
Abstract
Submillimeter-scale redox archives in organic-rich mudrocks elucidate dynamic environmental stress shaping early marine animal ecosystems. During the late Cambrian, benthic animals, including trilobites, thrived in intermittently oxygen-deficient (predominantly euxinic) outer shelf environment where the astronomically-tuned Scandinavian Alum Shale was deposited. We present ultrahigh-resolution (0.25 mm) molybdenum and uranium isotope (δ98Mo-δ238U) and concentration profiles, revealing millennial-scale redox fluctuations that intermittently disrupted bottom-water euxinia. Varying ocean circulation, redox buffer supplies, and/or reduced productivity likely created short-lived aerobic niches for opportunistic benthic animals. Distinct δ98Mo-δ238U patterns indicate recurrent chemocline shoaling with toxic hydrogen sulfide incursions into surface waters, resembling metal drawdown in modern restricted basins. These fluctuations punctuated a million-year-long global oceanic anoxic event linked to the Steptoean Positive Carbon Isotope Excursion. Following this excursion, local redox conditions oscillated around a globally more oxygenated baseline, coinciding with animal diversifications in Baltica and worldwide. These millennial-scale redox changes underscore the need for higher-resolution geochemical approaches in paleoenvironmental reconstructions.
Similar content being viewed by others

Introduction
Episodes of organic-rich shale deposition occurred throughout Earth’s history1,2,3,4,5,6 with a notable abundance in the early Paleozoic when anoxic marine zones were more widespread and atmospheric pO2 likely was lower than today7,8. Organic-rich shales are primarily deposited in oxygen-poor environments, either under euxinic conditions where hydrogen sulfide (H2S) is present in the anoxic bottom waters, under ferruginous conditions (rare in the modern ocean) where anoxic bottom waters are sulfur-limited, or under low-oxygen conditions, where the bottom water O2 levels are significantly lower than in the surface ocean9,10,11,12,13,14. Redox fluctuations at the seafloor are expected to exert first-order control on habitability and perhaps also on biotic evolution, since animals are obligate aerobic organisms and H2S is toxic to essentially all eukaryotes. The redox conditions near the seafloor fluctuate because marine oxygen supply and consumption vary over a wide range of timescales from seasonal to millennial to millions of years12,15,16,17. Yet, rapidly changing redox conditions in the oceans challenge the interpretation of sedimentary paleoredox proxies extracted from bulk rock samples that may aggregate time over thousands to millions of years, highlighting the need for obtaining records at finer stratigraphic resolution with an accurately tuned time scale.
The late Cambrian Alum Shale in Scandinavia was deposited under overall euxinic conditions and, perhaps occasionally, ferruginous conditions10,18,19,20. However, like many other anoxic shales in the world21,22, the Alum Shale also bears evidence of benthic animal activity, including abundant trilobite communities23,24,25,26 (Fig. 1), indicative of periods with aerobic conditions at the seafloor. The Alum Shale offers opportunities to investigate in detail how redox conditions fluctuated at millennial time scales in the Cambrian ocean. Episodic oxygenation events have been suggested to have opened niches for bottom-dwelling animals15. Independent evidence for millennial oxygenation events in the Alum Shale Sea was inferred from recurrent decreases in sedimentary molybdenum (Mo) concentrations from well above 100 ppm to below 25 ppm detected at sub-millimeter stratigraphic resolution15 (0.2 mm; Fig. 1C). In addition, we here cut samples at sub-millimeter resolution and use isotopic proxies to advance previous efforts and improve reconstructions of the processes governing redox dynamics at the millennial scale.

A Stratigraphy, lithology, and fossils. The detailed biozone framework is established based on existing fossil records and further constrained by gamma log correlation with other fossiliferous Alum Shale cores36,76,77. Biozone abbreviations: Pr Protopeltura, Le Leptoplastus. B Photos of fossils found in the Alum Shale Formation, e.g., Olenus transversus trilobites (top) from the lower part of the Olenus Superzone, brachiopods (middle), and cross-section of Orusia lenticularis (bottom) from the Parabolina Superzone15,36. C High-resolution (0.2 mm) Mo concentration profile with stratigraphic levels of ten MREs investigated in this study. The gray dots and green curve represent XRF Mo concentrations calibrated against ICP-MS data15 and a LOESS fitted curve, respectively. D Organic carbon isotope profile across the SPICE event in the Billegrav-2 core76. E Million-year-scale δ98Mo and δ238U data from hand-size Alum Shale specimens across the SPICE event10.
Molybdenum and uranium concentrations are powerful paleoredox proxies, and so are their stable isotope compositions, δ98Mo and δ238U. Both elements occur as dissolved anionic compounds with a conservative behavior in oxic seawater, which is reflected in their long residence times in the Cambrian (~100 kyr)10,27,28 and modern ocean (~400 kyr)29,30. Both elements are preferentially enriched in sediments deposited under reducing bottom water conditions31. However, the link between Mo and U concentrations and the local redox conditions that control their removal is not straightforward, since muted enrichments can also occur in basins with limited supply of these metals. For example, the seawater supply of Mo and U from the open ocean might be outpaced by rapid removal into anoxic and sulfidic sediments within a basin. Additionally, the inventories of Mo and U in the open ocean would have been lower at times when marine anoxia was globally more widespread10,20. Paired δ98Mo and δ238U analyses in sediments have opened prospects of discriminating between these scenarios10,32,33,34; however, to our knowledge, they have not yet been investigated at the submillimeter scale. Here, we re-examined ten “Millennial Redox Events” (MRE, consisting of both oxygenation or deoxygenation scenarios) previously described as “Brief Oxygenation Events” from the Billegrav-2 Alum Shale core obtained on Bornholm, Denmark15 (see location in Supplementary Fig. 1), by applying paired U and Mo concentrations and stable isotope analyses at the submillimeter scale (as low as 0.25 mm stratigraphic resolution). The Billegrav-2 core has been chronologically dated using cyclostratigraphy35 anchored in the geological record36, providing interpolated estimates for the average durations of the MREs spanning many centuries to several millennia (~650–8000 yrs). These durations are interpreted by interpolating 405 kyr-eccentricity cycles over approximately 80 cm of stratigraphy, but the events may be significantly shorter, particularly if extended periods of non-deposition were dominant.
Molybdenum and uranium isotopes
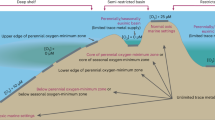
The long residence times of U and Mo in the Cambrian ocean are much longer than the ocean mixing time of ∼1.5 kyr37, implying that oceans are well mixed with respect to Mo and U and that significant changes in the global ocean inventories of these elements occur on timescales of over ~105 yrs. Therefore, millennial-scale changes in Mo and U concentrations and their isotope compositions (here, expressed over millimeters of stratigraphy) in sedimentary rocks must reflect changes in the local environment rather than changes in global seawater composition or metal inventories. In euxinic basins, Mo and U are efficiently scavenged and buried18,38,39, whereas burial rates are significantly lower in suboxic environments, where sulfide accumulates at depth inside the sediments. Figure 2 outlines three conceptually distinct MRE scenarios and how the sedimentary δ98Mo, δ238U, Mo, and U concentrations would be expected to change when the supply or burial rates vary in the basin.

The zones in green, orange, and purple represent oxic, anoxic (where Fe(III), Mn(IV), and nitrate reduction dominates), and sulfidic (where sulfate reduction prevails) zones, respectively. Black solid and dashed arrow lines represent relative more or less Mo and U fluxes to the sediment, respectively. SWI sediment-water interface, SW sea water. A During a Millenial Oxygenation Event in the water column (MOEWC), Mo burial on settling partiles will be dramatically reduced and lower δ98Mo values in the sediments (greater Mo isotope offset from contemporaneous seawater) are expected, whereas most U burial likely occurs below sediment-water interface where conditions are permanently sulfidic, resulting in smaller changes. B During a more extensive oxygenation event that also introduces O2 into the sediments (MOESED), U and δ238U willl also be affected. The U isotope offset in oxic settings is smaller than under euxinic conditions, which leads to lower δ238U closer to crustal composition and oceanic input. C During a Millenial Deoxygenation Event (MDE) where H2S levels in the water column rises and cause removal of Mo and U via settling particles to the sediments, which will draw down the Mo and U inventory in the overlying water column (given slow replenishment) resulting in low sedimentary Mo and U concentrations and sedimentary δ98Mo and δ238U compositions approaching seawater values.
As a first scenario, a redox-stratified euxinic basin interrupted by periodic millennial-scale bottom water oxygenation or vanishing H2S levels in the water column (MOEWC; Fig. 2A) will slow Mo sequestration from the water column into the sediments. In anoxic and euxinic settings, dissolved Mo (MoO4S2-) reacts with H2S to form thiomolybdates (MoO4-xS2-, x = 1–4), which are removed through precipitation with sinking iron sulfides and organic matter38,40,41. Each step of the conversion from molybdate towards tetrathiomolybdates is associated with stable isotope fractionation18,42,43. When H2S level exceeds 11 µM, Mo will be quantitatively removed via particle scavenging into the sediments44, producing sedimentary δ98Mo compositions approaching that of the overlying seawater45,46,47. However, in weakly euxinic or intermittently oxygenated water columns (H2S < 11 µM or fluctuating), Mo removal can be incomplete, yielding a negative δ98Mo offset in sediments relative to seawater, because the particle reactive Mo fraction in the basin consists of isotopically fractionated thiomolybdate48. Unlike Mo, uranium sequestration begins at around the Fe(II)–Fe(III) redox boundary in the absence of H2S33,49,50 and continues through the sulfidic zone. Additionally, reductive U sequestration typically occurs in the uppermost few centimeters of the sediments where soluble U(VI) from seawater diffuses across the sediment-water interface51, whereas Mo removal mainly occurs at the O2/H2S chemocline. Therefore, the MOEWC scenario could leave the U concentration and δ238U offset relative to overlying seawater largely unaffected as long as the pore fluids remained anoxic and sulfidic at depth inside the sediments. Given the differences, the coupling using Mo and U elemental abundances and isotopes has the potential to more precisely bracket the oceanic redox conditions.
In a second and more extensive oxygenation scenario, where the sediment porewaters also become oxygenated to a significant depth (MOESED; Fig. 2B), we expect sedimentary U and δ238U to change. With oxygenated porewaters, lower U-enrichments and lower δ238U are expected, as seen in modern sediments deposited under oxic waters with sulfide accumulating at depth51,52. In this scenario, the δ238U composition of the sediments will typically be closer to coeval seawater with a smaller isotopic offset than observed in euxinic settings (~0.6–0.8‰33,53). If Mo and U were removed via manganese oxides during MOESED events, this would also be accompanied by lower δ98Mo values and lower δ238U values54,55.
As a third and quite opposite scenario, we propose a Millennial Deoxygenation Event (MDE; Fig. 2C), where Mo and U are removed even faster and more completely in the basin relative to the basinal supply from the open ocean (e.g., declining deepwater renewal rate). This rapid removal would eventually lower the sedimentary Mo and U concentrations. As stated above, the oceanic Mo and U inventories cannot decrease significantly over such short time scales. During millennial-scale intensification of euxinia, elevated H2S concentrations and/or shoaling of the O2/H2S chemocline in the water column would enhance the formation of thiomolybdates, facilitating Mo removal via particle settling into the sediments38,40. Rising levels of aqueous H2S would, in general, imply that H2S also migrated towards the ocean surface, i.e., chemocline shoaling. At high enough [H2S] (>11 μM), quantitative Mo removal produces sediments with higher δ98Mo compositions close to that of the seawater source (i.e., open ocean) in a fashion similar to the trend observed with basinal restriction within euxinic basins today45,47. In the MDE scenario, we also expect the U isotopic offset between the water column and the sediment to diminish and the sedimentary δ238U to approach the composition of seawater input to the basin (Fig. 2C). Thus, the MDE is expected to produce a negative correlation of δ238U and δ98Mo, as observed in modern basins with sluggish ventilation33.
Results and discussion
A total of 127 Alum Shale samples from ten MREs of the Billegrav-2 core display U, δ238U, Mo, and δ98Mo values in ranges of 17–179 ppm, −0.27–0.30‰, 6–302 ppm, and 0.47–1.83‰, respectively (Supplementary Table 1). Five representative MREs are shown in Fig. 3 and described in the following text, and the remaining five MREs are discussed in the Supplementary Note 3. All ten MREs are classified as either MOEWC or MDE; no MOESED events were identified. For C10E14, we cut an additional sample with a direction perpendicular to the bedding plane, which could represent a bulk rock sample of this event. This sample contains Mo and U concentrations and δ98Mo and δ238U compositions of 59 ppm, 61 ppm, 0.78 ± 0.06‰ and −0.05 ± 0.07‰, respectively. The sub-millimeter bedding-parallel samples demonstrate significant variation of these proxies across the same stratigraphic range, with ranges of 6–76 ppm, 53–78 ppm, 0.75–1.35‰, and −0.24 to −0.04‰ (n = 10). This comparison demonstrates the substantial risk of missing short-term signals when using bulk rock analyses.

A Stratigraphy, lithology, and carbon isotopes of the Alum Shale Formation in the Billegrav-2 core. B Global genus-level diversity and relative diversification rate74,75. C Species diversity in the Alum Shale Formation, see data in Supplementary Table 2. D–G Abundances and isotope compositions of U and Mo at sub-millimeter resolution in the Alum Shale for five representative MREs. The geochemical profiles of the remaining five events are provided in Supplementary Fig. 2. Interpolated sedimentation rates (SR) are from Sørensen et al.35.
Bottom water oxygenation at millennial scale (MOEWC) and its drivers
The data patterns observed in C1E5, C1E6, and C9E6 are consistent with the expected characteristics of the MOEWC scenario (Figs. 2A and 3DE). These events contain δ98Mo decreases across stratigraphic thicknesses of 1.3–4.5 mm (i.e., lasting 650–2250 yrs; calculated based on ref. 35, which all correspond with a drop in Mo concentrations (from >170 to <25 ppm). Although we find evidence of increasing Al concentration that serves as a proxy for clastic inputs, the decreasing Mo concentrations cannot be explained only by sediment dilution due to changes in sedimentation rate, since the change of Al concentration is modest (Supplementary Fig. 3). Also, the Alum Shale mud was deposited exceedingly slowly (~1–4 mm kyr−1 post-compaction35,36), and even euxinic sediments deposited ~104 times faster also capture high Mo concentrations14,56. Additionally, U and Mo concentrations are exceptionally higher than those of upper continental crust (2.7 ppm U and 1.1 ppm Mo57), suggesting that the signals are authigenic rather than detrital in origin. Unlike the Mo-based proxies, the sedimentary δ238U values vary by only ~0.1–0.2‰ within a single MOEWC event (Fig. 3). Pyrite is present throughout the studied intervals15, thus we can infer that the sedimentary porewaters were persistently anoxic and sulfidic at depth with reductive U removal as the dominant U removal pathway taking place inside the sediments at all times. Such a setting is also observed in parts of the modern Baltic Sea58, where episodic inflows of oxygenated water are producing geochemical signatures similar to those in the Alum Shale.
The drivers for the MOEWC events could be changes in ocean circulation, redox buffer supplies, and/or biological productivity. A recent modeling study showed that the continental configuration in the late Cambrian could have permitted oscillations at millennial scales between two globally distinct ocean circulation regimes with dramatic consequences for benthic O2 levels worldwide (ranging between >100 μmol kg−1 and <45 μmol kg−1 in the abyssal oceans even if atmospheric pO2 were at modern-day levels59). Atmospheric pO2 levels were likely lower in the Cambrian7,28 with the surface oceans containing likely only ~10–40% of today’s maximum O2 concentration. Therefore, we envisage variations in the flux of oxygenated water into the Alum Shale basin, paced by shifts in ocean circulation, could have driven frequent switching between anoxic and oxic bottom water conditions in the region. Still, local sediment porewaters remained sulfidic at depth as evidenced by continuous pyrite formation at all times.
The Cambrian oceans were characterized by lower concentrations of redox buffers such as H2S and Fe2+, making millennial redox conditions sensitive to minor variations in the sulfide and iron contents at the basin scale. This inference is supported by a robust positive correlation (R2 = 0.84, n = 33) between Mo concentrations and the degree of pyritisation (DOPT) proxy [=(S/64)/(Fe/56), with lower DOPT values indicating non-euxinic conditions50] (Supplementary Fig. 4). The redox landscape in this part of the late Cambrian oceans included anoxic conditions with either free H2S (euxinic conditions) or Fe2+ (ferruginous conditions) prevailing in the water column, depending on the balance between H2S production and Fe2+ supply20. The MOEWC events coincide with decreases of sedimentary Fe, S, and DOPT values (Supplementary Fig. 3), indicating losses of both Fe2+ and H2S from the water column. Therefore, we infer that water column redox conditions most likely transited from euxinic to oxic over millennial timescales, and that these abrupt redox transitions were assisted or amplified by nonlinear positive feedbacks within the internal iron and sulfur cycles60.
In addition, MOEWC events could be linked to reduced biological productivity. The lower sedimentary content of the biolimiting nutrient P during the MOEWC indicates decreased biological production. We suggest that low sedimentary P concentrations were caused by reduced P recycling within the basin, as this also explains the delayed P recovery after the basin returned to euxinic conditions (Supplementary Fig. 3). Specifically, the benthic dissolved P flux would cease in oxic sediments through adsorption onto FeOOH particulates present. The recurring anoxic settings promote the P desorption from FeOOH minerals61, allowing P diffusion upward across the anoxic/oxic boundary. If this cycling dominates the P supply in the local basin, then the sedimentary P recovering trend awaits the slow rise in local P inventory, as seen in the less sharp increase of sedimentary P concentration relative to other redox proxies following MOEWC events (Supplementary Fig. 3). In this view, MOEWC periods appear to have influenced the nutrient availability in the basin even for a period after the millenial oxygenation events, and thus may have caused variation in photosynthetic productivity, organic matter remineralization in the water column, and organic matter export to sediments.
Millennial-scale release of hydrogen sulfide into surface waters (MDEs)
Given the long residence time of U and Mo in the Cambrian Ocean, faster and more complete removal during periods of intensified euxinia likely caused decreasing elemental concentrations in the sedimentary deposits, akin to that observed in restricted basins today. Widespread oceanic anoxia during the late Cambrian19,20,62 is characterized by lower U isotope values and reduced enrichments of both U and Mo, not only in the Scandinavian Alum Shale Basin10,20 but also in globally distributed records63,64. Multiple lines of evidence suggest the Alum Shale was always well connected to the open ocean, for example, modestly high Mo/TOC ratios indicate that the basin was no more than moderately restricted (e.g., comparable to modern Cariaco basin, or perhaps the Framvaren Fjord)10,65. Neodymium isotope evidence suggests the seawater mixing from the southeast Iapetus Ocean was a constant factor throughout Cambrian–Ordovician times65. And finally, Mo, U, and sulfate must have been delivered from the open oceans to maintain euxinic conditions in large parts of the Alum Shale basin for most of the time. That said, periodic shoaling of the chemocline could have moved H2S-bearing waters upwards in the water column, producing signatures similar to those observed in restricted basins, as shown in MDE scenarios C22E12 and C10E14 (Fig. 3FG). The strong negative covariation between δ98Mo and δ238U, e.g., for the C22E12 (r = −0.8, n = 12) with a slope of −1.3 (Supplementary Fig. 5), is similar to the negative regression line (slope ~ −1.5) observed in modern restricted euxinic environments33,66, suggesting MDEs represent periods with more complete Mo and U removal in the basin. That said, millennial-scale variations in water exchange (e.g., basinal restriction) likely contributed to variable H2S levels in the basin that ultimately were modulated by ocean circulation regime through wind stress, relative sea level, and seafloor bathymetry.
The δ98Mo and δ238U co-variation during MDE events offers an opportunity to estimate the compositions of open ocean seawater. Applying the model proposed by Lu et al. 32 yields the following estimates for coeval seawater: early Guzhangian (~500 Ma, C22E12) seawater would have had δ238U and δ98Mo values of –0.81 to –0.34‰ and >1.46‰, respectively. Late Paibian (~495 Ma, C10E14) seawater would have had δ238U and δ98Mo compositions of –0.83 to –0.67‰ and >1.35‰, respectively (Supplementary Fig. 5). The estimated seawater δ238U values are consistent with published δ238U data from upper Cambrian marine carbonates (–0.76 to –0.20‰)63 assuming those carbonates recorded seawater δ238U composition with a variable offset of +0.27 ± 0.28‰ (2 SD)52. Also, the estimated seawater δ98Mo is comparable to the maximum δ98Mo value of 1.33 ± 0.12‰ reported from coeval euxinic shales20 (Fig. 4).

The implication is that anoxic (and euxinic) conditions prevailed in late Cambrian oceans, as reflected by significantly lower oceanic δ238U and δ98Mo values than in the modern oceans (−0.39‰ and 2.34‰, respectively). We observe a single high peak δ98Mo value, reaching up to 1.83‰ (C23E10 despite a noisy trend; Supplementary Fig. 2E) in the early Guzhangian. This is higher than the estimate of >1.46‰ for contemporaneous seawater and may suggest a higher coeval seawater value (Fig. 4). Alternatively, the elevated sedimentary δ98Mo values may represent a temporary seawater δ98Mo overshoot caused by a “reservoir effect”, wherein preferential removal of light Mo isotopes outpaces the basin’s Mo supply67. This effect occurs when Mo burial with isotope fractionation is amplified in the euxinic basin, preferentially removing lighter Mo isotopes and leaving behind heavier isotopic residue in the basin for a short period. Consequently, the average δ98Mo value in the local basin can temporarily exceed that of the coeval open ocean. Therefore, we urge caution when using Mo-U isotope trends during plausible MDEs like C23E10 to gauge the composition of open ocean seawater. In any case, the consistently lower δ98Mo values observed during all MDEs, compared to modern seawater and restricted black shales, provide compelling evidence for a more extensively euxinic global ocean state during the late Cambrian than exists today (Fig. 4).
Relationship between local MREs and Myr-scale global redox and biotic changes
A major carbon cycle perturbation event in the late Cambrian, termed the Steptoean Positive Carbon Isotope Excursion (SPICE, from ~497.5 to ~494.5 Ma), covers a period of extreme oxygen deficiency in the global oceans10,19,63. In line with this global trend, the δ238U distributions of all ten MREs before, during, and after the SPICE event show a clear negative shift (Fig. 5), reflecting enhanced U burial in anoxic sediments worldwide20,36,63. These observations suggest that, although the studied MREs reflect local redox fluctuations, they also bear evidence of a changing global ocean redox state. In contrast, the δ98Mo records show no obvious trend across the SPICE event, implying that the δ98Mo fractionations derived from H2S fluctuations within the Alum Shale Sea had a greater influence on the sedimentary δ98Mo than changes in the open ocean composition. The MOEWC events occurred more frequently after the SPICE event (Fig. 3), aligning with the trend towards more oxygenated oceans globally in the aftermath of the SPICE event10,62. This comparison demonstrates the potential of submillimeter-scale geochemical analyses in uncovering connections between short-term local and long-term global redox variations in ancient oceans.

Distributions of (A) U concentrations, (B) δ238U values, (C) Mo concentrations, and (D) δ98Mo values for ten millennial redox events before, during, and after the SPICE event. The dashed black line within each panel represents the median value of that distribution. Before the SPICE event: C16E7, C22E9, C22E12 and C23E10; during the SPICE event: C10E14 and C12E12; after the SPICE event: C1E5, C1E6, C9E6 and C3E7. See stratigraphic levels of these ten events in Fig. 1C.
The millennial redox trends align with changes in animal diversification in the Furongian of the Baltic Alum Shale as well as in global compilations (Fig. 3B, C). Although only ten events were studied here, the intermittent MOEWC and MDE events during the Miaolingian shifted to predominantly MDE events during the early Furongian SPICE event. These changes coincided with a significant decline in diversity in Baltica, reaching its lowest point. After the SPICE event, MOEWC events gradually took over, each linked to a recovery in biodiversity in Baltica (Fig. 3C). Also, the elevated occurrences of local MOEWC through the Furongian correlate well with increases in global biotic richness and relative diversification rates (Fig. 3B). We suggest that the MOEWC events potentially created seafloor niches for aerobic, opportunistic animals. These events extended for ~650–2250 yrs, which must have been long enough for pioneer communities to colonize the seafloor and for faunal recovery, because it typically takes less than two years for mature metazoan and macrofaunal communities, and only days or weeks for small, opportunistic meiofaunal animals to colonize the seafloor68. Both trilobites and brachiopods (such as Orusia lenticularis) living in the Alum Shale Sea had planktonic larvae stage69, enabling them to migrate and survive the harsh living conditions in the Cambrian oceans. In addition, the oxygen demands of Cambrian organisms were likely lower70. Therefore, the MOEWC events have allowed opportunistic benthic communities to flourish intermittently before becoming locally or regionally extirpated when anoxic conditions re-emerged.
Materials and methods
Samples
Billegrav-2 is a fully cored scientific well with a total depth of 125.9 m, made by the Geological Survey of Denmark and Greenland (GEUS) and the University of Copenhagen on southern Bornholm, Denmark. The Alum Shale Formation was encountered between 95.4 and 122.2 m. For the geological background of the Alum Shale, see Supplementary Note 1. The Billegrav-2 core was split through the core center (55 mm core diameter) at ten shale intervals, including MREs (Fig. 1C). The ten 4–10 cm-long core pieces all demonstrate significant Mo concentration declines and/or contain benthic animal fossils.
XRF core scanning
To accurately locate the events, ten core pieces were scanned for elemental profiles at a stratigraphic resolution of 0.2 mm using an Itrax X-ray fluorescence core scanner (XRF-CS) at the Globe Institute, University of Copenhagen15. The measurements were performed on the split interior, plain, cleaned core surface. The applied voltage and current of Rh energy are 30 kV and 50 mA, respectively, and the exposure time is 100 s. Absorption peaks were identified and peak areas were quantified using the Q-Spec software CoreScanner 8.6.4 Rh from Cox Analytical Systems. Elemental concentrations were first exported with Peak Area - Concentration relationship obtained with factory settings using the SGR-1 standard (Green River Shale). To correct MoXRF-CS, we applied documented calibration for the Alum Shale (MoID-ICP-MS = 2 × MoXRF-CS – 5 ppm15; Supplementary Fig. 6A). The deviation of the slope from unity highlights the need for independent control measurements to ensure accurate trace element concentration profiles from XRF-CS. All XRF-CS Mo data reported here have been corrected using this calibration. A good correlation was also found between U concentrations obtained in XRF-CS and the isotope dilution data, UID-ICP-MS = 2.6 × UXRF-CS + 45 ppm (Supplementary Fig. 6B).
Submillimeter-scale U-Mo isotopes
To explore the MREs further using metal isotope analysis, we cut 127 ultra-narrow slots (spaced as closely as 0.25 mm apart) from the ten identically sized shale pieces along their bedding plane using a 0.25 mm-thick diamond wire, and collected the resulting powder from each slot for analyses. The shale powder was completely digested in concentrated hydrochloric and nitric acid following the procedure described by Dickson et al. 34. Uranium and molybdenum isotopes and their concentrations were analyzed by MC-ICP-MS at Royal Holloway University of London, UK. The sample solutions were spiked using 97Mo–100Mo and IRMM 3636a 233U–236U and purified using AG1-X8 anion resin and UTEVA resin. Uncertainties for individual δ98Mo analyses are propagated from the sample standard error and the bracketing standards (NIST 3134). Uncertainties for δ238U are the two-fold standard deviations of long-term external reproducibility of USGS Devonian Ohio Shale (SDO-1, ±0.07‰, 2 SD, n = 9) over the three months of Alum Shale measurements. For more details, see Supplementary Note 2. The results are listed in Supplementary Table 1 and Supplementary information.
Animal diversity
Species-level richness in each biozone from the Baltic Alum Shale Formation is based on refs. 21,25,71 with a few adjustments according to refs. 72,73. The ages for the lower boundary of biozones are based on ref. 36. The Baltic richness data are listed in Supplementary Table 2. The global generic richness data are from refs. 74,75.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data are available in the article and/or the Supplementary Information. The original geochemical data are publicly available at Zenodo (https://doi.org/10.5281/zenodo.16810480).
References
Jenkyns, H. C. Geochemistry of oceanic anoxic events. Geochem. Geophys. Geosyst. 11, 1–30 (2010).
Zou, C. et al. Unveiling the oldest industrial shale gas reservoir: insights for the enrichment pattern and exploration direction of lower Cambrian shale gas in the Sichuan basin. Engineering 42, 278–294 (2024).
Jin, Z. et al. Organic carbon cycling and black shale deposition: an Earth system science perspective. Natl. Sci. Rev. 10, nwad243 (2023).
Luo, Q. et al. Dispersed organic matter from pre-Devonian marine shales: a review on its composition, origin, evolution, and potential for hydrocarbon prospecting. Earth Sci. Rev. 261, 105027 (2025).
Fu, F. et al. Oceanic redox variations during the Cambrian age 2–3 in the Yangtze Block of South China: evidence from pyrite-based proxies. Mar. Pet. Geol. 179, 107438 (2025).
Zou, C. et al. Unconventional petroleum sedimentology: a key to understanding unconventional hydrocarbon accumulation. Engineering 18, 62–78 (2022).
Dahl, T. W. et al. Devonian rise in atmospheric oxygen correlated to the radiations of terrestrial plants and large predatory fish. Proc. Natl. Acad. Sci. USA107, 17911–17915 (2010).
Lenton, T. M., Daines, S. J. & Mills, B. J. W. COPSE reloaded: an improved model of biogeochemical cycling over Phanerozoic time. Earth Sci. Rev. 178, 1–28 (2017).
Cheng, M. et al. Marine Mo biogeochemistry in the context of dynamically euxinic mid-depth waters: a case study of the lower Cambrian Niutitang shales, South China. Geochim. Cosmochim. Acta 183, 79–93 (2016).
Zhao, Z. et al. Dynamic oceanic redox conditions across the late Cambrian SPICE event constrained by molybdenum and uranium isotopes. Earth Planet. Sci. Lett. 604, 118013 (2023).
Wei, G. et al. Global marine redox evolution from the late Neoproterozoic to the early Paleozoic constrained by the integration of Mo and U isotope records. Earth Sci. Rev. 214, 103506 (2021).
Sperling, E. A. et al. A long-term record of early to mid-Paleozoic marine redox change. Sci. Adv. 7, eabf4382 (2021).
Canfield, D. E. et al. Ferruginous conditions dominated later Neoproterozoic deep-water chemistry. Science 321, 949–952 (2008).
Zhao, Z. et al. Weathering-induced organic matter enrichment in marine-continental transitional shale: a case study on the early Permian Taiyuan Formation in the Ordos Basin, China. Int. J. Coal Geol. 291, 104562 (2024).
Dahl, T. W. et al. Brief oxygenation events in locally anoxic oceans during the Cambrian solves the animal breathing paradox. Sci. Rep. 9, 11669 (2019).
German, C. R. & Elderfield, H. Rare earth elements in Saanich Inlet, British Columbia, a seasonally anoxic basin. Geochim. Cosmochim. Acta 53, 2561–2571 (1989).
Wei, G. et al. Highly dynamic marine redox state through the Cambrian explosion highlighted by authigenic δ238U records. Earth Planet. Sci. Lett. 544, 116361 (2020).
Dahl, T. W. et al. The behavior of molybdenum and its isotopes across the chemocline and in the sediments of sulfidic Lake Cadagno, Switzerland. Geochim. Cosmochim. Acta 74, 144–163 (2010).
Gill, B. C. et al. Geochemical evidence for widespread euxinia in the Later Cambrian ocean. Nature 469, 80–83 (2011).
Gill, B. C. et al. Redox dynamics of later Cambrian oceans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 581, 110623 (2021).
Weidner, T., Nielsen, A. T. & Ebbestad, J. O. R. in Middle Cambrian Agnostoids and Trilobites from the Lower Allochthon, Swedish Caledonides Fossils and Strata 1–121 (Scandinavian University Press, 2023).
Uchman, A., Bąk, K. & Rodríguez-Tovar, F. J. Ichnological record of deep-sea palaeoenvironmental changes around the Oceanic Anoxic Event 2 (Cenomanian–Turonian boundary): an example from the Barnasiówka section, Polish Outer Carpathians. Palaeogeogr. Palaeoclimatol. Palaeoecol. 262, 61–71 (2008).
Schovsbo, N. H. Why barren intervals? A taphonomic case study of the Scandinavian Alum Shale and its faunas. Lethaia 34, 271–285 (2001).
Ahlberg, P. et al. Cambrian high-resolution biostratigraphy and carbon isotope chemostratigraphy in Scania, Sweden: first record of the SPICE and DICE excursions in Scandinavia. Lethaia 42, 2–16 (2009).
Nielsen, A. T., Høyberget, M. & Ahlberg, P. The Furongian (upper Cambrian) Alum Shale of Scandinavia: revision of zonation. Lethaia 53, 462–485 (2020).
Clarkson, E. N. K., Ahlberg, P. & Taylor, C. M. Faunal dynamics and microevolutionary investigations in the Upper CambrianOlenusZone at Andrarum, Skåne, Sweden. GFF 120, 257–267 (1998).
Dahl, T. W. et al. Reorganisation of Earth’s biogeochemical cycles briefly oxygenated the oceans 520 Myr ago. Geochem. Perspect. Lett. 3, 210–220 (2017).
Dahl, T. W. et al. Atmosphere-ocean oxygen and productivity dynamics during early animal radiations. Proc. Natl. Acad. Sci. USA 116, 19352–19361 (2019).
Ku, T.-L., Mathieu, G. G. & Knauss, K. G. Uranium in open ocean: concentration and isotopic composition. Deep Sea Res. 24, 1005–1017 (1977).
Miller, C. A., Peucker-Ehrenbrink, B., Walker, B. D. & Marcantonio, F. Re-assessing the surface cycling of molybdenum and rhenium. Geochim. Cosmochim. Acta 75, 7146–7179 (2011).
Algeo, T. J. & Tribovillard, N. Environmental analysis of paleoceanographic systems based on molybdenum–uranium covariation. Chem. Geol. 268, 211–225 (2009).
Lu, X. et al. Estimating ancient seawater isotope compositions and global ocean redox conditions by coupling the molybdenum and uranium isotope systems of euxinic organic-rich mudrocks. Geochim. Cosmochim. Acta 290, 76–103 (2020).
Brüske, A. et al. Correlated molybdenum and uranium isotope signatures in modern anoxic sediments: Implications for their use as paleo-redox proxy. Geochim. Cosmochim. Acta 270, 449–474 (2020).
Dickson, A. J. et al. Isotopic constraints on ocean redox at the end of the Eocene. Earth Planet. Sci. Lett. 562, 116814 (2021).
Sørensen, A. L. et al. Astronomically forced climate change in the late Cambrian. Earth Planet. Sci. Lett. 548, 116475 (2020).
Zhao, Z. et al. Synchronizing rock clocks in the late Cambrian. Nat. Commun. 13, 1990 (2022).
Sarmiento, J. L. & Gruber, N. Ocean Biogeochemical Dynamics Vol. 528 (Princeton University Press, 2006).
Dahl, T. W. et al. Evidence of molybdenum association with particulate organic matter under sulfidic conditions. Geobiology 15, 311–323 (2017).
Bone, S. E., Dynes, J. J., Cliff, J. & Bargar, J. R. Uranium(IV) adsorption by natural organic matter in anoxic sediments. Proc. Natl. Acad. Sci. USA 114, 711–716 (2017).
Helz, G. R., Bura-Nakić, E., Mikac, N. & Ciglenečki, I. New model for molybdenum behavior in euxinic waters. Chem. Geol. 284, 323–332 (2011).
Helz, G. R. Dissolved molybdenum asymptotes in sulfidic waters. Geochem. Perspect. Lett. 19, 23–26 (2021).
Tossell, J. A. Calculating the partitioning of the isotopes of Mo between oxidic and sulfidic species in aqueous solution. Geochim. Cosmochim. Acta 69, 2981–2993 (2005).
Kerl, C. F. et al. Experimental confirmation of isotope fractionation in thiomolybdates using ion chromatographic separation and detection by multicollector ICPMS. Anal. Chem. 89, 3123–3129 (2017).
Erickson, B. E. & Helz, G. R. Molybdenum(VI) speciation in sulfidic waters. Geochim. Cosmochim. Acta 64, 1149–1158 (2000).
Neubert, N., Nägler, T. F. & Böttcher, M. E. Sulfidity controls molybdenum isotope fractionation into euxinic sediments: evidence from the modern Black Sea. Geology 36, 775–778 (2008).
Nägler, T. F. et al. Molybdenum isotope fractionation in pelagic euxinia: evidence from the modern Black and Baltic Seas. Chem. Geol. 289, 1–11 (2011).
Noordmann, J. et al. Uranium and molybdenum isotope systematics in modern euxinic basins: Case studies from the central Baltic Sea and the Kyllaren fjord (Norway). Chem. Geol. 396, 182–195 (2015).
Poulson Brucker, R. L., McManus, J., Severmann, S. & Berelson, W. M. Molybdenum behavior during early diagenesis: Insights from Mo isotopes. Geochem. Geophys. Geosyst. 10, 1–25 (2009).
Algeo, T. J. & Maynard, J. B. Trace-element behavior and redox facies in core shales of Upper Pennsylvanian Kansas-type cyclothems. Chem. Geol. 206, 289–318 (2004).
Algeo, T. J. & Li, C. Redox classification and calibration of redox thresholds in sedimentary systems. Geochim. Cosmochim. Acta 287, 8–26 (2020).
Andersen, M. B., Stirling, C. H. & Weyer, S. Uranium isotope fractionation. Rev. Mineral. Geochem. 82, 799–850 (2017).
Zhang, F. et al. Uranium isotopes in marine carbonates as a global ocean paleoredox proxy: a critical review. Geochim. Cosmochim. Acta 287, 27–49 (2020).
Weyer, S. et al. Natural fractionation of 238U/235U. Geochim. Cosmochim. Acta 72, 345–359 (2008).
Barling, J. & Anbar, A. D. Molybdenum isotope fractionation during adsorption by manganese oxides. Earth Planet. Sci. Lett. 217, 315–329 (2004).
Goto, K. T. et al. Uranium isotope systematics of ferromanganese crusts in the Pacific Ocean: implications for the marine 238U/235U isotope system. Geochim. Cosmochim. Acta 146, 43–58 (2014).
Scott, C. & Lyons, T. W. Contrasting molybdenum cycling and isotopic properties in euxinic versus non-euxinic sediments and sedimentary rocks: Refining the paleoproxies. Chem. Geol. 324, 19–27 (2012).
Rudnick, R. L. & Gao, S. in Treatise on Geochemistry Vol. 3 (eds H. D. Holland & K. K. Turekian) 1–64 (Elsevier, 2003).
Hardisty, D. S. et al. A Holocene history of dynamic water column redox conditions in the Landsort Deep, Baltic Sea. Am. J. Sci. 316, 713–745 (2016).
Pohl, A. et al. Continental configuration controls ocean oxygenation during the Phanerozoic. Nature 608, 523–527 (2022).
van de Velde, S. J., Reinhard, C. T., Ridgwell, A. & Meysman, F. J. R. Bistability in the redox chemistry of sediments and oceans. Proc. Natl. Acad. Sci. USA 117, 33043–33050 (2020).
Colman, A. S. & Holland, H. D. in Marine Authigenesis: From Global to Microbial (eds Craig R. Glenn, Liliane Prévôt-Lucas, & Jacques Lucas) 53–75 (SEPM Society for Sedimentary Geology, 2000).
Saltzman, M. R. et al. Pulse of atmospheric oxygen during the late Cambrian. Proc. Natl. Acad. Sci. USA 108, 3876–3881 (2011).
Dahl, T. W. et al. Uranium isotopes distinguish two geochemically distinct stages during the later Cambrian SPICE event. Earth Planet. Sci. Lett. 401, 313–326 (2014).
He, R. et al. The dynamic ocean redox evolution during the late Cambrian SPICE: Evidence from the I/Ca proxy. Glob. Planet. Change 233, 104354 (2024).
Sturesson, U. L. F. et al. Neodymium isotopic composition of Cambrian–Ordovician biogenic apatite in the Baltoscandian Basin: implications for palaeogeographical evolution and patterns of biodiversity. Geol. Mag. 142, 419–439 (2005).
Bura-Nakić, E. et al. Coupled Mo-U abundances and isotopes in a small marine euxinic basin: constraints on processes in euxinic basins. Geochim. Cosmochim. Acta 222, 212–229 (2018).
Dahl, T. W. et al. Sulfidic anoxia in the oceans during the Late Ordovician mass extinctions – insights from molybdenum and uranium isotopic global redox proxies. Earth Sci. Rev. 220, 103748 (2021).
Sperling, E. A. et al. Oxygen, facies, and secular controls on the appearance of Cryogenian and Ediacaran body and trace fossils in the Mackenzie Mountains of northwestern Canada. Geol. Soc. Am. Bull. 128, 558–575 (2016).
Nielsen, A. T. A review of Ordovician agnostid genera (Trilobita). Trans. R. Soc. Edinb. 87, 463–501 (1997).
Mills, D. B. et al. Oxygen requirements of the earliest animals. Proc. Natl. Acad. Sci. USA 111, 4168–4172 (2014).
Żylińska, A., Weidner, T., Ahlgren, J. & Ahlberg, P. Exotic trilobites from the uppermost Cambrian series 3 and lower Furongian of Sweden. Acta Geol. Pol. 65, 21–67 (2015).
Nielsen, A. T. & Andersen, L. F. Furongian (upper Cambrian) trilobites and agnostoids from the Alum Shale Formation of Bornholm, Denmark: revised taxonomy and biostratigraphy. Bull. Geol. Soc. Den. 69, 123–213 (2021).
Ahlberg, P. & Ahlgren, J. Agnostids from the upper Cambrian of Västergötland, Sweden. GFF 118, 129–140 (1996).
Rasmussen, C. M. Ø, Kröger, B., Nielsen, M. L. & Colmenar, J. Cascading trend of early Paleozoic marine radiations paused by Late Ordovician extinctions. Proc. Natl. Acad. Sci. USA 116, 7207–7213 (2019).
Kroger, B., Franeck, F. & Rasmussen, C. M. O. The evolutionary dynamics of the early Palaeozoic marine biodiversity accumulation. Proc. Natl. Acad. Sci. USA 286, 20191634 (2019).
Zhao, Z. et al. High-resolution carbon isotope chemostratigraphy of the middle Cambrian to lowermost Ordovician in southern Scandinavia: implications for global correlation. Glob. Planet. Change 209, 103751 (2022).
Nielsen, A. T. et al. Gamma-ray log correlation and stratigraphic architecture of the Cambro-Ordovician Alum Shale formation on Bornholm, Denmark: evidence for differential syndepositional isostasy. Bull. Geol. Soc. Den. 66, 237–273 (2018).
Acknowledgements
This study was financially supported by the Science Fund for Creative Research Groups of the National Natural Science Foundation of China (42321002), Joint Funds of the National Natural Science Foundation of China (U23B20155), the Carlsberg Foundation (CF16–0876), Young Scientists Fund of the National Natural Science Foundation of China (42303004), the National Key R&D Program of China (2021YFA0718200), and the Fundamental Research Funds for the Central Universities (2025XJDC01). We thank Editor Alice Drinkwater and three anonymous reviewers for their insightful comments and constructive suggestions, which have greatly improved this work. No permissions were required for sample collections.
Author information
Authors and Affiliations
Contributions
T.W.D. conceived the idea. T.W.D. and Z.Z. designed the study. Z.Z., A.T.N., N.H.S., T.W.D., and I.W.H. collected the samples. Z.Z., I.W.H., A.J.D., and A.B. performed the elemental and isotopic analyses. A.T.N. compiled the species-level richness dataset. T.W.D. and C.Z. supervised the research and guided data interpretation. Z.Z., Z.J., P.J., and Z.G. prepared the data visualizations. S.D., C.Z., T.W.D., Z.J., and Z.Z. acquired funding for the project. Z.Z. wrote the initial manuscript draft with contributions from T.W.D., A.J.D., C.Z., S.D., A.T.N., Z.J., P.J., Z.G., A.B., N.H.S., and I.W.H. All authors contributed substantially to data acquisition, interpretation, and discussion of the manuscript content.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Ganqing Jiang and the other anonymous reviewer(s) for their contribution to the peer review of this work. Primary handling editors: Mojtaba Fakhraee, Carolina Ortiz Guerrero, and Alice Drinkwater. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhao, Z., Hougård, I.W., Zou, C. et al. Sub-millimeter molybdenum and uranium isotopes track millennial redox events in the Cambrian ocean. Commun Earth Environ 6, 766 (2025). https://doi.org/10.1038/s43247-025-02722-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-025-02722-2
