
Abstract
Background
Exposure to repetitive head impacts (RHI) has been shown to be associated with cognitive impairment and markers of neurodegeneration in professional male soccer players. However, data on the effects of RHI in female players are very limited. In this exploratory study, we investigated changes in blood biomarkers along with the cognitive status of elite female soccer players and compared the effect of RHI with outcomes in male players.
Methods
In a cohort study, elite female soccer players (age 16–41 years) performed training without heading the ball (n = 36) and training including heading (n = 30). Athletes were evaluated for the parameters of physical activity (heart rate) and neuropsychological testing (focused attention and cognitive flexibility) before and 1 h after each training. The levels of plasma tau, pTau181, pTau217 and microRNAs were determined at four timepoints: before training, 1 h, 24 h, and 48 h after each training session. Parameters in females were compared with control male players (n = 32).
Results
The levels of tau, pTau181 and pTau217 are significantly increased in the plasma of female soccer players 1 hour after physical exercise (tau, 1.4-fold; pTau181, 1.3-fold; pTau217, 1.2-fold) and repetitive head impacts (tau, 1.2-fold; pTau181, 1.3-fold; pTau217, 1.3-fold) compared to pre-training levels. The ratio of pTau181 to tau fold change is significantly higher 1 hour after heading and remains elevated specifically in the heading group even after 48 h. The analysis of deregulated microRNAs in plasma reveals enrichment of neuroprotective pathways after heading training. Female soccer players show worse cognitive functions after heading compared to exercise and also compared to male soccer players.
Conclusions
RHIs may lead to acute disbalance of tau and phosphorylated tau in plasma, and are associated with short-term decline in cognitive performance and neuroprotective pathways in elite female soccer players. The sexual dimorphism in response to RHI may reflect biological risk factors potentially relevant to the different degree of susceptibility to neurological disorders in females and males in later life.
Plain language summary
Majority of neuroscience research is conducted on male subjects, creating a gap in the knowledge. Here we investigated the effect of repetitive non-concussive head impacts (RHI) in elite female soccer players. The athletes were evaluated for the parameters of physical activity, cognition and levels of plasma tau protein which are known biomarkers of neurodegeneration. Our data demonstrate that repetitive heading of the ball is associated with biochemical alterations, reduction in focused attention, and cognitive flexibility. Here we also show that females are more sensitive to head impacts when compared to males and that the RHI exposure is associated with neuroprotective signaling. These findings link specific molecular pathways, potentially operating in the early stage of brain diseases. Deeper exploration of these pathways could be a promising approach to identify new biomarkers and therapeutic targets in the treatment of neurodegeneration.
Similar content being viewed by others

Introduction
The lack of efficient treatment for neurocognitive decline and neurodegenerations resulting in dementia is a major health and societal problem on a global scale1,2. Despite more females suffering from neurodegeneration, most studies on the mechanisms of the disease have been conducted on males. This can lead to neglection of important etiopathogenic mechanisms associated with neurodegeneration and cognitive decline in older age, as data from males are not directly applicable to the female population3,4,5.
Several large and well-controlled cross-sectional epidemiological studies on former professional soccer players clearly suggest that repetitive head impacts, even if mild and not causing immediate symptoms, are important risk factors for neurological diseases and dementia6,7,8.
The increased prevalence of dementia shown in male soccer players6,7, can be expected to emerge also in female players, as the number of players has increased rapidly over the past decade9. This increase can be further driven by the fact that female soccer players were shown to have a higher relative risk of concussion10,11,12,13 and higher incidence of concussion caused by heading the ball, when compared to male players14.
Repetitive headers in soccer can lead to acute impairment of short-term learning15,16,17,18 along with long-term impairment of cognitive performance, which has recently been documented in retired professional soccer players19. While it is accepted that there is an association between the heading frequency and cognitive decline, the causality has been questioned20,21. Well designed and controlled experimental studies, such as the study presented here, can bring further evidence to elucidate these associations.
Apart from the psychological evaluation, specific molecular markers in peripheral blood, would be useful to precisely assess the physiological changes incurred by the head impacts.
Phosphorylated tau proteins have recently been evaluated as promising candidates for the diagnosis and prognosis of acute or chronic TBI and also for the differentiation of AD and other types of neurodegeneration22,23,24,25. Their increased levels do not reflect the complexity of molecular processes induced by or associated with concussion. This can be achieved by profiling the peripheral microRNAs (miRNAs)26,27,28,29,30. However, experimental findings on miRNAs and their bioinformatic analysis of molecular events associated with repetitive nonconcussive head impacts are very limited26,31. Moreover, we are not aware of even a single such publication specifically focused on females.
In our previous research, we showed increased plasma tau levels and impaired cognitive performance after physical exercise and heading training in male soccer players32,33.
The purpose of this study is to evaluate the acute effects of mild repetitive head impacts (mRHI) in female soccer players. We show that females are more sensitive to repetitive head impacts than males, while this sensitivity is associated with alterations in cognition and peripheral fluid biomarkers. In a well-controlled study, the short-term cognitive abilities along with circulating tau, phosphorylated tau, miRNAs and signalling pathways after mRHI are compared with non-head-impact physical exercise. In addition, we discuss acute changes in cognition and biomarkers in females with findings observed in males and further explore sex-related alterations in response to mRHI.
Methods
Participants
Study participants were college soccer players at Comenius University in Bratislava and senior category players in various elite Slovak soccer clubs. A total of 36 female soccer players between the ages of 16 and 41 years participated in the study. The ethical review board of Comenius University in Bratislava, Faculty of Physical Education and Sport approved this study with human participants (#1/2022), and all involved players and their legal representatives (applies to participants under the age of 18) provided written informed consent prior to beginning the study. The study was conducted from March 1, 2023 to September 30, 2023 and followed the Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) reporting guideline for cohort studies.
Study design
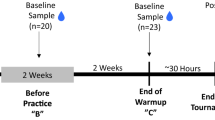
Training sessions were designed during 3 consecutive weeks, with maximal heart rate determination at week 0 followed by soccer training without heading the ball (week 1) and subsequent soccer training including heading drills (week 2). Training intensity was monitored using heart rate sensors (Polar Electro Oy, Kempele, Finland) at 5 s intervals. To determine maximum heart rate, we used a running field test recommended for sports games. The test comprised a warm-up (10 min) and 3 consecutive running exercises performed on a soccer field (95 × 42.5 m). Athletes were instructed to complete each block of 275 meters at their highest pace (as fast as possible), with a 2 min active recovery interval – walking34,35. The highest heart rate attained during this test was determined as the maximal heart rate. Most players reached their maximum heart rate in the second series of the test. The data including maximal heart rate acquired during week 0 training served as an indicator for designing the consecutive soccer training and heading training. The intensity of physical exertion was aimed at maintaining physical activity within the aerobic interval ( ~ 75 % of maximal heart rate) allowing simultaneous performance of soccer exercise and accomplishment of ball heading tasks. The players followed a resting regime 24 h before and between blood collection time points.
Outcomes
The primary study outcomes were the plasma levels of total tau protein, pTau181, pTau217, miRNAs and cognitive status (attention and flexibility) of the elite female soccer players. Study participants were assessed for primary outcomes at 4 time points, before (baseline) and after (1, 24, and 48 h) the 2 types of training, soccer exercise without heading the ball and soccer exercise including low-intensity repetitive headers. Outcomes were assessed at the specified timepoints following the entire completion of the study. The outcomes of male to female comparisons were the fold change of total tau protein and pTau181, fold change of pTau181 relative to tau fold change, and the cognitive status of male and female soccer players assessed at 3 time points before (baseline) and after (1 h, 24 h) the 2 types of training, soccer exercise without heading the ball and soccer exercise including low-intensity repetitive headers.
Soccer training
Soccer training without heading the ball (exercise group, abbreviated as E) with a total duration of 85 min comprised a warm-up including running (7 min), exercises without the ball and exercises with the ball (67 min), two exercise blocks of 2 different small-sided games (SSG) (each SSG lasted 2 min with a 2 min rest interval, a total of 6 min, pitch size 25 × 18 m), and a final compensatory exercise (5 min). Players were not allowed to do headers, while occasional player-to-player body contacts that represent an integral part of the game were not avoided.
Heading training
The soccer training with heading exercises (heading group, abbreviated as H + E) followed a modified training protocol. The warm-up included two sets of three-meter hands passes the ball with underhand throw to the head of a teammate (20 min). The main H + E training included 6 blocks of heading exercise with a total duration of 60 min: 1. Heading after hands passes the ball with underhand throw in a sitting position (sit with the spread legs) for two sets, each set with 10 attempts (5 min); 2. Heading in the square for two sets of 10 attempts lasting 3 min with a 30 s rest interval (7 min); 3. Heading in the zig-zag exercises for four sets of 10 attempts, each set lasting 3 min with a 30 s rest interval (15 min); 4. Heading over the semi-active defender after hands passes the ball with underhand throw of 10 attempts (15 min); 5. Heading in pairs for two sets, each set lasted 1 min with a 30 s rest interval (3 min); 6. Four small-sided games with a given pitch size (25 × 18 m) and a rule that a goal can only be scored with a header. During the SSG, hands passes the ball with underhand throw and head passes were continuously alternated. Each SSG lasted 2 min with a 2 min rest interval (15 min total), followed by a final compensatory exercise (5 min). Heading training included a pool of low-intensity headers that correspond to the practice of heading techniques in youth to adult categories. Game situations with high-intensity head impacts, such as headers after corner kicks, goalkeeper kickouts, or long-distance passes, were not played in this study. Training units were videotaped to cross-check the number of headers for each player.
Officially tested FIFA Quality Pro marked balls were used during the entire study. The ball parameters were in accordance with the International Football Association Board 2022/23 Laws of the game, Law 236.
Biochemical procedures
Venous blood was collected by professional medical personnel at 4 timepoints: before training, 1 h, 24 h, and 48 h after each training unit. The blood collection was performed in a nonfasting state using K2EDTA tubes (Cat. No. 455045, Greiner Bio-One, Kremsmünster, Austria). All samples were centrifuged within 60 minutes from the time of collection, at 2000 x g for 10 min at 4 °C. Prepared plasma aliquots were stored at −80 °C until analysis.
Tau protein concentrations were assayed by digital ELISA using the Simoa Tau 2.0 Advantage kit (Cat. No. 101552, Quanterix Corp., Billerica, MA, USA). Tau protein phosphorylated at threonine 181 (pTau181) and threonine 217 (pTau217) was measured using the Simoa pTau-181 V2.1 Advantage Kit (Quanterix, Cat. No. 104111) and ALZpath Simoa p-Tau 217 v2 Assay Kit (Quanterix, Cat. No. 104371). All samples were blinded prior to analysis and assayed in a single measurement in duplicates on the HD-X Analyzer (Quanterix Corp, MA, USA). The mean intra-assay coefficients of variation of tau, pTau181 and pTau217 measurements were 5, 8 and 6%, respectively.
MicroRNA profiling
Total RNA was extracted from 400 µL of plasma using the miRNeasy Serum/Plasma Advanced Kit (Cat.No. 217204, Qiagen, Hilden, Germany) according to the manufacturer’s protocol. To increase the yield of ribonucleic acids, 2 µg of glycogen was used as a carrier for each sample (Cat. R0551, Thermo Scientific, Vilnius, Lithuania). The resulting RNA was eluted in 25 µL of RNAse-free water. Analysis of miRNA levels was performed using pooled samples within each group. Each equivolumic pool comprised five randomly selected individual samples, with every timepoint being represented by three pools. This strategy enabled cost-effective miRNA discovery and has been previously used in other miRNA biomarker studies26,37.
The integrity and profile of extracted RNA were analyzed by 2100 Bioanalyzer using the RNA 6000 Pico Kit (Cat. No. 5067-1513, Agilent Technologies, Waldbronn, Germany). Reverse transcription of template RNA (4 µL) in each pool was performed using the miRCURY LNA RT Kit (Qiagen, Cat. No. 339340). cDNA samples were assessed for hemolysis evaluating the delta Cq values for miR-23a-3p – miR-451a38.
Expression profiles of 179 individual miRNAs were quantified using the Human serum/plasma miRCURY LNA miRNA PCR array (Qiagen, Cat. No. 339325, YAHS-106Y-A) and miRCURY LNA SYBR Green PCR Kit (Qiagen, Cat. No. 339347) amplified by QuantStudio 6 Flex Real-Time PCR System (Applied Biosystems, Singapore).
The fold change of miRNA expression at each timepoint was calculated using the 2-ΔΔCq method, compared to its respective baseline group (before training) with the geometric mean of Cq values as an endogenous control. Relative quantity and statistical significance for all evaluated miRNAs are summarized in Supplementary Data 1 and Supplementary Data 2.
Bioinformatics
We queried mirDIP ver. 5.3 (https://ophid.utoronto.ca/mirDIP)39 to identify gene targets of deregulated miRNAs. Pathway enrichment analysis was performed using pathDIP ver. 5 (https://ophid.utoronto.ca/pathDIP)40, using miRNA target genes as input, and using only literature-curated (core) pathway memberships. Statistically significant pathways (adjusted P < 0.05; FDR: BH-method) were further considered and grouped into pathway categories, as introduced in Pastrello et al.40. Interactions among miRNA target genes were retrieved using IID ver. 2021-05 (https://ophid.utoronto.ca/iid)41. Resulting networks were further annotated with Alzheimer’s disease-related genes from IID, as introduced in Kotlyar et al.42. All networks were built, annotated and analyzed using NAViGaTOR ver. 3.0.1943. The stacked bar chart ver. 32.3.0 and the scatter plot ver. 22.2.2 were used to prepare the plots in Fig. 2, using Fluorish studio (https://flourish.studio) (Canva UK Operations Limited, London, UK). The figures were exported in SVG format and finalized in CorelDraw ver. 24.0.0.301 to include legends.
Psychological testing
Neuropsychological examination of athletes after soccer and heading training involved testing of visual scanning and working memory using the Trail Making Test (TMT)44,45.
All participants were tested before (baseline) and after each training sessions (1 h), at timepoints aligned with blood collection. The assessment consisted of two parts: TMT-A focused on intensity and sustaining of attention; TMT-B focused on cognitive flexibility in executive functioning. In the TMT-A, each participant was instructed to draw a line between 25 randomly arranged circles in ascending order (1-2-3 etc.). In TMT-B, the subject was asked to connect the circles while alternating numbers and letters in consecutive order (1-A-2-B etc.). Different variations of TMT tests were used in each of the exercises, further eliminating possible practice effect. To achieve this, the spatial order of the numeric and alphanumeric targets was pseudo-randomized across test variants. Each version preserved the core structure and cognitive demands of the original TMT-A and TMT-B, including the number of items, the alternation rule (in TMT-B), the general spatial dispersion, and the relative density of elements. The randomization procedure was constrained to ensure psychometric equivalence across all test forms. Specifically, target locations were generated using a controlled randomization algorithm that maintained comparable path lengths, angular complexity, and spatial clustering patterns. Care was taken to avoid overly linear or overly compact arrangements, as these could reduce cognitive and visuomotor challenge. As a result, each version imposed a similar cognitive workload in terms of visual scanning, attention switching, sequencing, and motor planning. The alternative forms of TMT tests used in our study represent a standard screening tool with consistent difficulty, high reliability, and are suitable for sequential testing of executive functions46,47. As both training sessions were organized in consecutive weeks, cognitive performance was assessed before both trainings to minimize the practice effect in this short time frame. Duration time for each task was recorded, and results are displayed as a fold change from baseline (Fig. 1e, f).

a Schematic illustration of the study protocol, sampling and analysis. b Monitoring of maximal heart rate revealed no difference between exercise and heading groups. Athletes performed exercise (E) with higher physical activity compared with heading (H + E) training, represented by the higher (c) mean heart rate and higher (d) percentage of maximal heart rate. e Both training sessions were associated with decreases of focused attention of athletes when compared to baseline (black dotted line) as assessed by the Trail Making Test A (TMT-A), while the worsening was higher in the heading group. f Heading session also showed negative outcomes on cognitive flexibility (TMT-B) of athletes compared to baseline and exercise. Psychological testing revealed that heading training was associated with worse cognitive abilities than exercise without repetitive headers. g Analysis of total tau, pTau181 and pTau217 in plasma revealed no significant difference between exercise and heading groups at baseline. Horizontal black lines denote the mean and 95% CI. h Significant increase in the fold change of plasma tau from baseline was observed in athletes 1 hour after both training sessions, while there was a significant difference between E and H + E training. The higher change in tau protein normalized to baseline (black dotted line) in the exercise group was significantly decreased in the heading group after 48 hours. Significant change in plasma (i) pTau181 and (j) pTau217 from baseline was observed 1 hour after both types of training. The increased change in phosphorylated tau protein normalized to baseline within 24 hours. k The ratio of pTau181 to tau fold change was significantly higher 1 hour and 48 hours after heading when compared with exercise. Similar, yet nonsignificant increase of (l) pTau217 to tau fold change ratio in the heading group was also observed. The color of the P value in (h-j) indicates within-group significance between baseline and corresponding time points, as per legend. The interaction between groups is indicated by lines and the corresponding P value in black color. Statistical significance (b-d) was assessed using the two-tailed Mann-Whitney test. Statistical significance (e-l) was assessed using the two-way ANOVA with Tukey’s multiple comparison test. Exercise group (n = 36), heading group (n = 30) in tau and pTau181 analysis and both groups (n = 11) in pTau217 analysis.
Statistics and reproducibility
Statistical analyses were performed using GraphPad Prism (version 10.2.2) software (GraphPad Software, Boston, MA, USA). Data distribution was tested using the D’Agostino-Pearson omnibus normality test to determine whether parametric or nonparametric tests should be used. Two-tailed Mann-Whitney test was used for the analyses of demographic characteristics and physical exercise. As the groups did not differ significantly in demographic characteristics (Table 1), no adjustment for these variables was applied in statistical evaluation of biomarkers and cognition. The plasma protein concentration and cognitive status is expressed as a fold change at 1, 24, and 48 h from its own baseline for each athlete. Group changes are visualized as mean with a 95% confidence interval (CI). The mean fold change from baseline was compared in both training arms and at all timepoints using 2-way ANOVA considering the time (baseline, 1 hour, 24 hours, and 48 hours) as a within-participant factor and group (exercise, heading) as a between-participant factor, using Tukey multiple comparisons test. Multiplicity adjusted P values for each comparison were considered significant at the P < 0.05 level. For comparison of miRNA expression between the two training arms, unpaired two-tailed Student’s t-test was used. Correlation between tau biomarkers and the number of headers was assessed with the Spearman correlation coefficients. The ability of plasma proteins to discriminate athletes with mild repetitive head impacts vs exercise controls was assessed using the receiver operating characteristics (ROC) curve and associated area under the curve (AUC). Correlations were considered significant at 2-tailed P < 0.05 level.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Results
Demographic information
Study participants included 36 elite soccer players (all females of White European ancestry). After maximal heart rate determination in week 0, all players completed soccer training without heading the ball in week 1 (hereafter referred to as “exercise group”), with 30 of them also completing soccer training including heading exercises in week 2 (hereafter referred to as “heading group”; Fig. 1a). The groups did not differ significantly in demographic characteristics (Table 1). Individuals with a history of TBI were excluded.
Training parameters
The intensity of sports activity was monitored by heart rate monitors during both training units. Maximal heart rate of female soccer players was similar for exercise group (E) and heading group (H + E) (E = 191.6 bpm; 95% CI, 189–194 bpm vs H + E = 191.8 bpm; 95% CI, 189–195 bpm; P = 0.90). However, the mean heart rate in the exercise group was significantly higher compared to the heading group (E = 146.8 bpm; 95% CI, 143–151 bpm vs H + E = 140.4 bpm; 95% CI, 136–145 bpm; P = 0.047). Correspondingly, both types of training differed in the mean percentage of maximal heart rate (E = 76%, 95% CI, 75–78% vs H + E = 73%; 95% CI, 71–76%; P = 0.03), indicating higher physical activity during exercises without heading the ball (Fig. 1b-d). The mean (SD) performed low-intensity headers among players during H + E training was 152 (15) (median [IQR], 154 [144–161]) (Table 2).
Cognitive outcomes
Neuropsychological testing of female soccer players identified a deterioration of focused attention (TMT-A) in both training groups compared to pre-training baseline (E = 1.8-fold; 95% CI, 1.7–2.0; P < 0.001 and H + E = 3.4-fold; 95% CI, 3.1–3.7; P < 0.001), while decrease in cognitive performance was significantly higher in the heading group (P < 0.001, Fig. 1e). On the other hand, the examination of cognitive flexibility (TMT-B) identified significantly worse score only in the heading group (H + E = 1.5-fold; 95% CI, 1.4–1.7; P < 0.001), while the score in exercise group was not significantly altered (E = 1.1-fold; 95% CI, 1.0–1.2; P = 0.60) compared to baseline. Cognitive flexibility was significantly worse after heading, even in contrast to the exercise without headers (P < 0.001, Fig. 1f). No correlation was observed between the number of headers and decrease in focused attention (r = −0.02, P = 0.93) or cognitive flexibility (r = −0.22, P = 0.24). The TMT scores are listed in Supplementary Table 1.
Plasma neuroproteins after exercise and repetitive heading in female soccer players
To investigate the effect of mRHI, female soccer players completed exercises with or without heading the ball, followed by analysis of neuronal markers at multiple time points (Fig. 1a). The mean plasma concentration of tau, pTau181 and pTau217 assessed at baseline were similar in both training groups (Fig. 1g, Supplementary Table 2). Quantitative immunoassay identified a significant relative increase in all forms of tau in plasma compared to baseline 1 hour after physical exercise and heading training (exercise group, tau, 1.4-fold; 95% CI, 1.3–1.5; pTau181, 1.3-fold; 95% CI, 1.2–1.4; pTau217, 1.2-fold; 95% CI, 1.1–1.3; heading group, tau, 1.2-fold; 95% CI, 1.1–1.3; pTau181, 1.3-fold; 95% CI, 1.2–1.4; pTau217, 1.3-fold; 95% CI, 1.3–1.4) (Fig. 1h-j). However, the dynamics of tau biomarkers differed between training groups and time points. The fold change of plasma tau protein gradually decreased over time, with values in the heading group being about 20% lower compared with the exercise group, with a significant difference at 1 h (P = 0.002) and 48 hours (P = 0.006). Interestingly, tau fold change in the heading group significantly decreased below baseline at 48 h (0.9-fold; 95% CI, 0.8–1.0; P = 0.041). No significant difference in pTau181 and pTau217 was found between the two training groups, and their relative content normalized to pre-training levels within 24 hours (Fig. 1i, j). Finally, we observed a higher pTau:tau fold change ratio for both phosphoepitopes in the heading group compared with the exercise group at all time points, and this ratio was significant for the pTau181 epitope at 1 h (exercise group, 0.95-fold; 95% CI, 0.9–1.0; heading group, 1.1-fold; 95% CI, 1.0–1.2; P = 0.026) and 48 h (exercise group, 0.95-fold; 95% CI, 0.9–1.0; heading group, 1.1-fold; 95% CI, 1.0–1.1; P = 0.005; Fig. 1k, l). Mean raw concentrations of the assayed proteins are listed in Supplementary Table 2.
Deregulation of peripheral microRNAs after exercise and repetitive heading
We performed an exploratory targeted microRNA (miRNA) analysis to investigate the transcriptional signatures of repetitive headers while controlling for the effects of physical exercise. Significant differences in miRNA expression were observed between exercise without headers and heading training. MiRNA profiling revealed 5 down-regulated and 6 up-regulated miRNAs in at least one time point in response to exercise, while 9 miRNAs were decreased, and 4 miRNAs increased after heading training. Among the differentially expressed miRNAs, two miRNAs were shared between the exercise and heading training. MiR-301a-3p showed the opposite pattern of expression after exercise in contrast to heading, while miR-421 was down-regulated after both types of soccer training (Table 3). A complete list of all evaluated miRNAs is provided in Supplementary Data 1 and Supplementary Data 2.
Signaling pathways
To characterize the molecular signaling associated with repetitive head impacts and exercise, we performed pathway enrichment analysis. First, we identified target genes for all deregulated miRNAs by querying the mirDIP portal. Next, we used a pathDIP database to perform pathway enrichment analysis using all identified genes. The resulting 582 enriched pathways, 187 for exercise and 397 for heading, were grouped into 7 representative pathway types and 53 pathway ontology-based categories. Cluster analysis revealed that heading training is specifically associated with an increased number of highly significant pathways, with genetic and environmental information processing along with disease signaling being the most overrepresented pathway types (Fig. 2a). Further stratification revealed that the majority of identified enriched pathways is shared between exercise and heading, and only a fraction of enriched pathways is unique to particular training. The top-scored pathways unique for heading are associated with diseases of the immune system, metabolic diseases, cardiovascular diseases, cell cycle disorders and aging, while the pathways unique for exercise include signaling associated with the sensory system and regulation of the circadian clock (Fig. 2b). Additionally, analysis of shared pathway categories identified several pathways that are predominantly associated with heading compared to exercise. Specifically, the immunostimulatory signaling including IL-3, IL-6, B-cell and T-cell receptors, post-translational protein modifications and protein degradation were highly abundant in heading. The bioinformatic analysis also revealed enrichment of CNS-associated pathways, such as the neuronal development and regeneration pathways including synaptic signaling, neurite outgrowth, axon guidance, release of neurotransmitters and cell survival stimuli that were specifically enriched in heading compared to exercise (Fig. 2c).

a Annotation of pathways associated with deregulated plasma miRNAs after heading (purple) and exercise (orange) revealed that heading training is specifically associated with an increased number of disease-related and genetic/environmental information processing pathways when compared to exercise. Node size corresponds to the gene ratio and x-axis denotes the statistical significance using the false discovery rate (Benjamini-Hochberg method), as per legend. b Stratification of identified significantly enriched pathways after heading and exercise into pathway categories. c Subnetwork analysis of shared categories revealed that heading training is specifically associated with immunostimulatory, neuronal development and cell survival pathways. Edge color denotes the type of training, edge thickness corresponds to the number of pathways per category, and edge transparency signifies statistical significance using the false discovery rate (Benjamini-Hochberg method), as per legend. d Analysis of gene-disease associations revealed a higher number of proteins associated with neurodegeneration and increased number of interactions linked to Alzheimer’s disease (AD) after heading compared to exercise. Node color corresponds to gene ontology-biological process, node size and transparency represents the number of interacting partners, as per legend. Edge color only highlights intra- and inter-group interactions.
Protein-protein interaction analysis
To investigate the link between repetitive head impacts and neurodegeneration, we have queried the IID database to determine physical protein interactions of the protein targets of deregulated miRNAs. The entire network with 10,758 interactions was further filtered by relevance to neurodegeneration, using IID annotation from the Disease Ontology and DisGeNET database of gene-disease associations. The resulting network comprised 129 proteins connected with 788 interactions relevant to Alzheimer’s disease (AD). A higher number of neurodegeneration-associated proteins, together with the increased number of their mutual AD-relevant interactions, was identified in heading compared to exercise (Fig. 2d). This group also includes proteins that are involved in various neurodegenerative disorders, e.g., SNCA in Parkinson’s disease (PD) and TARDBP in Amyotrophic lateral sclerosis (ALS). Furthermore, the group of interacting proteins specifically enriched after heading includes proteins associated with cell-cell interactions and intracellular scaffold (ANK3, CD2AP, CFL2, FERMT2, FRMD6, NEFM), inflammatory response (CD69, SOCS6, ST18), synaptic vesicle transport and release of neurotransmitters (GRIN2A, SLC17A7, SNAP25, SYNJ1), neuronal development and axon guidance (EPHA4, DCX, DLL1, DPYSL2, MEF2A, NR4A2) and APP processing (ADAM9, IDE, VPS26A).
Differences between female and male players in response to heading and exercise
Recently, we obtained cognitive scores and plasma biomarker data on a cohort of elite male soccer players under highly similar training protocol33. To further analyze the differences in the response to repetitive heading between female and male soccer players, we compared the outcomes of cognitive testing and tau protein levels in these groups (Fig. 3a). The basic characteristics of training units, including the parameters of physical performance and the number of headers, are compared in Table 4. Training units involving male soccer players used a higher number of headers (256) compared to females (152), modelling the higher frequency of headers in male competitions48,49,50. Analysis of physical performance revealed higher heart rate (exercise females EF, 147 bpm; 95% CI, 143–151 bpm vs exercise males EM, 138 bpm; 95% CI, 134–143 bpm; P = 0.002) (heading females H + EF, 140 bpm; 95% CI, 136–145 bpm vs heading males H + EM, 131 bpm; 95% CI, 126–136 bpm; P = 0.003) and percentage of maximal heart rate (EF, 77%; 95% CI, 75–78% vs EM, 73%; 95% CI, 71–75%; P = 0.007) (H + EF, 73%; 95% CI, 71–76% vs H + EM, 69%; 95% CI, 67–72%; P = 0.019) in female players compared to male players during both types of training, although still in the aerobic interval and consistent with sex-specific differences in cardiac parameters51. Neuropsychological examination using both parts of the TMT identified significantly worse post-exercise scores in male players compared to female players (TMT- A, EF, 1.8-fold; 95% CI, 1.7–2.0 vs EM 3.1-fold; 95% CI, 2.7-3.4; P < 0.001) (TMT-B, EF, 1.1-fold; 95% CI, 1.0–1.2 vs EM 1.7-fold; 95% CI, 1.4–1.9; P < 0.001). Heading training resulted in worse scores of TMT-A (H + EF, 3.4-fold; 95% CI, 3.1-3.7 vs H + EM 1.4-fold; 95% CI, 1.2–1.5; P < 0.001) and TMT-B (H + EF, 1.5-fold; 95% CI, 1.4–1.7 vs H + EM 1.3-fold; 95% CI, 1.1–1.5; P = 0.023) specifically in female players. A comparison of female and male players revealed statistically significant sexual dimorphism in response to exercise and heading training (Fig. 3b, c), with female soccer players showing higher deficits in focused attention and cognitive flexibility, despite performing a significantly lower number of headers.

a Schematic illustration of the experimental setup. Psychological testing revealed significantly worse scores in (b) focused attention and (c) cognitive flexibility in male (M) soccer players compared to female (F) players after exercise (E), while heading training (H + E) showed greater negative effects on cognitive performance in females compared to males. Fold change for the male cohort was calculated using a pre-exercise baseline. d Analysis of tau fold change in plasma showed no difference between male and female players. e Significant difference in pTau181 was observed in males 1 hour after exercise and 1 to 24 hours after heading compared to females. f The ratio of pTau181:tau fold change was significantly higher 1 to 24 hour after exercise and 1 hour post-heading in male players compared to females. The dotted line in (b-f) represents the pre-training baseline and the horizontal black lines denote the mean with 95% CI. Number of headers correlated with (g) the change in plasma pTau181 and (h) plasma pTau181:tau fold change ratio in a combined group of soccer players at 1 hour post-heading. The dotted line (g, h) shows the 95% CI bands. i The ROC analysis showed high accuracy for plasma pTau181:tau fold change ratio and Tau fold change to distinguish exposure to repetitive head impacts (n = 58, F = 30, M = 28) from exercise in a group of male and female athletes (n = 68, F = 36, M = 32).
Comparison of changes in plasma tau protein revealed no difference between the groups at 1 to 24 hours post-training (Fig. 3d). However, the change in pTau181 was significantly higher in male soccer players compared to female players at 1 h post-exercise (E1F, 1.3-fold; 95% CI, 1.2–1.4 vs E1M, 1.4-fold; 95% CI, 1.3–1.5; P = 0.03) and 1 to 24 h post-heading training (H + E1F, 1.3-fold; 95% CI, 1.2–1.4 vs H + E1M, 1.5-fold; 95% CI, 1.4–1.7; P = 0.01; H + E24F, 1.0-fold; 95% CI, 0.9–1.1 vs H + E24M, 1.2-fold; 95% CI, 1.1-1.3; P = 0.01) (Fig. 3e). We also observed a specific increase in the pTau181:tau ratio in males compared to females 1 to 24 h post-exercise (E1F, 1.0-fold; 95% CI, 0.9–1.0 vs E1M, 1.1-fold; 95% CI, 1.0–1.2; P = 0.002; E24F, 1.0-fold; 95% CI, 0.9–1.1 vs E24M, 1.1-fold; 95% CI, 1.0–1.2; P = 0.02) and 1 hour post-heading (H + E1F, 1.1-fold; 95% CI, 1.0–1.2 vs H + E1M, 1.2-fold; 95% CI, 1.1–1.2; P = 0.046) (Fig. 3f, Supplementary Table 3).
Predictive power of tau plasma biomarkers for mRHI
Further analyses assessed the correlation between changes in tau proteins and repetitive headers in a combined group of elite soccer players (n = 58), including all females (n = 30) and males (n = 28) who performed headers. Firstly, the total number of headers correlated with the pTau181 fold change (r = 0.38, P = 0.003) and pTau181:tau fold change ratio (r = 0.36, P = 0.006) at 1 h timepoint (Fig. 3g, h). No correlation was observed between the number of headers and the change in total tau (r = 0.12, P = 0.38) in plasma, indicating that exposure to mRHI was associated with a higher peripheral pTau181:tau fold change ratio. Further, as our results suggest that tau biomarkers could differentiate athletes with mRHI from controls, we performed ROC analysis. The plasma pTau181:tau fold change ratio showed high accuracy for distinguishing exposure to mRHI from soccer training without headers in a group of female and male athletes (AUC = 0.690, P < 0.001), whereas tau protein change showed lower, but significant predictive accuracy (AUC = 0.605, P = 0.041), while pTau181 change alone had poor accuracy (AUC = 0.548, P = 0.35) (Fig. 3i, Supplementary Table 4). The ROC analysis of pTau181:tau fold change ratio in the mixed cohort was also independently reproduced in females (AUC = 0.716, P = 0.003) and males (AUC = 0.664, P = 0.029), indicating general predictive accuracy regardless of sex. Thus, we surmise that quantification of the pTau181:tau fold change ratio shows potential as an acute biomarker of mild repetitive head impacts.
Discussion
In this study, we analyzed the effect of mild repetitive head impacts (mRHI) in female soccer players on the levels of selected biomarkers and short-term neurocognitive outcomes. We provide evidence suggesting that mRHI in women can lead to significant decrease in short-term cognitive functions, along with increases of pTau181 to total tau fold change ratio. Simultaneously, the panel of miRNAs altered after the impact indicates a modulation of signaling pathways involved in neuronal, immune and several other metabolic processes involved in neurodegeneration.
As most published studies in human biomedicine originate from analyses of male subjects, research in neuroscience and sports-related disciplines suffer from a significant gender disparity5,52. This prospective study sheds light on the association of mental and molecular processes that occur in females after mRHI. Physical exercise alone has previously been associated with acute impairment of neuropsychological abilities such as focused attention and cognitive flexibility33,53,54,55. Therefore, it is necessary to consider the physiological responses to exercise also without the presence of head impacts. Our analysis identified altered short-term cognition in response to exercise alone and after exercise including repetitive head impacts.
Female soccer players manifested worse cognitive functions after heading compared to baseline and also compared to the male soccer players. We clearly showed that female players are more sensitive to mRHI and have a higher acute susceptibility to mRHI sequelae in soccer, compared to male players. The effect is also confirmed by the fact that the number of headers in the female group was lower than in the male group. These findings are consistent with a generally accepted higher susceptibility of females to perceive the consequences of head impacts18,56, and may also be in parallel with a higher prevalence and severity of post-concussion symptoms reported in females57,58.
As the molecular mechanisms leading to dementia are linked to abnormal phosphorylation of tau protein, assays profiling various pTau epitopes represent a viable strategy for diagnostics and prognostics of brain injury23, lifetime exposure to RHI59, but also seem to be informative for assessment of effects after nonconcussive RHI60. Both phosphorylated tau epitopes, pTau181 and pTau217, investigated in this research are also used for diagnostic prediction of mild cognitive impairment and early-stage dementia25,61,62,63,64,65,66.
Several studies have not reported plasma tau protein as a specific marker for RHI and sport related brain injuries60,67. In our study, we observed a significant increase in tau after exercise and heading training in female soccer players, which is consistent with previously published findings in a male cohort33. However, there is no difference in tau between the sexes, and there also appears to be no association between the number of headers and total tau levels when both female and male players are combined into one cohort. On the other hand, the altered profile of phosphorylated tau and pTau181:tau fold change ratio is significantly higher in males compared to females and is positively associated with the frequency of headers in a combined cohort, indicating that higher mRHI exposure may lead to a relative increase in pTau181 in the plasma of soccer players. A similar trend of increased phosphorylated tau after repetitive headers was also identified for pTau217. These findings are in line with studies reporting increased pTau181 and pTau231 levels after acute and chronic RHI exposure33,59,68. However, it should be noted that relatively small magnitude of change observed in our exploratory study should be further validated in a larger cohort.
Controlling for the physical exercise in our study clearly demonstrates that cycles in the level of tau and phosphorylated tau can be only partially attributed to the influence of physical activity, while these biomarkers are affected by mRHI. As shown, the quantitative change in the pTau181:tau fold change ratio specifically differentiates players with mRHI from players performing aerobic exercise without headers even after 48 h. Considering the longitudinal and repetitive nature of nonconcussive head impacts in exposed players during their professional career, we hypothesize that these oscillating amplitudes of the pTau:tau ratio may represent the first sign of a molecular imbalance leading to the onset of neurocognitive disorders later in life.
The circulating miRNAs miR-19a-3p, miR-93-3p, miR-155-5p, miR-223-5p and miR-338-3p observed in our study have been previously reported as deregulated after different types of aerobic exercise69,70,71,72,73. Altered profiles of miR-19b-3p, miR-378a-3p and miR-505-3p were shown to be associated with training-induced adaptations and metabolism in skeletal muscle74,75, providing supporting evidence for our findings. The panel of heading-specific miRNAs identified in this study is in agreement with other studies reporting decreased miR-144-5p in the plasma of soccer players performing repetitive headers31, but also further extends the evidence on blood miRNAs that are deregulated in response to repetitive head impacts26. However, at the level of miRNA, there is only a limited overlap with recent findings in sport-related concussions26,28,76,77. The lack of overlap in miRNA data between concussion and RHI indicates a distinct physiological response to various intensities and frequencies of head impacts. Nevertheless, the pathway enrichment analysis revealed a major involvement of CNS-specific signaling in response to mRHI, highlighting neurotrophic, brain-reparative, and immune response-regulatory pathways, as previously reported in mild to severe TBI27,78,79,80. In addition, this brain-specific signaling is associated with a higher number of protein interactions observed after mRHI, which are typically involved not only in normal brain functioning, but are also linked with various neurodegenerative diseases including AD, PD and ALS. The signaling events involved after mRHI point to recovery mechanisms typically associated with TBI, suggesting that even mild RHI may have a detrimental effect on the brain as neuroprotective signals emerge in the periphery.
Maintaining similar experimental conditions for male and female cohorts permitted direct comparison of cognitive and biomarker outcomes and disclosed sexual dimorphism in response to physical activity and mRHI. The effect of physical activity and mRHI on increased tau phosphorylation was confirmed on two epitopes, pTau181 and pTau217, which are relevant for tau pathophysiology. Nevertheless, there are some limitations related to factors that were not collected as a part of this exploratory study and that need to be addressed in future research to support our findings. Lifetime exposure to RHI during players’ previous professional careers may have influenced the acute responses measured in this study. Counter-balancing the E and H + E protocols by splitting groups and reversing the order of training units could provide an additional level of control to evaluate the potential confounding effect of exercise. The correlation between miRNA levels and RHI exposure considering the intensity and time after impacts, as well as the verification of protein interactions and pathways to assess their predictive power for RHI, could be further investigated in future studies. Longer follow-up periods would be necessary to determine whether the observed changes persist over time and to assess potential chronic effects of RHI. Finally, due to the sandwich design of the tau assay, potential proteolytic processing of the tau protein may result in a loss of detection power. We acknowledge that this shortcoming is inherently common to all sandwich antibody assays and needs to be addressed e.g. by single-epitope analytical approaches such as quantitative mass spectrometry, which are however not yet available for large-scale use. Consequently, counterintuitive relative reduction in plasma tau observed after the heading compared with the exercise-only group needs to be re-examined in further studies for possible tau protein truncation. As tau truncation is considered a risk factor in the development of tau aggregates, this will be important for elucidating the link between repetitive head impacts and neurodegeneration.
Conclusively, the results of our exploratory study in elite female soccer players suggest that mild repetitive head impacts may lead to (1) acute disbalance of plasma tau and pTau181:tau fold change ratio, (2) short-term decline in cognitive performance, and (3) initiation of neuroprotective pathways. Compared to males, female players appeared to be more sensitive to post-mRHI sequelae, manifested by a higher impact on cognition and an altered pTau profile. Albeit being exploratory and requiring further confirmation in replicated experiments, our findings provide insight into acute responses to mRHI, and are potentially relevant for further biomarker research and risk assessment of developing neurocognitive or neurodegenerative disorders due to mRHI exposure in later life, reflecting the specific conditions of the female population.
Data availability
All source data are available in the main text or in the supplementary tables. Supplementary Data 1 shows the Panel of evaluated miRNAs during soccer exercise without headers. Supplementary Data 2 shows the Panel of evaluated miRNAs during soccer exercise including heading of the ball. Supplementary Table 1 shows the TMT score of female soccer players. Supplementary Table 2 shows Protein concentrations in response to exercise and repetitive heading. Supplementary Table 3 shows Changes in plasma tau protein in females and males in response to exercise and repetitive heading. Supplementary Table 4 shows Receiver Operating Characteristic (ROC) analysis for biomarker identification. The source data for Fig. 1b-d is in Table 4, for Fig. 1e, f in Supplementary Table 1 and for Fig. 1g-l in Supplementary Table 2. The source data for Fig. 2a-c is in Supplementary Data 3 and for Fig. 2d in Supplementary Data 4. The source data for female cohort in Fig. 3b, c is in Supplementary Table 1 and source data for males are available in Cente et al. 202333, Table 3. The source data for Fig. 3d-f is provided in Supplementary Table 3 and for Fig. 3i in Supplementary Table 4.
References
2023 Alzheimer’s disease facts and figures. Alzheimers Dement 19, 1598–1695, (2023).
Gustavsson, A. et al. Global estimates on the number of persons across the Alzheimer’s disease continuum. Alzheimers Dement 19, 658–670 (2023).
Buckley, R. F., Gong, J. & Woodward, M. A call to action to address sex differences in Alzheimer disease clinical trials. JAMA Neurol. 80, 769–770 (2023).
Miramontes, S. et al. Alzheimer’s disease as a women’s health challenge: a call for action on integrative precision medicine approaches. NPJ Women’s. Health 2, 17 (2024).
Haupt, S., Carcel, C. & Norton, R. Neglecting sex and gender in research is a public-health risk. Nature 629, 527–530 (2024).
Mackay, D. F. et al. Neurodegenerative disease mortality among former professional soccer players. N. Engl. J. Med. 381, 1801–1808 (2019).
Ueda, P. et al. Neurodegenerative disease among male elite football (soccer) players in Sweden: a cohort study. Lancet Public Health 8, e256–e265 (2023).
Batty, G. D. et al. Dementia in former amateur and professional contact sports participants: population-based cohort study, systematic review, and meta-analysis. EClinicalMedicine 61, 102056 (2023).
Fédération Internationale de Football Association (FIFA). Women’s Football: Member Associations Survey Report 2023, <https://digitalhub.fifa.com/m/28ed34bd888832a8/original/FIFA-Women-s-Football-MA-Survey-Report-2023.pdf> (2023).
Covassin, T., Swanik, C. B. & Sachs, M. L. Sex differences and the incidence of concussions among collegiate athletes. J. Athl. Train. 38, 238–244 (2003).
Bretzin, A. C., Covassin, T., Wiebe, D. J. & Stewart, W. Association of sex with adolescent soccer concussion incidence and characteristics. JAMA Netw. Open 4, e218191 (2021).
Hardaker, N. J., Hume, P. A. & Sims, S. T. Differences in injury profiles between female and male athletes across the participant classification framework: a systematic review and meta-analysis. Sports Med 54, 1595–1665 (2024).
Cheng, J. et al. Sex-based differences in the incidence of sports-related concussion: systematic review and meta-analysis. Sports Health 11, 486–491 (2019).
Dave, U. et al. Systematic review and meta-analysis of sex-based differences for concussion incidence in soccer. Phys. Sportsmed. 50, 11–19 (2022).
Espahbodi, S. et al. Heading frequency and risk of cognitive impairment in retired male professional soccer players. JAMA Netw. Open 6, e2323822 (2023).
Di Virgilio, T. G. et al. Evidence for acute electrophysiological and cognitive changes following routine soccer heading. EBioMedicine 13, 66–71 (2016).
Kern, J., Gulde, P. & Hermsdorfer, J. A prospective investigation of the effects of soccer heading on cognitive and sensorimotor performances in semi-professional female players. Front Hum. Neurosci. 18, 1345868 (2024).
Ye, K. et al. Repetitive soccer heading adversely impacts short-term learning among adult women. J. Sci. Med Sport 25, 935–941 (2022).
Bruno, D. & Rutherford, A. Cognitive ability in former professional football (soccer) players is associated with estimated heading frequency. J. Neuropsychol. 16, 434–443 (2022).
Iverson, G. L. et al. Examining later-in-life health risks associated with sport-related concussion and repetitive head impacts: a systematic review of case-control and cohort studies. Br. J. Sports Med 57, 810–821 (2023).
Vann Jones, S. A., Breakey, R. W. & Evans, P. J. Heading in football, long-term cognitive decline and dementia: evidence from screening retired professional footballers. Br. J. Sports Med 48, 159–161 (2014).
Barthelemy, N. R. et al. Highly accurate blood test for Alzheimer’s disease is similar or superior to clinical cerebrospinal fluid tests. Nat. Med 30, 1085–1095 (2024).
Rubenstein, R. et al. Comparing Plasma Phospho Tau, Total Tau, and Phospho Tau-Total Tau ratio as acute and chronic traumatic brain injury biomarkers. JAMA Neurol. 74, 1063–1072 (2017).
Thijssen, E. H. et al. Diagnostic value of plasma phosphorylated tau181 in Alzheimer’s disease and frontotemporal lobar degeneration. Nat. Med 26, 387–397 (2020).
Thijssen, E. H. et al. Plasma phosphorylated tau 217 and phosphorylated tau 181 as biomarkers in Alzheimer’s disease and frontotemporal lobar degeneration: a retrospective diagnostic performance study. Lancet Neurol. 20, 739–752 (2021).
Sandmo, S. B. et al. Changes in circulating microRNAs following head impacts in soccer. Brain Inj. 36, 560–571 (2022).
Cente, M. et al. Traumatic MicroRNAs: deconvolving the signal after severe traumatic brain injury. Cell Mol. Neurobiol. 43, 1061–1075 (2023).
Wyczechowska, D. et al. Serum microRNAs associated with concussion in football players. Front Neurol. 14, 1155479 (2023).
Hicks, S. D. et al. Refinement of saliva microRNA biomarkers for sports-related concussion. J. Sport Health Sci. 12, 369–378 (2023).
Johnson, J. J. et al. Association of salivary microRNA changes with prolonged concussion symptoms. JAMA Pediatr. 172, 65–73 (2018).
Munoz, E. R. et al. Effects of purposeful soccer heading on circulating small extracellular vesicle concentration and cargo. J. Sport Health Sci. 10, 122–130 (2021).
Sandmo, S. B. et al. Neurofilament light and tau in serum after head-impact exposure in soccer. Brain Inj. 34, 602–609 (2020).
Cente, M. et al. Association of Nonconcussive Repetitive Head Impacts and Intense Physical Activity With Levels of Phosphorylated Tau181 and Total Tau in Plasma of Young Elite Soccer Players. JAMA Netw. Open 6, e236101 (2023).
Benson, R. & Connolly, D. Heart rate training. (Human Kinetics, 2011).
Benson, R. & Connolly, D. Heart rate training. Second edition. edn, (Human Kinetics, 2020).
The International Football Association Board. Laws of the Game 2022/2023, <https://downloads.theifab.com/downloads/laws-of-the-game-2022-23?l=en> (2022).
Cheng, H. H. et al. Circulating microRNA profiling identifies a subset of metastatic prostate cancer patients with evidence of cancer-associated hypoxia. PLoS One 8, e69239 (2013).
Blondal, T. et al. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods 59, S1–S6 (2013).
Hauschild, A. C. et al. MirDIP 5.2: tissue context annotation and novel microRNA curation. Nucleic Acids Res. 51, D217–D225 (2023).
Pastrello, C., Kotlyar, M., Abovsky, M., Lu, R. & Jurisica, I. PathDIP 5: improving coverage and making enrichment analysis more biologically meaningful. Nucleic Acids Res. 52, D663–D671 (2024).
Kotlyar, M. et al. IID 2021: towards context-specific protein interaction analyses by increased coverage, enhanced annotation and enrichment analysis. Nucleic Acids Res. 50, D640–D647 (2022).
Kotlyar, M., Pastrello, C., Malik, Z. & Jurisica, I. IID 2018 update: context-specific physical protein-protein interactions in human, model organisms and domesticated species. Nucleic Acids Res. 47, D581–D589 (2019).
Brown, K. R. et al. NAViGaTOR: network analysis, visualization and graphing Toronto. Bioinformatics 25, 3327–3329 (2009).
Reitan, R. M. Trail making test. Manual for administration, scoring, and interpretation. (Indiana University Press., 1956).
Reitan, R. M. & Wolfson, D. The trail making test as an initial screening procedure for neuropsychological impairment in older children. Arch. Clin. Neuropsychol. 19, 281–288 (2004).
Wagner, S., Helmreich, I., Dahmen, N., Lieb, K. & Tadic, A. Reliability of three alternate forms of the trail making tests a and B. Arch. Clin. Neuropsychol. 26, 314–321 (2011).
Mueller, S. T. & Piper, B. J. The Psychology Experiment Building Language (PEBL) and PEBL Test Battery. J. Neurosci. Methods 222, 250–259 (2014).
Langdon, S., Goedhart, E., Oosterlaan, J. & Konigs, M. Heading Exposure in Elite Football (Soccer): A Study in Adolescent, Young Adult, and Adult Male and Female Players. Med Sci. Sports Exerc 54, 1459–1465 (2022).
Beaudouin, F. et al. The UEFA Heading Study: Heading incidence in children’s and youth’ football (soccer) in eight European countries. Scand. J. Med Sci. Sports 30, 1506–1517 (2020).
Wahlquist, V. E. et al. Quantitative Analysis of Ball-Head Impact Exposure in Youth Soccer Players. J. Sports Sci. Med. 22, 591–596 (2023).
Pierre, S. T., Peirlinck, S. R. & Kuhl, M. E. Sex Matters: A Comprehensive Comparison of Female and Male Hearts. Front Physiol. 13, 831179 (2022).
D’Lauro, C. et al. Under-representation of female athletes in research informing influential concussion consensus and position statements: an evidence review and synthesis. Br J Sports Med, https://doi.org/10.1136/bjsports-2021-105045 (2022).
Moreira, A. et al. Mental fatigue impairs technical performance and alters neuroendocrine and autonomic responses in elite young basketball players. Physiol. Behav. 196, 112–118 (2018).
Nedelec, M. et al. Recovery in soccer: part I - post-match fatigue and time course of recovery. Sports Med. 42, 997–1015 (2012).
Sun, H., Soh, K. G., Roslan, S., Wazir, M. & Soh, K. L. Does mental fatigue affect skilled performance in athletes? A systematic review. PLoS One 16, e0258307 (2021).
Berz, K. et al. Sex-specific differences in the severity of symptoms and recovery rate following sports-related concussion in young athletes. Phys. Sportsmed. 41, 58–63 (2013).
Babakhanyan, I. et al. Gender disparities in neurobehavioral symptoms and the role of post-traumatic symptoms in us service Members following mild traumatic brain injury. J. Neurotrauma 41, e1687–e1696 (2024).
Starkey, N. J. et al. Sex differences in outcomes from mild traumatic brain injury eight years post-injury. PLoS One 17, e0269101 (2022).
Miner, A. E. et al. Examination of plasma biomarkers of amyloid, tau, neurodegeneration, and neuroinflammation in former elite American football players. Alzheimers Dement, https://doi.org/10.1002/alz.14231 (2024).
Lember, L. M. et al. The use of biofluid markers to evaluate the consequences of sport-related subconcussive head impact exposure: a scoping review. Sports Med Open 10, 12 (2024).
Janelidze, S. et al. Plasma P-tau181 in Alzheimer’s disease: relationship to other biomarkers, differential diagnosis, neuropathology and longitudinal progression to Alzheimer’s dementia. Nat. Med. 26, 379–386 (2020).
Barthelemy, N. R., Horie, K., Sato, C. & Bateman, R. J. Blood plasma phosphorylated-tau isoforms track CNS change in Alzheimer’s disease. J Exp Med. 217, https://doi.org/10.1084/jem.20200861 (2020).
Palmqvist, S. et al. Discriminative Accuracy of Plasma Phospho-tau217 for Alzheimer disease vs other neurodegenerative disorders. JAMA 324, 772–781 (2020).
Jonaitis, E. M. et al. Plasma phosphorylated tau 217 in preclinical Alzheimer’s disease. Brain Commun. 5, fcad057 (2023).
Palmqvist, S. et al. Prediction of future Alzheimer’s disease dementia using plasma phospho-tau combined with other accessible measures. Nat. Med. 27, 1034–1042 (2021).
Ashton, N. J. et al. Diagnostic accuracy of a plasma phosphorylated Tau 217 immunoassay for alzheimer disease pathology. JAMA Neurol. 81, 255–263 (2024).
Laverse, E. et al. Plasma glial fibrillary acidic protein and neurofilament light chain, but not tau, are biomarkers of sports-related mild traumatic brain injury. Brain Commun. 2, fcaa137 (2020).
Muraoka, S. et al. Proteomic Profiling of Extracellular Vesicles Separated from Plasma of Former National Football League Players at Risk for Chronic Traumatic Encephalopathy. Aging Dis. 12, 1363–1375 (2021).
Fernandez-Sanjurjo, M. et al. Next-generation sequencing reveals that miR-16-5p, miR-19a-3p, miR-451a, and miR-25-3p cargo in plasma extracellular vesicles differentiates sedentary young males from athletes. Eur. J. Sport Sci. 24, 766–776 (2024).
Narayan, V., Shivapurkar, N. & Baraniuk, J. N. Informatics Inference of Exercise-Induced Modulation of Brain Pathways Based on Cerebrospinal Fluid Micro-RNAs in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Netw. Syst. Med. 3, 142–158 (2020).
Agostini, S. et al. Evaluation of serum miRNAs expression in frail and robust subjects undergoing multicomponent exercise protocol (VIVIFRAIL). J. Transl. Med. 21, 67 (2023).
de Gonzalo-Calvo, D. et al. Circulating inflammatory miRNA signature in response to different doses of aerobic exercise. J. Appl Physiol. 119, 124–134 (2015).
Nielsen, S. et al. The miRNA plasma signature in response to acute aerobic exercise and endurance training. PLoS One 9, e87308 (2014).
Krammer, U. D. B. et al. MiRNA-based “fitness score” to assess the individual response to diet, metabolism, and exercise. J. Int Soc. Sports Nutr. 19, 455–473 (2022).
Massart, J. et al. Endurance exercise training-responsive miR-19b-3p improves skeletal muscle glucose metabolism. Nat. Commun. 12, 5948 (2021).
Shultz, S. R. et al. Decrease in Plasma miR-27a and miR-221 After Concussion in Australian Football Players. Biomark. Insights 17, 11772719221081318 (2022).
LaRocca, D. et al. Comparison of serum and saliva miRNAs for identification and characterization of mTBI in adult mixed martial arts fighters. PLoS One 14, e0207785 (2019).
Matyasova, K., Csicsatkova, N., Filipcik, P., Jurisica, I. & Cente, M. Peripheral microRNA alteration and pathway signaling after mild traumatic brain injur. y. Gen. Physiol. Biophys. 40, 523–539 (2021).
O’Connell, G. C., Smothers, C. G. & Winkelman, C. Bioinformatic analysis of brain-specific miRNAs for identification of candidate traumatic brain injury blood biomarkers. Brain Inj. 34, 965–974 (2020).
Di Pietro, V., Yakoub, K. M., Scarpa, U., Di Pietro, C. & Belli, A. MicroRNA signature of traumatic brain injury: from the biomarker discovery to the point-of-care. Front Neurol. 9, 429 (2018).
Acknowledgements
We would like to thank Silvia Putekova, PhD, Jana Marcinkova, PhD, and Gabriela Doktorova, PhD, of the Department of Nursing of the Trnava University for their help with blood sampling. We would like to thank Juliana Melichova, PhD, Dominika Plancikova, PhD, Anna Vanacka, BSc, Vladislava Libiakova, BSc, of the Department of Public Health of the Trnava University, Filip Jendrol, PhD, of the Department of Psychology of the Trnava University and others for their help with cognitive test applications and its organisation. This research was cofunded by the Slovak Research and Development Agency (grant Nos. APVV-22-0613, APVV-22-0470, APVV-20-0615, APVV-21-0254), the Ministry of Education, Research, Development and Youth of the Slovak Republic (grants VEGA 2/0086/23, 2/0141/23, 2/0153/22 and ERA-NET Neuron JTC2019 Neu-Vasc) and by the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 873127 - InterTAU. IJ was in part supported by grants from ERA-NET Neuron II/2023/77/Progress, Natural Sciences Research Council (NSERC RGPIN-2024-04314), CIHR (#519474), Canada Foundation for Innovation (CFI #225404, #30865), and Ontario Research Fund (RDI #34876, RE010-020). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Study design, J.P., P.P., I.T., M.Ma., M.S., M.C., and P.F.; investigation, J.P., P.P., I.J., I.T., J.H., P.S., M.Mi., and M.Sk.; data processing, J.P., M.C., I.J., and M.Ma.; writing–original draft preparation, M.C. and P.F.; writing–review and editing, M.C., P.F., I.J., M.Ma., J.P., P.P, and R.S.; funding acquisition, M.Ma., P.F., M.C., I.J., J.P., P.P., and R.S. All authors revised and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Medicine thanks Xintong Ge, Matthew J. Rogatzki and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cente, M., Perackova, J., Peracek, P. et al. Changes in peripheral blood and cognitive performance after nonconcussive head impacts in elite female soccer players. Commun Med 5, 417 (2025). https://doi.org/10.1038/s43856-025-01127-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43856-025-01127-8
