity and biodiversity conservation in Yunnan, China. Biodivers. Conserv. 13, 813–826 (2004).
Lee, W. H. & Abdullah, S. A. Framework to develop a consolidated index model to evaluate the conservation effectiveness of protected areas. Ecol. Indic. 102, 131–144 (2019).
Senior, R. A. et al. Global shortfalls in documented actions to conserve biodiversity. Nature 630, 387–391 (2024).
Jantke, K., Kuempel, C. D., McGowan, J., Chauvenet, A. L. & Possingham, H. P. Metrics for evaluating representation target achievement in protected area networks. Diversity Distrib. 25, 170–175 (2019).
Jetz, W. et al. Include biodiversity representation indicators in area-based conservation targets. Nat. Ecol. Evol. 6, 123–126 (2022).
China’s State Council. China’s First 5 National Parks Guard Wildlife. english.www.gov.cn/policies (2021).
Resource and Environmental Science Data Platform. National Nature Reserve Boundary Data. https://www.resdc.cn/data.aspx?DATAID=272 (2022).
Ministry of Ecology and Environment. List of Nature Reserves in 2017. https://www.mee.gov.cn/ywgz/zrstbh/zrbhdjg/201905/P020190514616282907461.pdf (2019).
Protected Area Platform. Nature Park. http://www.cnnpark.com/res-np-f-site.html (2022).
IUCN and UNEP-WCMC. The World Database on Protected Areas (WDPA) [On-line], [09/2022]. www.protectedplanet.net (2022).
Wang, F. et al. Over 1/4 of China’s terrestrial area significantly contributed both to biodiversity conservation and carbon neutrality, requiring protection. Sci. Total Environ. 912, 169070 (2024).
IUCN. IUCN Red List of Threatened Species. Version 2022. https://www.iucnredlist.org (2022).
Brooks, T. M. et al. Measuring Terrestrial Area of Habitat (AOH) and Its Utility for the IUCN Red List. Trends Ecol. Evolution 34, 977–986 (2019).
Peng, Q. et al. One-third of lands face high conflict risk between biodiversity conservation and human activities in China. J. Environ. Manage. 299, https://doi.org/10.1016/j.jenvman.2021.113449 (2021).
National Park Service. The Future of Species, www.nps.gov (2022).
National Forestry and Grassland Administration. China’s Key Protected Species Lists. http://www.forestry.gov.cn/main/5461/20210205/122418860831352.html (2021).
R Core Team. R: A Language and Environment for Statistical Computing (R Core Team, 2022).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant No. 51978365, Grant No. 52378060, and Grant No. 52408075). Pei Wang, Xiaoshan Wang, Hao Wang and Ruijia Zhu contributed to expressing our ideas and refining the manuscript.
Author information
Authors and Affiliations
Contributions
T.H.T. and R.Y. conceived the study; T.H.T. designed the research with F.W. and R.Y.; T.H.T., F.W., and S.H. collected data; T.H.T. performed the analysis; T.H.T., F.W., and M.S. interpreted results and wrote the manuscript; Y.C., S.H., Y.Z., Z.Z., and R.Y. commented and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Diogo Alagador and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Heike Langenberg and Aliénor Lavergne. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tseng, TH., Wang, F., Sun, M. et al. Protected areas unevenly contribute to terrestrial vertebrate habitat conservation in China. Commun Earth Environ 6, 313 (2025). https://doi.org/10.1038/s43247-025-02281-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02281-6
This article is cited by
-
Projected human land-use pressures and natural habitat conversion risk within global terrestrial protected areas
Nature Ecology & Evolution (2026)
