
Abstract
Continental Cretaceous-Paleogene boundary sections are best known from North America, where they invariably exhibit a marked shift in sedimentary facies at or very near the boundary level. Uppermost Cretaceous strata typically reflect water-logged soils and unstable meandering-river deposits, whereas lowermost Paleogene strata typically reflect coal swamps and broad, stable meander-belt deposits. Causal links between facies shifts at the Cretaceous-Paleogene boundary and the end-Cretaceous mass extinction have been largely dismissed. Here, we present five new Cretaceous-Paleogene boundary sections identified via iridium anomalies in the Bighorn and Williston basins and assess the sedimentological changes that occur at North American Cretaceous-Paleogene boundaries. We hypothesize that the geographically widespread Cretaceous–Paleogene facies shifts were driven by the extinction of dinosaur megafauna. Large-bodied dinosaurs likely promoted open vegetation structure, prompting fluvial avulsion and clastic sediment input to distal floodplains. After the end-Cretaceous mass extinction, dense forests could establish, stabilizing meander belts and starving the floodplain of clastic sediment, favoring the accumulation of organic-rich strata. More empirical data are needed, but facies change in continental Cretaceous-Paleogene boundary sections suggests dinosaurs were ecosystem engineers that promoted habitat openness in the Late Cretaceous, and their extinction likely led to a dramatic reorganization of ecosystem structure in the earliest Paleogene.
Similar content being viewed by others

Introduction
The Cretaceous-Paleogene boundary (KPB) is defined by an elevated iridium anomaly along with other ejecta from the Chicxulub asteroid impact ca. 66 million years ago1,2. The causes and biotic consequences of the end-Cretaceous mass extinction (EKME) have received considerable attention3,4,5, and that sudden disappearance of large swaths of the biosphere had reverberating effects on the geosphere, at least in the marine realm6. Did the most famous victims of the EKME—dinosaur megafauna—leave a similar imprint on the continental rock record?
Herbivorous terrestrial megafauna (>60 kg)7 are ecosystem engineers—they actively modify and ultimately structure their environment by maintaining habitat openness and vegetational heterogeneity via trampling and uprooting, suppressing the establishment of dense, monodominant forests7,8,9. When herbivorous megafauna are removed from areas climatically suited for dense forests, those forests can encroach, decreasing both habitat openness and spatial vegetation heterogeneity7,8. Vegetation cover (i.e., open vs. densely forested) in turn controls fluvial geomorphology: in more sparsely or patchily vegetated landscapes, river meanders are unstable, crevasse splays and avulsions are frequent, and meander-belt persistence is ephemeral, whereas dense, forested vegetation stabilizes meander belts and restricts splaying and avulsion10,11. Disappearance of megafauna can thus abruptly restructure vegetation and terrestrial ecosystems7,8, consequently altering fluvial dynamics and their expression in the sedimentological record10,11.
The extinction of non-avian dinosaurs at the KPB is the most profound example of megafauna being abruptly removed from fluvial environments. In the Western Interior of North America (Western Interior, hereafter), the disappearance of dinosaur megafauna coincides with a shift in fluvial sedimentary facies that is mappable in outcrop and thus formalized in stratigraphic nomenclature denoting contacts between uppermost Cretaceous (e.g., Lance and Hell Creek) and Paleocene (e.g., Fort Union) formations3,5,12. Direct evidence of the Chicxulub asteroid impact frequently coincides with those formational contacts (Fig. 1), especially in the Williston Basin, which hosts the most complete continental KPB sections in the world12. There, a thin, red claystone interval at the base of the lowest coal or lignite (simply ‘coals’, hereafter) of the Fort Union Formation (i) frequently yields elevated iridium anomalies, shocked quartz, and altered microtektite spherules12 and (ii) is a few cm below a volcanic ash bed coincident in age with Chicxulub tektites from Haiti13,14. Similarly abrupt KPB facies shifts—sharp changes in lithology, fluvial architecture, and floodplain deposition that are stark in outcrop appearance—have been documented throughout the Western Interior, and they may be a global phenomenon, but the present incompleteness of continental KPB records elsewhere prevents that unequivocal assessment here (see Supplementary Discussion). The current consensus is that, despite their synchronicity, KPB facies shifts are unrelated to the EKME and instead reflect chance overprinting of a global catastrophic event on what was otherwise a time-transgressive paleoenvironmental trend15,16,17,18,19. Here, we report on five newly discovered KPB sections from the Western Interior that further support the coincidence between continental facies shifts and the EKME, prompting us to reevaluate pre- and post-KPB sedimentology15,16,17,18,19,20,21 and the mechanisms underlying those geological changes.

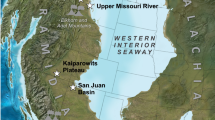
Map of continental Cretaceous-Paleogene boundary (KPB) locations in the Western Interior of North America with new KPB sections highlighted in red boxes (left panel) and typical KPB-coincident facies changes (right panel) (map from Ron Blakey [Deep Time Maps]; note that this artistic reconstruction is meant to illustrate major landscape features in the Western Interior near the KPB, not to imply support for landscape interpretations; see Supplementary Table 1 for a full list of KPB localities). The pie chart indicates the relationship between impact-identified KPBs and local facies changes. Despite being geographically and paleoenvironmentally disparate, most KPBs are previously recognized to coincide with a facies shift, either at a formational contact (red stars) or within a formation (yellow circles). In the right panel, note that the uppermost Cretaceous strata are dominated by overbank mudstone beds and thin fluvial-channel or crevasse-splay sandstone beds. In contrast, lowermost Paleogene strata are dominated by thick, cliff-forming sandstone beds representing multistoried fluvial-channel deposits (Raton and Bighorn basin panels) or thin, gently dipping, alternating mudstone-sandstone beds representing point-bar deposits (Williston Basin panels). Putative ‘exceptions’ to the KPB-coincident facies change (white-to-blue symbols), which we challenge below (see ‘Abiotic drivers are insufficient to explain KPB facies shifts’ section and Supplementary Discussion), are localities where reported KPBs lie within centimeters below an intraformational coal (a, white pentagon), atop a formation-defining coal (b, light-blue triangle), below a formation-defining coal (c, inverted royal-blue triangle), or within an intraformational coal (d, dark blue square). Note that in some places multiple KPBs have been reported from the same set of outcrops (see Supplementary Table 1). See main text and Supplementary Discussion for details. Photograph credit: PK (Raton Basin), LNW (Bighorn Basin, Hell Creek), TST (Mill Iron).
Results and discussion
New KPB sections prompt a new paradigm
It is remarkable that, across ~30° latitude (~37°–65° N), ~15° longitude (~104°–117° W) (Fig. 1), and diverse paleoenvironmental settings12,22,23, continental KPBs in the Western Interior can be reliably discovered by targeting obvious facies shifts. To test the prevalence of that phenomenon, we sampled for iridium anomalies at formational contacts that were plausible KPB candidates based on paleontological evidence (see ‘Methods’), yet captured distinct paleolandscape settings23. These included: Lance-Fort Union formational contacts in the Bighorn Basin, Wyoming (Supplementary Fig. 1; Supplementary Fig. 2), a region bounded by Laramide mountain belts, and the Hell Creek-Fort Union formational contact at Mill Iron, near Ekalaka, Montana (Supplementary Fig. 3), a region far from elevated topography (Fig. 1). We discovered elevated iridium anomalies, which we attribute to the Chicxulub asteroid impact and thus the KPB1, at all five of the sampled formational contacts (Supplementary Table 2). These represent the first KPBs identified in the Bighorn Basin and the southernmost KPB in the Williston Basin (Fig. 1). Together, they reinforce the pattern that continental KPBs invariably coincide with a facies shift across disparate paleoenvironments (see Supplementary Discussion).
These observations prompt us to challenge the prevailing notion that KPB facies shifts are independent of the EKME. Instead, we propose a new hypothesis: dinosaur megafauna were ecosystem engineers during the Late Cretaceous, and their sudden extinction caused an abrupt shift in continental facies at the KPB that persisted into the early Paleocene (the ‘Dinosaurs as Ecosystem Engineers’ hypothesis). We demonstrate further support for this hypothesis below, focusing especially on the Williston Basin since it hosts most of the known continental KPB sections, has superb three-dimensional outcrop exposures that most other KPB sections lack, and has featured prominently in debates about the EKME12,19,22.
KPB facies shifts reflect changes in fluvial systems, not widespread ponding
Continental facies shifts at the KPB most obviously reflect two aspects of fluvial systems: (i) river-meander-belt size and stability and (ii) the amount of clastic sediment (clay, silt, sand) deposited on the distal floodplain. Uppermost Cretaceous meander belts are characteristically narrow and unstable, delivering a steady supply of clastics to the distal floodplain; lower Paleogene meander belts are characteristically wide and stable, starving the distal floodplains of clastics and permitting the accumulation of concentrated organic debris. Those differences between pre- and post-KPB fluvial systems are epitomized by the Hell Creek/Lance and Fort Union formations, respectively, of the Williston (Hell Creek) and Powder River (Lance) basins (Fig. 2). The Lance/Hell Creek and Fort Union formations are all meandering-river deposits, evidenced by the preponderance of fine-grained clastics, erosive and asymmetrical channel bases, abandoned-channel deposits (oxbow lakes), and lateral-accretion beds15,16,17,18,19 (Fig. 2; Supplementary Fig. 4). The Hell Creek and Lance formations are characterized by (i) predominantly small (~1–3-m thick, ~5–10-m wide), single-storied fluvial-sandstone beds (small, short-lived river channels or crevasse splays), with large (~5–10-m thick, ~0.1–0.5-km wide), multistoried fluvial sandstone beds being rare, interbedded with (ii) massive, green-gray mudstone beds that exhibit root traces, slickensides, gleying, and gray–yellow mottling (immature, water-logged soils; i.e., hydromorphic paleosols)3,15,16,18,19,20. The lower Fort Union Formation is characterized by (i) large (~8–30-m thick, 0.1–1-km wide), multistoried fluvial sandstone beds (large, amalgamating, proximal river channels) and ‘variegated beds’ (associated point bars; see below) and (iii) coals (swamp-dominated, organic-rich distal floodplains)3,16,19,24.

Continental facies typical of the uppermost Cretaceous and lowermost Paleogene, epitomized by the Hell Creek Formation (A–D) and Tullock Member of the Fort Union Formation (E–J) in the Hell Creek region of northeastern Montana (Fig. 1). The upper (A), middle (B), and lower (C) portions of the Hell Creek Formation are all dominated by fine-grained clastic sediments, interpreted as hydromorphic paleosols, and small fluvial channels or crevasse splays (which sometimes scour into those paleosols, as in D). The Tullock Member of the Fort Union Formation is characterized by laterally persistent coals, ‘variegated beds’, and multistoried proximal fluvial-channel deposits (E). We interpret the ‘variegated beds’—packages of thinly interbedded coarse-fine clastic sediments—as lateral-accretion or point-bar deposits associated with large meandering channels because they (i) dip at a 5–25° angle and coarsen down-dip and fine upwards (F), (ii) exhibit paleoflow indicators that are perpendicular to the dip direction (G), and frequently rework and cross-cut each other (H). The KPB claystone bed is frequently overlain by a coal and then a series of those cross-cutting lateral-accretion beds (I, J). Triangles in B and F indicate fining-upwards sequences; note the difference between those in the Hell Creek versus Fort Union. Arrows in E and F highlight depositional dip directions. Carb. carbonaceous, LA lateral-accretion set, MS multistoried proximal fluvial-channel deposit.
The sudden, geographically widespread appearance of coals is the most conspicuous feature of lower Paleogene facies in the Western Interior and defines the base of the Fort Union Formation3,12,16,24. Even in the absence of coals, however, the inorganic sedimentology of the Fort Union Formation is readily distinguished from the underlying Hell Creek or Lance formations, largely due to the ‘variegated beds’—brightly colored, thinly interbedded claystone, siltstone, and fine-grained sandstone beds that are often under- and overlain by coals and were thus likely deposited laterally and coevally to coal deposits elsewhere16,17. These ‘variegated beds’ have historically been interpreted as pond or lake deposits16,18,19, and Fort Union deposition has thus been attributed to various mechanisms to increase ‘wetness’ in the early Paleocene, resulting in mostly ponded–marshy accumulation of clastic and organic sediments12,15,16,17,18,19,20. We collectively term these proposed mechanisms, which include increased precipitation16,17,20 and/or a rise in water table16,18,19,20, the ‘Wet Paleocene’ hypothesis.
New observations from our work in the Hell Creek region of northeastern Montana, in conjunction with previous research24,25,26, cast doubt on the ‘Wet Paleocene’ hypothesis: (i) Hell Creek Formation paleosols denote the prior existence of a very high water table, and (ii) most ‘variegated beds’ of the Fort Union Formation are not pond deposits but rather lateral-accretion deposits associated with large, broadly meandering rivers27 (Fig. 2; Supplementary Fig. 4). There is substantial support for the first point19,20,24—most overbank deposits in the Hell Creek Formation represent ponds or hydromorphic paleosols. The second point, to our knowledge, has not been fully appreciated. In the dozens of outcrops studied across the eastern Montana portion of the Williston Basin, we found that the ‘variegated beds’ consist of alternating fine- and coarse-grained sediments that dip at ~5–25°, coarsen down-dip, frequently grade into proximal channel-sandstone deposits, and exhibit paleoflow indicators (e.g., ripple marks) that are perpendicular to dip direction (Fig. 2)24,26. Further, although paleoflow directions remain roughly east–northeast across the KPB in our study area (Hell Creek mean = 109°, n = 166; Fort Union mean = 63.5°, n = 310), the standard deviations from those mean paleoflow directions are substantially higher in the Fort Union Formation (SD = 69.3°) compared to the Hell Creek Formation (SD = 52.9°), suggestive of a more pronounced meandering system in the early Paleocene. Thus, instead of representing pond or lake deposits, the ‘variegated beds’ represent point-bar deposits (and potentially counterpoint bar deposits, see Supplementary Discussion) associated with the lateral migration of fluvial-channel meanders27. This interpretation is consistent with the available paleoclimatic data, which indicates neither an abrupt nor persistent change in precipitation intensity across the KPB23 (see Supplementary Discussion).
Abiotic drivers are insufficient to explain KPB facies shifts
Transgression of the Cannonball Sea (Fig. 1) is most commonly invoked to explain the ‘Wet Paleocene’ interpretation of KPB facies change in the Williston Basin12,15,18,19 (the ‘Marine Transgression’ hypothesis), largely based on instances of minor (cm–m scale) stratigraphic offset between purported KPBs and the Hell Creek-Fort Union formational contact (i.e., the facies change is time-transgressive, not synchronous with the KPB). However, most of those KPBs are identified using non-impact-related tools or proxies like palynology or carbon-isotope stratigraphy12, which are sensitive to reworking28 and lithology25 and thus insufficiently precise to place the KPB at ≤ 2 m scales (this has been noted previously19,22). Palynology is still an important tool for constraining the approximate stratigraphic position of the KPB at the outcrop scale, and pollen is often the best tool available in certain locations12,19, despite its propensity for remobilization by fluvial action19,22,28 (see Supplementary Discussion for a fuller discussion about the merits and shortcomings of using pollen or carbon isotopes to place the KPB).
Although some impact-identified KPBs are located a few cm below, within, or atop a coal (Fig. 1; Supplementary Table 2), those putative examples of time-transgressive KPB facies change assume that coals are the sole sedimentological indicator of that change12. Yet, a preponderance of coals in lower Paleocene strata is merely one symptom of the systematic shift in the nature of fluvial systems at the KPB (Fig. 2). Indeed, coal-bearing units do occur in the Late Cretaceous21,29,30, including infrequently in the Hell Creek Formation3,14; that some KPBs would be preserved in Cretaceous coal swamps that persisted into the early Paleocene should therefore be expected (Supplementary Fig. 5; see Supplementary Discussion).
Further, magnetostratigraphy indicates that Cannonball Sea transgression began no earlier than chron C29n and occurred most substantially during chron C28r31,32, thus postdating the KPB (in the middle of chron C29r) by ca. 300–900 ka14 (Supplementary Fig. 5). Marine transgression should also result in high-accommodation features in the continental record, such as hydromorphic paleosols, low channel:floodplain ratios, and single-story channels29,30; a pattern that more closely matches the Hell Creek Formation (which does interfinger with marine deposits to the east) than the Fort Union Formation15,19 (Fig. 2; Supplementary Fig. 5). Lastly, the ‘Marine Transgression’ hypothesis fails to explain why facies changes analogous to those in the Williston Basin coincide precisely with the KPB in geographically distant areas like the Alberta and Powder River basins12,17,22, rather than tracking the advance of Cannonball Sea15 (Supplementary Fig. 5), or why facies changes occur at the KPB in areas like the Bighorn Basin that were separated from the rest of the Western Interior by Laramide arches (Fig. 1). Thus, the preponderance of evidence casts doubt on the ‘Marine Transgression’ hypothesis; the KPB and lower Fort Union more likely coincided with a marine regression or lowstand, both locally31 and globally33.
The Laramide Orogeny has also been proposed to explain KPB facies shifts21, but that process was spatially diachronous and insufficiently abrupt to generate the observed stratigraphic patterns across such a large geographic range (Fig. 1)—Laramide deformation advanced from the southwest to the northeast of the Western Interior over the course of the Late Cretaceous through early Paleogene, and there is no evidence of a continent-wide pulse of basement-cored block uplifts coincident with the KPB34. Landscape denudation via Chicxulub-associated wildfires, in contrast, could be both abrupt and widespread19,35, but such a mechanism cannot explain the persistence of those facies changes throughout the lower Fort Union Formation for a temporal duration exceeding 1 million years14,24,26. Thus, no previous hypotheses adequately explain the observed facies shifts across the KPB (see Supplementary Discussion).
Dinosaur megafauna as ecosystem engineers
We propose that the sedimentological and stratigraphic features that define Upper Cretaceous versus lower Paleogene continental strata are the result of ecosystem engineering by dinosaur megafauna in the Cretaceous and their disappearance in the Paleogene (Fig. 3). The ‘Dinosaurs as Ecosystem Engineers’ hypothesis better explains the synchroneity, style, and persistence of continental facies change and is linked to a causal mechanism that is well-documented in Quaternary terrestrial ecosystems7,8,9,10,11. Mammalian megafauna profoundly affect landscapes and vegetation7,8, but dinosaurian megafauna were many times larger, and the spatial extent of ecosystem engineering scales positively with body size36. Even smaller herbivorous dinosaurs likely reached body masses of ~1000 kg, and some (e.g., Triceratops horridus) may have approached 15,000 kg37, so their impact on terrestrial vegetation structure and fluvial dynamics was likely profound.

Large-bodied dinosaurs likely promoted an open vegetation structure that permitted frequent fluvial avulsion and clastic sediment input to distal floodplains. After the end-Cretaceous mass extinction, dense, closed-canopy forests could take root, stabilizing meander belts and starving the distal floodplain of clastic sediment, promoting the accumulation of organic-rich strata.
Not only were individuals of these species massive, but taphonomic studies indicate most ceratopsians and hadrosaurs were herd animals38. These herds, along with rarer large-bodied predators (e.g., Tyrannosaurus rex, ~8000 kg37), likely maintained open or patchily forested habitats during the Late Cretaceous via trampling and uprooting39. Consequently, fluvial systems would flood and avulse frequently40, resulting in: (i) short-lived meander belts and (ii) a frequent supply of clastic sediment to the distal floodplain, consistent with the (i) small, single-storied fluvial channels, thin crevasse-splay sandstone beds, and (ii) hydromorphic paleosols that characterize Upper Cretaceous strata. When those megafauna went extinct, so too did their influence on vegetation cover, allowing dense forests to take root, resulting in stable and long-lived river-meander belts that would have: (i) allowed prolonged reworking of channel meanders and (ii) starved the distal floodplain of clastics thus promoting organic accumulation, consistent with the (i) large, multistoried sandstone channels with abundant adjacent lateral-accretion bedding and (ii) laterally continuous coals that characterize lower Paleocene strata (Fig. 3).
Whether or not non-avian dinosaurs were in decline prior to the EKME remains debated41,42. Nevertheless, in the Western Interior, large-bodied dinosaurs, especially ceratopsians, persisted up to the KPB and their disappearance during the EKME was geologically abrupt43. Indeed, in nearly all of the KPB localities discussed here, dinosaurian megafauna are well-known from below the boundary and disappear at the KPB. Further, proposals of dinosaur decline specifically refer to taxonomic richness, not relative abundance—the former is germane to evolutionary interpretations, but the latter is most relevant to ecological impacts, and there is no evidence that large-bodied dinosaurs were numerically depauperate prior to the EKME. Lastly, although mammals underwent adaptive radiations in the Late Cretaceous23 and increased in body size quickly in the earliest Paleocene44, mammals remained relatively small for much of the early Paleogene45; as such, mammals did not likely substantially impact the structure of standing vegetation until the Eocene or Oligocene7.
Importantly, the immediate consequences of the Chicxulub asteroid impact on standing vegetation cover were relatively short-lived and only evident at the cm scale in the rock record12,22. Although plant taxonomic recovery from the EKME likely took thousands to millions of years44,46,47, lower taxonomic richness does not preclude forest density or canopy structure39. Indeed, the warm, monsoonal climates of the earliest Paleocene of the Western Interior23 would have allowed the establishment of dense forest vegetation in a matter of decades to centuries post-KPB44. The ‘Dinosaurs as Ecosystem Engineers’ hypothesis can therefore explain the pace, style, and persistence of the continental KPB facies shift and is consistent with the vertebrate3,4,5,19,26 and plant12,22,39,44,46,47 fossil records (see Supplementary Discussion).
Regional variation in the KPB facies shift
The facies shifts described above reflect landscape-scale changes in fluvial geomorphology that are likely synchronous with the KPB18,19; yet, KPB-coincident facies changes are not identical in mountain-proximal versus mountain-distal portions of the Western Interior (Fig. 4). Those in the Williston Basin are the best studied, and they broadly match the KPB facies shifts observed in other mountain-distal basins, such as the Powder River, Alberta, and Brackett basins12,17 (Fig. 4; but see Supplementary Discussion). The KPB in the Willow Creek Formation of the Alberta Foothills was deposited off the flank of the Sevier thrust belt (Fig. 1) and thus reflects a unique facies change reflective of locally well-drained, semiarid conditions48. KPBs in the Raton Formation of the Raton Basin (Fig. 1) mark the contact between the underlying ‘lower coal zone’ and overlying ‘barren series’ and superjacent ‘upper coal zone’21,49; the former is characterized by single-storied channels and hydromorphic paleosols, whereas the ‘barren series’ is dominated by multistoried, laterally aggraded, channel-sandstone beds21,49. KPB facies shifts in the Bighorn Basin closely match those in the Raton Basin (Fig. 4; see Supplementary Discussion), differing mainly in an absence of coals in the underlying Lance Formation. KPBs from the Denver Basin may exhibit a facies change like those in the Raton and Bighorn basins; however, the only impact-identified Denver Basin KPBs come from a drill core and a poorly exposed outcrop in the central and eastern basin44, respectively, precluding precise assessment of the KPB facies change currently (but see Supplementary Discussion). We contend that those similarities between the Raton, Bighorn, and potentially Denver basins reflect their proximity to emerging Laramide mountain belts (Figs. 1 and 4).

Diagram illustrating the abrupt continental facies changes that occur at impact-identified KPBs (red line) across the Western Interior (see Fig. 1), with approximate latitude given beneath each area to emphasize the geographic extent of those facies changes (note that these are modern latitudes, and diagrams are illustrative, not to precise stratigraphic scale). Question marks on different facies symbols represent uncertainties or ambiguities about those facies; the question mark on the KPB line in the Denver Basin represents uncertainty about the precise location of the KPB in the mountain-proximal portions of that basin. Note that these are generalized schematics that do not capture all sedimentological or stratigraphic details of a given area—for a more detailed discussion of these different localities, see the Supplementary Discussion. AB Alberta, CAN Canada, CR Castle River area of southern Alberta, CV Coal Valley area of western Alberta, Dist. mountain-distal portions of the Denver Basin, ID’d identified, lat. lateral, MS multistoried, Prox. mountain-proximal portions of Denver Basin, RDV Red Deer Valley Alberta, SS single-storied, UCZ ‘upper coal zone’, USA United States of America.
That continental KPB facies shifts in mountain-proximal settings are not identical to those in mountain-distal settings is expected given that (i) the effects of vegetation and ecosystem engineering on fluvial geomorphology10,50, and (ii) the drivers of river avulsion51, both of which are central to the ‘Dinosaurs as Ecosystem Engineers’ hypothesis, differ markedly in mountain-distal versus mountain-proximal settings (Fig. 3). Those differences could explain the abrupt appearance of laterally expansive and vertically thick proximal channel deposits overlying the KPB in places like the Raton21,49 and Bighorn basins (see Supplementary Discussion).
Implications for the evolution of landscapes and biota across the KPB
The ‘Dinosaurs as Ecosystem Engineers’ hypothesis explains both the geographically widespread coincidence between the KPB and continental facies shifts and provides a mechanism for those changes. Nonetheless, more empirical data are needed to robustly evaluate its legitimacy; in particular, more quantitative assessments of fluvial geomorphology and plant-canopy structure across the KPB throughout the Western Interior. Our hypothesis is falsifiable and testable in numerous ways, such as: (i) Continental, impact-identified KPBs should always be closely associated with a sedimentary facies change; (ii) Forest canopy structure, which is commonly inferred via light-sensitive proxies52, should be relatively open in the latest Cretaceous and relatively closed in the earliest Paleocene39; and (iii) Fluvial deposits overlying the KPB should reflect more broadly meandering and temporally long-lived rivers relative to those below the KPB, at least in mountain-distal settings.
If true, the ‘Dinosaurs as Ecosystem Engineers’ hypothesis has important implications for the evolution of terrestrial ecosystems. For example, early flowering plants were weedy opportunists that thrived in frequently disturbed environments53, and avulsion-prone fluvial systems of the Cretaceous maintained by dinosaur megafauna may have promoted their early ecological proliferation. The closing of forest canopies predicted by the ‘Dinosaurs as Ecosystem Engineers’ hypothesis is also consistent with observed increases in angiosperm-seed size after the KPB, attributed to the emergence of dense forests following the extinction of dinosaur megafauna54,55,56. Lastly, most Mesozoic mammals were likely terrestrial or semifossorial57, matching expectations for an open rather than densely forested habitat58. Widespread emergence of dense forest cover in the Paleocene may have therefore prompted the post-KPB rise of arboreal and frugivorous mammals58, such as primates.
In sum, this work highlights the need for continued research on the Cretaceous–Paleogene transition in continental settings. It is increasingly apparent that abiotic drivers like climate change and tectonics have shaped the distribution and evolution of life23; yet, as demonstrated here, life itself can also sculpt our planet. By bearing these relationships in mind, we may find that Earth’s climate, landscapes, and biota have been tightly entwined throughout geologic history.
Methods
The five new Cretaceous-Paleogene boundary (KPB) sections reported here are within the Bighorn Basin of northwestern Wyoming (USA) and the southeast Montana (USA) portion of the Williston Basin (Fig. 1). Defining an impactite-level resolution for the KPB typically includes three elements12: (i) physical evidence of the impact, (ii) occurrence of that impact evidence within magnetochron C29r, and (iii) biostratigraphic evidence of faunal or floral turnover. All five of these new KPBs occur at formational contacts and satisfy criteria (i) and (iii)—they were primarily identified via iridium anomalies exceeding local background limits of <0.1 ppb1 (Supplementary Table 2), and they are consistent with the vertebrate-fossil records from below and above those iridium anomalies. Magnetostratigraphic data (criterion [ii]) for these sections are pending, but there is no evidence for major depositional hiatuses or erosional unconformities in these sections that would preclude their occurrence in chron C29r strata14.
In the Bighorn Basin, elevated iridium anomalies were detected at four localities: Little Polecat Dome and Elk Basin, which are in the northern Bighorn Basin, and Kirby Ditch and Sand Draw, which are in the southern Bighorn Basin (Supplementary Fig. 1). The Little Polecat Dome locality (44.87430° N; 108.72791° W) is situated on the northeast side of Polecat Bench, ~13 km north-northeast of the town of Powell, along a road cut that passes across the contact between the Lance and Fort Union formations, which is defined by the first laterally persistent coal bed59,60,61. Approximately 70 m below the Lance-Fort Union contact is the Dumbbell Hill vertebrate-fossil locality, which yields a mammalian fauna ascribed to the latest Cretaceous Lancian North American Land-Mammal ‘age’ (NALMA)59,60,62, and the Lance Formation yields ceratopsian dinosaur fossils throughout the Little Polecat Dome exposures59,60,61. Approximately 0.75 km west-northwest of the Little Polecat Dome KPB section is the Mantua Quarry, a vertebrate-fossil locality that yields a mammalian fauna ascribed to the earliest Paleocene Puercan NALMA60, which is hosted at the base of the so-called ‘Mantua Lentil’59, an ~10–30-m-thick fluvial sandstone deposit that occurs near the base of the Fort Union Formation and sometimes immediately overlies or scours through the lowest Fort Union coal61. As such, the lowest coal of the Fort Union Formation at Little Polecat Dome has been called the ‘Mantua coal’ (Fig. 4). Given that Lancian mammals and ceratopsian dinosaurs are found in the Lance Formation below and Puercan mammals are found in the Fort Union Formation above, the ‘Mantua coal’ has long been considered to effectively mark the KPB59,60,61.
Elk Basin is a large, anticlinal structure that spans the Wyoming-Montana border. At its core are exposures of the Cody Shale, and along the rim are exposures of the Fort Union Formation; as such, Elk Basin captures much of the Upper Cretaceous, from the Santonian–Campanian to the latest Maastrichtian, and into the Paleocene63. The Elk Basin locality reported here (44.96885° N; 108.89359° W) is located ~2 km southwest of Silvertip Mine, up a narrow, roughly east–west-oriented coulee that cuts through the lower Fort Union and upper Lance formations. The lowest, formation-defining coal of the Fort Union Formation here is distorted via cm-scale soft-sediment deformation and intercalates with lenses of sand and mud varying from 10 cm to 1.5 m thick. Interpretations of the site are ongoing, but the lower ‘coaly’ portions below the KPB include chunks of carbonized wood and other organic debris that are not consistent with the typical, laminated coals of the Fort Union Formation. Near the top of the ‘coaly’ zone, it is no longer interdigitated with clastic sediments and more closely reflects a ‘typical’ Fort Union coal. As such, we interpret that upper coal to be the true base of the Fort Union Formation, but future work in the area is needed to understand the convolute bedding locally. Nonetheless, the ‘coaly’ interval is topped by an enormously thick channel-sandstone complex that is both morphologically and stratigraphically analogous to the Mantua Lentil at Little Polecat Dome. Paleontologically, non-avian dinosaur fossils are abundant in the underlying Lance Formation; however, to date, no Puercan-aspect mammals have been discovered (or at least reported) from the lower Fort Union Formation in Elk Basin (although Torrejonian mammals likely occur higher in the section of the Montana side of Elk Basin61).
Kirby Ditch is a small tributary of the Bighorn River that cuts through mostly lower Fort Union Formation sediments ~2 km west of the town of Kirby (accessible via Cowboy Mine Road) and is part of the larger, mapped feature referred to as Cedar Mountain. Moving down section from Kirby Ditch, approximately perpendicular to bedding dip, along Cowboy Mine Road ~0.5 km, the Kirby Ditch KPB locality is located ~60 m uphill (43.80703° N; 108.15554° W). There, the lowest coal of the Fort Union Formation is thin, and ~10 m uphill from it is an enormous channel-sandstone deposit that caps the hillside. The huge, lower Fort Union sandstone bed near Kirby Ditch has produced a Puercan-mammal assemblage known as the Leidy Quarry60,64, although we have not yet relocated that locality. As such, the large sandstone bed in the lower Fort Union of Cedar Mountain has been suggested to be coeval with the Mantua Lentil at Polecat Bench60. Other Puercan, Torrejonian, and Tiffanian mammal localities have been discovered in the larger Cedar Mountain area65.
Sand Draw refers to a stretch of badlands along Sand Draw Road, which extends between the town of Kirby to the east and Highway 120 just south of Grass Creek to the west63. Sand Draw Road runs more-or-less along the contact between the Meeteetse (south of the road) and Lance (north of the road) formations, and the Lance exposures to the north are capped by an enormous channel-sandstone deposit indicative of the lowermost Fort Union Formation (e.g., the Mantua Lentil). The Sand Draw KPB locality (43.84197° N; 108.34282° W) is approximately halfway along Sand Draw Road, ~14 km west-northwest of Kirby, and is located near the top of the bluff to the north of the road. There, the Mantua Lentil-like Fort Union sandstone bed cuts through the lowermost coal of the Fort Union Formation, including near where sediment samples were collected for iridium analysis (Supplementary Fig. 2). Ceratopsian dinosaur bones, teeth, and trace fossils (i.e., a coprolite) are abundant within the Lance Formation locally up to at least 5 m of the base of the Fort Union Formation. To date, no Puercan-mammal localities have been discovered in the lower Fort Union Formation at Sand Draw.
Mill Iron refers to an exposure of badlands straddling the Montana-South Dakota border; locally, most of the exposure, including the sampled interval (45.836470° N; 104.057126° W), is located on the Montana side. This area is best known for the nearby ‘Mill Iron’ Paleoindian site66, but the area also exposes the contact between the Hell Creek and Fort Union formations67, which is proposed as nearly coincident with the KPB based on circumstantial evidence68. This specific area has received little published geological or paleontological analysis to date, but the formational contact, as recognized by the lowest coal, is traceable laterally for hundreds of meters. The two formations in the area are also visually distinct at the outcrop scale (Fig. 1; Supplementary Fig. 3). We trenched and sampled a short section (~1.5 m) across the lowest coal, which revealed an ~1 cm thick pink claystone bed at the base of the coal (Supplementary Fig. 3).
To assess whether iridium anomalies indicative of the KPB were present in the Bighorn Basin, Wyoming, and at Mill Iron near Ekalaka, Montana, we targeted 0.5–1.5-m long sections spanning fromational contacts (Supplementary Fig. 2; Supplementary Fig. 3). In the Bighorn Basin, these were Lance-Fort Union contacts; at Mill Iron, this was the Hell Creek-Fort Union contact. To expose fresh rock, we dug ~0.25–0.5-m-deep trenches using plastic or wooden tools to avoid metal contamination, and bulk-sediment samples were taken at ~5–50-cm intervals across the formational contacts (note that wedding rings were also removed). Our sampling scheme primarily targeted reddish claystone units immediately underlying the lowest Fort Union Formation-defining coal and a series of fine-grained background facies underlying and/or overlying those units. Given this sample spacing, illustrated in Supplementary Fig. 2 and Supplementary Fig. 3, we cannot exclude the possibility that larger magnitude iridium anomalies are present stratigraphically between samples. Each sediment sample was filled into approximately half a gallon zip-lock bag. All Bighorn Basin samples were collected by LNW. Those from Little Polecat Dome were collected in 2022, Elk Basin were collected in 2023, Kirby Ditch and Sand Draw were collected in 2024. Mill Iron samples were collected by TST in 2024.
We sent bulk-sediment samples to Activation Laboratories Ltd. (Actlabs; Ancaster, Ontario, Canada) to measure trace amounts (ppb) of iridium and other platinum group elements using Instrumental Neutron Activation Analysis (INAA). Specifically, these samples were analyzed using the ‘5S – Short-Lived Isotopes – INAA’. According to the Actlabs website (https://actlabs.com/geochemistry/exploration-geochemistry/inaa/): ‘1 g samples are weighed into medium poly vials and are irradiated at a thermal flux of 7 × 1012 n cm−2 s−1 in a computer-controlled rabbit system, which sends the INAA vials directly to the source of irradiation, then back to the fume hood in the lab for analysis. After appropriate decay times to allow aluminum and other very short-lived isotopes to decay, the samples are counted sequentially for the analyte(s) of interest. Values are corrected for decay and compared to a standard calibration. Two standards are analyzed with every work order. Duplicates are analyzed when samples are provided.’ For a full report on these analyses, see Supplementary Table 2.
Our interpretations of KPB facies shifts come primarily from direct field observations and sedimentological data from the Hell Creek area of northeastern Montana (Williston Basin) and the Bighorn Basin of northwestern Wyoming. The bulk of our Hell Creek data was collected during the 2017–2021 field seasons, and the Bighorn Basin data were collected during the 2022–2024 field seasons. All of the sedimentological and stratigraphic features described here were identified in the field via trenching, hand sampling, and detailed description of lithology. Brunton compasses were used to measure paleoflow directions from trough cross-beds or ripple marks (preserved in well-indurated sandstone beds) and the strike and dip of inclined lateral-accretion surfaces. Coarser surveying techniques were supplemented by trenching, logging, and measuring surface sections to further characterize sedimentary sequences. Although limited in geographic scope, our interpretations are consistent with other sedimentological and stratigraphic data reported by previous studies across the Williston Basin12,15,17,18,19.
In the Hell Creek area, we examined nearly every reported, impact-identified KPB section and the sedimentology and stratigraphy of the surrounding areas, including: Iridium Hill, Brownie Butte, Lerbekmo (also known as the Hell Creek Road locality), Herpijunk Promontory, and Billy Creek (Supplementary Table 2). Studies of these KPB sections were supplemented by studies of Hell Creek and Fort Union sedimentology elsewhere in Garfield and McCone counties. See for example: Sprain et al.14,69, Fendley et al.70, Tobin et al.25, Wilson et al.71, Weaver et al.24, Claytor et al.26, and Wilson Deibel et al.40.
In the Bighorn Basin, Lance and Fort Union sedimentology were assessed in all of the KPB localities described above; however, the bulk of our sedimentological and stratigraphic data to date derive from Little Polecat Dome and Elk Basin. Nonetheless, the patterns in facies change studied in the northern Bighorn Basin are consistent with those observed in the southern Bighorn Basin.
The chronostratigraphic diagrams in Supplementary Fig. 5 were assembled using data from Sprain et al.14, Johnson et al.15, Rogers et al.29, Peppe et al.32,72, Hartman and Kihm73, Catuneanu and Sweet74, Murphy et al.75, and Secord et al.76.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Data from Supplementary Tables 1 and 2 can be found on Dryad: https://doi.org/10.5061/dryad.j9kd51crj.
References
Goderis, S. et al. Globally distributed iridium layer preserved within the Chicxulub impact structure. Sci. Adv. 7, eabe3647 (2021).
Fischer-Gödde, M. et al. Ruthenium isotopes show the Chicxulub impactor was a carbonaceous-type asteroid. Science 385, 752–756 (2024).
Archibald, J. D. A study of Mammalia and geology across the Cretaceous-Tertiary boundary in Garfield County. Mont. Univ. Calif. Publ. Geol. Sci. 122, 1–286 (1982).
Archibald, J. D. et al. Cretaceous extinctions: multiple causes. Science 328, 973 (2010).
Wilson, G. P. Mammalian extinction, survival, and recovery dynamics across the Cretaceous-Paleogene boundary in northeastern Montana, USA. In Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas (eds Wilson, G. P., Clemens, W. A., Horner, J. R. & Hartman, J. H.). Geol. Soc. Amer. Spec. Pap. 503, 365–392 (2014).
Bralower, T. J. et al. Origin of a global carbonate layer deposited in the aftermath of the Cretaceous-Paleogene boundary impact. Earth Planet. Sci. Lett. 548, 116476 (2020).
Pringle, R. M. et al. Impacts of large herbivores on terrestrial ecosystems. Curr. Biol. 33, R584–R610 (2023).
Smith, F. A. et al. After the mammoths: the ecological legacy of late Pleistocene megafauna extinctions. Camb. Prism Extinct. 1, 1–23 (2023).
Erwin, D. H. Macroevolutionary dynamics of ecosystem-engineering and niche construction. Palaeontol 67, e12718 (2024).
Charlton, R. Fundamentals of Fluvial Geomorphology (1st ed.). (Routledge, London, 2007).
Gurnell, A. Plants as river system engineers. Earth Surf. Process. Landf. 39, 4–25 (2014).
Nichols, D. J. & Johnson, K. R. Plants and the K-T boundary (Cambridge University Press, Cambridge, 2008).
Renne, P. R. et al. State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact. Science 350, 76–78 (2015).
Sprain, C. J., Renne, P. R., Clemens, W. A. & Wilson, G. P. Calibration of chron C29r: New high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana. Geol. Soc. Am. Bull. 130, 1615–1644 (2018).
Johnson, K. R., Nichols, D. J. & Hartman, J. H. Hell Creek Formation: a 2001 synthesis. In The Hell Creek Formation and the Cretaceous-Tertiary Boundary in the Northern Great Plains: An Integrated Continental Record of the End Cretaceous (eds Hartman, J. H., Johnson, K. R. & Nichols, D. J.). Geol. Soc. Am. Spec. Pap. 361, 503–510 (2002).
Fastovsky, D. E. Paleoenvironments of vertebrate-bearing strata during the Cretaceous-Paleogene transition, eastern Montana and western North Dakota. Palaios 2, 282–295 (1987).
Sweet, A. R. & Braman, D. R. The K-T boundary and contiguous strata in western Canada: interactions between paleoenvironments and palynological assemblages. Cretac. Res. 13, 31–79 (1992).
Wiest, L. A., Lukens, W. E., Driese, S. G., Peppe, D. J. & Forman, S. L. Landscape evolution across the Cretaceous/Paleogene boundary in southwestern North Dakota. USA Cretac. Res. 112, 104470 (2020).
Fastovsky, D. E. & Bercovici, A. The Hell Creek Formation and its contribution to the Cretaceous–Paleogene extinction: a short primer. Cretac. Res. 57, 368–390 (2016).
Fastovsky, D. E. & McSweeney, K. Paleosols spanning the Cretaceous-Paleogene transition, eastern Montana and western North Dakota. Geol. Soc. Am. Bull. 99, 66–77 (1987).
Flores, R. M. & Pillmore, C. L. Tectonic control on alluvial paleoarchitecture of the Cretaceous and Tertiary Raton Basin, Colorado and New Mexico. In Recent Developments in Fluvial Sedimentology (eds Ethridge, F. G., Flores, R. M. & Harvey, M. D.). SEPM Spec. Pub. 39, 311–320 (1987).
Vajda, V. & Bercovici, A. The global vegetation pattern across the Cretaceous–Paleogene mass extinction interval: a template for other extinction events. Glob. Planet. Change 122, 29–49 (2014).
Weaver, L. N., Kelson, J. R., Holder, R. M., Niemi, N. A. & Badgley, C. On the role of tectonics in stimulating the Cretaceous diversification of mammals. Earth Sci. Rev. 248, 104630 (2024).
Weaver, L. N. et al. Revised stratigraphic relationships within the lower Fort Union Formation (Tullock Member, Garfield County, Montana, USA) provide a new framework for examining post K–Pg mammalian recovery dynamics. Palaios 37, 104–127 (2022).
Tobin, T. S. et al. Analyzing sources of uncertainty in terrestrial organic carbon isotope data: a case study across the K-Pg boundary in Montana, USA. Palaeogeog. Palaeoclimatol. Palaeoecol. 574, 110451 (2021).
Claytor, J. R., Weaver, L. N., Tobin, T. S. & Wilson Mantilla, G. P. New mammalian local faunas from the first ca. 80 ka of the Paleocene in northeastern Montana and a revised model of biotic recovery from the Cretaceous–Paleogene mass extinction. J. Vertebr. Paleontol. 42, e2222777 (2022).
Thomas, R. G. et al. Inclined heterolithic stratification—terminology, description, interpretation and significance. Sediment. Geol. 53, 123–179 (1987).
Korasidis, V. A., Wing, S. L., Nelson, D. M. & Baczynski, A. A. Reworked pollen reduces apparent floral change during the Paleocene-Eocene thermal maximum. Geology 50, 1398–1402 (2022).
Rogers, R. R. et al. Age, correlation, and lithostratigraphic revision of the Upper Cretaceous (Campanian) Judith River Formation in its type area (north-central Montana), with a comparison of low-and high-accommodation alluvial records. J. Geol. 124, 99–135 (2016).
Holland, S. M. The structure of the nonmarine fossil record: predictions from a coupled stratigraphic–paleoecological model of a coastal basin. Paleobiology 48, 372–396 (2022).
Catuneanu, O. & Sweet, A. R. Maastrichtian–Paleocene foreland-basin stratigraphies, western Canada: a reciprocal sequence architecture. Can. J. Earth Sci. 36, 685–703 (1999).
Peppe, D. J., Johnson, K. R. & Evans, D. A. Magnetostratigraphy of the Lebo and Tongue River Members of the Fort Union Formation (Paleocene) in the northeastern Powder River Basin, Montana. Am. J. Sci. 311, 813–850 (2011).
Peters, S. E. Environmental determinants of extinction selectivity in the fossil record. Nature 454, 626–629 (2008).
Copeland, P., Currie, C. A., Lawton, T. F. & Murphy, M. A. Location, location, location: the variable lifespan of the Laramide orogeny. Geol 45, 223–226 (2017).
Kring, D. A. & Durda, D. D. Trajectories and distribution of material ejected from the Chicxulub impact crater: Implications for postimpact wildfires. J. Geophys. Res. 107, 6-1–6-22 (2002).
Albertson, L. K. et al. The ghosts of ecosystem engineers: legacy effects of biogenic modifications. Func. Ecol. 38, 52–72 (2022).
Benson, R. B. et al. Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage. PLoS Biol. 12, e1001853 (2014).
Rogers, R. R. Taphonomy of three dinosaur bone beds in the upper cretaceous two medicine formation of northwestern montana: evidence for drought-related mortality. Palaios 5, 394–413 (1990).
Carvalho, M. R. et al. Extinction at the end-Cretaceous and the origin of modern Neotropical rainforests. Science 372, 63–68 (2021).
Jones, L. S. & Gustason, E. R. Dinosaurs as possible avulsion enablers in the Upper Jurassic Morrison Formation, east-central Utah. Ichnos 13, 31–41 (2006).
Chiarenza, A. A. et al. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 117, 17084–17093 (2020).
Condamine, F. L., Guinot, G., Benton, M. J. & Currie, P. J. Dinosaur biodiversity declined well before the asteroid impact, influenced by ecological and environmental pressures. Nat. Comm. 12, 3833 (2021).
Lyson, T. R. et al. Dinosaur extinction: closing the ‘3 m gap’. Biol. Lett. 7, 925–928 (2011).
Lyson, T. R. et al. Exceptional continental record of biotic recovery after the Cretaceous–Paleogene mass extinction. Science 366, 977–983 (2019).
Alroy, J. The fossil record of North American mammals: evidence for a Paleocene evolutionary radiation. Syst. Biol. 48, 107–118 (1999).
Wilson Deibel, P. K., Wilson Mantilla, G. P. & Strömberg, C. A. Plant taxonomic turnover and diversity across the Cretaceous/Paleogene boundary in northeastern Montana. Paleobiol. 50, 1–19 (2024).
Wilf, P., Carvalho, M. R. & Stiles, E. The end-Cretaceous plant extinction: heterogeneity, ecosystem transformation, and insights for the future. Camb. Prism Extinct. 1, e14 (2023).
Jerzykiewicz, T. & Sweet, A. R. Sedimentological and palynological evidence of regional climatic changes in the Campanian to Paleocene sediments of the Rocky Mountain Foothills, Canada. Sediment. Geol. 59, 29–76 (1988).
Schwartz, T. M., Dechesne, M. & Zellman, K. L. Evidence for variable precipitation and discharge from Upper Cretaceous–Paleogene fluvial deposits of the Raton Basin, Colorado–New Mexico, USA. J. Sediment. Res. 91, 571–594 (2021).
Brazier, R. E. et al. Beaver: nature’s ecosystem engineers. WIREs Water 8, e1494 (2021).
Gearon, J. H. et al. Rules of river avulsion change downstream. Nature 634, 91–95 (2024).
Dunn, R. E., Strömberg, C. A., Madden, R. H., Kohn, M. J. & Carlini, A. A. Linked canopy, climate, and faunal change in the Cenozoic of Patagonia. Science 347, 258–261 (2015).
Wing, S. L. & Boucher, L. D. Ecological aspects of the Cretaceous flowering plant radiation. Ann. Rev. Earth Planet. Sci. 26, 379–421 (1998).
Wing, S. L. & Tiffney, B. H. The reciprocal interaction of angiosperm evolution and tetrapod herbivory. Rev. Palaeobot. Palynol. 50, 179–210 (1987).
Eriksson, O., Friis, E. M. & Löfgren, P. Seed size, fruit size, and dispersal systems in angiosperms from the Early Cretaceous to the Late Tertiary. Am. Nat. 156, 47–58 (2000).
Doughty, C. E. et al. Ecosystem engineers alter the evolution of seed size by impacting fertility and the understory light environment. Palaeontol 68, e70002 (2025).
Nations, J. A., Weaver, L. N. & Grossnickle, D. M. Locomotor trends among early mammals illuminated by predictors of arboreality from the mammalian appendicular skeleton. bioRxiv https://doi.org/10.1101/2025.07.09.663922 (2025).
Chen, M., Strömberg, C. A. & Wilson, G. P. Assembly of modern mammal community structure driven by Late Cretaceous dental evolution, rise of flowering plants, and dinosaur demise. Proc. Natl. Acad. Sci. 116, 9931–9940 (2019).
Jepsen, G. L. Paleocene faunas of the Polecat Bench Formation, Park County, Wyoming: part I. Proc. Am. Phil. Soc. 83, 217–340 (1940).
Gingerich, P. D., Rose, K. D. & Krause, D. W. Early Cenozoic mammalian faunas of the Clark’s Fork Basin-Polecat Bench area, northwestern Wyoming. In Early Cenozoic Paleontology and Stratigraphy of the Bighorn Basin, Wyoming (ed. Gingerich, P. D.). Univ. Mich. Mus. Paleontol. Pap. Paleontol. 24, 51–68 (1980).
Hickey, L. J. Paleocene stratigraphy and flora of the Clark’s Fork Basin. In Early Cenozoic Paleontology and Stratigraphy of the Bighorn Basin, Wyoming (ed. Gingerich, P. D.). Univ. Mich. Mus. Paleontol. Pap. Paleontol. 24, 33–49 (1980).
Cifelli, R. L., Eberle, J. J., Lofgren, D. L., Lillegraven, J. A. & Clemens, W. A. Mammalian biochronology of the latest Cretaceous. In Late Cretaceous and Cenozoic mammals of North America: biostratigraphy and geochronology (ed. Woodburne, M. O.). Columbia University Press, New York. 21–42 (2004).
Johnson, R. C., Keefer, W. R., Keighin, C. W. and Finn, T. M. Detailed outcrop studies of the upper part of the Upper Cretaceous Cody Shale and the Upper Cretaceous Mesaverde, Meeteetse, and Lance formations, Bighorn Basin, Wyoming. Cretaceous and Lower Tertiary Rocks of the Bighorn Basin, Wyoming and Montana: 49th Annual Field Conference Guidebook. 59–78 (1998).
Van Valen, L. The beginning of the age of mammals. Evol. Theory 4, 46–80 (1978).
Hartman, J. E. Paleontology and biostratigraphy of lower part of Polecat Bench Formation, southern Bighorn Basin, Wyoming. Contrib. Geol. Univ. Wyo. 24, 11–63 (1986).
Waters, M. R. & Stafford, T. W. Jr Redating the Mill Iron Site, Montana: a reexamination of Goshen complex chronology. Am. Antiq. 79, 541–548 (2014).
Vuke, S. M., Heffern, R. L., Bergantino, R. N., & Colton, R. B. Geologic map of the Powderville 30’ x 60’ quadrangle, eastern Montana. Montana Bureau of Mines & Geology, (2001).
Schwartz, J. M. Investigation of a Possible Cretaceous-Paleogene Boundary Layer in the Hell Creek Formation near Ekalaka, Montana, USA. Master’s thesis, Northern Illinois University 2021).
Sprain, C. J., Renne, P. R., Wilson, G. P. & Clemens, W. A. High-resolution chronostratigraphy of the terrestrial Cretaceous-Paleogene transition and recovery interval in the Hell Creek region, Montana. Geol. Soc. Am. Bullet. 127, 393–409 (2015).
Fendley, I. M. et al. Constraints on the volume and rate of Deccan Traps flood basalt eruptions using a combination of high-resolution terrestrial mercury records and geochemical box models. Earth Planet. Sci. Lett. 524, 115721 (2019).
Wilson, P. K., Wilson Mantilla, G. P. & Strömberg, C. A. Seafood salad: a diverse latest Cretaceous flora from eastern Montana. Cretac. Res. 121, 104734 (2021).
Peppe, D. J., Evans, D. A. D. & Smirnov, A. V. Magnetostratigraphy of the Ludlow Member of the Fort Union Formation (Lower Paleocene) in the Williston Basin, North Dakota. Geol. Soc. Am. Bullet. 121, 65–79 (2009).
Hartman, J. H. & Kihm, A. J. Bio-and magnetostratigraphy of the uppermost Cretaceous and lower Tertiary strata of North Dakota. Paleontologic. Soc. Spec. Pub. 8, 163–163 (1996).
Catuneanu, O. & Sweet, A. R. Maastrichtian-Paleocene foreland-basin stratigraphies, western Canada: a reciprocal sequence architecture. Can. J. Earth Sci. 36, 685–703 (1999).
Murphy, E. C., Hoganson, J. W., & Johnson, K. R. Lithostratigraphy of the Hell Creek Formation in North Dakota. In The Hell Creek Formation and the Cretaceous-Tertiary Boundary in the Northern Great Plains: An Integrated Continental Record of the End Cretaceous (eds Hartman, J. H., Johnson, K. R. & Nichols, D. J.). Geol. Soc. Amer. Spec. Pap. 361, 9–34 (2002).
Secord, R. et al. Geochronology and mammalian biostratigraphy of middle and upper Paleocene continental strata, Bighorn Basin, Wyoming. Am. J. Sci. 306, 211–245 (2006).
Acknowledgements
We are grateful to the following for discussions that improved this manuscript: G.P. Wilson Mantilla, A. Licht, T.R. Lyson, T. Jerzykiewicz, C. Badgley, P.D. Gingerich, W. A. Clemens, R.R. Rogers, R.E. Dunn, R.S. Barclay, S.L. Wing, E.D. Currano, J.N. Milligan, B.Z. Foreman, A.J. Lowe, H.Z. Fulghum, B.T. Hovatter, J.A. Wilson Mantilla, and R.E. Sweedler. This manuscript was substantially improved thanks to reviews by K.R. Johnson, M. Gibling, and M.J. Benton. Fieldwork in the Bighorn Basin was made possible by T. Churchill, P.D. Gingerich, B.H. Smith, H.Z. Fulghum, Cretaceous Bighorn Project field teams, A. Truman and the Ten Sleep Brewery staff, the U.S. Bureau of Land Management for providing paleontological permits, and B. Breithaupt for facilitating those permits. Fieldwork in Montana was made possible by Hell Creek Land Owners, especially the Engdahl, Tharp, Twitchell, and Thomas families, Hell Creek Bar staff, Hell Creek Project field teams, the U.S. Bureau of Land Management and the Montana State Department of Natural Resources and Conservation who provided paleontological permits, and G. Liggett, D. Melton, and P. Rennie for facilitating those permits. Fieldwork in the Raton Basin was made possible for P.K. by P. Claeys, C.B. Senel, and J. Vellekoop. Financial support was provided by the National Science Foundation (Graduate Research Fellowship and EAR-PF 2052992 to L.N.W.; EAR-2321343 to T.S.T.; Award 2321344 to C.J.S.; Graduate Research Fellowship to I.M.F.) and David B. Jones Foundation (L.N.W. and T.S.T.).
Author information
Authors and Affiliations
Contributions
L.N.W., T.S.T., and C.J.S. conceived of the study. L.N.W. and T.S.T. collected samples for iridium analysis. L.N.W., T.S.T., C.J.S., P.K.W.D., I.M.F., and P.K. contributed sedimentological and stratigraphic data and interpretations. P.K.W.D., V.A.K., and M.R.C. contributed paleobotanical expertise. L.N.W. wrote the manuscript and created the figures and tables with substantial input from all coauthors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Kirk R. Johnson, Martin R. Gibling and Michael J. Benton for their contribution to the peer review of this work. Primary Handling Editors: Carolina Ortiz Guerrero and Aliénor Lavergne. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Weaver, L.N., Tobin, T.S., Sprain, C.J. et al. Dinosaur extinction can explain continental facies shifts at the Cretaceous-Paleogene boundary. Commun Earth Environ 6, 712 (2025). https://doi.org/10.1038/s43247-025-02673-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-025-02673-8
